
Prevention of L-Dopa-Induced Dyskinesias by MPEP Blockade of Metabotropic Glutamate Receptor 5 Is Associated with Reduced Inflammation in the Brain of Parkinsonian Monkeys

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Treatments
2.2. Motor Behavior Measures
2.3. Striatal DA and Serotonin (5-HT) Contents
2.4. Tissue Preparation and Western Blot
2.5. Statistical Analysis
3. Results
3.1. MPEP Inhibited the Development of LID in MPTP Monkeys
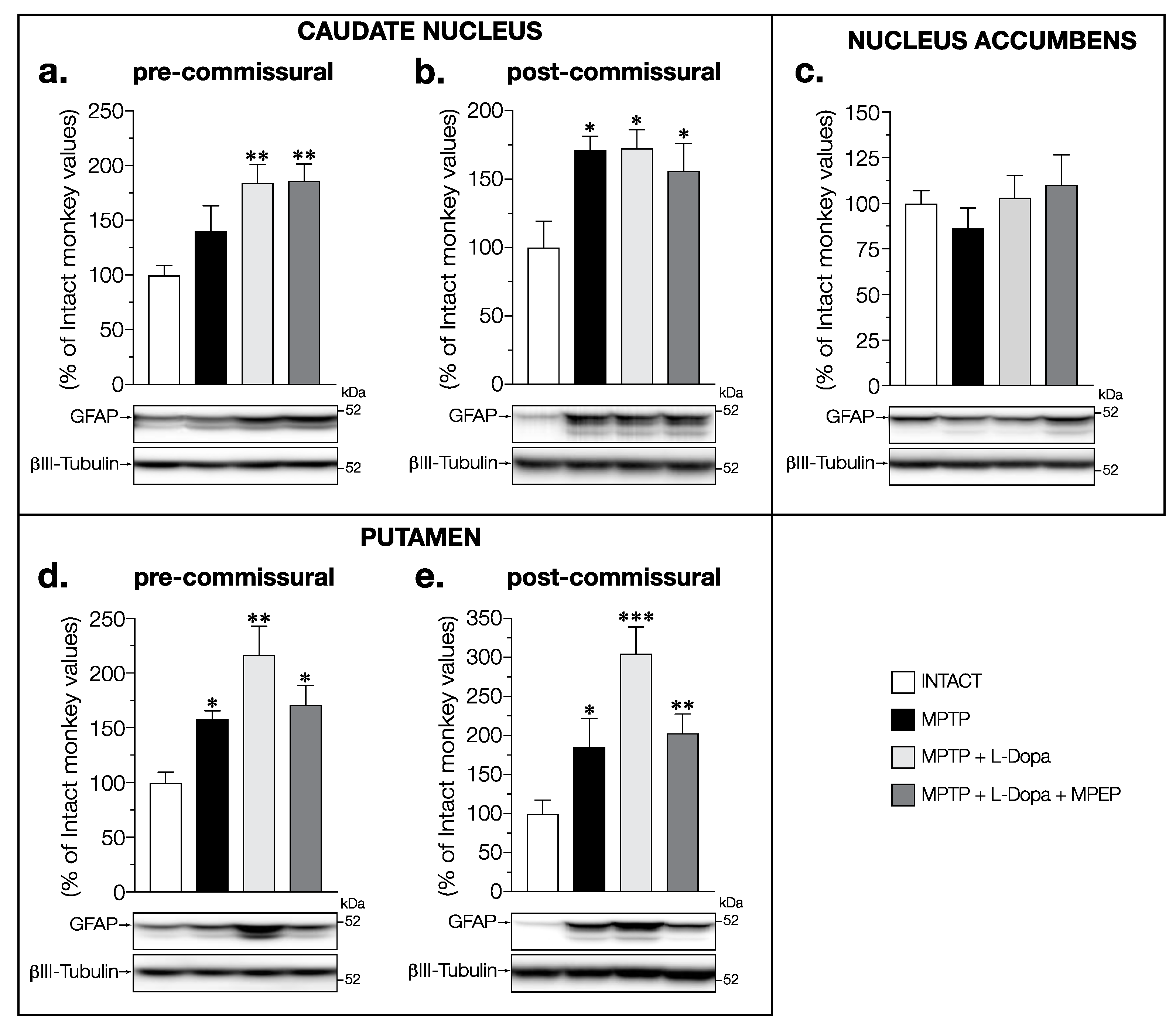
3.2. MPEP Attenuated the Increase in GFAP Levels Induced by L-Dopa in the Putamen and STN of MPTP-Lesioned Monkeys
3.3. MPEP Attenuated the Increase in Iba1 Levels Induced by L-Dopa in the Putamen, Substantia Nigra, and Globus Pallidus of MPTP-Lesioned Monkeys
3.4. MPEP Attenuated the Increase in CD68 Levels Induced by L-Dopa in the Putamen and Globus Pallidus of MPTP-Lesioned Monkeys
4. Discussion
4.1. Inflammation in PD and LID
4.2. The Astrocytes Marker GFAP Levels in the Brain of PD and of Animal Models of PD
4.3. The Astrocytes Marker GFAP Levels in the Brain of MPTP Monkeys, LID, and MPEP
4.4. Brain Microglia-Mediated Inflammation in PD and MPTP Monkeys
4.5. Brain Microglia-Mediated Inflammation: Positive Correlation with LID
4.6. MPEP Reduced Dyskinesias and Microglial Inflammation Markers in MPTP Monkeys
4.7. General Considerations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Blauwendraat, C.; Heilbron, K.; Vallerga, C.L.; Bandres-Ciga, S.; von Coelln, R.; Pihlstrom, L.; Simon-Sanchez, J.; Schulte, C.; Sharma, M.; Krohn, L.; et al. Parkinson’s disease age at onset genome-wide association study: Defining heritability, genetic loci, and alpha-synuclein mechanisms. Mov. Disord. 2019, 34, 866–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Draoui, A.; El Hiba, O.; Aimrane, A.; El Khiat, A.; Gamrani, H. Parkinson’s disease: From bench to bedside. Rev. Neurol. 2020, 176, 543–559. [Google Scholar] [CrossRef] [PubMed]
- Pang, S.Y.; Ho, P.W.; Liu, H.F.; Leung, C.T.; Li, L.; Chang, E.E.S.; Ramsden, D.B.; Ho, S.L. The interplay of aging, genetics and environmental factors in the pathogenesis of Parkinson’s disease. Transl. Neurodegener. 2019, 8, 23. [Google Scholar] [CrossRef] [PubMed]
- Olanow, C.W.; Stocchi, F. Levodopa: A new look at an old friend. Mov. Disord. 2018, 33, 859–866. [Google Scholar] [CrossRef] [PubMed]
- Bastide, M.F.; Meissner, W.G.; Picconi, B.; Fasano, S.; Fernagut, P.O.; Feyder, M.; Francardo, V.; Alcacer, C.; Ding, Y.; Brambilla, R.; et al. Pathophysiology of L-dopa-induced motor and non-motor complications in Parkinson’s disease. Prog. Neurobiol. 2015, 132, 96–168. [Google Scholar] [CrossRef]
