
Mechanical Control of Cell Migration by the Metastasis Suppressor Tetraspanin CD82/KAI1

 , , , , ,
, , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture and Transfections
2.3. Live Cell Imaging
2.4. Membrane Tether Measurements by AFM
2.5. Ultrastructural Analysis by Transmission Electron Microscopy (TEM)
2.6. Fabrication of Polydimethylsiloxane (PDMS) Gels
2.7. Immunofluorescence and Confocal Microscopy
2.8. Cell Lysates and Western Blots
2.9. RNA Extraction, Reverse Transcription and Real-Time RT-qPCR
- CYR61_For: ACCAAGAAATCCCCCGAACC
- CYR61_Rev: CGGGCAGTTGTAGTTGCATT
- CTGF_For: TTCCAAGACCTGTGGGAT
- CTGF_Rev: GTGCAGCCAGAAAGCTC
- AREG_For: CGAAGGACCAATGAGAGCCC
- AREG_Rev: AGGCATTTCACTCACAGGGG
- BIRC2_For: GTCAGAACACCGGAGGCATT
- BIRC2_Rev: TGACATCATCATTGCGACCCA
- CD82 For: ACTGGTTTCGTGGAAGGAAG
- CD82 _Rev: GCGCCCAGGATAAAGAAGAT
- YAP_For: GCTACAGTGTCCCTCGAACC
- YAP_REV: ACTTGGCATCAGCTCCTCTC
2.10. Statistical Analyses
3. Results
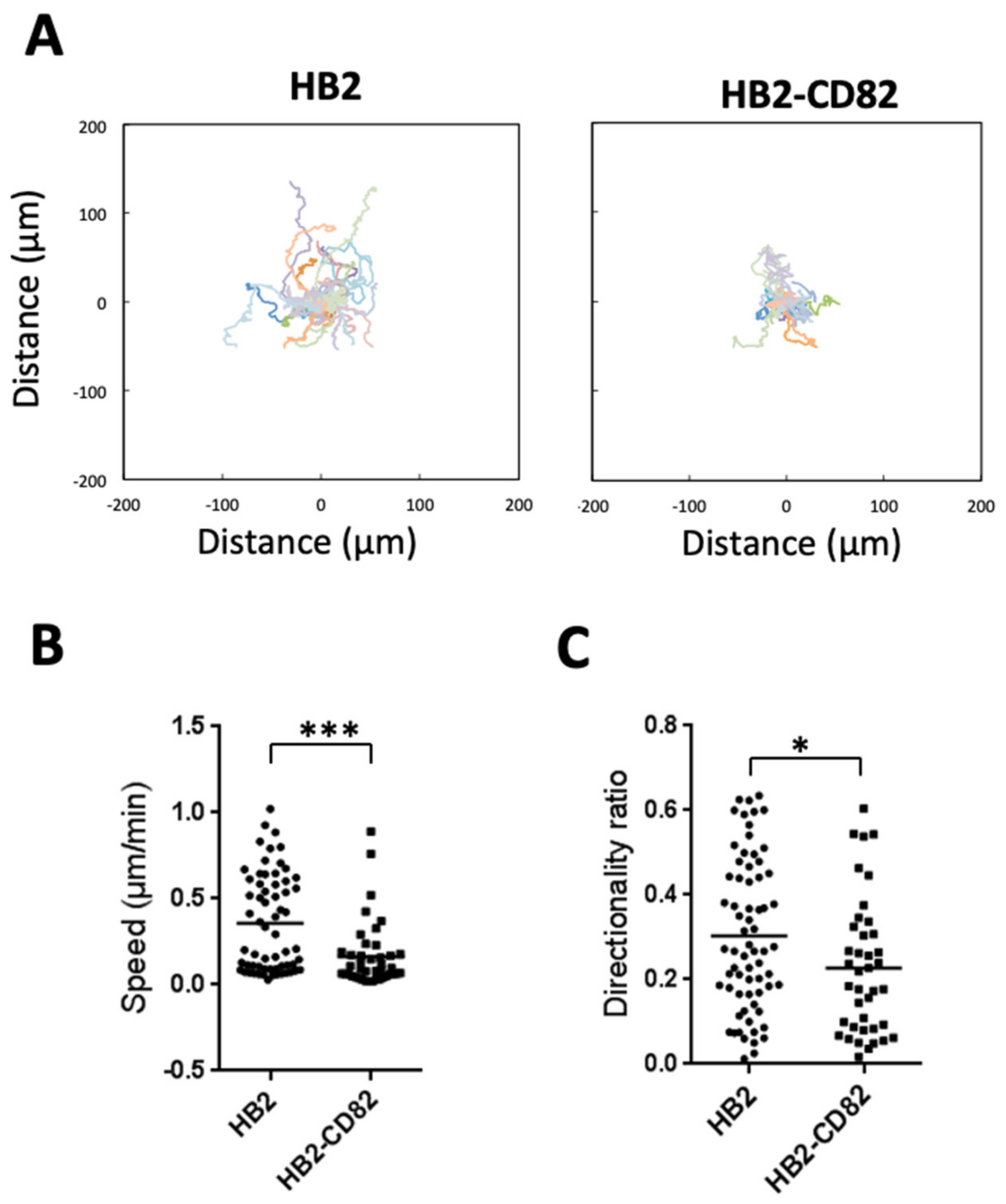
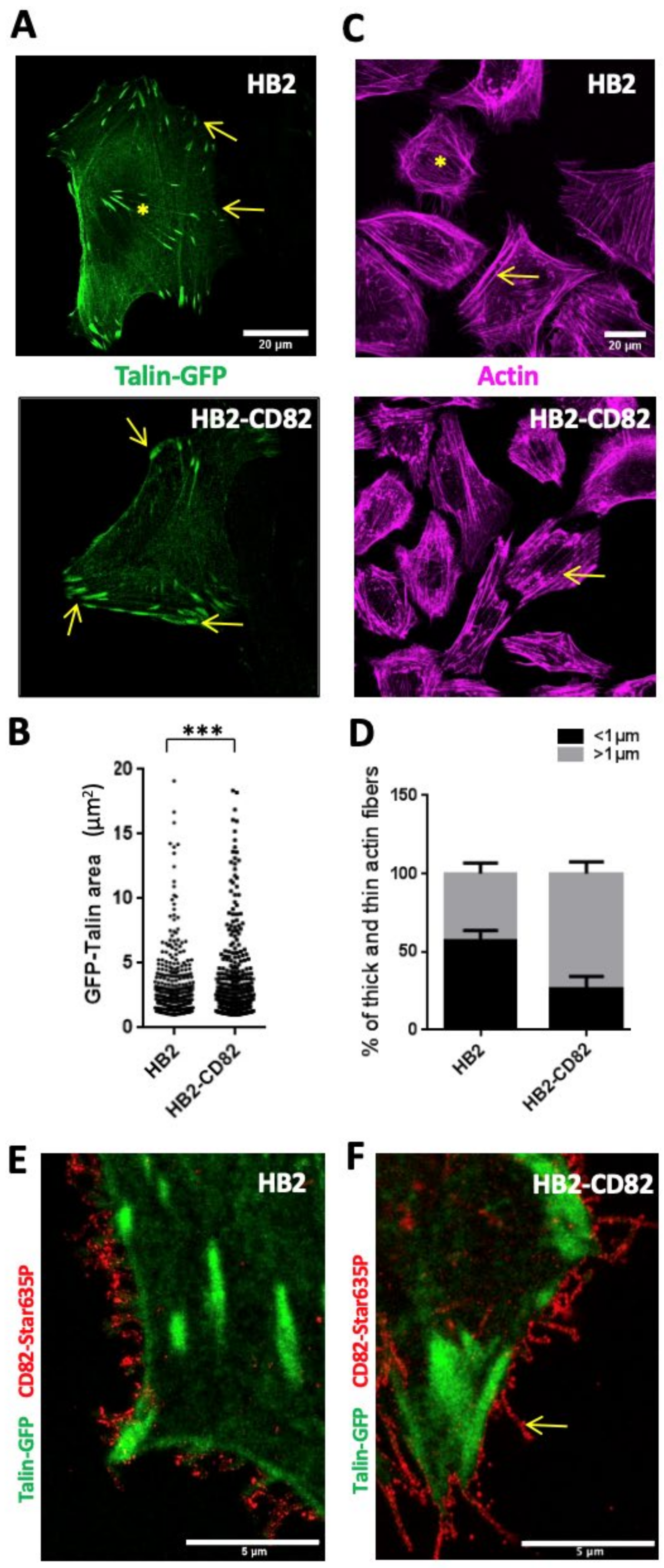
3.1. CD82 Regulates Persistent Cell Migration, Both Size and Dynamics of Focal Adhesions and Actin Polymerization
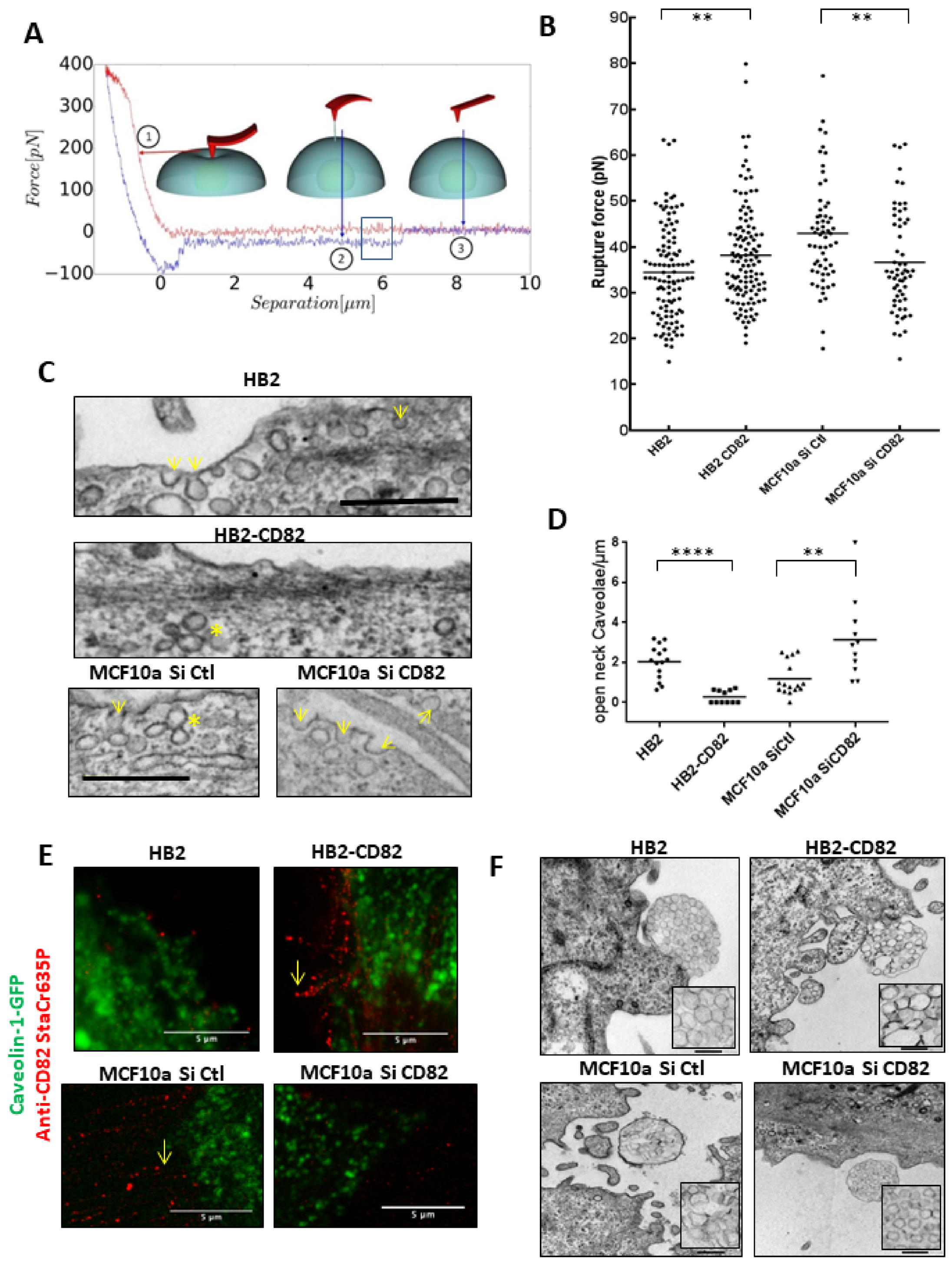
3.2. CD82 Regulates Membrane Tension and Cell Surface Caveolae Density
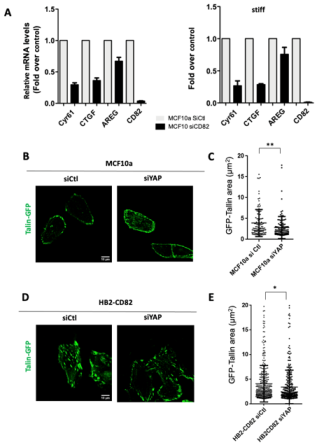
3.3. CD82 Regulates Caveolae Mechanosensing and YAP Nuclear Translocation and Activity
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Friedl, P.; Alexander, S. Cancer Invasion and the Microenvironment: Plasticity and Reciprocity. Cell 2011, 147, 992–1009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, J.; Yang, Q.; Huang, Q. Metastasis Suppressor Genes. Histol. Histopathol. 2013, 28, 285–292. [Google Scholar] [CrossRef]
- Tonoli, H.; Barrett, J.C. CD82 Metastasis Suppressor Gene: A Potential Target for New Therapeutics? Trends Mol. Med. 2005, 11, 563–570. [Google Scholar] [CrossRef] [PubMed]
