
Impact of Pharmacological and Non-Pharmacological Modulators on Dendritic Spines Structure and Functions in Brain


 , ,
, ,
Abstract
:1. Introduction
2. Dendritic Spines Biosynthesis
3. Effects of Pharmacological Interventions on Dendritic Spines
3.1. Cholinergic System Modulators
3.2. Effect of Glutamatergic Modulators on Dendritic Spines
3.3. Effect of GABAergic Agents on Dendritic Spines
3.4. Effect of Serotonergic Agents on Dendritic Spines
3.5. Effect of Adrenergic Agents on Dendritic Spines
3.6. Effect of Dopaminergic Agents on Dendritic Spines
4. Non-Therapeutic Modulators of Dendritic Spines
4.1. Impact of Calorie Restriction on Dendritic Spine
4.2. Effect of Pre-, Pro-, and Syn-Biotics on Dendritic Spines
4.3. Effect of Enriched Environment (EE) on Dendritic Spines
4.4. Effects of Yoga and Meditation on Dendritic Spine
- Dendritic Spine sprouting: “Recurving of distal dendritic arbors, short-segment branching, growth cone-like processes initiating at the neuronal dendritic process is spine sprouting” [39].
- Siddha medicine: “Siddha is an ancient traditional Indian system of medicine practiced in southern part of India. Siddha medicine is claimed to revitalize and rejuvenate dysfunctional organs that cause the disease. Kayakarpam, a special combination of medicine and life style, Varmam therapy, Vaasi (Pranayamam) and Muppu the universal Salt are the specialties of Siddha system of medicine. Thus, this system connects both spiritual and physical and treats the person as a whole i.e., it concentrates on the physical, psychological, social, and spiritual wellbeing of an individual” [180].
- Prebiotics: “Are the source of food for gut’s healthy bacteria or the food that induces growth or activity of beneficial microorganisms in the gut”.
- Probiotics: “Are the live microorganisms, when consumed improve or restore the gut flora”.
- Synbiotics: “Are mixture of both prebiotics and probiotics viz, fermented food that synergistically improve the gut flora” [181].
5. Conclusions
6. Limitations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McGuier, N.S.; Uys, J.D.; Mulholland, P.J. Chapter 9—Neural morphology and addiction. In Neural Mechanisms of Addiction; Torregrossa, M., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 123–135. ISBN 978-0-12-812202-0. [Google Scholar]
- Nimchinsky, E.A.; Sabatini, B.L.; Svoboda, K. Structure and function of dendritic spines. Annu. Rev. Physiol. 2002, 64, 313–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, A.; Proskauer, C.C.; Kaiserman-Abramof, I.R. The small pyramidal neuron of the rat cerebral cortex. The axon hillock and initial segment. J. Cell Biol. 1968, 39, 604–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rochefort, N.L.; Konnerth, A. Dendritic spines: From structure to in vivo function. EMBO Rep. 2012, 13, 699–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourne, J.; Harris, K.M. Do thin spines learn to be mushroom spines that remember? Curr. Opin. Neurobiol. 2007, 17, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Holtmaat, A.; Wilbrecht, L.; Knott, G.W.; Welker, E.; Svoboda, K. Experience-dependent and cell-type-specific spine growth in the neocortex. Nature 2006, 441, 979–983. [Google Scholar] [CrossRef] [PubMed]
- Paspalas, C.D.; Wang, M.; Arnsten, A.F.T. Constellation of HCN channels and CAMP regulating proteins in dendritic spines of the primate prefrontal cortex: Potential substrate for working memory deficits in schizophrenia. Cereb. Cortex 2013, 23, 1643–1654. [Google Scholar] [CrossRef] [Green Version]
- Engert, F.; Bonhoeffer, T. Dendritic spine changes associated with hippocampal long-term synaptic plasticity. Nature 1999, 399, 66–70. [Google Scholar] [CrossRef]
- Nägerl, U.V.; Eberhorn, N.; Cambridge, S.B.; Bonhoeffer, T. Bidirectional activity-dependent morphological plasticity in hippocampal neurons. Neuron 2004, 44, 759–767. [Google Scholar] [CrossRef] [Green Version]
- Kwon, H.-B.; Sabatini, B.L. Glutamate induces de novo growth of functional spines in developing cortex. Nature 2011, 474, 100–104. [Google Scholar] [CrossRef]
- Yuste, R.; Bonhoeffer, T. Morphological changes in dendritic spines associated with long-term synaptic plasticity. Annu. Rev. Neurosci. 2001, 24, 1071–1089. [Google Scholar] [CrossRef] [Green Version]
- Collin, C.; Miyaguchi, K.; Segal, M. Dendritic spine density and LTP induction in cultured hippocampal slices. J. Neurophysiol. 1997, 77, 1614–1623. [Google Scholar] [CrossRef] [Green Version]
- Meng, Y.; Zhang, Y.; Tregoubov, V.; Janus, C.; Cruz, L.; Jackson, M.; Lu, W.-Y.; MacDonald, J.F.; Wang, J.Y.; Falls, D.L.; et al. Abnormal spine morphology and enhanced LTP in LIMK-1 knockout mice. Neuron 2002, 35, 121–133. [Google Scholar] [CrossRef] [Green Version]
- Bosch, M.; Castro, J.; Saneyoshi, T.; Matsuno, H.; Sur, M.; Hayashi, Y. Structural and molecular remodeling of dendritic spine substructures during long-term potentiation. Neuron 2014, 82, 444–459. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Homma, K.J.; Poo, M. Shrinkage of dendritic spines associated with long-term depression of hippocampal synapses. Neuron 2004, 44, 749–757. [Google Scholar] [CrossRef] [Green Version]
- Hayama, T.; Noguchi, J.; Watanabe, S.; Takahashi, N.; Hayashi-Takagi, A.; Ellis-Davies, G.C.R.; Matsuzaki, M.; Kasai, H. GABA promotes the competitive selection of dendritic spines by controlling local Ca2+ signaling. Nat. Neurosci. 2013, 16, 1409–1416. [Google Scholar] [CrossRef]
- Oh, W.C.; Hill, T.C.; Zito, K. Synapse-specific and size-dependent mechanisms of spine structural plasticity accompanying synaptic weakening. Proc. Natl. Acad. Sci. USA 2013, 110, E305–E312. [Google Scholar] [CrossRef] [Green Version]
- Wiegert, J.S.; Oertner, T.G. Long-Term Depression triggers the selective elimination of weakly integrated synapses. Proc. Natl. Acad. Sci. USA 2013, 110, E4510–E4519. [Google Scholar] [CrossRef] [Green Version]
- Pchitskaya, E.; Bezprozvanny, I. Dendritic spines shape analysis—Classification or clusterization? Perspective. Front. Synaptic Neurosci. 2020, 12, 31. [Google Scholar] [CrossRef]
- Woolfrey, K.M.; Srivastava, D.P. Control of dendritic spine morphological and functional plasticity by small GTPases. Neural Plast. 2016, 2016, 3025948. [Google Scholar] [CrossRef] [Green Version]