- Guerra, A.; Suppa, A.; D’Onofrio, V.; Di Stasio, F.; Asci, F.; Fabbrini, G.; Berardelli, A. Abnormal cortical facilitation and L-dopa-induced dyskinesia in Parkinson’s disease. Brain Stimul. 2019, 12, 1517–1525. [Google Scholar] [CrossRef]
- Picconi, B.; Hernandez, L.F.; Obeso, J.A.; Calabresi, P. Motor complications in Parkinson’s disease: Striatal molecular and electrophysiological mechanisms of dyskinesias. Mov. Disord. 2018, 33, 867–876. [Google Scholar] [CrossRef]
- Mellone, M.; Zianni, E.; Stanic, J.; Campanelli, F.; Marino, G.; Ghiglieri, V.; Longhi, A.; Thiolat, M.L.; Li, Q.; Calabresi, P.; et al. NMDA receptor GluN2D subunit participates to levodopa-induced dyskinesia pathophysiology. Neurobiol. Dis. 2019, 121, 338–349. [Google Scholar] [CrossRef]
- Paoletti, P. Molecular basis of NMDA receptor functional diversity. Eur. J. Neurosci. 2011, 33, 1351–1365. [Google Scholar] [CrossRef]
- Fox, S.H.; Katzenschlager, R.; Lim, S.Y.; Ravina, B.; Seppi, K.; Coelho, M.; Poewe, W.; Rascol, O.; Goetz, C.G.; Sampaio, C. The Movement Disorder Society Evidence-Based Medicine Review Update: Treatments for the motor symptoms of Parkinson’s disease. Mov. Disord. 2011, 26 (Suppl. S3), S2–S41. [Google Scholar] [CrossRef]
- Rascol, O.; Fabbri, M.; Poewe, W. Amantadine in the treatment of Parkinson’s disease and other movement disorders. Lancet Neurol. 2021, 20, 1048–1056. [Google Scholar] [CrossRef]
- Masilamoni, G.J.; Smith, Y. Metabotropic glutamate receptors: Targets for neuroprotective therapies in Parkinson disease. Curr. Opin. Pharmacol. 2018, 38, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Awad, H.; Hubert, G.W.; Smith, Y.; Levey, A.I.; Conn, P.J. Activation of metabotropic glutamate receptor 5 has direct excitatory effects and potentiates NMDA receptor currents in neurons of the subthalamic nucleus. J. Neurosci. 2000, 20, 7871–7879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conn, P.J.; Battaglia, G.; Marino, M.J.; Nicoletti, F. Metabotropic glutamate receptors in the basal ganglia motor circuit. Nat. Rev. Neurosci. 2005, 6, 787–798. [Google Scholar] [CrossRef] [PubMed]
- Jin, D.Z.; Xue, B.; Mao, L.M.; Wang, J.Q. Metabotropic glutamate receptor 5 upregulates surface NMDA receptor expression in striatal neurons via CaMKII. Brain Res. 2015, 1624, 414–423. [Google Scholar] [CrossRef] [Green Version]
- Voulalas, P.J.; Holtzclaw, L.; Wolstenholme, J.; Russell, J.T.; Hyman, S.E. Metabotropic glutamate receptors and dopamine receptors cooperate to enhance extracellular signal-regulated kinase phosphorylation in striatal neurons. J. Neurosci. 2005, 25, 3763–3773. [Google Scholar] [CrossRef] [Green Version]
