

Redox Balance Differentially Affects Biomechanics in Permeabilized Single Muscle Fibres—Active and Passive Force Assessments with the Myorobot
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Handling and Single Fibre Preparation
2.2. System Electronics and Software
2.3. Bioactive Solutions
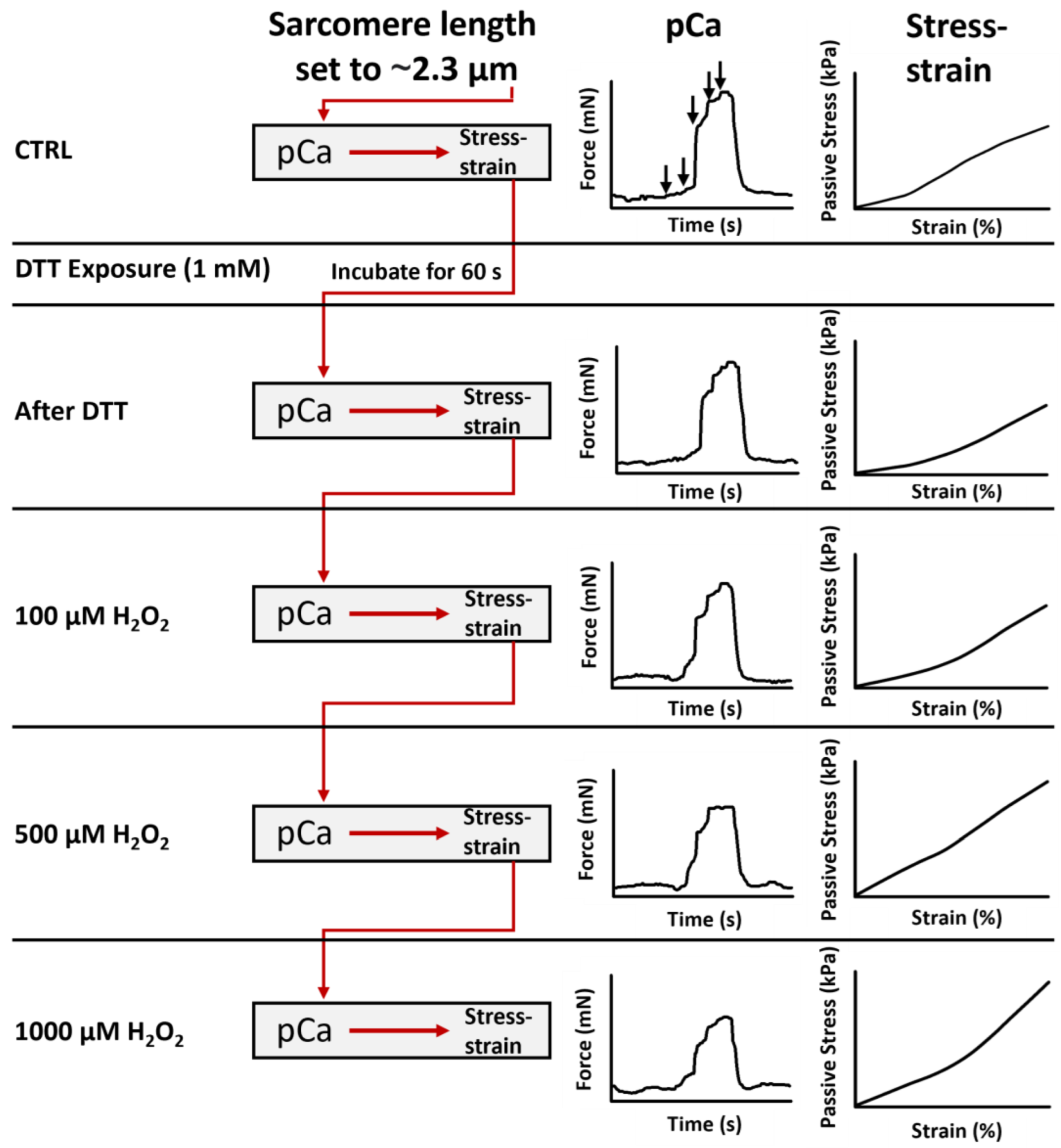
2.4. Active and Passive Force Assessment Protocols
3. Data Analysis
4. Results
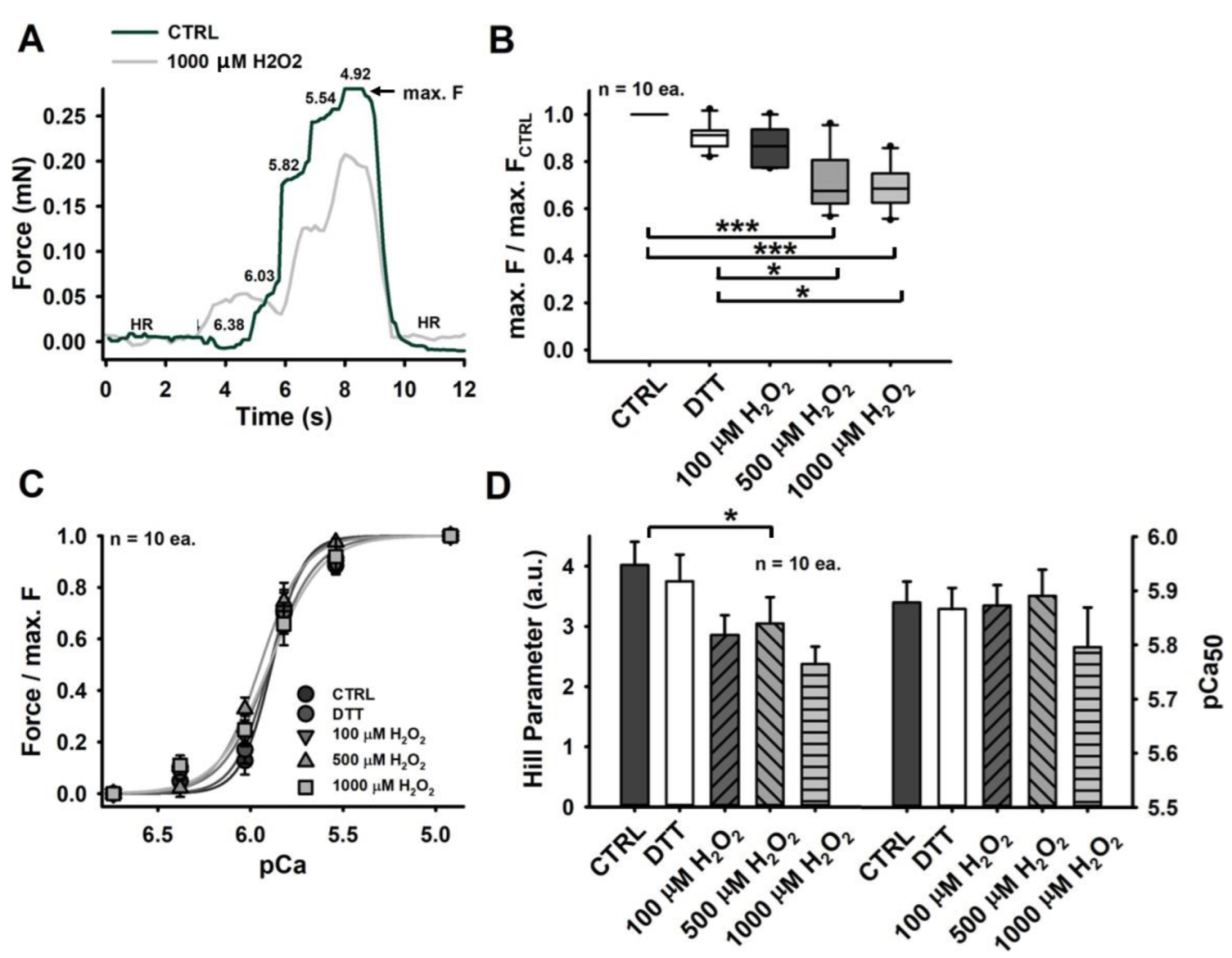
4.1. Maximum Ca2+ Saturated Force Is Diminished in Permeabilized Single Fibres in an Oxidizing Environment
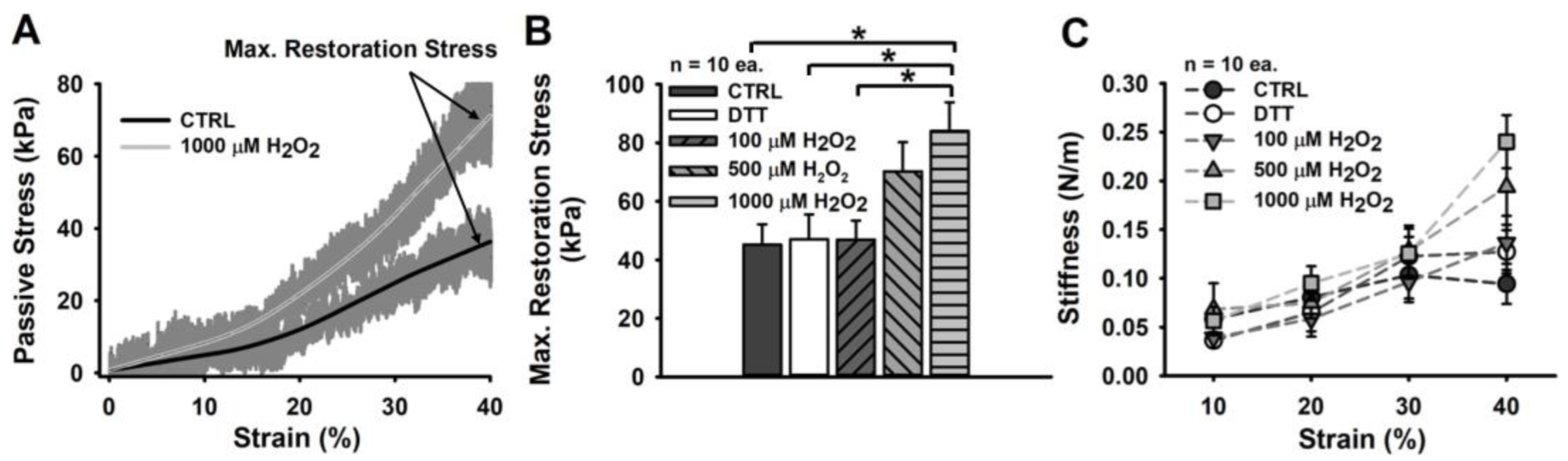
4.2. An Oxidizing Redox Balance Imposes a Non-Linear Fibre Stiffening onto Its Passive Strain Resistance
5. Discussion
5.1. Reduced Maximum Ca2+ Activated Force Does Not Originate from an Altered Ca2+ Sensitivity Inpermeabilized Muscle Fibres
5.2. An Oxidized Redox Balance Promotes Increased Stress with Strain in Permeabilized Muscle Fibres
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| EDL | M. Extensor Digitorum Longus |
| CTRL | Control |
| FT | Force Transducer |
| VC | Voice Coil |
| HA | High Activating (Solution) |
| HR | High Relaxing (Solution) |
| LR | Low Relaxing (Solution) |
| HKS | High-Potassium Solution |
| EDTA | Ethylene glycol-bis(β-aminoethylether))-N,N,N′,N′-tetraacetic acid |
| HDTA | Hexamethylenediaminetetraacetic acid |
| DTT | Dithiothreitol |
References
- Moopanar, T.R.; Allen, D.G. The activity-induced reduction of myofibrillar Ca2+ sensitivity in mouse skeletal muscle is reversed by dithiothreitol. J. Physiol. 2006, 571, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Reid, M.B. Invited Review: Redox modulation of skeletal muscle contraction: What we know and what we don’t. J. Appl. Physiol. 2001, 90, 724–731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamb, G.D.; Posterino, G.S. Effects of oxidation and reduction on contractile function in skeletal muscle fibres of the rat. J. Physiol. 2003, 546, 149–163. [Google Scholar] [CrossRef] [PubMed]
- Andrade, F.H.; Reid, M.B.; Allen, D.G.; Westerblad, H. Effect of hydrogen peroxide and dithiothreitol on contractile function of single skeletal muscle fibres from the mouse. J. Physiol. 1998, 509, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Darnley, G.M.; Duke, A.M.; Steele, D.S.; MacFarlane, N.G. Effects of reactive oxygen species on aspects of excitation-contraction coupling in chemically skinned rabbit diaphragm muscle fibres. Exp. Physiol. 2001, 86, 161–168. [Google Scholar] [CrossRef] [Green Version]
