
Neuromolecular Underpinnings of Negative Cognitive Bias in Depression


Affective Cognitive Neuroscience Laboratory, Department of Pharmacology, Maj Institute of Pharmacology Polish Academy of Sciences, Smetna Street 12, 31-343 Krakow, Poland
*
Author to whom correspondence should be addressed.
Cells 2021, 10(11), 3157; https://0-doi-org.brum.beds.ac.uk/10.3390/cells10113157
Submission received: 27 August 2021
/
Revised: 9 November 2021
/
Accepted: 10 November 2021
/
Published: 13 November 2021
(This article belongs to the Special Issue Neurobiology of Depression: Molecular and Cellular Mechanisms)
Abstract
:This selective review aims to summarize the recent advances in understanding the neuromolecular underpinnings of biased cognition in depressive disorder. We begin by considering the cognitive correlates of depressed mood and the key brain systems implicated in its development. We then review the core findings across two domains of biased cognitive function in depression: pessimistic judgment bias and abnormal response to negative feedback. In considering their underlying substrates, we focus on the neurochemical mechanisms identified by genetic, molecular and pharmacological challenge studies. We conclude by discussing experimental approaches to the treatment of depression, which are derived largely from an improved understanding of its cognitive substrates.
Keywords:
depression; cognitive; dopamine; serotonin; noradrenaline; judgment bias; feedback sensitivity1. Introduction
Depression is the leading cause of disability in the 21st century, affecting an estimated 350 million people worldwide [1]. The number of people prescribed antidepressant medications, the first-line treatment for depressive disorder, increases each year, and the market for these medications is valued at billions of dollars [2]. However, the prevalence of depression has not decreased since accurate record-keeping began. One reason for this paradox is the failure of science to find a compelling biomedical explanation for depression and adequately address how and why this devastating condition occurs.
One of the most important yet still underappreciated symptoms of depression is aberrant cognition. Indeed, cognitive problems have been included in the diagnostic criteria for mood disorders, and according to the Diagnostic and Statistical Manual (DSM 5), the specific criteria for depression include a reduced ability to concentrate, disturbed memory and indecisiveness [3]. These cognitive problems are usually persistent, recurrent and significantly disrupt quality of life [4].
The symptomatology of depression suggests a processing bias toward negative aspects of the environment. Indeed, depressed individuals are more likely to recall negative autobiographical memories, and when they do recall positive experiences, they are overly general and lacking in detail [5]. Individuals also exhibit impaired recognition of happy facial expressions [6] and respond more rapidly to sad versus happy word targets [7]. The two most important characteristics of negatively biased cognition include pessimistic judgment bias [8,9] and catastrophic reactions to negative feedback [10,11].
Among the brain regions that have been most commonly implicated in the biased cognition associated with depression are the ventromedial (vlPFC) and orbitofrontal (OFC) cortices [12]. It has been suggested that the vlPFC and adjacent OFC are critical for making an association between a reinforced outcome and a given action, as well as for behavioral guidance once the expectancy of an outcome is violated [13]. A study by Wheeler and Fellows (2008) demonstrated that OFC is critical for learning from negative feedback [14]. Other important neuroanatomical correlates of negative processing bias include the dorsal and ventral striatum, the anterior insula extending to the lateral OFC and the cingulate cortex [12]. It has been suggested that the dorsal regions of the striatum are involved in forming habitual action selection following positive feedback, while the ventral regions mediate learning the value of positive feedback [15]. The insula has been suggested to be involved in the evaluation of negative outcomes and in the processing of changes in previously learned actions [16], while the anterior and posterior cingulate cortices are involved in post-feedback performance adjustments [17].
This selective minireview aims to summarize recent advances in understanding the neuromolecular underpinnings of biased cognition in depressive disorder. We begin by reviewing the core findings across two domains of biased cognitive function in depression: pessimistic judgment bias and abnormal response to negative feedback. In considering their underlying substrates, we focus on the neurochemical mechanisms identified by genetic, molecular and pharmacological challenge studies. We conclude by discussing potential experimental approaches to the treatment of depression, which are derived largely from an improved understanding of its cognitive substrates.
2. Pessimistic Judgment Bias
This term refers to the tendency to overestimate the likelihood of experiencing negative events while underestimating the likelihood of positive events [18]. This attitude of expecting the worst is a prominent cognitive feature of depression and can have considerable ramifications at both personal and societal levels [18,19].
The two major cognitive theories of depression, Seligman’s learned helplessness theory [20,21] and Beck’s cognitive model [22,23], emphasize the importance of hopelessness and pessimism about the future in the etiology, maintenance and treatment of depression. Moreover, according to both theories, a pessimistic view of the future and hopelessness—the belief that desirable outcomes are highly improbable and that aversive outcomes are very probable—are seen as underlying causes of depressive symptoms. Growing experimental evidence supports this notion. A study by Alloy and colleagues [24] demonstrated that depressed individuals, contrary to nondepressed controls, forecast the future for both the self and others in a pessimistic manner. In 2014, Korn and colleagues [25] demonstrated an absence of optimistic bias in belief updating in depressed individuals, and this absence correlated with symptom severity. A study by Zenger and colleagues [26] provided experimental evidence that elevated pessimism is a risk factor for anxiety and depression. Interestingly, a similar association was observed in preclinical studies in rats [27,28].
Although the theoretical rationale and experimental evidence regarding the association between pessimistic judgment bias and depression are relatively strong, information about the neuromolecular background of this interaction is still meager and requires elucidation.
One of the promising neuromolecular candidates involved in the regulation of expectations about future outcomes is dopamine (DA), a key neuromodulator in reward learning and reward-seeking behavior both in humans [29] and in animals [30]. In 2012, Sharot and colleagues [31] demonstrated for the first time that administration of a drug that enhances dopaminergic function, 3, 4-dihydroxy-L-phenylalanine (L-DOPA), impairs the ability to update belief in response to undesirable information about the future, generating optimism. Interestingly, L-DOPA was also shown to reduce pessimistic expectations by altering the value of information about potential losses, inducing, in this way, bias toward the information about potential gains [32]. Animal studies confirmed the important role of DA in the mediation of judgment bias. Thanks to the introduction of the ambiguous-cue interpretation (ACI) paradigm [33,34], a test allowing for measurement of cognitive judgment bias in animals, Rygula and colleagues demonstrated in 2014 that acute administration of the dopaminergic booster d-amphetamine produces optimism in rats [35]. This observation was confirmed several years later by Hales and colleagues [36]. Interestingly, published studies reported no optimism-inducing effects of another dopaminergic drug, cocaine [36,37]. Additionally, in 2017, Golebiowska and Rygula demonstrated, in the same animal model, that the effects of L-DOPA administration depend on the basal valence of cognitive judgment bias of experimental animals [38]. In that study, L-DOPA caused a pessimistic shift in animals classified as optimistic but had no effects on those classified as pessimistic. The inconsistency in the effects of L-DOPA between humans and animals was postulated to stem from the fact that the human study explored the valence of long-term expectations, while the animals responded to immediate rewards or punishments in the ACI paradigm. In the abovementioned study by Golebiowska and Rygula, similarly, twisted effects were observed following administration of another dopaminergic drug, the dopamine D2 receptor antagonist haloperidol [38]. The effects of haloperidol were reported to be opposite for animals classified as optimistic and pessimistic. The optimists became more pessimistic, while the pessimists became more optimistic. Because similar effects were observed following treatment with a serotonergic compound, the selective serotonin reuptake inhibitor (SSRI) escitalopram [38], it has been suggested that the effects of acute dopaminergic and serotonergic manipulations on pessimistic/optimistic interpretation of ambiguous cues may, at least in animals, depend on the basal valence of cognitive judgment bias [38].
The second key neurotransmitter implicated in cognitive judgment bias is serotonin (5-HT). Although in the abovementioned study by Sharot and colleagues [31], administration of the SSRI citalopram did not impact updating of beliefs about future life events, this does not imply that 5-HT function does not influence optimistic/pessimistic judgment bias. Indeed, several preclinical studies demonstrated that pharmacological modulation of 5-HT function by the administration of drugs from the family of SSRIs can change the interpretation of ambiguity in animals. In the study by Rygula and colleagues from 2014, administration of a low dose of the SSRI citalopram significantly biased animals toward the negative interpretation of the ambiguous cues in the ACI paradigm [35]. In contrast, acute administration of higher doses induced optimistic judgment bias [35]. To explain these contrasting effects, it has been proposed that the pessimistic shift observed following administration of the low dose of citalopram resulted from the temporary silencing of 5-HT system activity through stimulation of the serotonin 5-HT1A autoreceptors in the raphe nuclei [39,40]. According to this hypothesis, administration of higher doses of citalopram bypasses this inhibitory mechanism, allowing for a net increase in prefrontal 5-HT levels [41,42,43,44] and optimistic shift in judgment [35]. In 2013, Anderson and colleagues demonstrated pro-optimistic effects of chronic but not acute administration of another SSRI drug—fluoxetine [45]. These results were repeated and extended by Hales and colleagues [36], who demonstrated that the pro-optimistic effects of fluoxetine on the interpretation of ambiguous cues were most pronounced after 2 and 3 weeks of treatment [36]. A study by Doyle and colleagues [46] demonstrated that administration of the 5-HT inhibitor p-chlorophenylalanine induces pessimistic-like judgment bias in sheep. A similar effect was observed following 5-HT depletion in the ACI paradigm in pigs [47].