- Morin, N.; Morissette, M.; Gregoire, L.; Gomez-Mancilla, B.; Gasparini, F.; Di Paolo, T. Chronic treatment with MPEP, an mGlu5 receptor antagonist, normalizes basal ganglia glutamate neurotransmission in L-DOPA-treated parkinsonian monkeys. Neuropharmacology 2013, 73, 216–231. [Google Scholar] [CrossRef]
- Ouattara, B.; Gasparini, F.; Morissette, M.; Gregoire, L.; Samadi, P.; Gomez-Mancilla, B.; Di Paolo, T. Effect of L-Dopa on metabotropic glutamate receptor 5 in the brain of parkinsonian monkeys. J. Neurochem. 2010, 113, 715–724. [Google Scholar] [CrossRef]
- Ouattara, B.; Gregoire, L.; Morissette, M.; Gasparini, F.; Vranesic, I.; Bilbe, G.; Johns, D.R.; Rajput, A.; Hornykiewicz, O.; Rajput, A.H.; et al. Metabotropic glutamate receptor type 5 in levodopa-induced motor complications. Neurobiol. Aging 2011, 32, 1286–1295. [Google Scholar] [CrossRef]
- Sanchez-Pernaute, R.; Wang, J.Q.; Kuruppu, D.; Cao, L.; Tueckmantel, W.; Kozikowski, A.; Isacson, O.; Brownell, A.L. Enhanced binding of metabotropic glutamate receptor type 5 (mGluR5) PET tracers in the brain of parkinsonian primates. Neuroimage 2008, 42, 248–251. [Google Scholar] [CrossRef] [Green Version]
- Litim, N.; Morissette, M.; Di Paolo, T. Metabotropic glutamate receptors as therapeutic targets in Parkinson’s disease: An update from the last 5 years of research. Neuropharmacology 2017, 115, 166–179. [Google Scholar] [CrossRef]
- Morin, N.; Gregoire, L.; Gomez-Mancilla, B.; Gasparini, F.; Di Paolo, T. Effect of the metabotropic glutamate receptor type 5 antagonists MPEP and MTEP in parkinsonian monkeys. Neuropharmacology 2010, 58, 981–986. [Google Scholar] [CrossRef] [PubMed]
- Morin, N.; Gregoire, L.; Morissette, M.; Desrayaud, S.; Gomez-Mancilla, B.; Gasparini, F.; Di Paolo, T. MPEP, an mGlu5 receptor antagonist, reduces the development of L-DOPA-induced motor complications in de novo parkinsonian monkeys: Biochemical correlates. Neuropharmacology 2013, 66, 355–364. [Google Scholar] [CrossRef]
- Morin, N.; Jourdain, V.A.; Morissette, M.; Gregoire, L.; Di Paolo, T. Long-term treatment with l-DOPA and an mGlu5 receptor antagonist prevents changes in brain basal ganglia dopamine receptors, their associated signaling proteins and neuropeptides in parkinsonian monkeys. Neuropharmacology 2014, 79, 688–706. [Google Scholar] [CrossRef] [PubMed]
- Morissette, M.; Morin, N.; Gregoire, L.; Rajput, A.; Rajput, A.H.; Di Paolo, T. Brain alpha7 nicotinic acetylcholine receptors in MPTP-lesioned monkeys and parkinsonian patients. Biochem. Pharmacol. 2016, 109, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Armentero, M.T.; Fancellu, R.; Nappi, G.; Bramanti, P.; Blandini, F. Prolonged blockade of NMDA or mGluR5 glutamate receptors reduces nigrostriatal degeneration while inducing selective metabolic changes in the basal ganglia circuitry in a rodent model of Parkinson’s disease. Neurobiol. Dis. 2006, 22, 1–9. [Google Scholar] [CrossRef]