- Emes, R.D.; Pocklington, A.J.; Anderson, C.N.G.; Bayes, A.; Collins, M.O.; Vickers, C.A.; Croning, M.D.R.; Malik, B.R.; Choudhary, J.S.; Armstrong, J.D.; et al. Evolutionary expansion and anatomical specialization of synapse proteome complexity. Nat. Neurosci. 2008, 11, 799–806. [Google Scholar] [CrossRef] [Green Version]
- De Roo, M.; Klauser, P.; Mendez, P.; Poglia, L.; Muller, D. Activity-dependent PSD formation and stabilization of newly formed spines in hippocampal slice cultures. Cereb. Cortex 2008, 18, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Kayser, M.S.; Nolt, M.J.; Dalva, M.B. EphB receptors couple dendritic filopodia motility to synapse formation. Neuron 2008, 59, 56–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.-L.; Lei, Y.-T.; Hong, C.-J.; Hsueh, Y.-P. Syndecan-2 induces filopodia and dendritic spine formation via the Neurofibromin-PKA-Ena/VASP pathway. J. Cell Biol. 2007, 177, 829–841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arstikaitis, P.; Gauthier-Campbell, C.; Carolina Gutierrez Herrera, R.; Huang, K.; Levinson, J.N.; Murphy, T.H.; Kilimann, M.W.; Sala, C.; Colicos, M.A.; El-Husseini, A. Paralemmin-1, a Modulator of filopodia induction is required for spine maturation. Mol. Biol. Cell 2008, 19, 2026–2038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuno, H.; Okabe, S.; Mishina, M.; Yanagida, T.; Mori, K.; Yoshihara, Y. Telencephalin slows spine maturation. J. Neurosci. Off. J. Soc. Neurosci. 2006, 26, 1776–1786. [Google Scholar] [CrossRef] [Green Version]
- Honkura, N.; Matsuzaki, M.; Noguchi, J.; Ellis-Davies, G.C.R.; Kasai, H. The subspine organization of actin fibers regulates the structure and plasticity of dendritic spines. Neuron 2008, 57, 719–729. [Google Scholar] [CrossRef] [Green Version]
- Basu, S.; Lamprecht, R. The role of actin cytoskeleton in dendritic spines in the maintenance of long-term memory. Front. Mol. Neurosci. 2018, 11, 143. [Google Scholar] [CrossRef] [Green Version]
- Borovac, J.; Bosch, M.; Okamoto, K. Regulation of actin dynamics during structural plasticity of dendritic spines: Signaling messengers and actin-binding proteins. Mol. Cell. Neurosci. 2018, 91, 122–130. [Google Scholar] [CrossRef]
- Koskinen, M.; Hotulainen, P. Measuring F-actin properties in dendritic spines. Front. Neuroanat. 2014, 8, 74. [Google Scholar] [CrossRef] [Green Version]
- Kilinc, D. The emerging role of mechanics in synapse formation and plasticity. Front. Cell. Neurosci. 2018, 12, 483. [Google Scholar] [CrossRef]
- Runge, K.; Cardoso, C.; de Chevigny, A. Dendritic Spine Plasticity: Function and Mechanisms. Front. Synaptic Neurosci. 2020, 12, 36. [Google Scholar] [CrossRef]
- Berry, K.P.; Nedivi, E. Spine Dynamics: Are they all the same? Neuron 2017, 96, 43–55. [Google Scholar] [CrossRef] [Green Version]
- Duncan, B.W.; Mohan, V.; Wade, S.D.; Truong, Y.; Kampov-Polevoi, A.; Temple, B.R.; Maness, P.F. Semaphorin3F drives dendritic spine pruning through Rho-GTPase signaling. bioRxiv 2021, 1–18. [Google Scholar] [CrossRef]
- Fiala, J.C.; Spacek, J.; Harris, K.M. Dendritic spine pathology: Cause or consequence of neurological disorders? Brain Res. Rev. 2002, 39, 29–54. [Google Scholar] [CrossRef]
- Norrholm, S.D.; Ouimet, C.C. Altered dendritic spine density in animal models of depression and in response to antidepressant treatment. Synapse 2001, 42, 151–163. [Google Scholar] [CrossRef]
- Hayashi, K.; Shirao, T. Change in the Shape of dendritic spines caused by overexpression of drebrin in cultured cortical neurons. J. Neurosci. 1999, 19, 3918–3925. [Google Scholar] [CrossRef]
- Greenwood, S.M.; Mizielinska, S.M.; Frenguelli, B.G.; Harvey, J.; Connolly, C.N. Mitochondrial dysfunction and dendritic beading during neuronal toxicity. J. Biol. Chem. 2007, 282, 26235–26244. [Google Scholar] [CrossRef] [Green Version]
- Scheibel, A.B.; Tomiyasu, U. Dendritic sprouting in Alzheimer’s presenile dementia. Exp. Neurol. 1978, 60, 1–8. [Google Scholar] [CrossRef]
- Bloss, E.B.; Janssen, W.G.; Ohm, D.T.; Yuk, F.J.; Wadsworth, S.; Saardi, K.M.; McEwen, B.S.; Morrison, J.H. Evidence for reduced experience-dependent dendritic spine plasticity in the aging prefrontal cortex. J. Neurosci. 2011, 31, 7831–7839. [Google Scholar] [CrossRef]
- Maiti, P.; Manna, J.; Ilavazhagan, G.; Rossignol, J.; Dunbar, G.L. Molecular Regulation of dendritic spine dynamics and their potential impact on synaptic plasticity and neurological diseases. Neurosci. Biobehav. Rev. 2015, 59, 208–237. [Google Scholar] [CrossRef]
- Danglot, L.; Freret, T.; Roux, N.L.; Nême, N.N.; Burgo, A.; Hyenne, V.; Roumier, A.; Contremoulins, V.; Dauphin, F.; Bizot, J.-C.; et al. Vezatin is essential for dendritic spine morphogenesis and functional synaptic maturation. J. Neurosci. 2012, 32, 9007–9022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishii, K.; Nagaoka, A.; Kishida, Y.; Okazaki, H.; Yagishita, S.; Ucar, H.; Takahashi, N.; Saito, N.; Kasai, H. In vivo volume dynamics of dendritic spines in the neocortex of wild-type and Fmr1 KO Mice. eNeuro 2018, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borczyk, M.; Śliwińska, M.A.; Caly, A.; Bernas, T.; Radwanska, K. Neuronal plasticity affects correlation between the size of dendritic spine and its postsynaptic density. Sci. Rep. 2019, 9, 1693. [Google Scholar] [CrossRef] [PubMed]
- Sala, C.; Segal, M. Dendritic spines: The locus of structural and functional plasticity. Physiol. Rev. 2014, 94, 141–188. [Google Scholar] [CrossRef]
- Südhof, T.C. Neuroligins and neurexins link synaptic function to cognitive disease. Nature 2008, 455, 903–911. [Google Scholar] [CrossRef] [Green Version]
- Bhatt, P.C.; Pathak, S.; Kumar, V.; Panda, B.P. Attenuation of Neurobehavioral and neurochemical abnormalities in animal model of cognitive deficits of Alzheimer’s disease by fermented soybean nanonutraceutical. Inflammopharmacology 2018, 26, 105–118. [Google Scholar] [CrossRef]