Along with DA and 5-HT, noradrenaline (NA) is the third neurotransmitter involved in mediating optimistic and pessimistic biases. In preclinical studies using animal models, administration of drugs boosting NA neurotransmission consistently biases cognitive judgment toward pessimism. In the study by Rygula and colleagues in 2014, the NA reuptake blocker desipramine at all tested doses rendered the rats pessimistic [35]. A study testing mazindol, another noradrenergic drug with mixed noradrenergic and dopaminergic mechanism of action, reported similar results [37]. A decrease in the positive processing of the ambiguous cue in the ACI paradigm was also reported by Anderson and colleagues after acute administration of the selective noradrenaline reuptake inhibitor (SNRI) reboxetine [45]. The same drug, in combination with the stress hormone corticosterone, was reported by Enkel and colleagues to produce pessimism in their landmark study with the ACI paradigm in rats [33].
Although judgment bias was initially seen mainly as a derivative of a current affective state, which depends on the environment to which an individual is exposed, rather than a phenotypic trait, recent research unequivocally demonstrated that optimism and pessimism can be considered stable and enduring phenotypic traits [28,48,49,50,51,52]. Nonetheless, research on the genetic background of judgment bias is still scarce. In a recent study using the ACI paradigm in zebrafish, Espigares and colleagues found that telomerase-deficient fish (tert−/−) were more pessimistic in response to ambiguous stimuli than their wild-type conspecifics [53]. These telomerase-deficient mutants have shorter telomeres than their wild-type siblings and develop degenerative phenotypes characterized by, e.g., increased inflammation, which is common in aged organisms. It has been suggested that this inflammation may be responsible for the altered judgment bias. Indeed, a correlational link between pessimism and inflammation has already been reported in humans [54] and animals [48]. In the latter study, trait pessimism was reported to be associated with decreased proliferative activity of splenocytes and increased production of interleukin-(IL)1β and IL-4, activin A, l-selectin, interferon (IFN)-γ and some chemokines and receptors for advanced glycation end products [48].
While the involvement of individual genes in the development of cognitive distortions and depression has not been widely proven, there are reports of the role of 5-HT transporter (SERT) polymorphisms in the development of depression, suggesting their important role in shaping cognitive biases associated with this disorder. Indeed, individuals carrying the short allele of the SERT gene are characterized by negative cognitive bias, as they tend to display enhanced attention toward negative information and interpret ambiguous stimuli in a more pessimistic way [55,56]. Although the effects observed in humans are generally supported by studies using animal models, some studies provided mixed results. In 2014, Kloke and colleagues reported a trend for homozygous SERT knockout mice to display pessimistic bias [57]. However, this finding was not confirmed in a recent study by Krakenberg and colleagues, who did not observe interactions between the SERT genotype and biased judgment [58]. It has been suggested that, at least in mice, the association between the SERT genotype and judgment bias is not straightforward. Other factors, including multiple genes and environmental influences, are implicated in the modulation of genotype and bias.
A recent study by Boddington and collaborators suggested that individual differences in biased cognition can be partially explained by variations in brain gene expression [59]. They analyzed the expression of several dopaminergic and serotonergic genes in the prefrontal cortex of red junglefowls and reported that chicks with higher dopamine D1 receptor expression were more optimistic, while chicks with higher serotonin 5-HT2A receptor expression tended to be less optimistic. These results further suggested the involvement of monoaminergic systems in cognitive judgment bias.
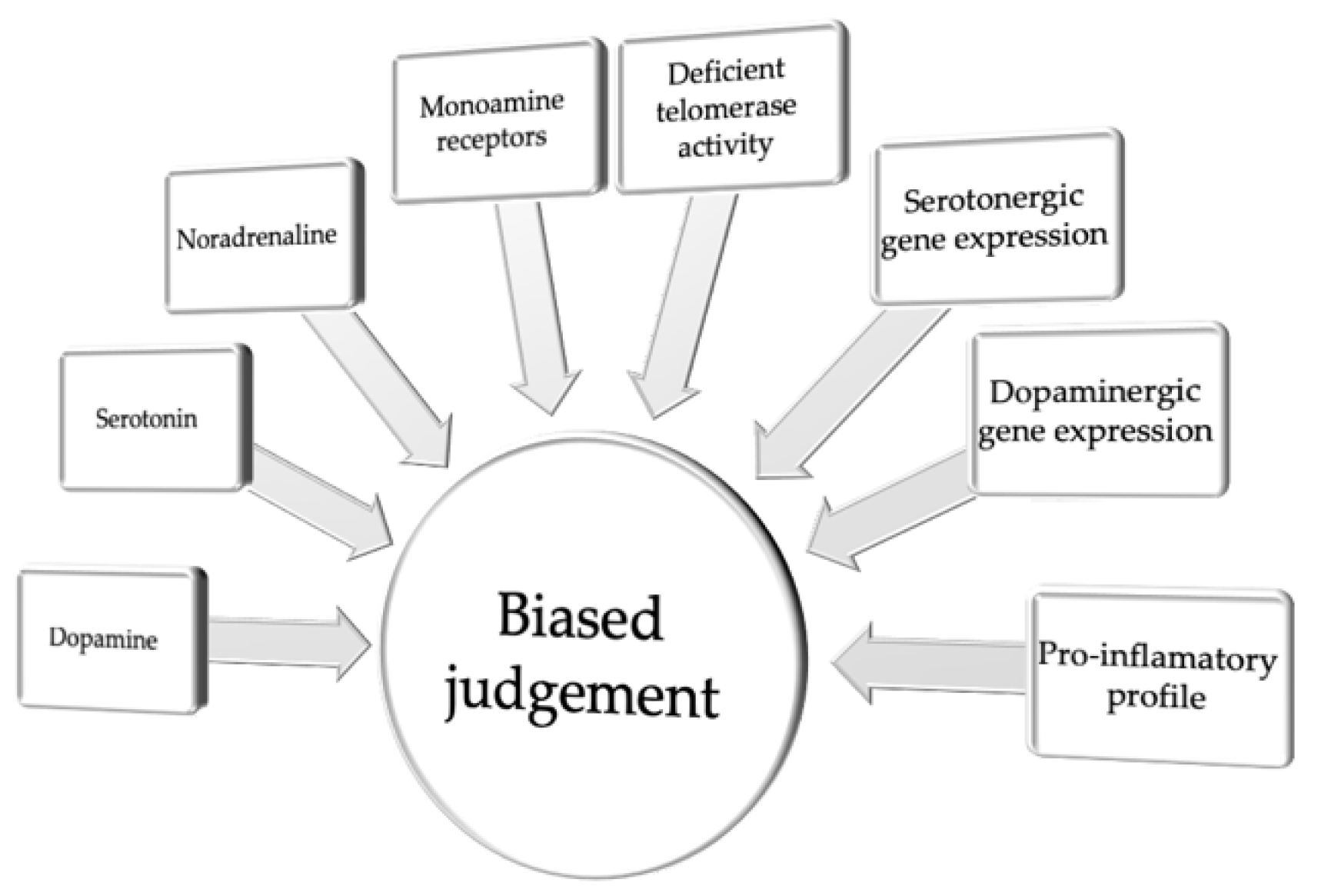
Taken together, although the mechanisms contributing to biased judgment in depression remain poorly understood, the majority of the studies conducted to date point to a common neuromolecular and cellular background of emotional regulation and cognitive judgment bias. This background includes various alterations in the function of the monoaminergic system function in the brain and associated physiological and cellular processes, such as a proinflammatory profile, variability in serotonergic and dopaminergic gene expression, or altered telomerase activity (Figure 1 and Table 1).
