
Dihydromyricetin Attenuates Depressive-like Behaviors in Mice by Inhibiting the AGE-RAGE Signaling Pathway

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Animals
2.2. Chemicals
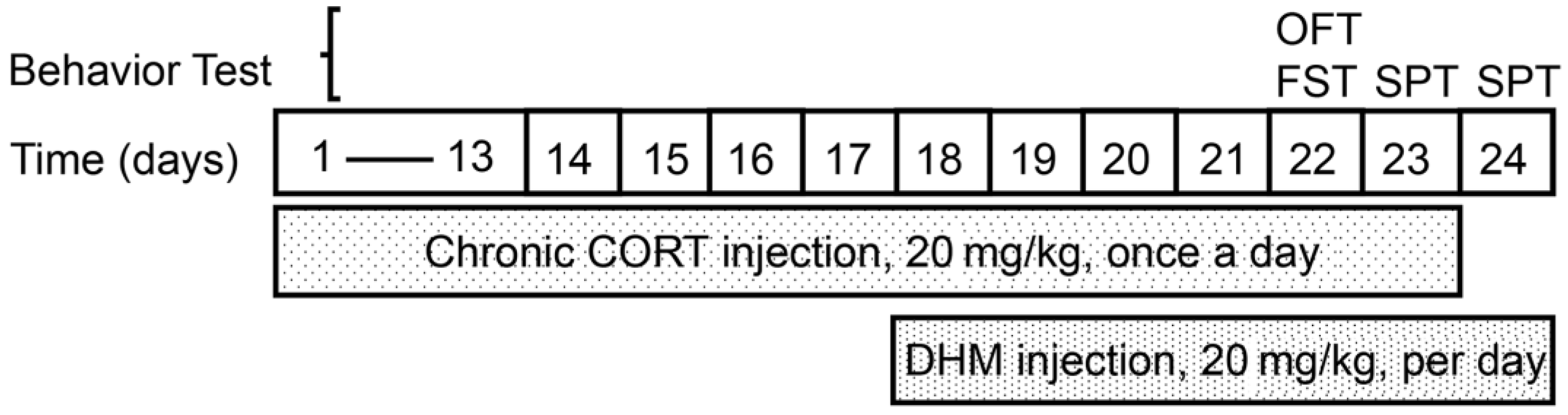
2.3. Animal Treatment
2.4. Behavior Tasks
2.4.1. Open Field Test (OFT)
2.4.2. Forced Swimming Test (FST)