- Posterino, G.S.; Cellini, M.A.; Lamb, G.D. Effects of oxidation and cytosolic redox conditions on excitation-contraction coupling in rat skeletal muscle. J. Physiol. 2003, 547, 807–823. [Google Scholar] [CrossRef]
- Smith, M.A.; Reid, M.B. Redox modulation of contractile function in respiratory and limb skeletal muscle. Respir. Physiol. Neurobiol. 2006, 151, 229–241. [Google Scholar] [CrossRef]
- Willingham, T.B.; Ajayi, P.T.; Glancy, B. Subcellular Specialization of Mitochondrial Form and Function in Skeletal Muscle Cells. Front. Cell Dev. Biol. 2021, 9, 757305. [Google Scholar] [CrossRef] [PubMed]
- Murphy, R.M.; Dutka, T.L.; Lamb, G.D. Hydroxyl radical and glutathione interactions alter calcium sensitivity and maximum force of the contractile apparatus in rat skeletal muscle fibres. J. Physiol. 2008, 586, 2203–2216. [Google Scholar] [CrossRef]
- Callahan, L.A.; She, Z.W.; Nosek, T.M. Superoxide, hydroxyl radical, and hydrogen peroxide effects on single-diaphragm fiber contractile apparatus. J. Appl. Physiol. 2001, 90, 45–54. [Google Scholar] [CrossRef]
- Reid, M.B.; Haack, K.E.; Franchek, K.M.; Valberg, P.A.; Kobzik, L.; West, M.S. Reactive oxygen in skeletal muscle. I. Intracellular oxidant kinetics and fatigue in vitro. J. Appl. Physiol. 1992, 73, 1797–1804. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.L.; Yeo, D.; Kang, C.; Zhang, T. The role of mitochondria in redox signaling of muscle homeostasis. J. Sport Health Sci. 2020, 9, 386–393. [Google Scholar] [CrossRef] [PubMed]
- Zuo, L.; Christofi, F.L.; Wright, V.P.; Liu, C.Y.; Merola, A.J.; Berliner, L.J.; Clanton, T.L. Intra- and extracellular measurement of reactive oxygen species produced during heat stress in diaphragm muscle. Am. J. Physiol.-Cell Physiol. 2000, 279, C1058–C1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brotto, M.A.; Nosek, T.M. Hydrogen peroxide disrupts Ca2+ release from the sarcoplasmic reticulum of rat skeletal muscle fibers. J. Appl. Physiol. 1996, 81, 731–737. [Google Scholar] [CrossRef]
- MacFarlane, N.G.; Miller, D.J. Depression of peak force without altering calcium sensitivity by the superoxide anion in chemically skinned cardiac muscle of rat. Circ. Res. 1992, 70, 1217–1224. [Google Scholar] [CrossRef] [Green Version]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine; Oxford University Press: Oxford, UK, 2015. [Google Scholar] [CrossRef]