3. Biased Sensitivity to Feedback
People suffering from depression often ruminate over perceived failures and criticism [60]. A growing body of evidence shows that depressed individuals also have an exaggerated response to negative feedback during laboratory testing [11,61,62]. This effect was demonstrated for the first time by Elliot and colleagues [11], who found that if depressed individuals responded incorrectly on a given trial (trial N) of a simple memory task, they were disproportionately likely to fail the subsequent trial (N + 1). This “catastrophic response to perceived failure” was postulated to have an impact upon cognitive ability on any tasks that deliver performance-contingent feedback. Moreover, this effect appeared specific to depression because it was not seen in healthy controls or in any other neuropsychiatric condition [11]. The deleterious effects of hypersensitivity to negative feedback on task performance were later identified on a probabilistic reversal learning (PRL) task [61,62], during which subjects must learn to disregard misleading negative information (for review, see [63]). Apart from hypersensitivity to negative feedback, a growing number of studies examining cognitive processes in depression have suggested that depressed individuals also show hyposensitivity to positive feedback and altered processing of positively valenced information [45,64,65,66]. As postulated by Beck [23], people suffering from depression generally tend to distort environmental information negatively and thereby fail to accurately perceive or utilize positive information to modulate their behavioral responses [67,68]. Experimental evidence seems to support this notion, and numerous studies demonstrated that depressed and bipolar patients show decreased reward learning and generally reduced hedonic capacity [69,70,71].
Although studies conducted over the last two decades have shed some light on the neuromolecular background of altered sensitivity to feedback in depression, neurochemical correlates are still far from fully understood. One of the natural and most frequently studied neurotransmitters in this context is DA. In an already classic study from 2004, Frank and colleagues demonstrated that decreased availability of DA, which can be observed, e.g., in Parkinson’s patients off medication, is associated with better learning from negative feedback than from positive feedback [72]. Importantly, DA medication reversed this bias, making patients more sensitive to positive than negative outcomes. The computational model of reinforcement learning applied in the study allowed for the prediction of the abovementioned effects [72]. In a model proposed one year later, Frank postulated that low DA availability shifts the neurochemical balance in the basal ganglia toward an indirect (“NoGo”) pathway and impairs learning from positive feedback in comparison to learning from punishment [73]. In contrast, high DA availability leads to direct (“Go”) pathway overactivity and improves learning from positive feedback compared to learning from negative feedback [73]. A growing number of pharmacological, neuroimaging and genetic studies seem to support this model. Pharmacological studies have demonstrated that modulation of dopamine D2 receptors affects learning only from positive feedback but not from negative feedback [29,70,71,74], suggesting their specific involvement in learning from a reward. However, other studies in humans [75,76] and in animals [77,78,79,80] pointed to the specific role of dopamine D2 receptors in avoiding negative outcomes [73,81]. A recent study by Lim and collaborators [82] confirmed the blunting effects of dopamine D2 receptor agonism on learning from negative feedback in healthy participants and reported similar effects on learning from punishment, following the administration of the dopamine D2/3 receptor antagonist amisulpride. Such a nonselective effect of dopamine D2/3 receptor antagonism has already been previously reported [83,84], suggesting that these receptors are generally involved in feedback-based learning. Indeed, an elegant study by Cox and colleagues [76] demonstrated, using positron-emission tomography (PET), that individual differences in dopamine D1 and D2 receptor binding predict the effectiveness of learning from positive and negative feedback, respectively, and that DA depletion improves learning from negative feedback via dopamine D2 receptor signaling.
As proposed by Frank and O’reilley [81] and supported by the abovementioned studies, higher levels of DA (e.g., during unexpected rewards) switch the balance within the DA nigro-striatal circuit towards higher sensitivity of dopamine D1 receptors to its endogenous ligand, leading to activation of the (direct) “Go” pathway. On the contrary, DA depletion (e.g., during lack of reward) switches the balance toward higher sensitivity of dopamine D2 receptors and leads to activation of the (indirect) “No-Go” pathway. Therefore, activation of dopamine D1 receptors improves learning from positive feedback, while activation of dopamine D2 receptors improves learning from negative feedback.
Research by Cools and collaborators [85] revealed that subjects with high basal DA synthesis in the striatum show relatively better reversal learning from unexpected rewards than from unexpected punishments, whereas subjects with a low basal level of DA synthesis show the reverse pattern. In 2007, Frank and colleagues [86] demonstrated that three polymorphisms in genes associated with DA function contribute to reward and avoidance learning in humans. A polymorphism in the DARPP-32 gene (which encodes for a dopamine- and cAMP-regulated phosphoprotein that is a crucial mediator of the biochemical effects of DA) predicted relatively better probabilistic reward learning; the C957T polymorphism of the dopamine D2 receptor gene, associated with striatal dopamine D2 receptor function, predicted the degree to which participants learned to avoid choices that had been probabilistically associated with negative outcomes; and the Val/Met polymorphism of the catechol-O-methyltransferase (COMT) gene, associated with prefrontal cortical DA function, predicted participants’ ability to rapidly adapt behavior on a trial-to-trial basis. A specific role of another dopamine D2 receptor gene polymorphism (DRD2-TAQ-IA) in feedback-based learning in a human neuroimaging paradigm was demonstrated in 2007 by Klein and colleagues [87] and in 2009 by Jocham and colleagues [88]. In the former, A1 allele carriers with lower dopamine D2 receptor densities learned to avoid actions with negative consequences less efficiently than those without the allele. In the latter, the A1 subjects demonstrated an impaired ability to sustain a newly rewarded response after a reversal of the stimulus-reward contingency in the PRL task and showed a generally decreased tendency to stick with a rewarded response.
A growing number of studies have also implicated 5-HT in the modulation of sensitivity to feedback. Published reports suggest that increasing 5-HT transmission leads to a reduced sensitivity to aversive outcomes, whereas decreasing 5-HT transmission, by way of either acute tryptophan depletion, presynaptic receptor stimulation or upregulation of SERT, leads to an increased sensitivity to aversive outcomes (reviewed by [63]). In the pioneering study by Chamberlain and colleagues [89], a low, acute dose of the SSRI citalopram, which was postulated to activate presynaptic 5-HT autoreceptors and in this way downregulate 5-HT transmission, increased the tendency to switch the response following misleading negative feedback in the PRL task, mimicking the increased sensitivity to negative feedback observed in depression [61]. This effect of acute SSRI treatment was replicated 12 years later by Skandali and colleagues using escitalopram [90]. Similar effects of reduced 5-HT function were reported following acute tryptophan depletion, a procedure that has been used extensively to study the effect of low 5-HT levels in humans [91,92]. Additionally, a complex report by den Ouden and collaborators [93] revealed that allelic variation in SERT predicts negative feedback sensitivity (behavioral adaptation following punishment). Specifically, L′ homozygosity, which has been linked with increased SERT binding and decreased levels of extracellular 5-HT [94], was associated with increased sensitivity to negative feedback [93]. Studies in humans are complemented by results from animal models. In 2010, Bari and colleagues [95] repeated the effect observed by Chamberlain and collaborators in humans [89] using a pre-clinical version of the PRL test in rats treated with citalopram, while the study by Ineichen and colleagues demonstrated that in mice, a genetic reduction in SERT function leads to reduced sensitivity to negative feedback [96]. In 2015, Rygula and colleagues showed that selective 5-HT depletion in the amygdala increases sensitivity to aversive feedback and reduces punishment-induced response suppression in nonhuman primates [97], and a study by Phillips and colleagues revealed that sensitivity to positive feedback can be modulated by pharmacological targeting of serotonin receptor 5-HT2C [98]. In the latter study, administration of the serotonin 5-HT2C receptor antagonist SB 242084 reduced sensitivity to positive feedback, while administration of the serotonin 5-HT2C receptor agonist WAY 163909 resulted in changes associated with increased sensitivity to positive feedback and decreased sensitivity to negative feedback. It has been proposed that oversensitivity to negative feedback associated with low levels of tonic 5-HT could represent either enhanced prediction error signals, brought about by an increased signal-to-noise ratio of phasic 5-HT bursts [99], or attenuated punishment-induced response suppression, which can be defined as an instrumental process that inhibits behavior by virtue of the link between responses and the aversive outcomes they produce and a Pavlovian process that reflexively suppresses behavior [100].
In 2017, Rychlik and colleagues reported the role of glutamatergic neurotransmission in mediating sensitivity to feedback [101]. In this study, acute treatment with the prototypic, fast-acting antidepressant, N-methyl-D-aspartate (NMDA) receptor antagonist ketamine, significantly and persistently diminished the sensitivity of rats to negative feedback in the preclinical version of the PRL paradigm in a manner similar to that observed following the administration of higher doses of the SSRI citalopram [95].
Recent research with a pharmacogenetic model of reduced neurogenesis and a translationally relevant PRL paradigm demonstrated novel functions for adult-born neurons in sensitivity to rewards and negative feedback [102]. In that study, transgenic male rats that lacked adult neurogenesis were impaired in the use of probabilistic reward feedback to guide choice toward more profitable options. The observed effect was speculated to be due to either the specific loss of newborn neurons or to downstream changes that may have arisen over several weeks of neurogenesis ablation and pointed at hippocampal function in the modulation of sensitivity to feedback [102].