2.4.3. Sucrose Preference Test (SPT)
2.5. Western Blotting
2.6. Enzyme-Linked Immunosorbent Assay (ELISA)
2.7. Network Pharmacology Analyses
2.7.1. Prediction of DHM and Depression Targets
2.7.2. Construction of Venn Diagram and DHM Targets Depression Network
2.7.3. Construction of Protein–Protein Interaction (PPI) Network
2.7.4. Gene Ontology (GO) Function and Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway Enrichment
2.8. Statistical Analysis
3. Results
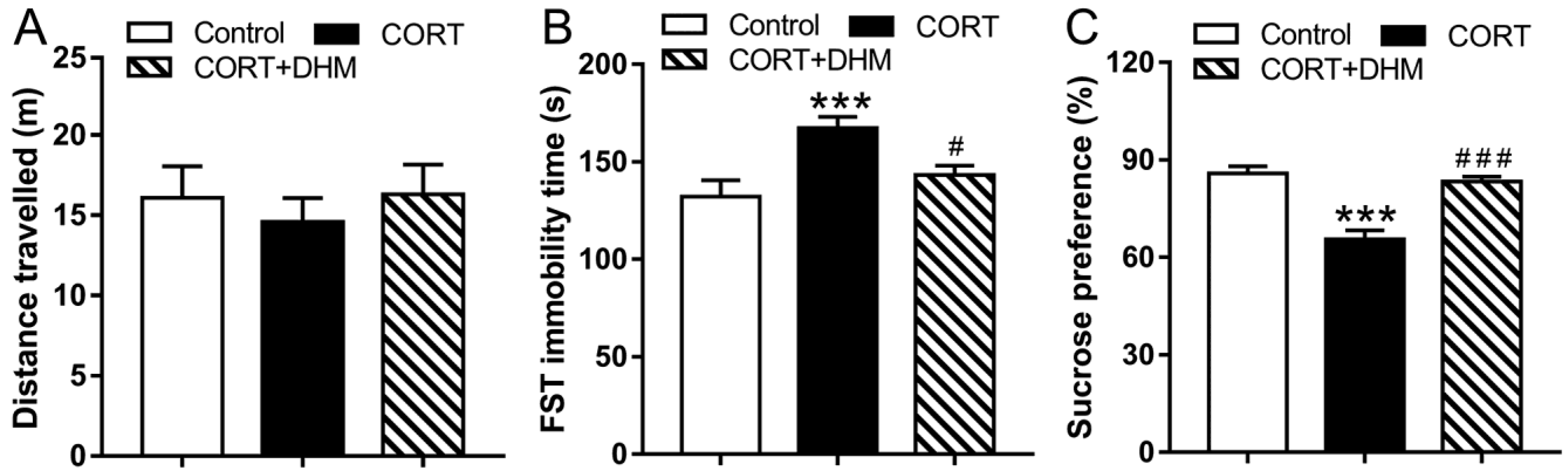
3.1. Dihydromyricetin Alleviated CORT-Induced Depressive-like Behaviors in Mice
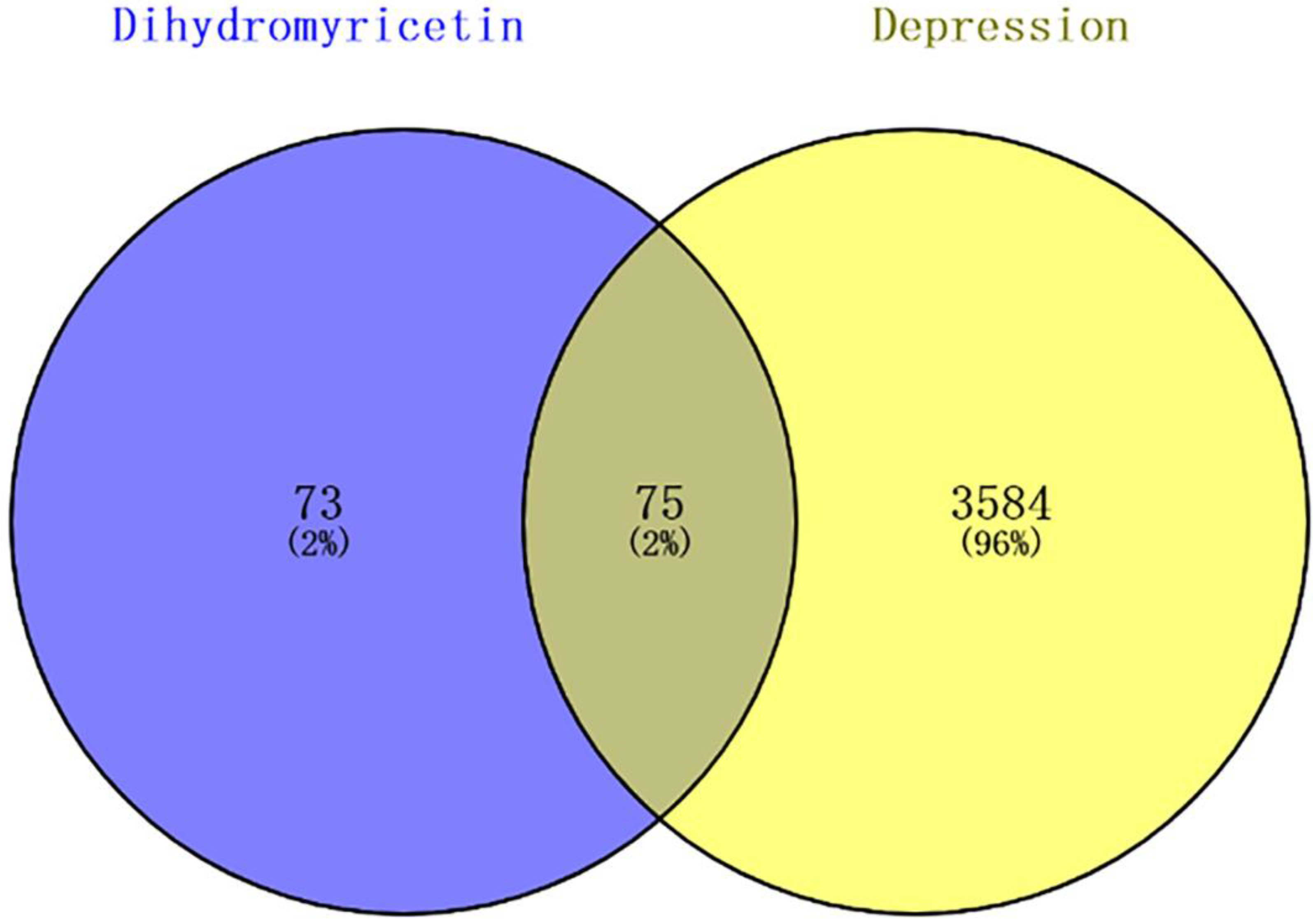