- Dorostkar, M.M.; Zou, C.; Blazquez-Llorca, L.; Herms, J. analyzing dendritic spine pathology in Alzheimer’s disease: Problems and opportunities. Acta Neuropathol. 2015, 130, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Toledano, A.; Alvarez, M.I. Lesions and dysfunctions of the nucleus basalis as Alzheimer’s disease models: General and critical overview and analysis of the long-term changes in several excitotoxic models. Curr. Alzheimer Res. 2004, 1, 189–214. [Google Scholar] [CrossRef]
- Champtiaux, N.; Changeux, J.-P. Knockout and knockin mice to investigate the role of nicotinic receptors in the central nervous system. Prog. Brain Res. 2004, 145, 235–251. [Google Scholar] [CrossRef]
- Pugh, P.C.; Berg, D.K. Neuronal Acetylcholine receptors that bind alpha-bungarotoxin mediate neurite retraction in a calcium-dependent manner. J. Neurosci. Off. J. Soc. Neurosci. 1994, 14, 889–896. [Google Scholar] [CrossRef]
- Chattipakorn, S.C.; Leech, T.; Apaijai, N.; Higgins, L.; Jinawong, K.; Palee, S.; Maneechote, C.; Jaiwongkam, T.; Chattipakorn, N. Pretreatment with metformin reduced dendritic spine loss following cardiac ischaemia/reperfusion injury by preventing amyloid beta aggregation, brain inflammation and mitochondrial dysfunction. Eur. Heart J. 2020, 41, ehaa946.3142. [Google Scholar] [CrossRef]
- Ongnok, B.; Khuanjing, T.; Chunchai, T.; Kerdphoo, S.; Jaiwongkam, T.; Chattipakorn, N.; Chattipakorn, S.C. Donepezil provides neuroprotective effects against brain injury and Alzheimer’s pathology under conditions of cardiac ischemia/reperfusion injury. Biochim. Biophys. Acta Mol. Basis Dis. 2021, 1867, 165975. [Google Scholar] [CrossRef]
- Alcantara-Gonzalez, F.; Juarez, I.; Solis, O.; Martinez-Tellez, I.; Camacho-Abrego, I.; Masliah, E.; Mena, R.; Flores, G. Enhanced dendritic spine number of neurons of the prefrontal cortex, hippocampus and nucleus accumbens in old rats after chronic donepezil administration. Synapse 2010, 64, 786–793. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.; Tran, K.C.; Zeng, A.Y.; Massa, S.M.; Longo, F.M. Small Molecule Modulation of the P75 Neurotrophin Receptor Inhibits Multiple Amyloid Beta-Induced Tau Pathologies. Sci. Rep. 2020, 10, 20322. [Google Scholar] [CrossRef]
- Rajendran, K.; Chellappan, D.R.; Sanaranarayanan, S.; Ramakrishnan, V.; Krishnan, U.M. Investigations on a Polyherbal Formulation for Treatment of Cognitive Impairment in a Cholinergic Dysfunctional Rodent Model. Neurochem. Int. 2020, 141, 104890. [Google Scholar] [CrossRef]
- Lecca, D.; Bader, M.; Tweedie, D.; Hoffman, A.F.; Jung, Y.J.; Hsueh, S.-C.; Hoffer, B.J.; Becker, R.E.; Pick, C.G.; Lupica, C.R.; et al. (-)-Phenserine and the Prevention of pre-programmed cell death and neuroinflammation in mild traumatic brain injury and Alzheimer’s disease challenged mice. Neurobiol. Dis. 2019, 130, 104528. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, X.C.; Zhang, H.Y. Huperzine a Alleviates synaptic deficits and modulates amyloidogenic and nonamyloidogenic pathways in APPswe/PS1dE9 transgenic mice. J. Neurosci. Res. 2012, 90, 508–517. [Google Scholar] [CrossRef]
- Kang, L.; Tian, M.K.; Bailey, C.D.C.; Lambe, E.K. Dendritic Spine density of prefrontal layer 6 pyramidal neurons in relation to apical dendrite sculpting by nicotinic acetylcholine receptors. Front. Cell. Neurosci. 2015, 9, 398. [Google Scholar] [CrossRef] [Green Version]
- Oda, A.; Yamagata, K.; Nakagomi, S.; Uejima, H.; Wiriyasermkul, P.; Ohgaki, R.; Nagamori, S.; Kanai, Y.; Tanaka, H. Nicotine induces dendritic spine remodeling in cultured hippocampal neurons. J. Neurochem. 2014, 128, 246–255. [Google Scholar] [CrossRef] [Green Version]
- Lozada, A.F.; Wang, X.; Gounko, N.V.; Massey, K.A.; Duan, J.; Liu, Z.; Berg, D.K. Induction of Dendritic spines by Β2-containing nicotinic receptors. J. Neurosci. 2012, 32, 8391–8400. [Google Scholar] [CrossRef]
- Schätzle, P.; Ster, J.; Verbich, D.; McKinney, R.A.; Gerber, U.; Sonderegger, P.; María Mateos, J. Rapid and reversible formation of spine head filopodia in response to muscarinic receptor activation in CA1 pyramidal cells. J. Physiol. 2011, 589, 4353–4364. [Google Scholar] [CrossRef] [Green Version]
- Korkotian, E.; Segal, M. Bidirectional regulation of dendritic spine dimensions by glutamate receptors. Neuroreport 1999, 10, 2875–2877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korkotian, E.; Segal, M. Release of Calcium from stores alters the morphology of dendritic spines in cultured hippocampal neurons. Proc. Natl. Acad. Sci. USA 1999, 96, 12068–12072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanderklish, P.W.; Edelman, G.M. Dendritic Spines elongate after stimulation of group 1 metabotropic glutamate receptors in cultured hippocampal neurons. Proc. Natl. Acad. Sci. USA 2002, 99, 1639–1644. [Google Scholar] [CrossRef] [Green Version]
- Maletic-Savatic, M.; Malinow, R.; Svoboda, K. Rapid dendritic morphogenesis in CA1 hippocampal dendrites induced by synaptic activity. Science 1999, 283, 1923–1927. [Google Scholar] [CrossRef] [Green Version]
- Hasbani, M.J.; Schlief, M.L.; Fisher, D.A.; Goldberg, M.P. Dendritic spines lost during glutamate receptor activation reemerge at original sites of synaptic contact. J. Neurosci. Off. J. Soc. Neurosci. 2001, 21, 2393–2403. [Google Scholar] [CrossRef] [Green Version]
- Fischer, M.; Kaech, S.; Wagner, U.; Brinkhaus, H.; Matus, A. Glutamate receptors regulate actin-based plasticity in dendritic spines. Nat. Neurosci. 2000, 3, 887–894. [Google Scholar] [CrossRef]
- Pereira, A.C.; Lambert, H.K.; Grossman, Y.S.; Dumitriu, D.; Waldman, R.; Jannetty, S.K.; Calakos, K.; Janssen, W.G.; McEwen, B.S.; Morrison, J.H. Glutamatergic Regulation prevents hippocampal-dependent age-related cognitive decline through dendritic spine clustering. Proc. Natl. Acad. Sci. USA 2014, 111, 18733–18738. [Google Scholar] [CrossRef] [Green Version]
- LaCrosse, A.L.; Taylor, S.B.; Nemirovsky, N.E.; Gass, J.T.; Olive, M.F. MGluR5 Positive and negative allosteric modulators differentially affect dendritic spine density and morphology in the prefrontal cortex. CNS Neurol. Disord. Drug Targets 2015, 14, 476–485. [Google Scholar] [CrossRef] [Green Version]
- Kuhlmann, N.; Wagner Valladolid, M.; Quesada-Ramírez, L.; Farrer, M.J.; Milnerwood, A.J. Chronic and acute manipulation of cortical glutamate transmission induces structural and synaptic changes in co-cultured striatal neurons. Front. Cell. Neurosci. 2021, 15, 23. [Google Scholar] [CrossRef]