A study by Vaselic and colleagues revealed that reward processing and learning can be influenced by the sex hormone estradiol, which increases sensitivity to positive feedback and that these effects can be partially modulated by striatal DA transporter (DAT1) genes and personality traits related to reward sensitivity [103]. Similar feedback modulating effects were observed following acute administration of the neuropeptide oxytocin, which acutely increased the sensitivity of rats to positive feedback [104].
The studies by Bryce and Floresco [105] and Dieterich [106] revealed the role of stress hormones in sensitivity to feedback. In the former, increased corticotropin releasing factor (CRF) signaling reduced negative feedback sensitivity in rats, while in the latter, chronic corticosterone administration decreased the sensitivity of mice to positive feedback, which has been interpreted as a robust blunting of positive processing [106].
4. Implications for Treatment
Contemporary pharmacological treatment strategies for depressive disorder are directed at drugs that block the reuptake of 5-HT (the SSRIs) and/or NA (the SNRIs) from the extracellular space, which is thought to enhance the neural activity of these systems over time. Although clinical evidence clearly shows that the beneficial effects of these drugs occur with prolonged (min. 4–6 weeks) treatment, this delay in antidepressant action seems to be counterintuitive, as the molecular, cellular and chemical effects occur very quickly after a drug is administered, and recent studies have demonstrated that beneficial effects on biased cognition are evident even after the first dose. The cognitive neuropsychological theory of antidepressant action [107,108] was developed in an attempt to understand this potential paradox and states that the induction of a more positive way of processing environmental stimuli (positive bias) leads to cognitive and psychological reconsolidation [107]. This theory is consistent with cognitive models of depression, which emphasize the importance of correcting negative biases in information processing in the successful treatment of this disorder [23,109]. Indeed, there is now a growing body of experimental evidence indicating that by targeting cognitive biases, antidepressants can affect emotional processing very early in treatment and independently of changes in subjective mood (reviewed by Serra and colleagues [110]). Studies in animals confirmed that acute administration of several widely prescribed antidepressants changes cognitive judgment bias in the ACI test (citalopram [35], desipramine [35] and reboxetine [36]) and the sensitivity of rats to performance feedback in the preclinical version of the PRL task (citalopram [95] and ketamine [101]). Monitoring this positive shift in emotional processing creates an opportunity for fast detection of agents with antidepressant potential. This could be applied to new molecules in the early phase of development or the repurposing of existing drugs. As mentioned by Godlewska and Harmer [111], the use of this simple measure could also allow relatively inexpensive screening of the best treatment regimen by testing different doses or treatment periods in smaller groups of individuals before running large clinical trials. Another emerging opportunity associated with studying the effects of antidepressant drugs on cognitive biases is to explain the mode of action of new fast-acting antidepressants, such as the NMDA receptor antagonist ketamine, which acts on mood within hours. It has been proposed that the impact on emotional processing by this group of drugs is different than that of conventional antidepressant drugs and that they might act by blocking the retrieval of negative memory associations. This effect could also modulate sensitivity to negative feedback [101].
5. Conclusions and Future Directions
The aforementioned advances in studying neuromolecular correlates of biased cognition in depression are both exciting and timely. We are sure that the next few years will see exciting discoveries emerge from a new focus on this level of inquiry. This progress must evolve from the collection of experimental data from various brain areas and neural networks through the integration of a biologically grounded theoretical framework with deeply analogous animal research using the newest neuromolecular techniques to human cognitive neuroscience and clinical psychiatry. Indeed, current single-cell genomic technology already allows us to obtain new molecular mechanistic insights from the brains of depressed patients. Other emerging techniques include transcranial focused ultrasound/radiation methods. These approaches can be improved by combining them with the molecular mechanisms suggested in our manuscript. It seems also crucial to combine the levels of inquiry tackling at the same time molecular mechanisms and whole circuits. When seeking a better understanding of the neuromolecular background of cognitive biases in depression, we need to actively commit to synergy.
Author Contributions
All authors contributed to conceptualization and writing of this manuscript, as well as have read and agreed to its published version. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Statutory Funds of the Maj Institute of Pharmacology Polish Academy of Sciences.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Smith, K. Mental health: A world of depression. Nature 2014, 515, 180–181. [Google Scholar] [CrossRef]
- Greenberg, P.E.; Fournier, A.A.; Sisitsky, T.; Pike, C.T.; Kessler, R.C. The economic burden of adults with major depressive disorder in the United States (2005 and 2010). J. Clin. Psychiatry 2015, 76, 155–162. [Google Scholar] [CrossRef] [Green Version]
- DSM-5. Diagnostic and Statistical Manual of Mental Disorders; American Psychiatric Association: Washington, DC, USA, 2013. [Google Scholar]
- Chamberlain, S.R.; Sahakian, B.J. Cognition in mania and depression: Psychological models and clinical implications. Curr. Psychiatry Rep. 2004, 6, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Brittlebank, A.D.; Scott, J.; Williams, J.M.; Ferrier, I.N. Autobiographical memory in depression: State or trait marker? Br. J. Psychiatry 1993, 162, 118–121. [Google Scholar] [CrossRef] [PubMed]
- Rubinow, D.R.; Post, R.M. Impaired recognition of affect in facial expression in depressed patients. Biol. Psychiatry 1992, 31, 947–953. [Google Scholar] [CrossRef]
- Murphy, F.C.; Sahakian, B.J.; Rubinsztein, J.S.; Michael, A.; Rogers, R.D.; Robbins, T.W.; Paykel, E.S. Emotional bias and inhibitory control processes in mania and depression. Psychol. Med. 1999, 29, 1307–1321. [Google Scholar] [CrossRef]
- Douglas, K.M.; Porter, R.J. Recognition of disgusted facial expressions in severe depression. Br. J. Psychiatry 2010, 197, 156–157. [Google Scholar] [CrossRef] [Green Version]
- Maniglio, R.; Gusciglio, F.; Lofrese, V.; Belvederi Murri, M.; Tamburello, A.; Innamorati, M. Biased processing of neutral facial expressions is associated with depressive symptoms and suicide ideation in individuals at risk for major depression due to affective temperaments. Compr. Psychiatry 2014, 55, 518–525. [Google Scholar] [CrossRef]
- Beats, B.C.; Sahakian, B.J.; Levy, R. Cognitive performance in tests sensitive to frontal lobe dysfunction in the elderly depressed. Psychol. Med. 1996, 26, 591–603. [Google Scholar] [CrossRef] [PubMed]
- Elliott, R.; Sahakian, B.J.; Herrod, J.J.; Robbins, T.W.; Paykel, E.S. Abnormal response to negative feedback in unipolar depression: Evidence for a diagnosis specific impairment. J. Neurol Neurosurg. Psychiatry 1997, 63, 74–82. [Google Scholar] [CrossRef]
- Clark, L.; Chamberlain, S.R.; Sahakian, B.J. Neurocognitive mechanisms in depression: Implications for treatment. Annu. Rev. Neurosci. 2009, 32, 57–74. [Google Scholar] [CrossRef] [Green Version]