- Van der Poel, C.; Stephenson, D.G. Reversible changes in Ca2+-activation properties of rat skeletal muscle exposed to elevated physiological temperatures. J. Physiol. 2002, 544, 765–776. [Google Scholar] [CrossRef]
- Prochniewicz, E.; Lowe, D.A.; Spakowicz, D.J.; Higgins, L.; O’Conor, K.; Thompson, L.V.; Ferrington, D.A.; Thomas, D.D. Functional, structural, and chemical changes in myosin associated with hydrogen peroxide treatment of skeletal muscle fibers. Am. J. Physiol.-Cell Physiol. 2008, 294, C613–C626. [Google Scholar] [CrossRef] [Green Version]
- Haug, M.; Ritter, P.; Michael, M.; Reischl, B.; Schurmann, S.; Prols, G.; Friedrich, O. Structure-Function Relationships in Muscle Fibres: MyoRobot Online Assessment of Muscle Fibre Elasticity and Sarcomere Length Distributions. IEEE Trans. Biomed. Eng. 2022, 69, 148–155. [Google Scholar] [CrossRef]
- Haug, M.; Meyer, C.; Reischl, B.; Prölß, G.; Nübler, S.; Schürmann, S.; Schneidereit, D.; Heckel, M.; Pöschel, T.; Rupitsch, S.J.; et al. MyoRobot 2.0: An advanced biomechatronics platform for automated, environmentally controlled skeletal muscle single fiber biomechanics assessment employing inbuilt real-time optical imaging. Biosens. Bioelectron. 2019, 138, 111284. [Google Scholar] [CrossRef]
- Launikonis, B.S.; Stephenson, D.G. Effect of saponin treatment on the sarcoplasmic reticulum of rat, cane toad and crustacean (Yabby) skeletal muscle. J. Physiol. 1997, 504, 425–437. [Google Scholar] [CrossRef] [Green Version]
- Posterino, G.S.; Lamb, G.D.; Stephenson, D.G. Twitch and tetanic force responses and longitudinal propagation of action potentials in skinned skeletal muscle fibres of the rat. J. Physiol. 2000, 527, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Haug, M.; Meyer, C.; Reischl, B.; Prölß, G.; Vetter, K.; Iberl, J.; Nübler, S.; Schürmann, S.; Rupitsch, S.J.; Heckel, M.; et al. The MyoRobot technology discloses a premature biomechanical decay of skeletal muscle fiber bundles derived from R349P desminopathy mice. Sci. Rep. 2019, 9, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedrich, O.; Yi, B.; Edwards, J.N.; Reischl, B.; Wirth-Huecking, A.; Buttgereit, A.; Lang, R.; Polyak, F.; Liu, I.; Wegner, F.v.; et al. IL-1α reversibly inhibits skeletal muscle ryanodine receptor. A novel mechanism for critical illness myopathy? Am. J. Respir. Cell Mol. Biol. 2014, 50, 1096–1106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamb, G.D.; Stephenson, D.G. Measurement of force and calcium release using mechanically skinned fibers from mammalian skeletal muscle. J. Appl. Physiol. 2018, 125, 1105–1127. [Google Scholar] [CrossRef] [PubMed]