- Charrin, S.; Jouannet, S.; Boucheix, C.; Rubinstein, E. Tetraspanins at a Glance. J. Cell. Sci. 2014, 127, 3641–3648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, Y.C.; Weissman, A.M. Dissecting the Diverse Functions of the Metastasis Suppressor CD82/KAI1. FEBS Lett. 2011, 585, 3166–3173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Odintsova, E.; Sugiura, T.; Berditchevski, F. Attenuation of EGF Receptor Signaling by a Metastasis Suppressor, the Tetraspanin CD82/KAI-1. Curr. Biol. 2000, 10, 1009–1012. [Google Scholar] [CrossRef] [Green Version]
- You, J.; Chang, R.; Liu, B.; Zu, L.; Zhou, Q. Nm23-H1 Was Involved in Regulation of KAI1 Expression in High-Metastatic Lung Cancer Cells L9981. J. Thorac. Dis. 2016, 8, 1217–1226. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.M.; Zhang, F.; Moshiach, S.; Zhou, B.; Huang, C.; Srinivasan, K.; Khurana, S.; Zheng, Y.; Lahti, J.M.; Zhang, X.A. Tetraspanin CD82 Inhibits Protrusion and Retraction in Cell Movement by Attenuating the Plasma Membrane-Dependent Actin Organization. PLoS ONE 2012, 7, e51797. [Google Scholar] [CrossRef]
- Lee, J.; Byun, H.-J.; Lee, M.-S.; Jin, Y.-J.; Jeoung, D.; Kim, Y.-M.; Lee, H. The Metastasis Suppressor CD82/KAI1 Inhibits Fibronectin Adhesion-Induced Epithelial-to-Mesenchymal Transition in Prostate Cancer Cells by Repressing the Associated Integrin Signaling. Oncotarget 2017, 8, 1641–1654. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, M.; Sugiura, T.; Abe, M.; Ishii, K.; Shirasuna, K. Regulation of C-Met Signaling by the Tetraspanin KAI-1/CD82 Affects Cancer Cell Migration. Int. J. Cancer 2007, 121, 1919–1929. [Google Scholar] [CrossRef]
- Termini, C.M.; Cotter, M.L.; Marjon, K.D.; Buranda, T.; Lidke, K.A.; Gillette, J.M. The Membrane Scaffold CD82 Regulates Cell Adhesion by Altering A4 Integrin Stability and Molecular Density. Mol. Biol. Cell 2014, 25, 1560–1573. [Google Scholar] [CrossRef] [PubMed]
- Odintsova, E.; Butters, T.D.; Monti, E.; Sprong, H.; Van Meer, G.; Berditchevski, F. Gangliosides Play an Important Role in the Organization of CD82-Enriched Microdomains. Biochem. J. 2006, 400, 315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bandyopadhyay, S.; Zhan, R.; Chaudhuri, A.; Watabe, M.; Pai, S.K.; Hirota, S.; Hosobe, S.; Tsukada, T.; Miura, K.; Takano, Y.; et al. Interaction of KAI1 on Tumor Cells with DARC on Vascular Endothelium Leads to Metastasis Suppression. Nat. Med. 2006, 12, 933–938. [Google Scholar] [CrossRef]