- Noonan, M.P.; Kolling, N.; Walton, M.E.; Rushworth, M.F. Re-evaluating the role of the orbitofrontal cortex in reward and reinforcement. Eur. J. Neurosci. 2012, 35, 997–1010. [Google Scholar] [CrossRef]
- Wheeler, E.Z.; Fellows, L.K. The human ventromedial frontal lobe is critical for learning from negative feedback. Brain 2008, 131, 1323–1331. [Google Scholar] [CrossRef] [PubMed]
- O’Doherty, J.; Dayan, P.; Schultz, J.; Deichmann, R.; Friston, K.; Dolan, R.J. Dissociable roles of ventral and dorsal striatum in instrumental conditioning. Science 2004, 304, 452–454. [Google Scholar] [CrossRef] [Green Version]
- Palminteri, S.; Justo, D.; Jauffret, C.; Pavlicek, B.; Dauta, A.; Delmaire, C.; Czernecki, V.; Karachi, C.; Capelle, L.; Durr, A.; et al. Critical roles for anterior insula and dorsal striatum in punishment-based avoidance learning. Neuron 2012, 76, 998–1009. [Google Scholar] [CrossRef] [Green Version]
- Tricomi, E.; Balleine, B.W.; O’Doherty, J.P. A specific role for posterior dorsolateral striatum in human habit learning. Eur. J. Neurosci. 2009, 29, 2225–2232. [Google Scholar] [CrossRef] [Green Version]
- Carver, C.S.; Scheier, M.F. Dispositional optimism. Trends Cogn. Sci. 2014, 18, 293–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheier, M.F.; Carver, C.S. Dispositional optimism and physical health: A long look back, a quick look forward. Am. Psychol. 2018, 73, 1082. [Google Scholar] [CrossRef] [PubMed]
- Maier, S.F.; Seligman, M.E. Learned helplessness: Theory and evidence. J. Exp. Psychol. Gen. 1976, 105, 3–46. [Google Scholar] [CrossRef]
- Abramson, L.Y.; Seligman, M.E.; Teasdale, J.D. Learned helplessness in humans: Critique and reformulation. J. Abnorm. Psychol. 1978, 87, 49–74. [Google Scholar] [CrossRef]
- Beck, A.T. Depression: Clinical, Experimental, and Theoretical Aspects; Harper and Row: New York, NY, USA, 1967. [Google Scholar]
- Beck, A.T. The evolution of the cognitive model of depression and its neurobiological correlates. Am. J. Psychiatry 2008, 165, 969–977. [Google Scholar] [CrossRef]
- Alloy, L.B.; Ahrens, A.H. Depression and pessimism for the future: Biased use of statistically relevant information in predictions for self versus others. J. Pers. Soc. Psychol. 1987, 52, 366–378. [Google Scholar] [CrossRef]
- Korn, C.W.; Sharot, T.; Walter, H.; Heekeren, H.R.; Dolan, R.J. Depression is related to an absence of optimistically biased belief updating about future life events. Psychol. Med. 2014, 44, 579–592. [Google Scholar] [CrossRef] [Green Version]
- Zenger, M.; Glaesmer, H.; Höckel, M.; Hinz, A. Pessimism Predicts Anxiety, Depression and Quality of Life in Female Cancer Patients. Jpn. J. Clin. Oncol. 2010, 41, 87–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papciak, J.; Popik, P.; Fuchs, E.; Rygula, R. Chronic psychosocial stress makes rats more ‘pessimistic’ in the ambiguous-cue interpretation paradigm. Behav. Brain Res. 2013, 256, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Rygula, R.; Papciak, J.; Popik, P. Trait pessimism predicts vulnerability to stress-induced anhedonia in rats. Neuropsychopharmacology 2013, 38, 2188–2196. [Google Scholar] [CrossRef] [Green Version]
- Pessiglione, M.; Seymour, B.; Flandin, G.; Dolan, R.J.; Frith, C.D. Dopamine-dependent prediction errors underpin reward-seeking behaviour in humans. Nature 2006, 442, 1042–1045. [Google Scholar] [CrossRef] [Green Version]
- Schultz, W.; Tremblay, L.; Hollerman, J.R. Reward prediction in primate basal ganglia and frontal cortex. Neuropharmacology 1998, 37, 421–429. [Google Scholar] [CrossRef]
- Sharot, T.; Guitart-Masip, M.; Korn, C.W.; Chowdhury, R.; Dolan, R.J. How dopamine enhances an optimism bias in humans. Curr. Biol. 2012, 22, 1477–1481. [Google Scholar] [CrossRef] [Green Version]
- Vellani, V.; de Vries, L.P.; Gaule, A.; Sharot, T. A selective effect of dopamine on information-seeking. Elife 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Enkel, T.; Gholizadeh, D.; von Bohlen Und Halbach, O.; Sanchis-Segura, C.; Hurlemann, R.; Spanagel, R.; Gass, P.; Vollmayr, B. Ambiguous-cue interpretation is biased under stress- and depression-like states in rats. Neuropsychopharmacology 2010, 35, 1008–1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harding, E.J.; Paul, E.S.; Mendl, M. Animal behaviour: Cognitive bias and affective state. Nature 2004, 427, 312. [Google Scholar] [CrossRef]
- Rygula, R.; Papciak, J.; Popik, P. The effects of acute pharmacological stimulation of the 5-HT, NA and DA systems on the cognitive judgement bias of rats in the ambiguous-cue interpretation paradigm. Eur. Neuropsychopharmacol. J. Eur. Coll. Neuropsychopharmacol. 2014, 24, 1103–1111. [Google Scholar] [CrossRef] [PubMed]
- Hales, C.A.; Houghton, C.J.; Robinson, E.S.J. Behavioural and computational methods reveal differential effects for how delayed and rapid onset antidepressants effect decision making in rats. Eur. Neuropsychopharmacol. 2017, 27, 1268–1280. [Google Scholar] [CrossRef]
- Rygula, R.; Szczech, E.; Papciak, J.; Nikiforuk, A.; Popik, P. The effects of cocaine and mazindol on the cognitive judgement bias of rats in the ambiguous-cue interpretation paradigm. Behav. Brain Res. 2014, 270, 206–212. [Google Scholar] [CrossRef]
- Golebiowska, J.; Rygula, R. Effects of acute dopaminergic and serotonergic manipulations in the ACI paradigm depend on the basal valence of cognitive judgement bias in rats. Behav. Brain Res. 2017, 327, 133–143. [Google Scholar] [CrossRef]
- Sharp, T.; Foster, G.A. In vivo measurement using microdialysis of the release and metabolism of 5-hydroxytryptamine in raphe neurones grafted to the rat hippocampus. J. Neurochem. 1989, 53, 303–306. [Google Scholar] [CrossRef] [PubMed]
- Sprouse, J.S.; Aghajanian, G.K. Electrophysiological responses of serotoninergic dorsal raphe neurons to 5-HT1A and 5-HT1B agonists. Synapse 1987, 1, 3–9. [Google Scholar] [CrossRef]
- Bymaster, F.P.; Zhang, W.; Carter, P.A.; Shaw, J.; Chernet, E.; Phebus, L.; Wong, D.T.; Perry, K.W. Fluoxetine, but not other selective serotonin uptake inhibitors, increases norepinephrine and dopamine extracellular levels in prefrontal cortex. Psychopharmacology 2002, 160, 353–361. [Google Scholar] [CrossRef]
- Koch, S.; Perry, K.W.; Nelson, D.L.; Conway, R.G.; Threlkeld, P.G.; Bymaster, F.P. R-fluoxetine increases extracellular DA, NE, as well as 5-HT in rat prefrontal cortex and hypothalamus: An in vivo microdialysis and receptor binding study. Neuropsychopharmacology 2002, 27, 949–959. [Google Scholar] [CrossRef] [Green Version]
- Invernizzi, R.; Belli, S.; Samanin, R. Citalopram’s ability to increase the extracellular concentrations of serotonin in the dorsal raphe prevents the drug’s effect in the frontal cortex. Brain Res 1992, 584, 322–324. [Google Scholar] [CrossRef]
- Pozzi, L.; Invernizzi, R.; Garavaglia, C.; Samanin, R. Fluoxetine increases extracellular dopamine in the prefrontal cortex by a mechanism not dependent on serotonin: A comparison with citalopram. J. Neurochem. 1999, 73, 1051–1057. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.H.; Munafo, M.R.; Robinson, E.S. Investigating the psychopharmacology of cognitive affective bias in rats using an affective tone discrimination task. Psychopharmacology 2013, 226, 601–613. [Google Scholar] [CrossRef]
- Doyle, R.E.; Hinch, G.N.; Fisher, A.D.; Boissy, A.; Henshall, J.M.; Lee, C. Administration of serotonin inhibitor p-Chlorophenylalanine induces pessimistic-like judgement bias in sheep. Psychoneuroendocrinology 2011, 36, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Stracke, J.; Otten, W.; Tuchscherer, A.; Puppe, B.; Düpjan, S. Serotonin depletion induces pessimistic-like behavior in a cognitive bias paradigm in pigs. Physiol. Behav. 2017, 174, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Curzytek, K.; Kubera, M.; Trojan, E.; Wojcik, K.; Basta-Kaim, A.; Detka, J.; Maes, M.; Rygula, R. The effects of pessimism on cell-mediated immunity in rats. Prog. Neuropsychopharmacol. Biol. Psychiatry 2018, 80, 295–303. [Google Scholar] [CrossRef]