- Ali, F.; Gerhard, D.M.; Sweasy, K.; Pothula, S.; Pittenger, C.; Duman, R.S.; Kwan, A.C. Ketamine disinhibits dendrites and enhances calcium signals in prefrontal dendritic spines. Nat. Commun. 2020, 11, 72. [Google Scholar] [CrossRef] [Green Version]
- Seo, M.; Hien, L.T.; Park, M.K.; Choi, A.J.; Seog, D.-H.; Kim, S.-H.; Park, S.W.; Lee, J.G. AMPA Receptor-MTORC1 signaling activation is required for neuroplastic effects of LY341495 in rat hippocampal neurons. Sci. Rep. 2020, 10, 993. [Google Scholar] [CrossRef]
- Yoon, Y.J.; Gokin, A.P.; Martin-Caraballo, M. Pharmacological manipulation of GABA-driven activity in ovo disrupts the development of dendritic morphology but not the maturation of spinal cord network activity. Neural Dev. 2010, 5, 11. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y. GABA-A receptor-dependent mechanisms prevent excessive spine elimination during postnatal maturation of the mouse cortex in vivo. FEBS Lett. 2014, 588, 4551–4560. [Google Scholar] [CrossRef] [Green Version]
- Salmon, C.K.; Pribiag, H.; Gizowski, C.; Farmer, W.T.; Cameron, S.; Jones, E.V.; Mahadevan, V.; Bourque, C.W.; Stellwagen, D.; Woodin, M.A.; et al. Depolarizing GABA transmission restrains activity-dependent glutamatergic synapse formation in the developing hippocampal circuit. Front. Cell. Neurosci. 2020, 14, 36. [Google Scholar] [CrossRef] [Green Version]
- Lachance-Touchette, P.; Choudhury, M.; Stoica, A.; Di Cristo, G.; Cossette, P. Single-cell genetic expression of mutant GABAA receptors causing human genetic epilepsy alters dendritic spine and GABAergic bouton formation in a mutation-specific manner. Front. Cell. Neurosci. 2014, 8, 317. [Google Scholar] [CrossRef] [Green Version]
- Lur, G.; Higley, M.J. Glutamate receptor modulation is restricted to synaptic microdomains. Cell Rep. 2015, 12, 326–334. [Google Scholar] [CrossRef] [Green Version]
- Giachero, M.; Calfa, G.D.; Molina, V.A. Hippocampal Dendritic spines remodeling and fear memory are modulated by GABAergic signaling within the basolateral amygdala complex. Hippocampus 2015, 25, 545–555. [Google Scholar] [CrossRef]
- Vitalis, T.; Ansorge, M.S.; Dayer, A.G. Serotonin Homeostasis and serotonin receptors as actors of cortical construction: Special attention to the 5-HT3A and 5-HT6 receptor subtypes. Front. Cell. Neurosci. 2013, 7, 93. [Google Scholar] [CrossRef] [Green Version]
- Bonnin, A.; Goeden, N.; Chen, K.; Wilson, M.L.; King, J.; Shih, J.C.; Blakely, R.D.; Deneris, E.S.; Levitt, P. A Transient placental source of serotonin for the fetal forebrain. Nature 2011, 472, 347–350. [Google Scholar] [CrossRef] [Green Version]
- Dayer, A. Serotonin-related pathways and developmental plasticity: Relevance for psychiatric disorders. Dialogues Clin. Neurosci. 2014, 16, 29–41. [Google Scholar] [PubMed]
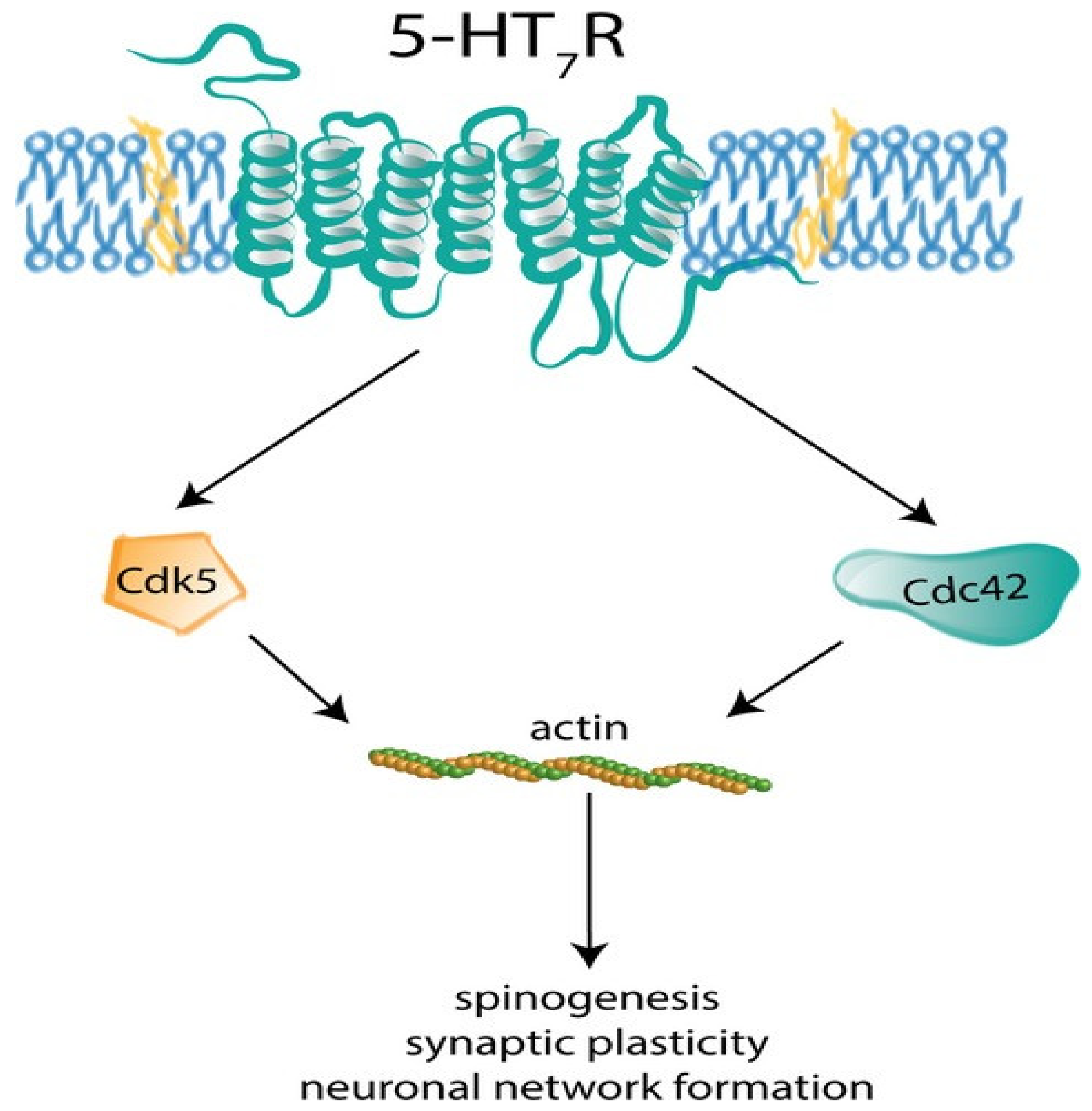
- Kobe, F.; Guseva, D.; Jensen, T.P.; Wirth, A.; Renner, U.; Hess, D.; Müller, M.; Medrihan, L.; Zhang, W.; Zhang, M.; et al. 5-HT7R/G12 signaling regulates neuronal morphology and function in an age-dependent manner. J. Neurosci. Off. J. Soc. Neurosci. 2012, 32, 2915–2930. [Google Scholar] [CrossRef] [PubMed]
- Hedlund, P.B.; Leopoldo, M.; Caccia, S.; Sarkisyan, G.; Fracasso, C.; Martelli, G.; Lacivita, E.; Berardi, F.; Perrone, R. LP-211 Is a brain penetrant selective agonist for the serotonin 5-HT(7) receptor. Neurosci. Lett. 2010, 481, 12–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bijata, M.; Labus, J.; Guseva, D.; Stawarski, M.; Butzlaff, M.; Dzwonek, J.; Schneeberg, J.; Böhm, K.; Michaluk, P.; Rusakov, D.A.; et al. Synaptic remodeling depends on signaling between serotonin receptors and the extracellular matrix. Cell Rep. 2017, 19, 1767–1782. [Google Scholar] [CrossRef] [Green Version]
- Jones, K.A.; Srivastava, D.P.; Allen, J.A.; Strachan, R.T.; Roth, B.L.; Penzes, P. Rapid Modulation of spine morphology by the 5-HT2A serotonin receptor through Kalirin-7 signaling. Proc. Natl. Acad. Sci. USA 2009, 106, 19575–19580. [Google Scholar] [CrossRef] [Green Version]
- Mi, Z.; Si, T.; Kapadia, K.; Li, Q.; Muma, N.A. Receptor-stimulated transamidation induces activation of Rac1 and Cdc42 and the regulation of dendritic spines. Neuropharmacology 2017, 117, 93–105. [Google Scholar] [CrossRef] [Green Version]
- Waller, J.A.; Chen, F.; Sánchez, C. Vortioxetine promotes maturation of dendritic spines in vitro: A comparative study in hippocampal cultures. Neuropharmacology 2016, 103, 143–154. [Google Scholar] [CrossRef]
- Speranza, L.; Labus, J.; Volpicelli, F.; Guseva, D.; Lacivita, E.; Leopoldo, M.; Bellenchi, G.C.; di Porzio, U.; Bijata, M.; Perrone-Capano, C.; et al. Serotonin 5-HT7 Receptor increases the density of dendritic spines and facilitates synaptogenesis in forebrain neurons. J. Neurochem. 2017, 141, 647–661. [Google Scholar] [CrossRef] [Green Version]
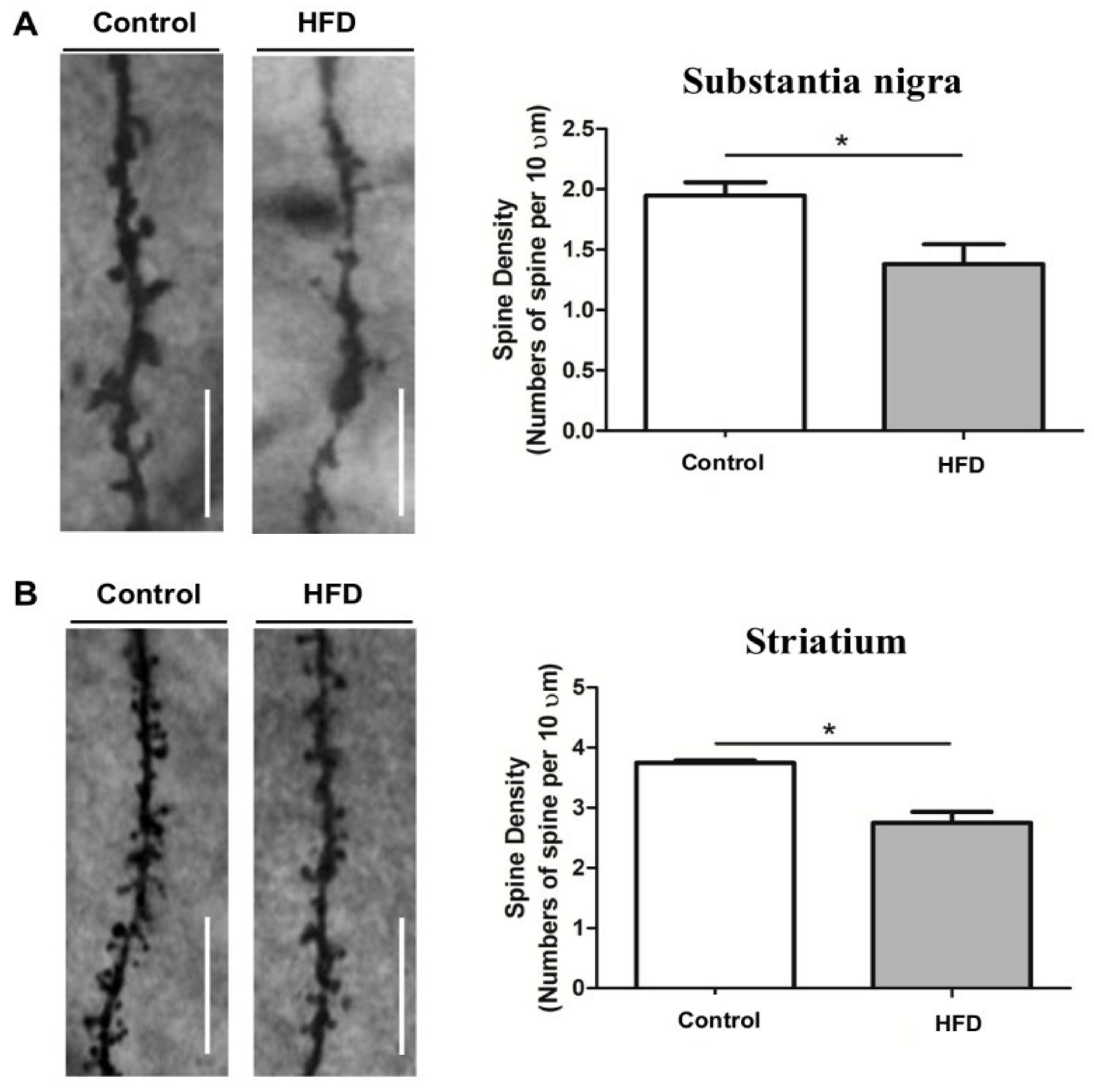
- Kao, Y.-C.; Wei, W.-Y.; Tsai, K.; Wang, L.-C. High fat diet suppresses peroxisome proliferator-activated receptors and reduces dopaminergic neurons in the substantia nigra. Int. J. Mol. Sci. 2019, 21, 207. [Google Scholar] [CrossRef] [Green Version]
- O’Dell, T.J.; Connor, S.A.; Guglietta, R.; Nguyen, P.V. β-adrenergic receptor signaling and modulation of long-term potentiation in the mammalian hippocampus. Learn. Mem. 2015, 22, 461–471. [Google Scholar] [CrossRef] [Green Version]
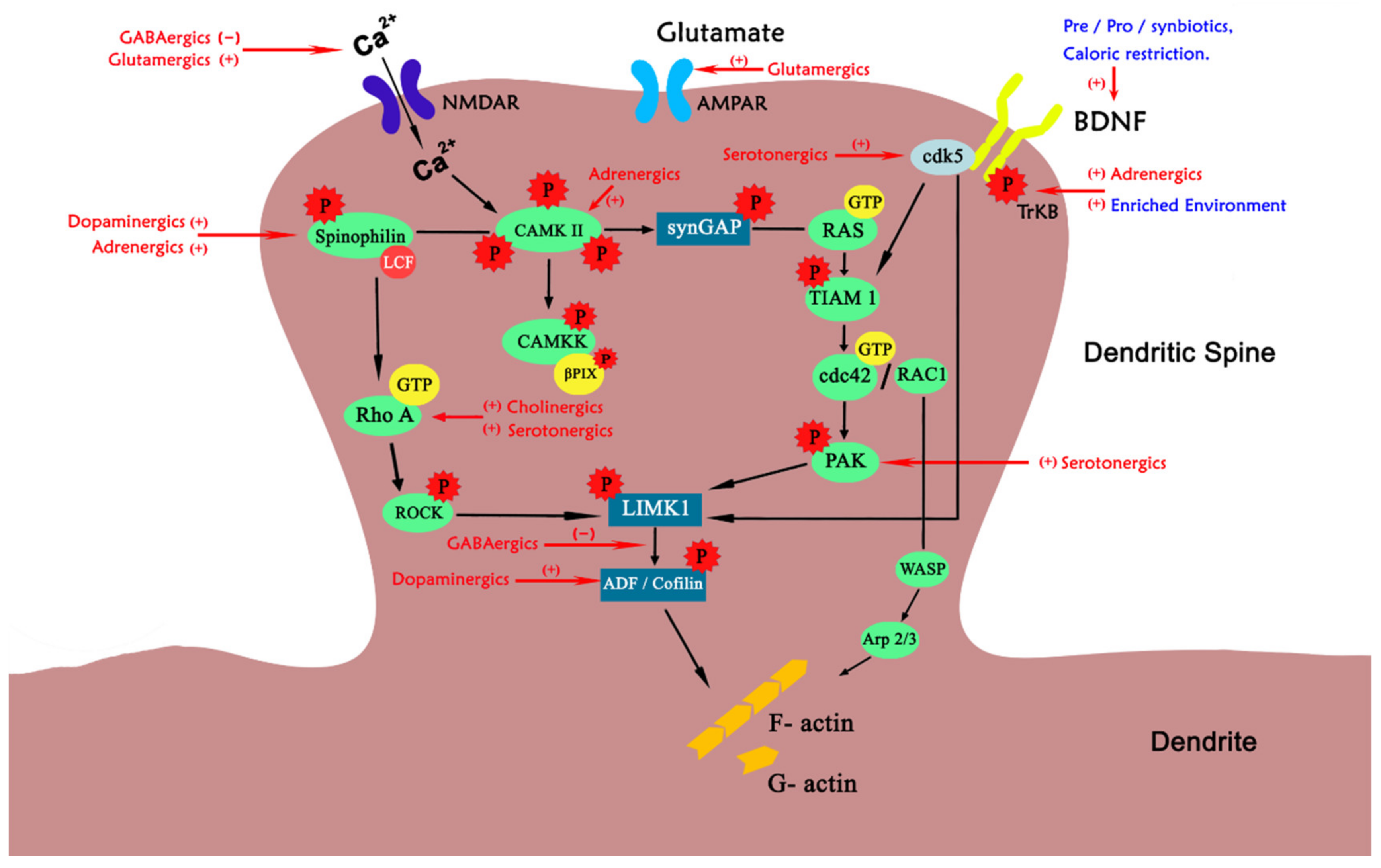
- Ji, Y.; Pang, P.T.; Feng, L.; Lu, B. Cyclic AMP controls BDNF-induced TrkB Phosphorylation and dendritic spine formation in mature hippocampal neurons. Nat. Neurosci. 2005, 8, 164–172. [Google Scholar] [CrossRef]
- Jȩdrzejewska-Szmek, J.; Luczak, V.; Abel, T.; Blackwell, K.T. β-adrenergic signaling broadly contributes to LTP induction. PLoS Comput. Biol. 2017, 13, e1005657. [Google Scholar] [CrossRef] [Green Version]
- Ren, W.-W.; Liu, Y.; Li, B.-M. Stimulation of α(2A)-adrenoceptors promotes the maturation of dendritic spines in cultured neurons of the medial prefrontal cortex. Mol. Cell. Neurosci. 2012, 49, 205–216. [Google Scholar] [CrossRef]