- Hill, A.V. The possible effects of the aggregation of the molecules of haemoglobin on its oxygen dissociation. J. Physiol. 1910, 40, 4–7. [Google Scholar]
- Haug, M.; Reischl, B.; Prölß, G.; Pollmann, C.; Buckert, T.; Keidel, C.; Schürmann, S.; Hock, M.; Rupitsch, S.; Heckel, M.; et al. The MyoRobot: A novel automated biomechatronics system to assess voltage/Ca2+ biosensors and active/passive biomechanics in muscle and biomaterials. Biosens. Bioelectron. 2018, 102, 589–599. [Google Scholar] [CrossRef]
- Moopanar, T.R.; Allen, D.G. Reactive oxygen species reduce myofibrillar Ca2+ sensitivity in fatiguing mouse skeletal muscle at 37 degrees C. J. Physiol. 2005, 564, 189–199. [Google Scholar] [CrossRef]
- Bruton, J.D.; Place, N.; Yamada, T.; Silva, J.P.; Andrade, F.H.; Dahlstedt, A.J.; Zhang, S.J.; Katz, A.; Larsson, N.G.; Westerblad, H. Reactive oxygen species and fatigue-induced prolonged low-frequency force depression in skeletal muscle fibres of rats, mice and SOD2 overexpressing mice. J. Physiol. 2008, 586, 175–184. [Google Scholar] [CrossRef]
- Plant, D.R.; Lynch, G.S.; Williams, D.A. Hydrogen peroxide modulates Ca2+-activation of single permeabilized fibres from fast- and slow-twitch skeletal muscles of rats. J. Muscle Res. Cell Motil. 2000, 21, 747–752. [Google Scholar] [CrossRef]
- Pollmann, C.; Haug, M.; Reischl, B.; Prölß, G.; Pöschel, T.; Rupitsch, S.J.; Clemen, C.S.; Schröder, R.; Friedrich, O. Growing Old Too Early: Skeletal Muscle Single Fiber Biomechanics in Ageing R349P Desmin Knock-in Mice Using the MyoRobot Technology. Int. J. Mol. Sci. 2020, 21, 5501. [Google Scholar] [CrossRef]
- Haug, M.; Reischl, B.; Nübler, S.; Kiriaev, L.; Mázala, D.A.G.; Houweling, P.J.; North, K.N.; Friedrich, O.; Head, S.I. Absence of the Z-disc protein a-actinin-3 impairs the mechanical stability of Actn3KO mouse fast-twitch muscle fibres without altering their contractile properties or twitch kinetics. Skelet. Muscle 2022, 12, 14. [Google Scholar] [CrossRef] [PubMed]
- Fink, R.H.; Stephenson, D.G.; Williams, D.A. Calcium and strontium activation of single skinned muscle fibres of normal and dystrophic mice. J. Physiol. 1986, 373, 513–525. [Google Scholar] [CrossRef] [PubMed]