- Drozd, R.; Cieslak, P.E.; Rychlik, M.; Rodriguez Parkitna, J.; Rygula, R. Cognitive Judgment Bias Interacts with Risk Based Decision Making and Sensitivity to Dopaminergic Challenge in Male Rats. Front. Behav. Neurosci. 2016, 10, 163. [Google Scholar] [CrossRef] [Green Version]
- Drozd, R.; Rojek-Sito, K.; Rygula, R. The trait ‘pessimism’ does not interact with cognitive flexibility but makes rats more vulnerable to stress-induced motivational deficits: Results from the attentional set-shifting task. Behav. Brain Res. 2017, 335, 199–207. [Google Scholar] [CrossRef]
- Rafa, D.; Kregiel, J.; Popik, P.; Rygula, R. Effects of optimism on gambling in the rat slot machine task. Behav. Brain Res. 2016, 300, 97–105. [Google Scholar] [CrossRef]
- Rygula, R.; Golebiowska, J.; Kregiel, J.; Kubik, J.; Popik, P. Effects of optimism on motivation in rats. Front. Behav. Neurosci. 2015, 9, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espigares, F.; Abad-Tortosa, D.; Varela, S.A.M.; Ferreira, M.G.; Oliveira, R.F. Short telomeres drive pessimistic judgement bias in zebrafish. Biol. Lett. 2021, 17, 20200745. [Google Scholar] [CrossRef]
- Roy, B.; Diez-Roux, A.V.; Seeman, T.; Ranjit, N.; Shea, S.; Cushman, M. Association of optimism and pessimism with inflammation and hemostasis in the Multi-Ethnic Study of Atherosclerosis (MESA). Psychosom. Med. 2010, 72, 134–140. [Google Scholar] [CrossRef]
- Beevers, C.G.; Wells, T.T.; Ellis, A.J.; McGeary, J.E. Association of the serotonin transporter gene promoter region (5-HTTLPR) polymorphism with biased attention for emotional stimuli. J. Abnorm Psychol. 2009, 118, 670–681. [Google Scholar] [CrossRef] [Green Version]
- Fox, E.; Ridgewell, A.; Ashwin, C. Looking on the bright side: Biased attention and the human serotonin transporter gene. Proc. Biol. Sci. 2009, 276, 1747–1751. [Google Scholar] [CrossRef] [PubMed]
- Kloke, V.; Schreiber, R.S.; Bodden, C.; Möllers, J.; Ruhmann, H.; Kaiser, S.; Lesch, K.-P.; Sachser, N.; Lewejohann, L. Hope for the best or prepare for the worst? Towards a spatial cognitive bias test for mice. PLoS ONE 2014, 9, e105431. [Google Scholar] [CrossRef] [Green Version]
- Krakenberg, V.; von Kortzfleisch, V.T.; Kaiser, S.; Sachser, N.; Richter, S.H. Differential Effects of Serotonin Transporter Genotype on Anxiety-Like Behavior and Cognitive Judgment Bias in Mice. Front. Behav. Neurosci. 2019, 13, 263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boddington, R.; Gómez Dunlop, C.A.; Garnham, L.C.; Ryding, S.; Abbey-Lee, R.N.; Kreshchenko, A.; Løvlie, H. The relationship between monoaminergic gene expression, learning, and optimism in red junglefowl chicks. Anim. Cogn. 2020, 23, 901–911. [Google Scholar] [CrossRef]
- Spasojević, J.; Alloy, L.B. Rumination as a common mechanism relating depressive risk factors to depression. Emotion 2001, 1, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Murphy, F.C.; Michael, A.; Robbins, T.W.; Sahakian, B.J. Neuropsychological impairment in patients with major depressive disorder: The effects of feedback on task performance. Psychol. Med. 2003, 33, 455–467. [Google Scholar] [CrossRef]
- Taylor Tavares, J.V.; Clark, L.; Furey, M.L.; Williams, G.B.; Sahakian, B.J.; Drevets, W.C. Neural basis of abnormal response to negative feedback in unmedicated mood disorders. Neuroimage 2008, 42, 1118–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rygula, R.; Noworyta-Sokolowska, K.; Drozd, R.; Kozub, A. Using rodents to model abnormal sensitivity to feedback in depression. Neurosci. Biobehav. Rev. 2018, 95, 336–346. [Google Scholar] [CrossRef]
- Henriques, J.B.; Glowacki, J.M.; Davidson, R.J. Reward fails to alter response bias in depression. J. Abnorm Psychol. 1994, 103, 460–466. [Google Scholar] [CrossRef]
- McFarland, B.R.; Klein, D.N. Emotional reactivity in depression: Diminished responsiveness to anticipated reward but not to anticipated punishment or to nonreward or avoidance. Depress Anxiety 2009, 26, 117–122. [Google Scholar] [CrossRef]
- Robinson, O.J.; Cools, R.; Carlisi, C.O.; Sahakian, B.J.; Drevets, W.C. Ventral striatum response during reward and punishment reversal learning in unmedicated major depressive disorder. Am. J. Psychiatry 2012, 169, 152–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gotlib, I.H.; Joormann, J. Cognition and depression: Current status and future directions. Annu. Rev. Clin. Psychol. 2010, 6, 285–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roiser, J.P.; Sahakian, B.J. Hot and cold cognition in depression. CNS Spectr. 2013, 18, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Steele, J.D.; Kumar, P.; Ebmeier, K.P. Blunted response to feedback information in depressive illness. Brain 2007, 130, 2367–2374. [Google Scholar] [CrossRef] [PubMed]
- Pizzagalli, D.A.; Goetz, E.; Ostacher, M.; Iosifescu, D.V.; Perlis, R.H. Euthymic patients with bipolar disorder show decreased reward learning in a probabilistic reward task. Biol. Psychiatry 2008, 64, 162–168. [Google Scholar] [CrossRef] [Green Version]
- Pizzagalli, D.A.; Iosifescu, D.; Hallett, L.A.; Ratner, K.G.; Fava, M. Reduced hedonic capacity in major depressive disorder: Evidence from a probabilistic reward task. J. Psychiatr. Res. 2008, 43, 76–87. [Google Scholar] [CrossRef] [Green Version]
- Frank, M.J.; Seeberger, L.C.; O’Reilly, R.C. By carrot or by stick: Cognitive reinforcement learning in parkinsonism. Science 2004, 306, 1940–1943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frank, M.J. Dynamic dopamine modulation in the basal ganglia: A neurocomputational account of cognitive deficits in medicated and nonmedicated Parkinsonism. J. Cogn. Neurosci. 2005, 17, 51–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisenegger, C.; Naef, M.; Linssen, A.; Clark, L.; Gandamaneni, P.K.; Müller, U.; Robbins, T.W. Role of dopamine D2 receptors in human reinforcement learning. Neuropsychopharmacology 2014, 39, 2366–2375. [Google Scholar] [CrossRef]
- Frank, M.J.; Hutchison, K. Genetic contributions to avoidance-based decisions: Striatal D2 receptor polymorphisms. Neuroscience 2009, 164, 131–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, S.M.; Frank, M.J.; Larcher, K.; Fellows, L.K.; Clark, C.A.; Leyton, M.; Dagher, A. Striatal D1 and D2 signaling differentially predict learning from positive and negative outcomes. Neuroimage 2015, 109, 95–101. [Google Scholar] [CrossRef]
- Hikida, T.; Kimura, K.; Wada, N.; Funabiki, K.; Nakanishi, S. Distinct Roles of Synaptic Transmission in Direct and Indirect Striatal Pathways to Reward and Aversive Behavior. Neuron 2010, 66, 896–907. [Google Scholar] [CrossRef] [Green Version]
- Kravitz, A.V.; Tye, L.D.; Kreitzer, A.C. Distinct roles for direct and indirect pathway striatal neurons in reinforcement. Nat. Neurosci. 2012, 15, 816–818. [Google Scholar] [CrossRef] [Green Version]
- Alsiö, J.; Phillips, B.U.; Sala-Bayo, J.; Nilsson, S.R.O.; Calafat-Pla, T.C.; Rizwand, A.; Plumbridge, J.M.; López-Cruz, L.; Dalley, J.W.; Cardinal, R.N.; et al. Dopamine D2-like receptor stimulation blocks negative feedback in visual and spatial reversal learning in the rat: Behavioural and computational evidence. Psychopharmacology 2019, 236, 2307–2323. [Google Scholar] [CrossRef] [Green Version]
- Verharen, J.P.H.; Adan, R.A.H.; Vanderschuren, L. Differential contributions of striatal dopamine D1 and D2 receptors to component processes of value-based decision making. Neuropsychopharmacology 2019, 44, 2195–2204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frank, M.J.; O’Reilly, R.C. A mechanistic account of striatal dopamine function in human cognition: Psychopharmacological studies with cabergoline and haloperidol. Behav. Neurosci. 2006, 120, 497–517. [Google Scholar] [CrossRef] [Green Version]