3.2. Target Prediction of DHM in Depression
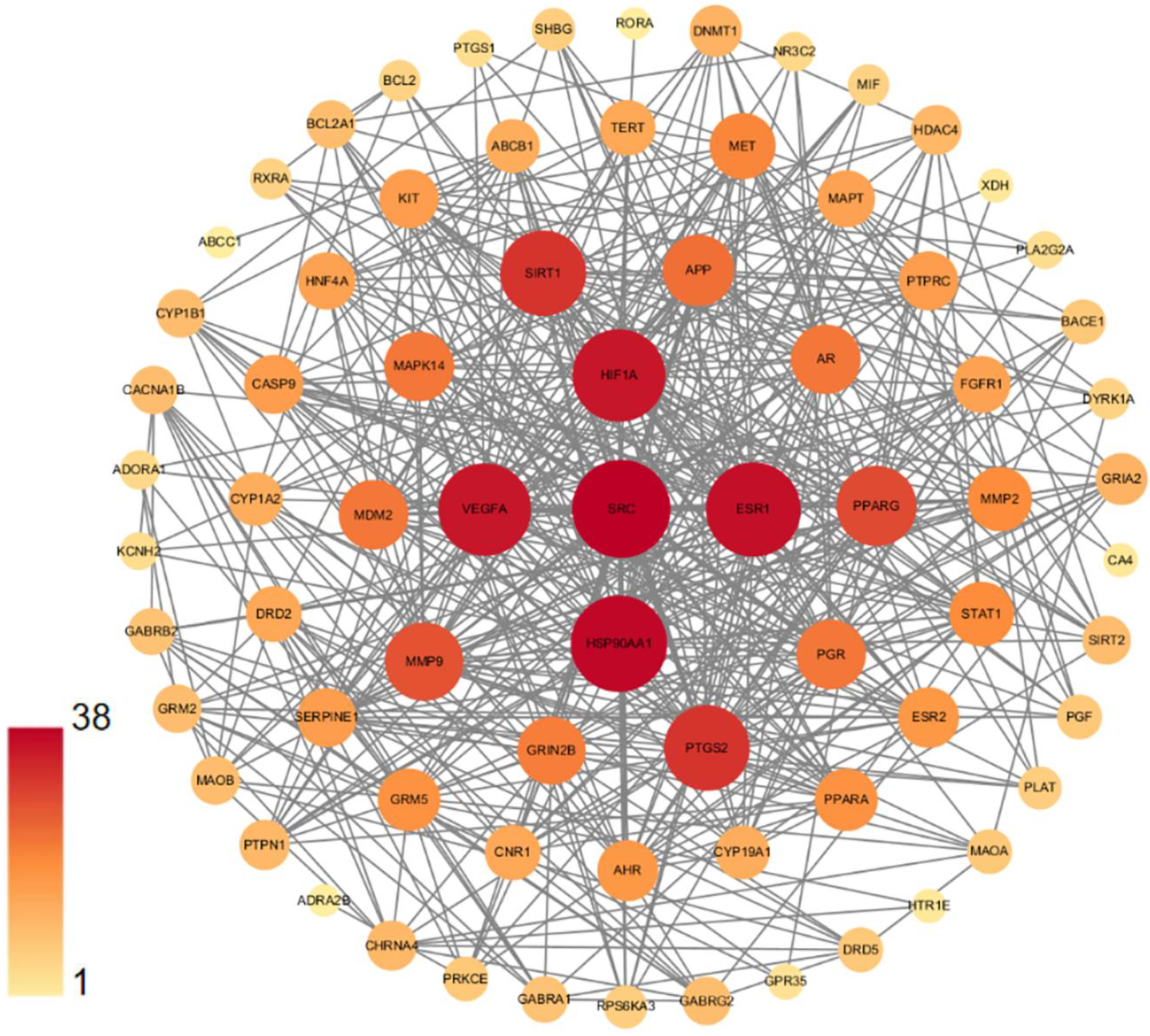
3.3. PPI Network of DHM Targets in Depression
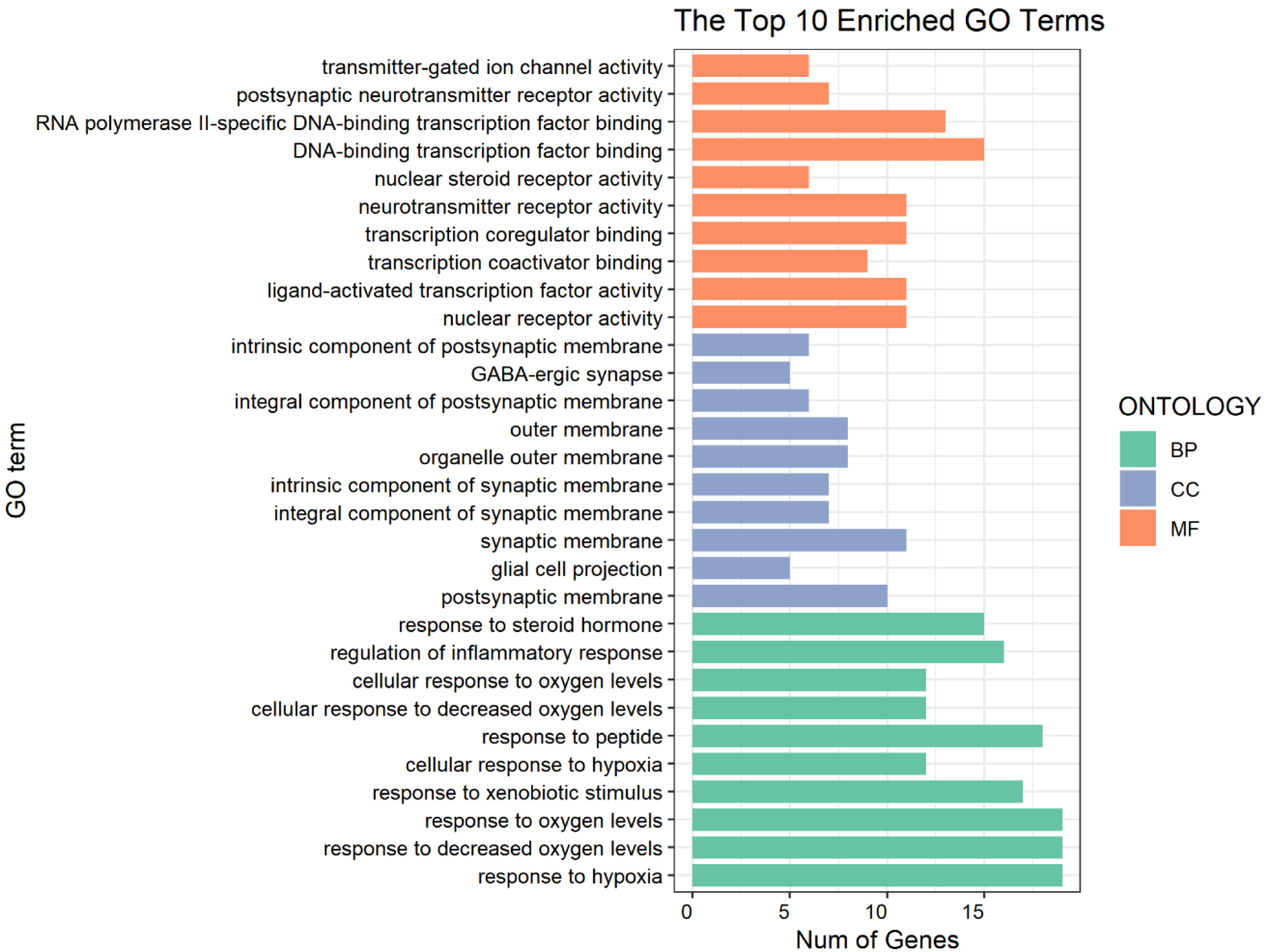
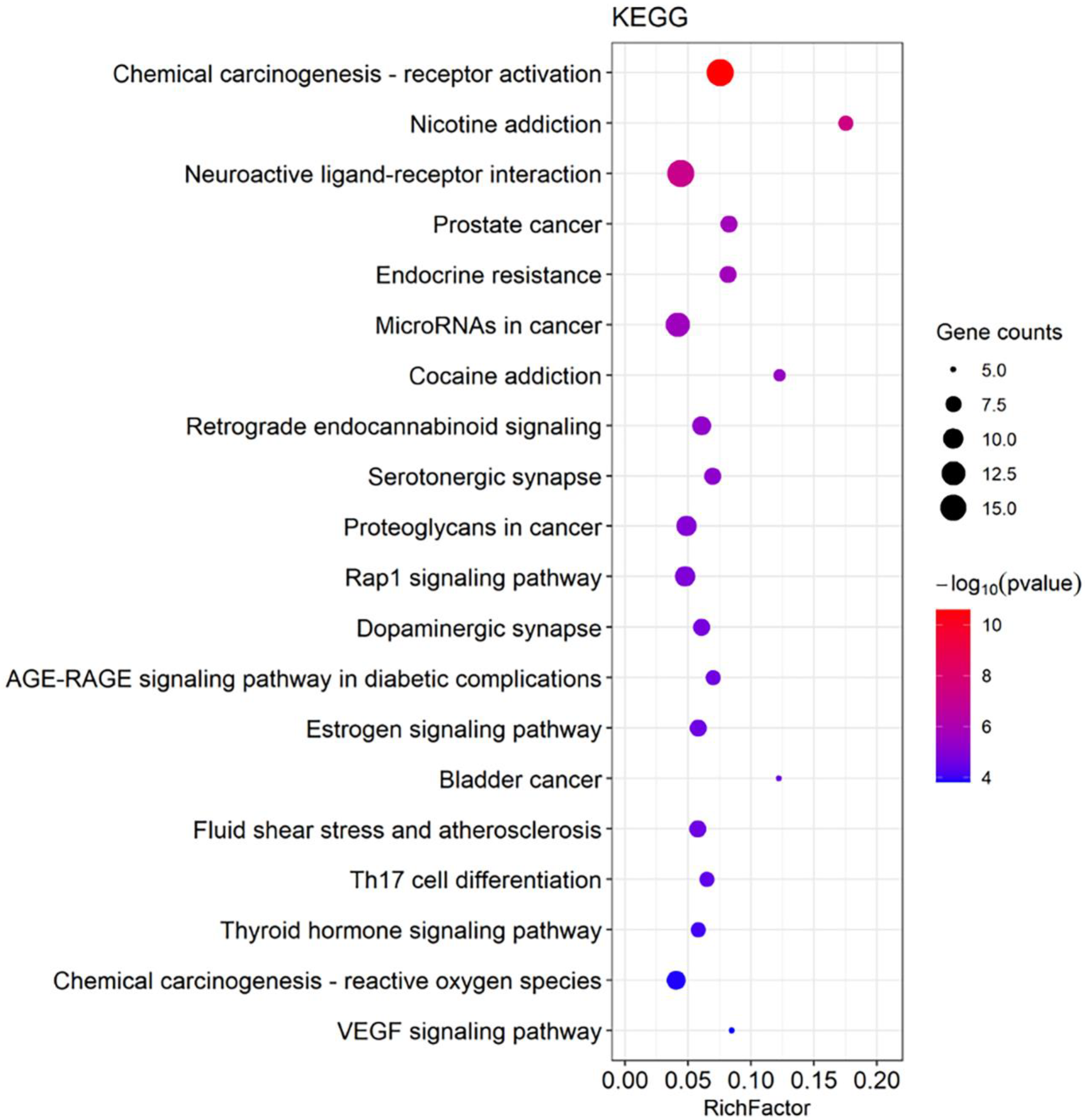
3.4. GO and KEGG Pathway Enrichment Analyses for DHM-Targeted Gene Function and Signaling Pathways