- Chan, S.; Seto, J.T.; MacArthur, D.G.; Yang, N.; North, K.N.; Head, S.I. A gene for speed: Contractile properties of isolated whole EDL muscle from an alpha-actinin-3 knockout mouse. Am. J. Physiol.-Cell Physiol. 2008, 295, C897–C904. [Google Scholar] [CrossRef] [Green Version]
- Choi, M.H.; Ow, J.R.; Yang, N.D.; Taneja, R. Oxidative Stress-Mediated Skeletal Muscle Degeneration: Molecules, Mechanisms, and Therapies. Oxidative Med. Cell. Longev. 2016, 2016, 6842568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, I.A.; Allen, D.G. The role of reactive oxygen species in the hearts of dystrophin-deficient mdx mice. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H1969–H1977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheresh, P.; Kim, S.J.; Tulasiram, S.; Kamp, D.W. Oxidative stress and pulmonary fibrosis. Biochim. Biophys. Acta 2013, 1832, 1028–1040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richter, K.; Kietzmann, T. Reactive oxygen species and fibrosis: Further evidence of a significant liaison. Cell Tissue Res. 2016, 365, 591–605. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Gonzalez, F.J.; Chandel, N.S.; Jain, M.; Budinger, G.R.S. Reactive oxygen species as signaling molecules in the development of lung fibrosis. Transl. Res. J. Lab. Clin. Med. 2017, 190, 61–68. [Google Scholar] [CrossRef]
- Alamdari, N.; Toraldo, G.; Aversa, Z.; Smith, I.; Castillero, E.; Renaud, G.; Qaisar, R.; Larsson, L.; Jasuja, R.; Hasselgren, P.O. Loss of muscle strength during sepsis is in part regulated by glucocorticoids and is associated with reduced muscle fiber stiffness. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 303, R1090–R1099. [Google Scholar] [CrossRef] [Green Version]
- Goossens, C.; Weckx, R.; Derde, S.; van Helleputte, L.; Schneidereit, D.; Haug, M.; Reischl, B.; Friedrich, O.; van den Bosch, L.; van den Berghe, G.; et al. Impact of prolonged sepsis on neural and muscular components of muscle contractions in a mouse model. J. Cachexia Sarcopenia Muscle 2021, 12, 443–455. [Google Scholar] [CrossRef]
- Judge, S.M.; Nosacka, R.L.; Delitto, D.; Gerber, M.H.; Cameron, M.E.; Trevino, J.G.; Judge, A.R. Skeletal Muscle Fibrosis in Pancreatic Cancer Patients with Respect to Survival. JNCI Cancer Spectr. 2018, 2, pky043. [Google Scholar] [CrossRef] [PubMed]