- Hains, A.B.; Yabe, Y.; Arnsten, A.F.T. Chronic stimulation of Alpha-2A-adrenoceptors with guanfacine protects rodent prefrontal cortex dendritic spines and cognition from the effects of chronic stress. Neurobiol. Stress 2015, 2, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Vidovic, M.; Chen, M.-M.; Lu, Q.-Y.; Song, Z.-M. Activation of A2A Adrenoceptors Alters Dendritic Spine Development and the Expression of Spinophilin in Cultured Cortical Neurones. Brain Res. 2008, 1199, 37–45. [Google Scholar] [CrossRef]
- Avila-Costa, M.R.; Colín-Barenque, L.; Montiel-Flores, E.; Aley-Medina, P.; Valdez, A.L.G.; Librado, J.L.O.; Martínez, E.F.; Martínez, V.A.; Mussali-Galante, P.; Fortoul, T.I. Bromocriptine treatment in a murine Parkinson’s model: Ultrastructural evaluation after dopaminergic deafferentation. Int. J. Neurosci. 2005, 115, 851–859. [Google Scholar] [CrossRef]
- Yao, W.-D.; Spealman, R.D.; Zhang, J. Dopaminergic signaling in dendritic spines. Biochem. Pharmacol. 2008, 75, 2055–2069. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Gu, J.; Wang, B.; Xie, M.; Huang, L.; Liu, Y.; Zhang, L.; Xue, J.; Guo, F.; Zhang, L.; et al. Activation of dopamine D1 receptors regulates dendritic morphogenesis through Rac1 and RhoA in prefrontal cortex neurons. Mol. Neurobiol. 2015, 51, 1024–1037. [Google Scholar] [CrossRef] [Green Version]
- Zagrebelsky, M.; Tacke, C.; Korte, M. BDNF signaling during the lifetime of dendritic spines. Cell Tissue Res. 2020, 382, 185–199. [Google Scholar] [CrossRef]
- Collo, G.; Cavalleri, L.; Bono, F.; Mora, C.; Fedele, S.; Invernizzi, R.W.; Gennarelli, M.; Piovani, G.; Kunath, T.; Millan, M.J.; et al. Ropinirole and pramipexole promote structural plasticity in human IPSC-derived dopaminergic neurons via BDNF and MTOR signaling. Neural Plast. 2018, 2018, 4196961. [Google Scholar] [CrossRef]
- Parajuli, L.K.; Wako, K.; Maruo, S.; Kakuta, S.; Taguchi, T.; Ikuno, M.; Yamakado, H.; Takahashi, R.; Koike, M. Developmental changes in dendritic spine morphology in the striatum and their alteration in an A53T α-Synuclein transgenic mouse model of Parkinson’s disease. eNeuro 2020, 7. [Google Scholar] [CrossRef] [PubMed]
- Ueno, T.; Nishijima, H.; Ueno, S.; Tomiyama, M. Spine Enlargement of pyramidal tract-type neurons in the motor cortex of a rat model of Levodopa-induced dyskinesia. Front. Neurosci. 2017, 11, 206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Funamizu, Y.; Nishijima, H.; Ueno, T.; Ueno, S.; Mizukami, H.; Yagihashi, S.; Tomiyama, M. Morphological dendritic spine changes of medium spiny neurons in the nucleus accumbens in 6-Hydroxydopamine-Lesioned Rats treated with Levodopa. Neurosci. Res. 2017, 121, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Gagnon, D.; Petryszyn, S.; Sanchez, M.G.; Bories, C.; Beaulieu, J.M.; De Koninck, Y.; Parent, A.; Parent, M. Striatal neurons expressing D1 and D2 receptors are morphologically distinct and differently affected by dopamine denervation in mice. Sci. Rep. 2017, 7, 41432. [Google Scholar] [CrossRef] [Green Version]
- Arroyo-García, L.E.; Vázquez-Roque, R.A.; Díaz, A.; Treviño, S.; De La Cruz, F.; Flores, G.; Rodríguez-Moreno, A. The effects of non-selective dopamine receptor activation by apomorphine in the mouse hippocampus. Mol. Neurobiol. 2018, 55, 8625–8636. [Google Scholar] [CrossRef]
- Russo, E.; De Fazio, S.; De Fazio, P.; Amorosi, A.; Perrotta, I.; De Sarro, G.; Donato, G. Apomorphine-Induced neurodegeneration in mongolian gerbil hippocampus. Schizophr. Res. 2007, 95, 223–227. [Google Scholar] [CrossRef]
- Razgado-Hernandez, L.F.; Espadas-Alvarez, A.J.; Reyna-Velazquez, P.; Sierra-Sanchez, A.; Anaya-Martinez, V.; Jimenez-Estrada, I.; Bannon, M.J.; Martinez-Fong, D.; Aceves-Ruiz, J. The transfection of BDNF to dopamine neurons potentiates the effect of dopamine D3 Receptor agonist recovering the striatal innervation, dendritic spines and motor behavior in an aged rat model of Parkinson’s disease. PLoS ONE 2015, 10, e0117391. [Google Scholar] [CrossRef]
- Graziane, N.M.; Yuen, E.Y.; Yan, Z. Dopamine D4 receptors regulate GABAA receptor trafficking via an actin/cofilin/myosin-dependent mechanism. J. Biol. Chem. 2009, 284, 8329–8336. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.-D.; Deutch, A.Y. Dopamine depletion of the prefrontal cortex induces dendritic spine loss: Reversal by atypical antipsychotic drug treatment. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2008, 33, 1276–1286. [Google Scholar] [CrossRef] [Green Version]
- Garcia, B.G.; Neely, M.D.; Deutch, A.Y. Cortical regulation of striatal medium spiny neuron dendritic remodeling in Parkinsonism: Modulation of glutamate release reverses dopamine depletion–induced dendritic spine loss. Cereb. Cortex 2010, 20, 2423–2432. [Google Scholar] [CrossRef] [Green Version]
- Fasano, C.; Bourque, M.-J.; Lapointe, G.; Leo, D.; Thibault, D.; Haber, M.; Kortleven, C.; Desgroseillers, L.; Murai, K.K.; Trudeau, L.-É. Dopamine facilitates dendritic spine formation by cultured striatal medium spiny neurons through Both D1 and D2 dopamine receptors. Neuropharmacology 2013, 67, 432–443. [Google Scholar] [CrossRef]
- Hiday, A.C.; Edler, M.C.; Salek, A.B.; Morris, C.W.; Thang, M.; Rentz, T.J.; Rose, K.L.; Jones, L.M.; Baucum, A.J. Mechanisms and consequences of dopamine depletion-induced attenuation of the spinophilin/neurofilament medium interaction. PMC 2017, 2017, 4153076. [Google Scholar] [CrossRef] [Green Version]
- Areal, L.B.; Hamilton, A.; Martins-Silva, C.; Pires, R.G.W.; Ferguson, S.S.G. Neuronal scaffolding protein spinophilin is integral for cocaine-induced behavioral sensitization and ERK1/2 activation. Mol. Brain 2019, 12, 15. [Google Scholar] [CrossRef] [Green Version]
- Jung, G.; Kim, E.-J.; Cicvaric, A.; Sase, S.; Gröger, M.; Höger, H.; Sialana, F.J.; Berger, J.; Monje, F.J.; Lubec, G. Drebrin depletion alters neurotransmitter receptor levels in protein complexes, dendritic spine morphogenesis and memory-related synaptic plasticity in the mouse hippocampus. J. Neurochem. 2015, 134, 327–339. [Google Scholar] [CrossRef] [Green Version]
- Cannizzaro, C.; Talani, G.; Brancato, A.; Mulas, G.; Spiga, S.; Luca, M.A.D.; Sanna, A.; Marino, R.A.M.; Biggio, G.; Sanna, E.; et al. Dopamine restores limbic memory loss, dendritic spine structure, and NMDAR-dependent LTD in the Nucleus accumbens of alcohol-withdrawn rats. J. Neurosci. 2019, 39, 929–943. [Google Scholar] [CrossRef] [Green Version]