- Hur, J.; Choi, J.-I.; Lee, H.; Nham, P.; Kim, T.-W.; Chae, C.-W.; Yun, J.-Y.; Kang, J.-A.; Kang, J.; Lee, S.E.; et al. CD82/KAI1 Maintains the Dormancy of Long-Term Hematopoietic Stem Cells through Interaction with DARC-Expressing Macrophages. Cell Stem Cell 2016, 18, 508–521. [Google Scholar] [CrossRef] [Green Version]
- Mueller, J.; Szep, G.; Nemethova, M.; de Vries, I.; Lieber, A.D.; Winkler, C.; Kruse, K.; Small, J.V.; Schmeiser, C.; Keren, K.; et al. Load Adaptation of Lamellipodial Actin Networks. Cell 2017, 171, 188–200.e16. [Google Scholar] [CrossRef] [Green Version]
- Diz-Muñoz, A.; Krieg, M.; Bergert, M.; Ibarlucea-Benitez, I.; Muller, D.J.; Paluch, E.; Heisenberg, C.-P. Control of Directed Cell Migration in Vivo by Membrane-to-Cortex Attachment. PLoS Biol. 2010, 8, e1000544. [Google Scholar] [CrossRef] [PubMed]
- Pontes, B.; Monzo, P.; Gole, L.; Le Roux, A.-L.; Kosmalska, A.J.; Tam, Z.Y.; Luo, W.; Kan, S.; Viasnoff, V.; Roca-Cusachs, P.; et al. Membrane Tension Controls Adhesion Positioning at the Leading Edge of Cells. J. Cell Biol. 2017, 216, 2959–2977. [Google Scholar] [CrossRef] [Green Version]
- Sinha, B.; Köster, D.; Ruez, R.; Gonnord, P.; Bastiani, M.; Abankwa, D.; Stan, R.V.; Butler-Browne, G.; Vedie, B.; Johannes, L.; et al. Cells Respond to Mechanical Stress by Rapid Disassembly of Caveolae. Cell 2011, 144, 402–413. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Zucker, B.; Zhang, S.; Elias, S.; Zhu, Y.; Chen, H.; Ding, T.; Li, Y.; Sun, Y.; Lou, J.; et al. Migrasome Formation Is Mediated by Assembly of Micron-Scale Tetraspanin Macrodomains. Nat. Cell Biol. 2019, 21, 991–1002. [Google Scholar] [CrossRef]
- Parton, R.G.; Del Pozo, M.A.; Vassilopoulos, S.; Nabi, I.R.; Le Lay, S.; Lundmark, R.; Kenworthy, A.K.; Camus, A.; Blouin, C.M.; Sessa, W.C.; et al. Caveolae: The FAQs. Traffic 2020, 21, 181–185. [Google Scholar] [CrossRef]
- Moreno-Vicente, R.; Pavón, D.M.; Martín-Padura, I.; Català-Montoro, M.; Díez-Sánchez, A.; Quílez-Álvarez, A.; López, J.A.; Sánchez-Álvarez, M.; Vázquez, J.; Strippoli, R.; et al. Caveolin-1 Modulates Mechanotransduction Responses to Substrate Stiffness through Actin-Dependent Control of YAP. Cell Rep. 2018, 25, 1622–1635.e6. [Google Scholar] [CrossRef] [Green Version]