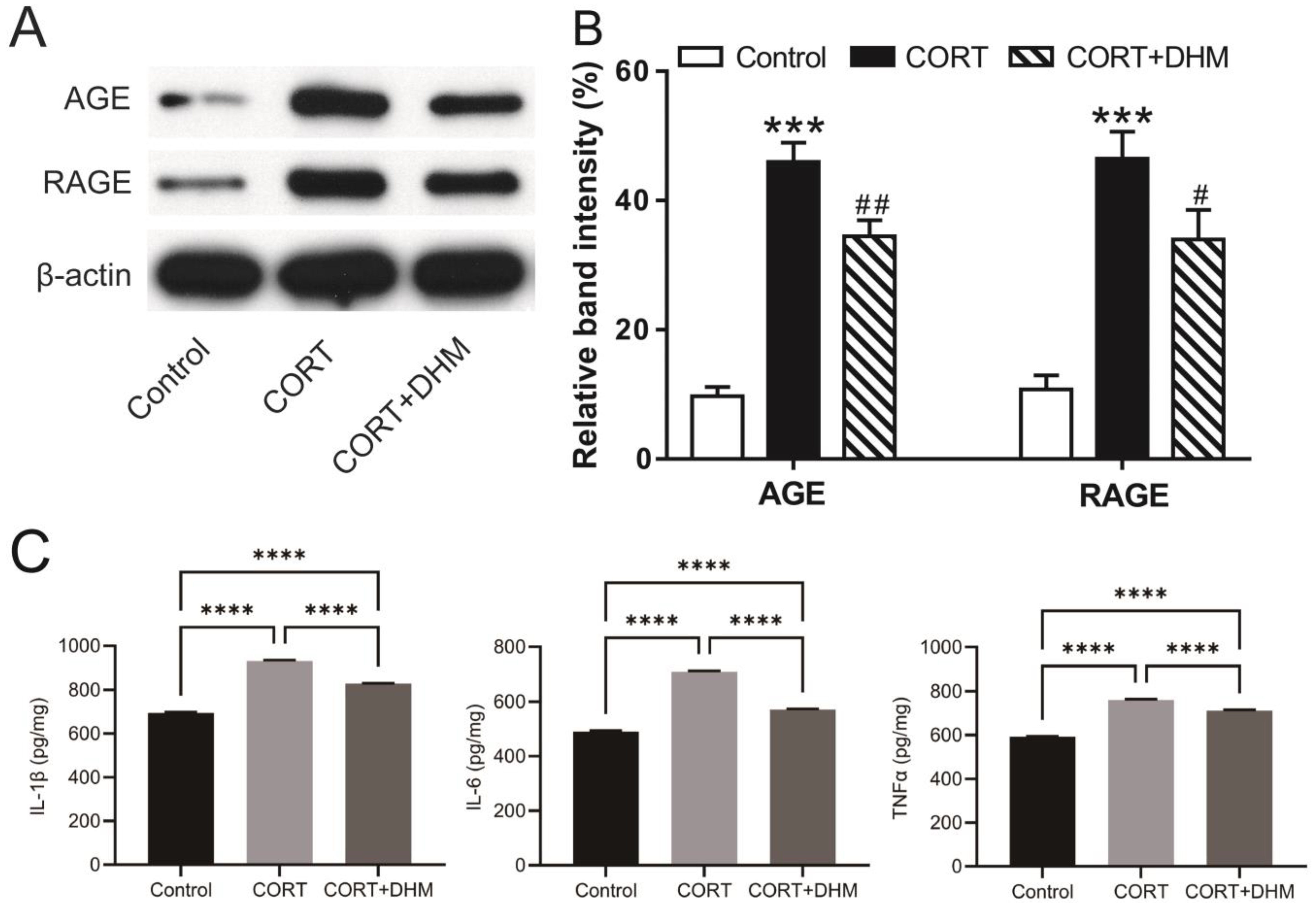
3.5. Inhibition of AGE-RAGE Signaling Mediated Inflammation in CORT-Exposed Mouse Hippocampus
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- WHO. Depression. 2016. Available online: http://apps.who.int/mediacentre/factsheets/fs369/en/index.html (accessed on 15 September 2022).