- Wahl, D.; Solon-Biet, S.M.; Wang, Q.-P.; Wali, J.A.; Pulpitel, T.; Clark, X.; Raubenheimer, D.; Senior, A.M.; Sinclair, D.A.; Cooney, G.J.; et al. Comparing the effects of low-protein and high-carbohydrate diets and caloric restriction on brain aging in mice. Cell Rep. 2018, 25, 2234–2243.e6. [Google Scholar] [CrossRef] [Green Version]
- Pani, G. Neuroprotective effects of dietary restriction: Evidence and mechanisms. Semin. Cell Dev. Biol. 2015, 40, 106–114. [Google Scholar] [CrossRef]
- Brandhorst, S.; Choi, I.Y.; Wei, M.; Cheng, C.W.; Sedrakyan, S.; Navarrete, G.; Dubeau, L.; Yap, L.P.; Park, R.; Vinciguerra, M.; et al. A periodic diet that mimics fasting promotes multi-system regeneration, enhanced cognitive performance, and healthspan. Cell Metab. 2015, 22, 86–99. [Google Scholar] [CrossRef] [Green Version]
- Stranahan, A.M.; Lee, K.; Martin, B.; Maudsley, S.; Golden, E.; Cutler, R.G.; Mattson, M.P. Voluntary exercise and caloric restriction enhance hippocampal dendritic spine density and BDNF levels in diabetic mice. Hippocampus 2009, 19, 951–961. [Google Scholar] [CrossRef]
- Moroi-Fetters, S.E.; Mervis, R.F.; London, E.D.; Ingram, D.K. Dietary Restriction suppresses age-related changes in dendritic spines. Neurobiol. Aging 1989, 10, 317–322. [Google Scholar] [CrossRef]
- Sripetchwandee, J.; Pipatpiboon, N.; Pratchayasakul, W.; Chattipakorn, N.; Chattipakorn, S.C. DPP-4 inhibitor and PPARγ agonist restore the loss of CA1 dendritic spines in obese insulin-resistant rats. Arch. Med. Res. 2014, 45, 547–552. [Google Scholar] [CrossRef] [PubMed]
- Torabi, T.; Azizzadeh Delshad, A.; Roghani, M. Dietary Restriction prevents dendritic changes of pyramidal neurons in hippocampal and prefrontal cortex in diabetic rat. J. Basic Clin. Pathophysiol. 2019, 7, 28–32. [Google Scholar] [CrossRef]
- Andrade, J.P.; Lukoyanov, N.V.; Paula-Barbosa, M.M. Chronic Food restriction is associated with subtle dendritic alterations in granule cells of the rat hippocampal formation. Hippocampus 2002, 12, 149–164. [Google Scholar] [CrossRef] [PubMed]
- Salas, M. Effects of early undernutrition on dendritic spines of cortical pyramidal cells in the rat. Dev. Neurosci. 1980, 3, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Benítez-Bribiesca, L.; De la Rosa-Alvarez, I.; Mansilla-Olivares, A. Dendritic spine pathology in infants with severe protein-calorie malnutrition. Pediatrics 1999, 104, e21. [Google Scholar] [CrossRef] [Green Version]
- Braniste, V.; Al-Asmakh, M.; Kowal, C.; Anuar, F.; Abbaspour, A.; Tóth, M.; Korecka, A.; Bakocevic, N.; Ng, L.G.; Guan, N.L.; et al. The gut microbiota influences blood-brain barrier permeability in mice. Sci. Transl. Med. 2014, 6, 263ra158. [Google Scholar] [CrossRef] [Green Version]
- Möhle, L.; Mattei, D.; Heimesaat, M.M.; Bereswill, S.; Fischer, A.; Alutis, M.; French, T.; Hambardzumyan, D.; Matzinger, P.; Dunay, I.R.; et al. Ly6C(Hi) monocytes provide a link between antibiotic-induced changes in gut microbiota and adult hippocampal neurogenesis. Cell Rep. 2016, 15, 1945–1956. [Google Scholar] [CrossRef] [Green Version]
- Matcovitch-Natan, O.; Winter, D.R.; Giladi, A.; Vargas Aguilar, S.; Spinrad, A.; Sarrazin, S.; Ben-Yehuda, H.; David, E.; Zelada González, F.; Perrin, P.; et al. Microglia development follows a stepwise program to regulate brain homeostasis. Science 2016, 353, aad8670. [Google Scholar] [CrossRef]
- Hoban, A.E.; Stilling, R.M.; Ryan, F.J.; Shanahan, F.; Dinan, T.G.; Claesson, M.J.; Clarke, G.; Cryan, J.F. Regulation of prefrontal cortex myelination by the microbiota. Transl. Psychiatry 2016, 6, e774. [Google Scholar] [CrossRef] [Green Version]
- Desbonnet, L.; Clarke, G.; Traplin, A.; O’Sullivan, O.; Crispie, F.; Moloney, R.D.; Cotter, P.D.; Dinan, T.G.; Cryan, J.F. Gut microbiota depletion from early adolescence in mice: Implications for brain and behaviour. Brain Behav. Immun. 2015, 48, 165–173. [Google Scholar] [CrossRef]
- O’Mahony, S.M.; Clarke, G.; Borre, Y.E.; Dinan, T.G.; Cryan, J.F. Serotonin, tryptophan metabolism and the brain-gut-microbiome axis. Behav. Brain Res. 2015, 277, 32–48. [Google Scholar] [CrossRef]
- Dinan, T.G.; Stanton, C.; Cryan, J.F. Psychobiotics: A novel class of psychotropic. Biol. Psychiatry 2013, 74, 720–726. [Google Scholar] [CrossRef]
- Sarkar, A.; Lehto, S.M.; Harty, S.; Dinan, T.G.; Cryan, J.F.; Burnet, P.W.J. Psychobiotics and the Manipulation of bacteria-gut-brain signals. Trends Neurosci. 2016, 39, 763–781. [Google Scholar] [CrossRef] [Green Version]
- Arentsen, T.; Raith, H.; Qian, Y.; Forssberg, H.; Diaz Heijtz, R. Host microbiota modulates development of social preference in mice. Microb. Ecol. Health Dis. 2015, 26, 29719. [Google Scholar] [CrossRef]
- Luczynski, P.; Whelan, S.O.; O’Sullivan, C.; Clarke, G.; Shanahan, F.; Dinan, T.G.; Cryan, J.F. Adult Microbiota-deficient mice have distinct dendritic morphological changes: Differential effects in the amygdala and hippocampus. Eur. J. Neurosci. 2016, 44, 2654–2666. [Google Scholar] [CrossRef] [Green Version]
- De Vincenti, A.P.; Ríos, A.S.; Paratcha, G.; Ledda, F. Mechanisms that modulate and diversify BDNF functions: Implications for hippocampal synaptic plasticity. Front. Cell. Neurosci. 2019, 13, 135. [Google Scholar] [CrossRef] [Green Version]
- Kellner, Y.; Gödecke, N.; Dierkes, T.; Thieme, N.; Zagrebelsky, M.; Korte, M. The BDNF effects on dendritic spines of mature hippocampal neurons depend on neuronal activity. Front. Synaptic Neurosci. 2014, 6, 5. [Google Scholar] [CrossRef] [Green Version]
- O’Sullivan, E.; Barrett, E.; Grenham, S.; Fitzgerald, P.; Stanton, C.; Ross, R.P.; Quigley, E.M.M.; Cryan, J.F.; Dinan, T.G. BDNF expression in the hippocampus of maternally separated rats: Does bifidobacterium breve 6330 alter BDNF levels? Benef. Microbes 2011, 2, 199–207. [Google Scholar] [CrossRef]
- Ranuh, R.; Athiyyah, A.F.; Darma, A.; Risky, V.P.; Riawan, W.; Surono, I.S.; Sudarmo, S.M. Effect of the probiotic lactobacillus plantarum IS-10506 on BDNF and 5HT stimulation: Role of intestinal microbiota on the gut-brain axis. Iran. J. Microbiol. 2019, 11, 145–150. [Google Scholar] [CrossRef]