- Qiao, Y.; Chen, J.; Lim, Y.B.; Finch-Edmondson, M.L.; Seshachalam, V.P.; Qin, L.; Jiang, T.; Low, B.C.; Singh, H.; Lim, C.T.; et al. YAP Regulates Actin Dynamics through ARHGAP29 and Promotes Metastasis. Cell Rep. 2017, 19, 1495–1502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mason, D.E.; Collins, J.M.; Dawahare, J.H.; Nguyen, T.D.; Lin, Y.; Voytik-Harbin, S.L.; Zorlutuna, P.; Yoder, M.C.; Boerckel, J.D. YAP and TAZ Limit Cytoskeletal and Focal Adhesion Maturation to Enable Persistent Cell Motility. J. Cell Biol. 2019, 218, 1369–1389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.; Kim, D.-H.; Shah, S.R.; Kim, H.-N.; Kim, P.; Quiñones-Hinojosa, A.; Levchenko, A. Switch-like Enhancement of Epithelial-Mesenchymal Transition by YAP through Feedback Regulation of WT1 and Rho-Family GTPases. Nat. Commun. 2019, 10, 2797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, J.; Cao, B.; Wang, Y.; Ma, C.; Zeng, Z.; Liu, L.; Li, X.; Tao, D.; Gong, J.; Xie, D. Hippo Component YAP Promotes Focal Adhesion and Tumour Aggressiveness via Transcriptionally Activating THBS1/FAK Signalling in Breast Cancer. J. Exp. Clin. Cancer Res. 2018, 37, 175. [Google Scholar] [CrossRef]
- Rausch, V.; Bostrom, J.R.; Park, J.; Bravo, I.R.; Feng, Y.; Hay, D.C.; Link, B.A.; Hansen, C.G. The Hippo Pathway Regulates Caveolae Expression and Mediates Flow Response via Caveolae. Curr. Biol. 2019, 29, 242–255.e6. [Google Scholar] [CrossRef]
- Rubinstein, E.; Le Naour, F.; Lagaudriere-Gesbert, C.; Billard, M.; Conjeaud, H.; Boucheix, C. CD9, CD63, CD81, and CD82 Are Components of a Surface Tetraspan Network Connected to HLA-DR and VLA Integrins. Eur. J. Immunol. 1996, 26, 2657–2665. [Google Scholar] [CrossRef]
- Vera, J.; Lartigue, L.; Vigneron, S.; Gadea, G.; Gire, V.; Del Rio, M.; Soubeyran, I.; Chibon, F.; Lorca, T.; Castro, A. Greatwall Promotes Cell Transformation by Hyperactivating AKT in Human Malignancies. Elife 2015, 4. [Google Scholar] [CrossRef]