- Shadrina, M.; Bondarenko, E.A.; Slominsky, P.A. Genetics factors in major depression disease. Front. Psychiatry 2018, 9, 334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levinstein, M.R.; Samuels, B.A. Mechanisms underlying the antidepressant response and treatment resistance. Front. Behav. Neurosci. 2014, 8, 208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Mao, Y.; Ding, L.; Zeng, X.A. Dihydromyricetin: A review on identification and quantification methods, biological activities, chemical stability, metabolism and approaches to enhance its bioavailability. Trends Food Sci. Technol. 2019, 91, 586–597. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, Y.; Luo, H.; Sun, L.; Xu, M.; Yu, J.; Zhou, Q.; Meng, G.; Yang, S. Recent update on the pharmacological effects and mechanisms of dihydromyricetin. Front. Pharmacol. 2018, 9, 1204. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Guo, H.; Li, Z.; Zhang, C.; Zhang, X.; Cui, Q.; Tian, J. Molecular level insight into the benefit of myricetin and dihydromyricetin uptake in patients with Alzheimer’s diseases. Front. Aging Neurosci. 2020, 12, 601603. [Google Scholar] [CrossRef]
- Guan, S.; Shen, Y.; Ge, H.; Xiong, W.; He, L.; Liu, L.; Yin, C.; Wei, X.; Gao, Y. Dihydromyricetin alleviates diabetic neuropathic pain and depression comorbidity symptoms by inhibiting P2 × 7 receptor. Front. Psychiatry 2019, 10, 770. [Google Scholar] [CrossRef]
- Demuyser, T.; Bentea, E.; Deneyer, L.; Albertini, G.; Massie, A.; Smolders, I. Disruption of the HPA-axis through corticosterone-release pellets induces robust depressive-like behavior and reduced BDNF levels in mice. Neurosci. Lett. 2016, 626, 119–125. [Google Scholar] [CrossRef]
- Yu, X.D.; Zhang, D.; Xiao, C.L.; Zhou, Y.; Li, X.; Wang, L.; He, Z.; Reilly, J.; Xiao, Z.-Y.; Shu, X. P-Coumaric Acid Reverses Depression-Like Behavior and Memory Deficit Via Inhibiting AGE-RAGE-Mediated Neuroinflammation. Cells 2022, 11, 1594. [Google Scholar] [CrossRef]
- Taylor, C.; Fricker, A.D.; Devi, L.A.; Gomes, I. Mechanisms of action of antidepressants: From neurotransmitter systems to signaling pathways. Cell. Signal. 2005, 17, 549–557. [Google Scholar] [CrossRef] [Green Version]
- Leonard, B.E. Impact of inflammation on neurotransmitter changes in major depression: An insight into the action of antidepressants. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2014, 48, 261–267. [Google Scholar] [CrossRef]
- Pilc, A.; Nowak, G. GABAergic hypotheses of anxiety and depression: Focus on GABA-B receptors. Drugs Today 2005, 41, 755–766. [Google Scholar] [CrossRef]
- Wang, H.; Kan, W.J.; Feng, Y.; Feng, L.; Yang, Y.; Chen, P.; Xu, J.J.; Si, T.M.; Zhang, L.; Wang, G.; et al. Nuclear receptors modulate inflammasomes in the pathophysiology and treatment of major depressive disorder. World J. Psychiatry 2021, 11, 1191. [Google Scholar] [CrossRef] [PubMed]
- Holsboer, F. The corticosteroid receptor hypothesis of depression. Neuropsychopharmacology 2000, 23, 477–501. [Google Scholar] [CrossRef] [Green Version]
- Sulser, F. The role of CREB and other transcription factors in the pharmacotherapy and etiology of depression. Ann. Med. 2002, 34, 348–356. [Google Scholar] [CrossRef]
- Zucker, R.S. Short-term synaptic plasticity. Annu. Rev. Neurosci. 1989, 12, 13–31. [Google Scholar] [CrossRef]
- Christie, B.R.; Kerr, D.S.; Abraham, W.C. Flip side of synaptic plasticity: Long-term depression mechanisms in the hippocampus. Hippocampus 1994, 4, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Numakawa, T.; Richards, M.; Nakajima, S.; Adachi, N.; Furuta, M.; Odaka, H.; Kunugi, H. The role of brain-derived neurotrophic factor in comorbid depression: Possible linkage with steroid hormones, cytokines, and nutrition. Front. Psychiatry 2014, 5, 136. [Google Scholar] [CrossRef]