- Bjørklund, G.; Dadar, M.; Aaseth, J.; Chirumbolo, S.; Pen, J.J. Cancer-associated Cachexia, Reactive Oxygen Species and Nutrition Therapy. Curr. Med. Chem. 2019, 26, 5728–5744. [Google Scholar] [CrossRef] [PubMed]
- Finkel, T. Redox-dependent signal transduction. FEBS Lett. 2000, 476, 52–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alegre-Cebollada, J.; Kosuri, P.; Giganti, D.; Eckels, E.; Rivas-Pardo, J.A.; Hamdani, N.; Warren, C.M.; Solaro, R.J.; Linke, W.A.; Fernández, J.M. S-glutathionylation of cryptic cysteines enhances titin elasticity by blocking protein folding. Cell 2014, 156, 1235–1246. [Google Scholar] [CrossRef] [Green Version]
- Hoshi, T.; Heinemann, S. Regulation of cell function by methionine oxidation and reduction. J. Physiol. 2001, 531, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.H.; Lee, K.; Kwak, G.H.; Park, Y.S.; Lee, K.J.; Hwang, K.Y.; Kim, H.Y. Evidence for the dimerization-mediated catalysis of methionine sulfoxide reductase A from Clostridium oremlandii. PLoS ONE 2015, 10, e0131523. [Google Scholar] [CrossRef]
- Meyer, G.A.; Lieber, R.L. Elucidation of extracellular matrix mechanics from muscle fibers and fiber bundles. J. Biomech. 2011, 44, 771–773. [Google Scholar] [CrossRef] [Green Version]
- Anderson, J.; Joumaa, V.; Stevens, L.; Neagoe, C.; Li, Z.; Mounier, Y.; Linke, W.; Goubel, F. Passive stiffness changes in soleus muscles from desmin knockout mice are not due to titin modifications. Pfluegers Arch. Eur. J. Physiol. 2002, 444, 771–776. [Google Scholar] [CrossRef]
- Mutungi, G.; Ranatunga, K.W. The viscous, viscoelastic and elastic characteristics of resting fast and slow mammalian (rat) muscle fibres. J. Physiol. 1996, 496, 827–836. [Google Scholar] [CrossRef]
- Gillies, A.R.; Lieber, R.L. Structure and function of the skeletal muscle extracellular matrix. Muscle Nerve 2011, 44, 318–331. [Google Scholar] [CrossRef]
- Wood, L.K.; Kayupov, E.; Gumucio, J.P.; Mendias, C.L.; Claflin, D.R.; Brooks, S.V. Intrinsic stiffness of extracellular matrix increases with age in skeletal muscles of mice. J. Appl. Physiol. 