- Gorelik, R.; Gautreau, A. Quantitative and Unbiased Analysis of Directional Persistence in Cell Migration. Nat. Protoc. 2014, 9, 1931–1943. [Google Scholar] [CrossRef]
- Veracini, L.; Simon, V.; Richard, V.; Schraven, B.; Horejsi, V.; Roche, S.; Benistant, C. The Csk-Binding Protein PAG Regulates PDGF-Induced Src Mitogenic Signaling via GM1. J. Cell Biol. 2008, 182, 603–614. [Google Scholar] [CrossRef] [Green Version]
- Sadoun, A.; Puech, P.-H. Quantifying CD95/Cl-CD95L Implications in Cell Mechanics and Membrane Tension by Atomic Force Microscopy Based Force Measurements. Methods Mol. Biol. 2017, 1557, 139–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gavara, N. A Beginner’s Guide to Atomic Force Microscopy Probing for Cell Mechanics. Microsc. Res. Technique. 2017, 80, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Saavedra, V.O.; Fernandes, T.F.D.; Milhiet, P.-E.; Costa, L. Compression, Rupture, and Puncture of Model Membranes at the Molecular Scale. Langmuir 2020, 36, 5709–5716. [Google Scholar] [CrossRef]
- Tejera, E.; Rocha-Perugini, V.; López-Martín, S.; Pérez-Hernández, D.; Bachir, A.I.; Horwitz, A.R.; Vázquez, J.; Sánchez-Madrid, F.; Yáñez-Mo, M. CD81 Regulates Cell Migration through Its Association with Rac GTPase. Mol. Biol. Cell 2013, 24, 261–273. [Google Scholar] [CrossRef] [Green Version]
- Odintsova, E.; van Niel, G.; Conjeaud, H.; Raposo, G.; Iwamoto, R.; Mekada, E.; Berditchevski, F. Metastasis Suppressor Tetraspanin CD82/KAI1 Regulates Ubiquitylation of Epidermal Growth Factor Receptor. J. Biol. Chem. 2013, 288, 26323–26334. [Google Scholar] [CrossRef] [Green Version]
- Lorentzen, A.; Becker, P.F.; Kosla, J.; Saini, M.; Weidele, K.; Ronchi, P.; Klein, C.; Wolf, M.J.; Geist, F.; Seubert, B.; et al. Single Cell Polarity in Liquid Phase Facilitates Tumour Metastasis. Nat. Commun. 2018, 9, 887. [Google Scholar] [CrossRef] [Green Version]
- Vicente-Manzanares, M.; Webb, D.J.; Horwitz, A.R. Cell Migration at a Glance. J. Cell Sci. 2005, 118, 4917–4919. [Google Scholar] [CrossRef] [Green Version]
- Burridge, K. Focal Adhesions: A Personal Perspective on a Half Century of Progress. FEBS J. 2017, 284, 3355–3361. [Google Scholar] [CrossRef] [Green Version]