- Beurel, E.; Toups, M.; Nemeroff, C.B. The bidirectional relationship of depression and inflammation: Double trouble. Neuron 2020, 107, 234–256. [Google Scholar] [CrossRef]
- Hui, J.J.; Xi, G.J.; Liu, S.S.; Li, X.L.; Geng, L.Y.; Teng, G.J.; Nie, B.B.; Shan, B.C.; Yan, J.; Dong, L.; et al. Blood oxygen level-dependent signals via fMRI in the mood-regulating circuit using two animal models of depression are reversed by chronic escitalopram treatment. Behav. Brain Res. 2016, 311, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Yin, A.Q.; Wang, F.; Zhang, X. Integrating endocannabinoid signaling in the regulation of anxiety and depression. Acta Pharmacol. Sin. 2019, 40, 336–341. [Google Scholar] [CrossRef]
- Perez, J.; Tardito, D.; Racagni, G.; Smeraldi, E.; Zanardi, R. Protein kinase A and Rap1 levels in platelets of untreated patients with major depression. Mol. Psychiatry 2001, 6, 44–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franklin, T.C.; Wohleb, E.S.; Zhang, Y.; Fogaça, M.; Hare, B.; Duman, R.S. Persistent increase in microglial RAGE contributes to chronic stress–induced priming of depressive-like behavior. Biol. Psychiatry 2018, 83, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Mehta, D.; Newport, D.J.; Frishman, G.; Kraus, L.; Rex-Haffner, M.; Ritchie, J.C.; Lori, A.; Knight, B.T.; Stagnaro, E.; Ruepp, A.; et al. Early predictive biomarkers for postpartum depression point to a role for estrogen receptor signaling. Psychol. Med. 2014, 44, 2309–2322. [Google Scholar] [CrossRef]
- Bhatt, S.; Nagappa, A.N.; Patil, C.R. Role of oxidative stress in depression. Drug Discov. Today 2020, 25, 1270–1276. [Google Scholar] [CrossRef]
- De Rossi, P.; Harde, E.; Dupuis, J.P.; Martin, L.; Chounlamountri, N.; Bardin, M.; Watrin, C.; Benetollo, C.; Pernet-Gallay, K.; Luhmann, H.J.; et al. A critical role for VEGF and VEGFR2 in NMDA receptor synaptic function and fear-related behavior. Mol. Psychiatry 2016, 21, 1768–1780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hudson, B.I.; Lippman, M.E. Targeting RAGE signaling in inflammatory disease. Annu. Rev. Med. 2018, 69, 349–364. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Hu, Y.; Qi, K.; Li, Y.; Chen, J.; Wang, R. Dihydromyricetin improves LPS-induced sickness and depressive-like behaviors in mice by inhibiting the TLR4/Akt/HIF1a/NLRP3 pathway. Behav. Brain Res. 2022, 423, 113775. [Google Scholar] [CrossRef]
- Ren, Z.; Yan, P.; Zhu, L.; Yang, H.; Zhao, Y.; Kirby, B.P.; Waddington, J.L.; Zhen, X. Dihydromyricetin exerts a rapid antidepressant-like effect in association with enhancement of BDNF expression and inhibition of neuroinflammation. Psychopharmacology 2018, 235, 233–244. [Google Scholar] [CrossRef]
- Ge, H.; Guan, S.; Shen, Y.; Sun, M.; Hao, Y.; He, L.; Liu, L.; Yin, C.; Huang, R.; Xiong, W.; et al. Dihydromyricetin affects BDNF levels in the nervous system in rats with comorbid diabetic neuropathic pain and depression. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Hopkins, A.L. Network pharmacology: The next paradigm in drug discovery. Nat. Chem. Biol. 2008, 4, 682–690. [Google Scholar] [CrossRef]
- Pan, H.T.; Xi, Z.Q.; Wei, X.Q.; Wang, K. A network pharmacology approach to predict potential targets and mechanisms of “Ramulus Cinnamomi (cassiae)–Paeonia lactiflora” herb pair in the treatment of chronic pain with comorbid anxiety and depression. Ann. Med. 2022, 54, 413–425. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Yang, P.; Zhang, L. Mode of action of Shan-Zhu-Yu (Cornus officinalis Sieb. et Zucc.) in the treatment of depression based on network pharmacology. Evid.-Based Complementary Altern. Med. 2020, 2020, 8838888. [Google Scholar] [CrossRef] [PubMed]