2014, 117, 363–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varland, S.; Vandekerckhove, J.; Drazic, A. Actin Post-translational Modifications: The Cinderella of Cytoskeletal Control. Trends Biochem. Sci. 2019, 44, 502–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koser, F.; Loescher, C.; Linke, W.A. Posttranslational modifications of titin from cardiac muscle: How, where, and what for? FEBS J. 2019, 286, 2240–2260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grützner, A.; Garcia-Manyes, S.; Kötter, S.; Badilla, C.L.; Fernandez, J.M.; Linke, W.A. Modulation of Titin-Based Stiffness by Disulfide Bonding in the Cardiac Titin N2-B Unique Sequence. Biophys. J. 2009, 97, 825–834. [Google Scholar] [CrossRef] [Green Version]
- Loescher, C.M.; Breitkreuz, M.; Li, Y.; Nickel, A.; Unger, A.; Dietl, A.; Schmidt, A.; Mohamed, B.A.; Kötter, S.; Schmitt, J.P.; et al. Regulation of titin-based cardiac stiffness by unfolded domain oxidation (UnDOx). Proc. Natl. Acad. Sci. USA 2020, 117, 24545–24556. [Google Scholar] [CrossRef]
- Corpeno, R.; Dworkin, B.; Cacciani, N.; Salah, H.; Bergman, H.M.; Ravara, B.; Vitadello, M.; Gorza, L.; Gustafson, A.M.; Hedström, Y.; et al. Time course analysis of mechanical ventilation-induced diaphragm contractile muscle dysfunction in the rat. J. Physiol. 2014, 592, 3859–3880. [Google Scholar] [CrossRef]
- Bibli, S.I.; Fleming, I. Oxidative Post-Translational Modifications: A Focus on Cysteine S-Sulfhydration and the Regulation of Endothelial Fitness. Antioxid. Redox Signal. 2021, 35, 1494–1514. [Google Scholar] [CrossRef]
- Herzog, W.; Schappacher, G.; DuVall, M.; Leonard, T.R.; Herzog, J.A. Residual Force Enhancement Following Eccentric Contractions: A New Mechanism Involving Titin. Physiology 2016, 31, 300–312. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Lang, P.; Linke, W.A. Titin stiffness modifies the force-generating region of muscle sarcomeres. Sci. Rep. 2016, 6, 24492. [Google Scholar] [CrossRef]
- Preetha, N.; Yiming, W.; Helmes, M.; Norio, F.; Siegfried, L.; Granzier, H. Restoring force development by titin/connectin and assessment of Ig domain unfolding. J. Muscle Res. Cell Motil. 2005, 26, 307–317. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Well | Bioactive Solution | Purpose |
|---|---|---|
| 1 | Saponin (0.01% w/v) | fibre skinning |
| 2 | LR | replace EGTA with HDTA, relax |
| 3 | HR | buffer excess Ca2+, relax |
| 4 | pCa 6.38 | initiate contraction |
| 5 | pCa 6.03 | initiate contraction |
| 6 | pCa 5.82 | initiate contraction |
| 7 | pCa 5.54 | initiate contraction |
| 8 | pCa 4.92 | initiate maximum contraction |
| 9 | DTT | reducing environment |
| 10 | LR (100 µM H2O2) | replace EGTA with HDTA, relax |