- Tochitani, S.; Ikeno, T.; Ito, T.; Sakurai, A.; Yamauchi, T.; Matsuzaki, H. Administration of non-absorbable antibiotics to pregnant mice to perturb the maternal gut microbiota is associated with alterations in offspring behavior. PLoS ONE 2016, 11, e0138293. [Google Scholar] [CrossRef]
- Xiao, J.; Wang, T.; Xu, Y.; Gu, X.; Li, D.; Niu, K.; Wang, T.; Zhao, J.; Zhou, R.; Wang, H.-L. Long-term probiotic intervention mitigates memory dysfunction through a novel H3K27me3-based mechanism in lead-exposed rats. Transl. Psychiatry 2020, 10, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chunchai, T.; Thunapong, W.; Yasom, S.; Wanchai, K.; Eaimworawuthikul, S.; Metzler, G.; Lungkaphin, A.; Pongchaidecha, A.; Sirilun, S.; Chaiyasut, C.; et al. Decreased microglial activation through gut-brain axis by prebiotics, probiotics, or synbiotics effectively restored cognitive function in obese-insulin resistant rats. J. Neuroinflamm. 2018, 15, 11. [Google Scholar] [CrossRef] [PubMed]
- Bocarsly, M.E.; Fasolino, M.; Kane, G.A.; LaMarca, E.A.; Kirschen, G.W.; Karatsoreos, I.N.; McEwen, B.S.; Gould, E. Obesity diminishes synaptic markers, alters microglial morphology, and impairs cognitive function. Proc. Natl. Acad. Sci. USA 2015, 112, 15731–15736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, S.; Dey, A.; Yu, X.; Stranahan, A.M. Dietary Obesity reversibly induces synaptic stripping by microglia and impairs hippocampal plasticity. Brain Behav. Immun. 2016, 51, 230–239. [Google Scholar] [CrossRef] [Green Version]
- Cani, P.D.; Amar, J.; Iglesias, M.A.; Poggi, M.; Knauf, C.; Bastelica, D.; Neyrinck, A.M.; Fava, F.; Tuohy, K.M.; Chabo, C.; et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes 2007, 56, 1761–1772. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.-A.; Gu, W.; Lee, I.-A.; Joh, E.-H.; Kim, D.-H. High fat diet-induced gut microbiota exacerbates inflammation and obesity in mice via the TLR4 signaling pathway. PLoS ONE 2012, 7, e47713. [Google Scholar] [CrossRef]
- Shirao, T.; Hanamura, K.; Koganezawa, N.; Ishizuka, Y.; Yamazaki, H.; Sekino, Y. The role of drebrin in neurons. J. Neurochem. 2017, 141, 819–834. [Google Scholar] [CrossRef] [Green Version]
- Koganezawa, N.; Hanamura, K.; Sekino, Y.; Shirao, T. The role of drebrin in dendritic spines. Mol. Cell. Neurosci. 2017, 84, 85–92. [Google Scholar] [CrossRef]
- Sekino, Y.; Koganezawa, N.; Mizui, T.; Shirao, T. Role of drebrin in synaptic plasticity. In Drebrin: From Structure and Function to Physiological and Pathological Roles; Advances in Experimental Medicine and Biology; Shirao, T., Sekino, Y., Eds.; Springer: Tokyo, Japan, 2017; pp. 183–201. ISBN 978-4-431-56550-5. [Google Scholar]
- Rudzki, L.; Szulc, A. “Immune gate” of psychopathology—The role of gut derived immune activation in major psychiatric disorders. Front. Psychiatry 2018, 9, 205. [Google Scholar] [CrossRef] [Green Version]
- Ishii, T.; Furuoka, H.; Kaya, M.; Kuhara, T. Oral administration of probiotic bifidobacterium breve improves facilitation of hippocampal memory extinction via restoration of aberrant higher induction of neuropsin in an MPTP-induced mouse model of Parkinson’s disease. Biomedicines 2021, 9, 167. [Google Scholar] [CrossRef]
- Talani, G.; Biggio, F.; Mostallino, M.C.; Locci, V.; Porcedda, C.; Boi, L.; Saolini, E.; Piras, R.; Sanna, E.; Biggio, G. Treatment with gut bifidobacteria improves hippocampal plasticity and cognitive behavior in adult healthy rats. Neuropharmacology 2020, 165, 107909. [Google Scholar] [CrossRef]
- Sale, A.; Putignano, E.; Cancedda, L.; Landi, S.; Cirulli, F.; Berardi, N.; Maffei, L. Enriched environment and acceleration of visual system development. Neuropharmacology 2004, 47, 649–660. [Google Scholar] [CrossRef]
- Scichilone, J.M.; Yarraguntla, K.; Charalambides, A.; Harney, J.P.; Butler, D. Environmental enrichment mitigates detrimental cognitive effects of ketogenic diet in weanling rats. J. Mol. Neurosci. 2016, 60, 1–9. [Google Scholar] [CrossRef]
- Diamond, M.C. Response of the brain to enrichment. An. Acad. Bras. Cienc. 2001, 73, 211–220. [Google Scholar] [CrossRef]
- Nithianantharajah, J.; Barkus, C.; Vijiaratnam, N.; Clement, O.; Hannan, A.J. Modeling brain reserve: Experience-dependent neuronal plasticity in healthy and Huntington’s disease transgenic mice. Am. J. Geriatr. Psychiatry 2009, 17, 196–209. [Google Scholar] [CrossRef]
- Estes, M.L.; McAllister, A.K. Maternal immune activation: Implications for neuropsychiatric disorders. Science 2016, 353, 772–777. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Rondón-Ortiz, A.N.; Lima, E.P.; Puracchio, M.; Roderick, R.C.; Kentner, A.C. Therapeutic efficacy of environmental enrichment on behavioral, endocrine, and synaptic alterations in an animal model of maternal immune activation. Brain Behav. Immun. Health 2020, 3, 100043. [Google Scholar] [CrossRef]
- Gong, X.; Lu, X.; Zhan, L.; Sui, H.; Qi, X.; Ji, Z.; Niu, X.; Liu, L. Role of the SNK-SPAR pathway in the development of Alzheimer’s disease. IUBMB Life 2010, 62, 214–221. [Google Scholar] [CrossRef]
- Wegner, A.M.; Nebhan, C.A.; Hu, L.; Majumdar, D.; Meier, K.M.; Weaver, A.M.; Webb, D.J. N-WASP and the Arp2/3 complex are critical regulators of actin in the development of dendritic spines and synapses. J. Biol. Chem. 2008, 283, 15912–15920. [Google Scholar] [CrossRef] [Green Version]
- Zhou, F.; Yin, G.; Gao, Y.; Ouyang, L.; Liu, S.; Jia, Q.; Yu, H.; Zha, Z.; Wang, K.; Xie, J.; et al. Insights into Cognitive deficits caused by low-dose toxic heavy metal mixtures and their remediation through a postnatal enriched environment in rats. J. Hazard. Mater. 2020, 388, 122081. [Google Scholar] [CrossRef]