- Totaro, A.; Panciera, T.; Piccolo, S. YAP/TAZ Upstream Signals and Downstream Responses. Nat. Cell Biol. 2018, 20, 888–899. [Google Scholar] [CrossRef]
- Vidi, P.-A.; Maleki, T.; Ochoa, M.; Wang, L.; Clark, S.M.; Leary, J.F.; Lelièvre, S.A. Disease-on-a-Chip: Mimicry of Tumor Growth in Mammary Ducts. Lab. Chip 2014, 14, 172–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elosegui-Artola, A.; Andreu, I.; Beedle, A.E.M.; Lezamiz, A.; Uroz, M.; Kosmalska, A.J.; Oria, R.; Kechagia, J.Z.; Rico-Lastres, P.; Le Roux, A.-L.; et al. Force Triggers YAP Nuclear Entry by Regulating Transport across Nuclear Pores. Cell 2017, 171, 1397–1410.e14. [Google Scholar] [CrossRef]
- Ege, N.; Dowbaj, A.M.; Jiang, M.; Howell, M.; Hooper, S.; Foster, C.; Jenkins, R.P.; Sahai, E. Quantitative Analysis Reveals That Actin and Src-Family Kinases Regulate Nuclear YAP1 and Its Export. Cell Syst. 2018, 6, 692–708.e13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Northcott, J.M.; Dean, I.S.; Mouw, J.K.; Weaver, V.M. Feeling Stress: The Mechanics of Cancer Progression and Aggression. Front. Cell Dev. Biol. 2018, 6, 17. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yan, Q.; Sun, P.; Liu, J.-W.; Go, L.; McDaniel, S.M.; Paller, A.S. Suppression of Epidermal Growth Factor Receptor Signaling by Protein Kinase C-Alpha Activation Requires CD82, Caveolin-1, and Ganglioside. Cancer Res. 2007, 67, 9986–9995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vences-Catalán, F.; Kuo, C.-C.; Sagi, Y.; Chen, H.; Kela-Madar, N.; van Zelm, M.C.; van Dongen, J.J.M.; Levy, S. A Mutation in the Human Tetraspanin CD81 Gene Is Expressed as a Truncated Protein but Does Not Enable CD19 Maturation and Cell Surface Expression. J. Clin. Immunol. 2015, 35, 254–263. [Google Scholar] [CrossRef]
- Lazareth, H.; Henique, C.; Lenoir, O.; Puelles, V.G.; Flamant, M.; Bollée, G.; Fligny, C.; Camus, M.; Guyonnet, L.; Millien, C.; et al. The Tetraspanin CD9 Controls Migration and Proliferation of Parietal Epithelial Cells and Glomerular Disease Progression. Nat. Commun. 2019, 10, 3303. [Google Scholar] [CrossRef]