| 11 | HR (100 µM H2O2) | buffer excess Ca2+, relax |
| 12 | pCa 6.38 (100 µM H2O2) | initiate contraction |
| 13 | pCa 6.03 (100 µM H2O2) | initiate contraction |
| 14 | pCa 5.82 (100 µM H2O2) | initiate contraction |
| 15 | idle | relaxing environment |
| 16 | pCa 5.54 (100 µM H2O2) | initiate contraction |
| 17 | pCa 4.92 (100 µM H2O2) | initiate maximum contraction |
| 18 | LR (500 µM H2O2) | replace EGTA with HDTA, relax |
| 19 | HR (500 µM H2O2) | buffer excess Ca2+, relax |
| 20 | pCa 6.38 (500 µM H2O2) | initiate contraction |
| 21 | pCa 6.03 (500 µM H2O2) | initiate contraction |
| 22 | pCa 5.82 (500 µM H2O2) | initiate contraction |
| 23 | pCa 5.54 (500 µM H2O2) | initiate contraction |
| 24 | pCa 4.92 (500 µM H2O2) | initiate maximum contraction |
| 25 | LR (1000 µM H2O2) | replace EGTA with HDTA, relax |
| 26 | HR (1000 µM H2O2) | buffer excess Ca2+, relax |
| 27 | pCa 6.38 (1000 µM H2O2) | initiate contraction |
| 28 | pCa 6.03 (1000 µM H2O2) | initiate contraction |
| 29 | pCa 5.82 (1000 µM H2O2) | initiate contraction |
| 30 | pCa 5.54 (1000 µM H2O2) | initiate contraction |
| 31 | pCa 4.92 (1000 µM H2O2) | initiate maximum contraction |
| Stress (kPa) | CTRL | DTT | 100 µM H2O2 | 500 µM H2O2 | 1000 µM H2O2 |
|---|---|---|---|---|---|
| Contractile Restoration stress at 140% L0 (∼3.2 µm SL) | 172 ± 20 # 45 ± 7 # | 156 ± 19 # 47 ± 8 # | 148 ± 17 # 47 ± 6 # | 130 ± 19 *# 70 ± 9 # | 118 ± 14 *# 84 ± 9 *# |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Michael, M.; Kovbasyuk, L.; Ritter, P.; Reid, M.B.; Friedrich, O.; Haug, M. Redox Balance Differentially Affects Biomechanics in Permeabilized Single Muscle Fibres—Active and Passive Force Assessments with the Myorobot. Cells 2022, 11, 3715. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11233715
Michael M, Kovbasyuk L, Ritter P, Reid MB, Friedrich O, Haug M. Redox Balance Differentially Affects Biomechanics in Permeabilized Single Muscle Fibres—Active and Passive Force Assessments with the Myorobot. Cells. 2022; 11(23):3715. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11233715
Chicago/Turabian StyleMichael, Mena, Larisa Kovbasyuk, Paul Ritter, Michael B. Reid, Oliver Friedrich, and Michael Haug. 2022. "Redox Balance Differentially Affects Biomechanics in Permeabilized Single Muscle Fibres—Active and Passive Force Assessments with the Myorobot" Cells 11, no. 23: 3715. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11233715