- McAllister, B.B.; Thackray, S.E.; de la Orta, B.K.G.; Gosse, E.; Tak, P.; Chipak, C.; Rehal, S.; Rascon, A.V.; Dyck, R.H. Effects of enriched housing on the neuronal morphology of mice that lack zinc transporter 3 (ZnT3) and vesicular zinc. Behav. Brain Res. 2020, 379, 112336. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Meng, X.; El Fatimy, R.; Sun, B.; Mai, D.; Zhang, J.; Arora, R.; Zeng, A.; Xu, P.; Qu, S.; et al. Environmental enrichment prevents a beta oligomer-induced synaptic dysfunction through Mirna-132 and Hdac3 signaling pathways. Neurobiol. Dis. 2020, 134, 104617. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhang, Q.; Yu, K.; Shen, X.; Wu, Y.; Wu, J. Enriched Environment promoted cognitive function via bilateral synaptic remodeling after cerebral ischemia. Front. Neurol. 2019, 10, 1189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murueta-Goyena, A.; Morera-Herreras, T.; Miguelez, C.; Gutierrez-Ceballos, A.; Ugedo, L.; Vicente Lafuente, J.; Bengoetxea, H. Effects of adult enriched environment on cognition, hippocampal-prefrontal plasticity and NMDAR subunit expression in MK-801-induced schizophrenia model. Eur. Neuropsychopharmacol. 2019, 29, 590–600. [Google Scholar] [CrossRef]
- Bose, M.; Muñoz-Llancao, P.; Roychowdhury, S.; Nichols, J.A.; Jakkamsetti, V.; Porter, B.; Byrapureddy, R.; Salgado, H.; Kilgard, M.P.; Aboitiz, F.; et al. Effect of the environment on the dendritic morphology of the rat auditory cortex. Synapse 2010, 64, 97–110. [Google Scholar] [CrossRef] [Green Version]
- Woolery, A.; Myers, H.; Sternlieb, B.; Zeltzer, L. A yoga intervention for young adults with elevated symptoms of depression. Altern. Ther. Health Med. 2004, 10, 60–63. [Google Scholar]
- Galantino, M.L.; Bzdewka, T.M.; Eissler-Russo, J.L.; Holbrook, M.L.; Mogck, E.P.; Geigle, P.; Farrar, J.T. The impact of modified hatha yoga on chronic low back pain: A pilot study. Altern. Ther. Health Med. 2004, 10, 56–59. [Google Scholar]
- Epstein, G.N.; Halper, J.P.; Barrett, E.A.M.; Birdsall, C.; McGee, M.; Baron, K.P.; Lowenstein, S. A pilot study of mind-body changes in adults with asthma who practice mental imagery. Altern. Ther. Health Med. 2004, 10, 66–71. [Google Scholar]
- Villemure, C.; Ceko, M.; Cotton, V.A.; Bushnell, M.C. Neuroprotective effects of yoga practice: Age-, experience-, and frequency-dependent plasticity. Front. Hum. Neurosci. 2015, 9, 281. [Google Scholar] [CrossRef] [Green Version]
- Froeliger, B.; Garland, E.L.; McClernon, F.J. Yoga meditation practitioners exhibit greater gray matter volume and fewer reported cognitive failures: Results of a preliminary voxel-based morphometric analysis. Evid. Based Complement. Altern. Med. ECAM 2012, 2012, 821307. [Google Scholar] [CrossRef]
- Hernández, S.E.; Dorta, R.; Suero, J.; Barros-Loscertales, A.; González-Mora, J.L.; Rubia, K. Larger whole brain grey matter associated with long-term sahaja yoga meditation: A detailed area by area comparison. PLoS ONE 2020, 15, e0237552. [Google Scholar] [CrossRef]
- Hernández, S.E.; Barros-Loscertales, A.; Xiao, Y.; González-Mora, J.L.; Rubia, K. Gray matter and functional connectivity in anterior cingulate cortex are associated with the state of mental silence during sahaja yoga meditation. Neuroscience 2018, 371, 395–406. [Google Scholar] [CrossRef]
- Babu, M.G.R.; Kadavigere, R.; Koteshwara, P.; Sathian, B.; Rai, K.S. Rajyoga Meditation induces grey matter volume changes in regions that process reward and happiness. Sci. Rep. 2020, 10, 16177. [Google Scholar] [CrossRef]
- Hernández, S.E.; Suero, J.; Rubia, K.; González-Mora, J.L. Monitoring the neural activity of the state of mental silence while practicing sahaja yoga meditation. J. Altern. Complement. Med. 2015, 21, 175–179. [Google Scholar] [CrossRef]
- Hendriks, T. The effects of sahaja yoga meditation on mental health: A systematic review. J. Complement. Integr. Med. 2018, 15, 0163. [Google Scholar] [CrossRef]
- Froeliger, B.E.; Garland, E.L.; Modlin, L.A.; McClernon, F.J. Neurocognitive correlates of the effects of yoga meditation practice on emotion and cognition: A pilot study. Front. Integr. Neurosci. 2012, 6, 48. [Google Scholar] [CrossRef] [Green Version]
- Kassem, M.S.; Lagopoulos, J.; Stait-Gardner, T.; Price, W.S.; Chohan, T.W.; Arnold, J.C.; Hatton, S.N.; Bennett, M.R. Stress-induced grey matter loss determined by MRI Is primarily due to loss of dendrites and their synapses. Mol. Neurobiol. 2013, 47, 645–661. [Google Scholar] [CrossRef]
- Thas, J.J. Siddha medicine—Background and principles and the application for skin diseases. Clin. Dermatol. 2008, 26, 62–78. [Google Scholar] [CrossRef]
- Pandey, K.R.; Naik, S.R.; Vakil, B.V. Probiotics, prebiotics and synbiotics—A review. J. Food Sci. Technol. 2015, 52, 7577–7587. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Sl. No | Neurochemical System | Drug/Chemical Treatment | Effect on DS | References | |||||
|---|---|---|---|---|---|---|---|---|---|
| Number | Density | Length | Size | Morphogenesis | Enlargement | ||||
| 1 | Cholinergic | Donepezil, LM11A-3, Brahmi Nei, Phenserine tartarate, Huperzine A, | ↑ | ↑ | ↑ | - | - | - | [53,55,56,57,58] |
| 2 | Glutamatergic | Riluzole, Ketamine, LY341495 | ↑ | ↑ | - | - | - | - | [69,72,73] |
| 3 | GABAergic | Bicuculline, mercaptopropionic acid | - | ↑ | - | - | - | - | [79] |
| 4 | Serotonergic | LP-211, Olanzapine, Vortioxetine, | ↑ | ↑ | - | ↑ | ↑ | ↑ | [84,87,88] |
| 5 | Adrenergic | Guanfacine, Bromocriptine, isoproterenol | - | ↑ | ↑ | ↑ | - | - | [93,94,97] |
| 6 | Dopaminergic | SKF81297, Ropinirole, Pramipexole | ↑ | ↑ | ↑ | - | ↑ | - | [99,101] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahalakshmi, A.M.; Ray, B.; Tuladhar, S.; Hediyal, T.A.; Raj, P.; Rathipriya, A.G.; Qoronfleh, M.W.; Essa, M.M.; Chidambaram, S.B. Impact of Pharmacological and Non-Pharmacological Modulators on Dendritic Spines Structure and Functions in Brain. Cells 2021, 10, 3405. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10123405
Mahalakshmi AM, Ray B, Tuladhar S, Hediyal TA, Raj P, Rathipriya AG, Qoronfleh MW, Essa MM, Chidambaram SB. Impact of Pharmacological and Non-Pharmacological Modulators on Dendritic Spines Structure and Functions in Brain. Cells. 2021; 10(12):3405. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10123405
Chicago/Turabian StyleMahalakshmi, Arehally M., Bipul Ray, Sunanda Tuladhar, Tousif Ahmed Hediyal, Praveen Raj, Annan Gopinath Rathipriya, M. Walid Qoronfleh, Musthafa Mohamed Essa, and Saravana Babu Chidambaram. 2021. "Impact of Pharmacological and Non-Pharmacological Modulators on Dendritic Spines Structure and Functions in Brain" Cells 10, no. 12: 3405. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10123405