- Yu, X.; Li, S.; Pang, M.; Du, P.; Xu, T.; Bai, T.; Yang, T.; Hu, J.; Zhu, S.; Wang, L.; et al. TSPAN7 Exerts Anti-Tumor Effects in Bladder Cancer Through the PTEN/PI3K/AKT Pathway. Front. Oncol. 2020, 10, 613869. [Google Scholar] [CrossRef]
- Wang, X.; Lin, M.; Zhao, J.; Zhu, S.; Xu, M.; Zhou, X. TSPAN7 promotes the migration and proliferation of lung cancer cells via epithelial-to-mesenchymal transition. OncoTargets Therapy. 2018, 11, 8815–8822. [Google Scholar] [CrossRef] [Green Version]
- Pike, L.J. Lipid Rafts: Bringing Order to Chaos. J. Lipid. Res. 2003, 44, 655–667. [Google Scholar] [CrossRef] [Green Version]
- Colom, A.; Derivery, E.; Soleimanpour, S.; Tomba, C.; Molin, M.D.; Sakai, N.; González-Gaitán, M.; Matile, S.; Roux, A. A Fluorescent Membrane Tension Probe. Nat. Chem. 2018, 10, 1118–1125. [Google Scholar] [CrossRef]
- Charrin, S.; Manié, S.; Thiele, C.; Billard, M.; Gerlier, D.; Boucheix, C.; Rubinstein, E. A Physical and Functional Link between Cholesterol and Tetraspanins. Eur. J. Immunol. 2003, 33, 2479–2489. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-Y.; Yoon, S.-J.; Freire-de-Lima, L.; Kim, J.-H.; Hakomori, S. Control of Cell Motility by Interaction of Gangliosides, Tetraspanins, and Epidermal Growth Factor Receptor in A431 versus KB Epidermoid Tumor Cells. Carbohydr. Res. 2009, 344, 1479–1486. [Google Scholar] [CrossRef]
- Regina Todeschini, A.; Hakomori, S. Functional Role of Glycosphingolipids and Gangliosides in Control of Cell Adhesion, Motility, and Growth, through Glycosynaptic Microdomains. Biochim. Biophys. Acta 2008, 1780, 421–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danglot, L.; Chaineau, M.; Dahan, M.; Gendron, M.-C.; Boggetto, N.; Perez, F.; Galli, T. Role of TI-VAMP and CD82 in EGFR Cell-Surface Dynamics and Signaling. J. Cell Sci. 2010, 123, 723–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shvets, E.; Bitsikas, V.; Howard, G.; Hansen, C.G.; Nichols, B.J. Dynamic Caveolae Exclude Bulk Membrane Proteins and Are Required for Sorting of Excess Glycosphingolipids. Nat. Commun. 2015, 6, 6867. [Google Scholar] [CrossRef]