- Lim, T.V.; Cardinal, R.N.; Bullmore, E.T.; Robbins, T.W.; Ersche, K.D. Impaired Learning From Negative Feedback in Stimulant Use Disorder: Dopaminergic Modulation. Int. J. Neuropsychopharmacol. 2021, 24, 867–878. [Google Scholar] [CrossRef]
- McCabe, C.; Huber, A.; Harmer, C.J.; Cowen, P.J. The D2 antagonist sulpiride modulates the neural processing of both rewarding and aversive stimuli in healthy volunteers. Psychopharmacology 2011, 217, 271–278. [Google Scholar] [CrossRef] [Green Version]
- Jocham, G.; Klein, T.A.; Ullsperger, M. Differential modulation of reinforcement learning by D2 dopamine and NMDA glutamate receptor antagonism. J. Neurosci. 2014, 34, 13151–13162. [Google Scholar] [CrossRef] [Green Version]
- Cools, R.; Frank, M.J.; Gibbs, S.E.; Miyakawa, A.; Jagust, W.; D’Esposito, M. Striatal dopamine predicts outcome-specific reversal learning and its sensitivity to dopaminergic drug administration. J. Neurosci. 2009, 29, 1538–1543. [Google Scholar] [CrossRef] [Green Version]
- Frank, M.J.; Moustafa, A.A.; Haughey, H.M.; Curran, T.; Hutchison, K.E. Genetic triple dissociation reveals multiple roles for dopamine in reinforcement learning. Proc. Natl. Acad. Sci. USA 2007, 104, 16311–16316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, T.A.; Neumann, J.; Reuter, M.; Hennig, J.; von Cramon, D.Y.; Ullsperger, M. Genetically determined differences in learning from errors. Science 2007, 318, 1642–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jocham, G.; Klein, T.A.; Neumann, J.; von Cramon, D.Y.; Reuter, M.; Ullsperger, M. Dopamine DRD2 polymorphism alters reversal learning and associated neural activity. J. Neurosci. 2009, 29, 3695–3704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chamberlain, S.R.; Muller, U.; Blackwell, A.D.; Clark, L.; Robbins, T.W.; Sahakian, B.J. Neurochemical modulation of response inhibition and probabilistic learning in humans. Science 2006, 311, 861–863. [Google Scholar] [CrossRef] [Green Version]
- Skandali, N.; Rowe, J.B.; Voon, V.; Deakin, J.B.; Cardinal, R.N.; Cormack, F.; Passamonti, L.; Bevan-Jones, W.R.; Regenthal, R.; Chamberlain, S.R.; et al. Dissociable effects of acute SSRI (escitalopram) on executive, learning and emotional functions in healthy humans. Neuropsychopharmacology 2018, 43, 2645–2651. [Google Scholar] [CrossRef] [Green Version]
- Evers, E.A.; Cools, R.; Clark, L.; van der Veen, F.M.; Jolles, J.; Sahakian, B.J.; Robbins, T.W. Serotonergic modulation of prefrontal cortex during negative feedback in probabilistic reversal learning. Neuropsychopharmacology 2005, 30, 1138–1147. [Google Scholar] [CrossRef]
- Cools, R.; Robinson, O.J.; Sahakian, B. Acute tryptophan depletion in healthy volunteers enhances punishment prediction but does not affect reward prediction. Neuropsychopharmacology 2008, 33, 2291–2299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Den Ouden, H.E.; Daw, N.D.; Fernandez, G.; Elshout, J.A.; Rijpkema, M.; Hoogman, M.; Franke, B.; Cools, R. Dissociable effects of dopamine and serotonin on reversal learning. Neuron 2013, 80, 1090–1100. [Google Scholar] [CrossRef] [Green Version]
- Willeit, M.; Praschak-Rieder, N. Imaging the effects of genetic polymorphisms on radioligand binding in the living human brain: A review on genetic neuroreceptor imaging of monoaminergic systems in psychiatry. Neuroimage 2010, 53, 878–892. [Google Scholar] [CrossRef]
- Bari, A.; Theobald, D.E.; Caprioli, D.; Mar, A.C.; Aidoo-Micah, A.; Dalley, J.W.; Robbins, T.W. Serotonin modulates sensitivity to reward and negative feedback in a probabilistic reversal learning task in rats. Neuropsychopharmacology 2010, 35, 1290–1301. [Google Scholar] [CrossRef] [PubMed]
- Ineichen, C.; Sigrist, H.; Spinelli, S.; Lesch, K.P.; Sautter, E.; Seifritz, E.; Pryce, C.R. Establishing a probabilistic reversal learning test in mice: Evidence for the processes mediating reward-stay and punishment-shift behaviour and for their modulation by serotonin. Neuropharmacology 2012, 63, 1012–1021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rygula, R.; Clarke, H.F.; Cardinal, R.N.; Cockcroft, G.J.; Xia, J.; Dalley, J.W.; Robbins, T.W.; Roberts, A.C. Role of Central Serotonin in Anticipation of Rewarding and Punishing Outcomes: Effects of Selective Amygdala or Orbitofrontal 5-HT Depletion. Cereb. Cortex. 2015, 25, 3064–3076. [Google Scholar] [CrossRef]
- Phillips, B.U.; Dewan, S.; Nilsson, S.R.O.; Robbins, T.W.; Heath, C.J.; Saksida, L.M.; Bussey, T.J.; Alsio, J. Selective effects of 5-HT2C receptor modulation on performance of a novel valence-probe visual discrimination task and probabilistic reversal learning in mice. Psychopharmacology 2018, 235, 2101–2111. [Google Scholar] [CrossRef] [Green Version]
- Cools, R.; Roberts, A.C.; Robbins, T.W. Serotoninergic regulation of emotional and behavioural control processes. Trends Cogn. Sci. 2008, 12, 31–40. [Google Scholar] [CrossRef]
- Robinson, O.J.; Cools, R.; Sahakian, B.J. Tryptophan depletion disinhibits punishment but not reward prediction: Implications for resilience. Psychopharmacology 2012, 219, 599–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rychlik, M.; Bollen, E.; Rygula, R. Ketamine decreases sensitivity of male rats to misleading negative feedback in a probabilistic reversal-learning task. Psychopharmacology 2017, 234, 613–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seib, D.R.; Espinueva, D.F.; Floresco, S.B.; Snyder, J.S. A role for neurogenesis in probabilistic reward learning. Behav. Neurosci. 2020, 134, 283–295. [Google Scholar] [CrossRef]
- Veselic, S.; Jocham, G.; Gausterer, C.; Wagner, B.; Ernhoefer-Reßler, M.; Lanzenberger, R.; Eisenegger, C.; Lamm, C.; Losecaat Vermeer, A. A causal role of estradiol in human reinforcement learning. Horm. Behav. 2021, 134, 105022. [Google Scholar] [CrossRef] [PubMed]
- Roberts, B.Z.; Young, J.W.; He, Y.V.; Cope, Z.A.; Shilling, P.D.; Feifel, D. Oxytocin improves probabilistic reversal learning but not effortful motivation in Brown Norway rats. Neuropharmacology 2019, 150, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Bryce, C.A.; Floresco, S.B. Central CRF and acute stress differentially modulate probabilistic reversal learning in male and female rats. Behav. Brain Res. 2021, 397, 112929. [Google Scholar] [CrossRef] [PubMed]
- Dieterich, A.; Srivastava, P.; Sharif, A.; Stech, K.; Floeder, J.; Yohn, S.E.; Samuels, B.A. Chronic corticosterone administration induces negative valence and impairs positive valence behaviors in mice. Transl. Psychiatry 2019, 9, 337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harmer, C.J.; Goodwin, G.M.; Cowen, P.J. Why do antidepressants take so long to work? A cognitive neuropsychological model of antidepressant drug action. Br. J. Psychiatry 2009, 195, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Godlewska, B.R. Cognitive neuropsychological theory: Reconciliation of psychological and biological approaches for depression. Pharm. Ther. 2019, 197, 38–51. [Google Scholar] [CrossRef]
- Beck, A.T. Cognitive therapy. A 30-year retrospective. Am. Psychol. 1991, 46, 368–375. [Google Scholar] [CrossRef]
- Serra, M.; Salgado-Pineda, P.; Delaveau, P.; Fakra, E.; Gasto, C.; Blin, O. Effects of antidepressant drugs on emotion. Clin. Neuropharmacol. 2006, 29, 170–185. [Google Scholar] [CrossRef]
- Godlewska, B.R.; Harmer, C.J. Cognitive neuropsychological theory of antidepressant action: A modern-day approach to depression and its treatment. Psychopharmacology 2021, 238, 1265–1278. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Neuromolecular and cellular correlates of biased judgment.