- Lu, G.; Li, J.; Zhang, H.; Zhao, X.; Yan, L.J.; Yang, X. Role and possible mechanisms of Sirt1 in depression. Oxid. Med. Cell. Longev. 2018, 2018, 8596903. [Google Scholar] [CrossRef] [Green Version]
- Bialek, K.; Czarny, P.; Watala, C.; Wigner, P.; Talarowska, M.; Galecki, P.; Szemraj, J.; Sliwinski, T. Novel association between TGFA, TGFB1, IRF1, PTGS2 and IKBKB single-nucleotide polymorphisms and occurrence, severity and treatment response of major depressive disorder. PeerJ 2020, 8, e8676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sundermann, E.E.; Maki, P.M.; Bishop, J.R. A review of estrogen receptor gene (ESR1) polymorphisms, mood, and cognition. Menopause 2010, 17, 874. [Google Scholar] [CrossRef] [Green Version]
- Xiang, X.; You, X.M.; Li, L.Q. Expression of HSP90AA1/HSPA8 in hepatocellular carcinoma patients with depression. Oncotargets Ther. 2018, 11, 3013. [Google Scholar] [CrossRef] [Green Version]
- Ergenc, M.; Ozacmak, H.S.; Turan, I.; Ozacmak, V.H. Melatonin reverses depressive and anxiety like-behaviours induced by diabetes: Involvement of oxidative stress, age, rage and S100B levels in the hippocampus and prefrontal cortex of rats. Arch. Physiol. Biochem. 2022, 128, 402–410. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, S.; Yang, S.; Chen, C.; Yang, Y.; Lin, M.; Liu, C.; Wang, W.; Zhou, X.; Ai, Q.; et al. Mechanism of dihydromyricetin on inflammatory diseases. Front. Pharmacol. 2021, 12, 794563. [Google Scholar] [CrossRef]
- Han, R.; Liu, Z.; Sun, N.; Liu, S.; Li, L.; Shen, Y.; Xiu, J.; Xu, Q. BDNF alleviates neuroinflammation in the Hippocampus of type 1 diabetic mice via blocking the aberrant HMGB1/RAGE/NF-_B pathway. Aging Dis. 2019, 10, 611. [Google Scholar] [CrossRef] [Green Version]
- Orgeta, V.; Tabet, N.; Nilforooshan, R.; Howard, R. 2017. Efficacy of antidepressants for depression in Alzheimer’s disease: Systematic review and meta-analysis. J. Alzheimer’s Dis. 2017, 58, 725–733. [Google Scholar] [CrossRef]
- Marsh, L. Depression and Parkinson’s disease: Current knowledge. Curr. Neurol. Neurosci. Rep. 2013, 13, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martín-Sánchez, A.; Piñero, J.; Nonell, L.; Arnal, M.; Ribe, E.M.; Nevado-Holgado, A.; Lovestone, S.; Sanz, F.; Furlong, L.I.; Valverde, O. Comorbidity between Alzheimer’s disease and major depression: A behavioural and transcriptomic characterization study in mice. Alzheimer’s Res. Ther. 2021, 13, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Ross, J.A.; Gliebus, G.; Van Bockstaele, E.J. 2018. Stress induced neural reorganization: A conceptual framework linking depression and Alzheimer’s disease. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2018, 85, 136–151. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, J.; Chen, B.; Wang, H.; Hu, S.; Yu, X.; Reilly, J.; He, Z.; You, Y.; Shu, X. Dihydromyricetin Attenuates Depressive-like Behaviors in Mice by Inhibiting the AGE-RAGE Signaling Pathway. Cells 2022, 11, 3730. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11233730
Huang J, Chen B, Wang H, Hu S, Yu X, Reilly J, He Z, You Y, Shu X. Dihydromyricetin Attenuates Depressive-like Behaviors in Mice by Inhibiting the AGE-RAGE Signaling Pathway. Cells. 2022; 11(23):3730. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11233730
Chicago/Turabian StyleHuang, Jun, Bin Chen, Hao Wang, Sheng Hu, Xudong Yu, James Reilly, Zhiming He, Yong You, and Xinhua Shu. 2022. "Dihydromyricetin Attenuates Depressive-like Behaviors in Mice by Inhibiting the AGE-RAGE Signaling Pathway" Cells 11, no. 23: 3730. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11233730