- Lee, J.H.; Park, S.R.; Chay, K.-O.; Seo, Y.-W.; Kook, H.; Ahn, K.Y.; Kim, Y.J.; Kim, K.K. KAI1 COOH-Terminal Interacting Tetraspanin (KITENIN), a Member of the Tetraspanin Family, Interacts with KAI1, a Tumor Metastasis Suppressor, and Enhances Metastasis of Cancer. Cancer Res. 2004, 64, 4235–4243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, P.; Godde, N.; Rubio, S.; Tekeste, M.; Vladar, E.K.; Axelrod, J.D.; Henderson, D.J.; Milgrom-Hoffman, M.; Humbert, P.O.; Hinck, L. VANGL2 Regulates Luminal Epithelial Organization and Cell Turnover in the Mammary Gland. Sci. Rep. 2019, 9, 7079. [Google Scholar] [CrossRef]
- Franklin, J.M.; Ghosh, R.P.; Shi, Q.; Reddick, M.P.; Liphardt, J.T. Concerted Localization-Resets Precede YAP-Dependent Transcription. Nat. Commun. 2020, 11, 4581. [Google Scholar] [CrossRef]
- Kantar, D.; Mur, E.B.; Mancini, M.; Slaninova, V.; Salah, Y.B.; Costa, L.; Forest, E.; Lassus, P.; Géminard, C.; Boissière-Michot, F.; et al. MAGI1 Inhibits the AMOTL2/P38 Stress Pathway and Prevents Luminal Breast Tumorigenesis. Sci. Rep. 2021, 11, 5752. [Google Scholar] [CrossRef]
- Srivastava, D.; de Toledo, M.; Manchon, L.; Tazi, J.; Juge, F. Modulation of Yorkie Activity by Alternative Splicing Is Required for Developmental Stability. EMBO J. 2021, 40, e104895. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ordas, L.; Costa, L.; Lozano, A.; Chevillard, C.; Calovoulos, A.; Kantar, D.; Fernandez, L.; Chauvin, L.; Dosset, P.; Doucet, C.; et al. Mechanical Control of Cell Migration by the Metastasis Suppressor Tetraspanin CD82/KAI1. Cells 2021, 10, 1545. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10061545
Ordas L, Costa L, Lozano A, Chevillard C, Calovoulos A, Kantar D, Fernandez L, Chauvin L, Dosset P, Doucet C, et al. Mechanical Control of Cell Migration by the Metastasis Suppressor Tetraspanin CD82/KAI1. Cells. 2021; 10(6):1545. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10061545
Chicago/Turabian StyleOrdas, Laura, Luca Costa, Anthony Lozano, Christopher Chevillard, Alexia Calovoulos, Diala Kantar, Laurent Fernandez, Lucie Chauvin, Patrice Dosset, Christine Doucet, and et al. 2021. "Mechanical Control of Cell Migration by the Metastasis Suppressor Tetraspanin CD82/KAI1" Cells 10, no. 6: 1545. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10061545