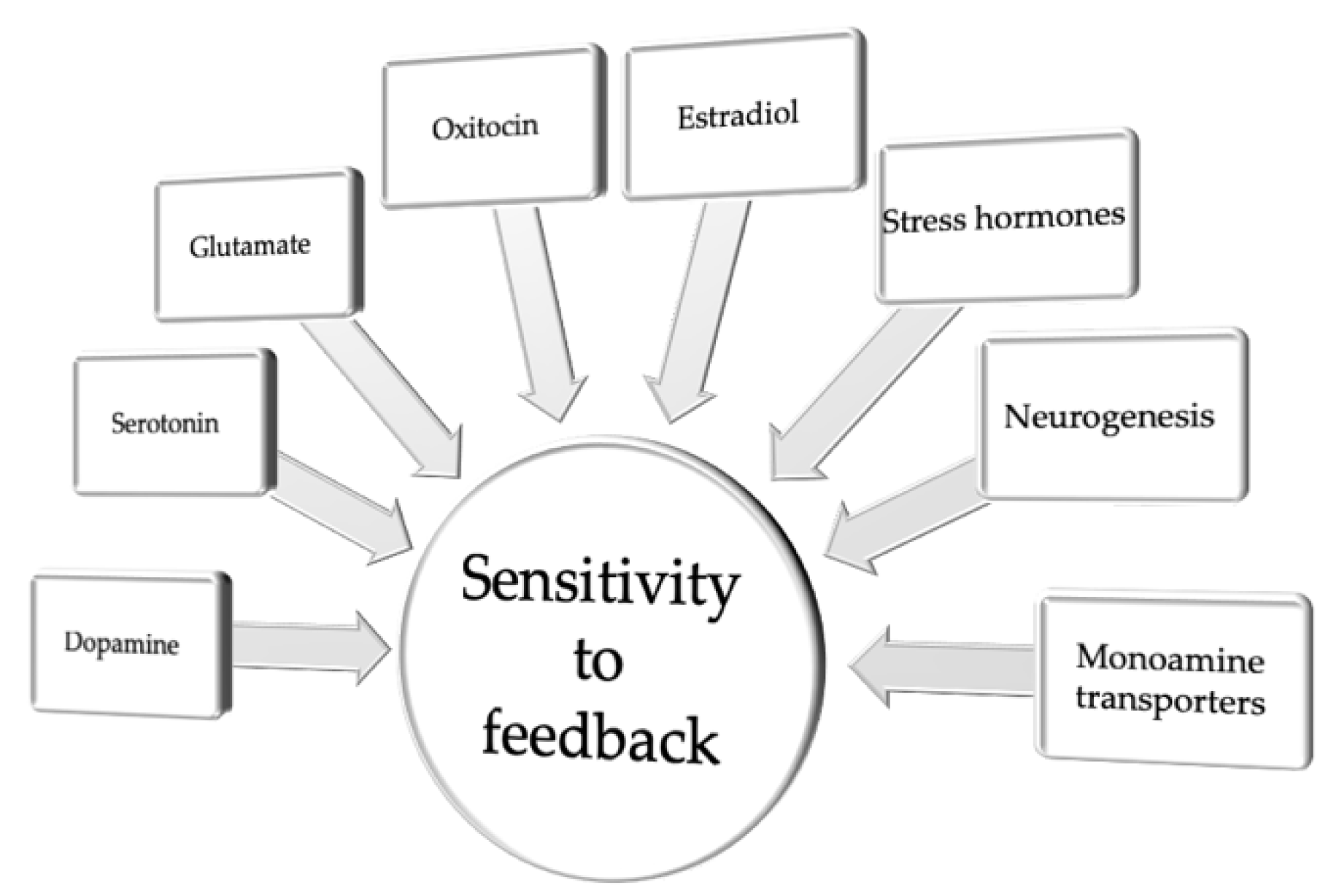
Figure 2.
Neuromolecular and cellular correlates of sensitivity to feedback.


{kind=link}
{kind=link}
Table 1.
Pharmacological manipulations targeting various neurotransmitter systems, which affect pessimistic judgment bias or sensitivity to feedback.
Table 1.
Pharmacological manipulations targeting various neurotransmitter systems, which affect pessimistic judgment bias or sensitivity to feedback.
| Pessimistic Judgment Bias | |||||
|---|---|---|---|---|---|
| Target System | Drug Used | Study Subject | Test | Behavioral Outcome | Reference |
| DA | L-DOPA | Human | BUT | Impaired ability to update belief in response to undesirable information about the future, higher optimism | [31] |
| DA | L-DOPA | Human | SMT | Shift bias toward the information about gains | [32] |
| DA | d-amph | Rat | ACI | Optimism | [35] |
| DA | L-DOPA | Rat | ACI | Pessimistic shift in animals classified as optimistic | [38] |
| DA | Halo | Rat | ACI | Optimists became more pessimistic, while pessimists became more optimistic | [38] |
| 5-HT | Escit | Rat | ACI | Pessimistic shift in animals classified as optimistic | [38] |
| 5-HT | Cit | Human | BUT | No effect | [31] |
| 5-HT | Cit | Rat | ACI | Negative interpretation of ambiguous cues (a low dose) or optimistic judgment bias (a high dose) | [35] |
| 5-HT | Flx | Rat | ATDT | Pro-optimistic effects of chronic treatment | [45] |
| 5-HT | Flx | Rat | ACI | Pro-optimistic effects of chronic treatment | [36] |
| 5-HT | pCPA | Sheep | SDT | Pessimistic judgment bias | [46] |
| 5-HT | pCPA | Pig | ACI | Pessimistic judgment bias | [47] |
| NA | Desi | Rat | ACI | Pessimistic judgment bias | [35] |
| NA/DA | Mazin | Rat | ACI | Pessimistic judgment bias | [37] |
| NA | Rbx | Rat | ACI | Decrease in the positive processing | [45] |
| NA | Rbx, Cort | Rat | ACI | Pessimistic judgment bias | [33] |
| Sensitivity to feedback | |||||
| Target System | Drug Used | Study Subject | Test | Behavioral Outcome | Reference |
| DA | L-DOPA | Human | PLT | Higher sensitivity to positive than negative outcomes in PD patients on medication | [72] |
| DA | L-DOPA, Halo | Human | G/NG | Subjects treated with L-DOPA have a greater propensity to choose the most rewarding action relative to subjects treated with haloperidol | [29] |
| DA | Sulp | Human | RLT | Impairment in reward choice performance | [74] |
| DA | APTD | Human | PST | Improved learning from negative outcomes | [76] |
| DA | Quin | Rat | SPRL | Impaired learning from negative feedback | [79] |
| DA | Raclo, Quin | Rat | PRL | Negative feedback learning depends on D2R signaling, whereas learning from positive feedback depends on D1R signaling | [80] |
| DA | Halo | Human | PST | Increased DA release during positive feedback enhanced Go learning for good choices | [81] |
| DA | Ami, Prami | Human | RL | Impaired learning from negative feedback | [82] |
| 5-HT | Cit | Human | PRL | Low dose increased tendency to switch the response following negative feedback | [89] |
| 5-HT | Escit | Human | PRL | Impaired learning with uncertain reinforcement and enhanced responsivity to negative feedback | [90] |
| 5-HT | ATD | Human | PRL | Increased punishment prediction | [92] |
| 5-HT | SB 242084 | Mice | PRL | Reduced sensitivity to positive feedback | [98] |
| 5-HT | WAY 163909 | Mice | PRL | Increased sensitivity to positive feedback and decreased sensitivity to negative feedback | [98] |
| glu | Ket | Rat | PRL | Diminished the sensitivity of rats to negative feedback | [101] |
5-HT—serotonin, ACI—the ambiguous interpretation test, Ami—amisulpride, APTD—acute phenylalanine and tyrosine depletion, ATD—acute tryptophan depletion, ATDT—the affective tone discrimination task, d-amph—d-amphetamine, BUT—the belief updating test, Cit—citalopram, Cort—corticosterone, D1R—dopamine D1 receptor, D2R—dopamine D2 receptor, DA—dopamine, Desi—desipramine, Escit—escitalopram, Flx—fluoxetine, glu—glutamate, G/NG—Go/NoGo task, Halo—haloperidol, Ket—ketamine, L-DOPA—3,4-dihydroxy-L-phenylalanine, Mazin—maziodol, NA—noradrenaline, pCPA—p-Chlorophenylalanine, PD—Parkinson’s disease, PLT—the procedural learning task, Prami—pramipeskole, PRL—the probabilistic reversal learning test, PST—the probabilistic selection task, Quin—quinpirole, Raclo—raclopride, Rbx—reboxetine, RL—reversal learning taks, RLT—the reinforcement learning task, SB 242084—selective antagonist of the 5-HT2C receptor, SDT—the spatial differentiation task, SMT—the stock market task, SPRL—the spatial reversal learning test, Sulp—sulpiride, WAY 163909—selective agonist for the serotonin 5-HT2C receptor.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Noworyta, K.; Cieslik, A.; Rygula, R. Neuromolecular Underpinnings of Negative Cognitive Bias in Depression. Cells 2021, 10, 3157. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10113157
AMA Style
Noworyta K, Cieslik A, Rygula R. Neuromolecular Underpinnings of Negative Cognitive Bias in Depression. Cells. 2021; 10(11):3157. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10113157
Chicago/Turabian StyleNoworyta, Karolina, Agata Cieslik, and Rafal Rygula. 2021. "Neuromolecular Underpinnings of Negative Cognitive Bias in Depression" Cells 10, no. 11: 3157. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10113157
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.