- Battaglia, G.; Busceti, C.L.; Molinaro, G.; Biagioni, F.; Storto, M.; Fornai, F.; Nicoletti, F.; Bruno, V. Endogenous activation of mGlu5 metabotropic glutamate receptors contributes to the development of nigro-striatal damage induced by 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine in mice. J. Neurosci. 2004, 24, 828–835. [Google Scholar] [CrossRef]
- Masilamoni, G.J.; Bogenpohl, J.W.; Alagille, D.; Delevich, K.; Tamagnan, G.; Votaw, J.R.; Wichmann, T.; Smith, Y. Metabotropic glutamate receptor 5 antagonist protects dopaminergic and noradrenergic neurons from degeneration in MPTP-treated monkeys. Brain 2011, 134, 2057–2073. [Google Scholar] [CrossRef] [Green Version]
- Vernon, A.C.; Zbarsky, V.; Datla, K.P.; Croucher, M.J.; Dexter, D.T. Subtype selective antagonism of substantia nigra pars compacta Group I metabotropic glutamate receptors protects the nigrostriatal system against 6-hydroxydopamine toxicity in vivo. J. Neurochem. 2007, 103, 1075–1091. [Google Scholar] [CrossRef]
- Biber, K.; Laurie, D.J.; Berthele, A.; Sommer, B.; Tolle, T.R.; Gebicke-Harter, P.J.; van Calker, D.; Boddeke, H.W. Expression and signaling of group I metabotropic glutamate receptors in astrocytes and microglia. J. Neurochem. 1999, 72, 1671–1680. [Google Scholar] [CrossRef]
- Byrnes, K.R.; Stoica, B.; Loane, D.J.; Riccio, A.; Davis, M.I.; Faden, A.I. Metabotropic glutamate receptor 5 activation inhibits microglial associated inflammation and neurotoxicity. Glia 2009, 57, 550–560. [Google Scholar] [CrossRef] [Green Version]
- Spampinato, S.F.; Copani, A.; Nicoletti, F.; Sortino, M.A.; Caraci, F. Metabotropic Glutamate Receptors in Glial Cells: A New Potential Target for Neuroprotection? Front. Mol. Neurosci. 2018, 11, 414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Chen, K.; Sloan, S.A.; Bennett, M.L.; Scholze, A.R.; O’Keeffe, S.; Phatnani, H.P.; Guarnieri, P.; Caneda, C.; Ruderisch, N.; et al. An RNA-sequencing transcriptome and splicing database of glia, neurons, and vascular cells of the cerebral cortex. J. Neurosci. 2014, 34, 11929–11947. [Google Scholar] [CrossRef]
- Aronica, E.; van Vliet, E.A.; Mayboroda, O.A.; Troost, D.; da Silva, F.H.; Gorter, J.A. Upregulation of metabotropic glutamate receptor subtype mGluR3 and mGluR5 in reactive astrocytes in a rat model of mesial temporal lobe epilepsy. Eur. J. Neurosci. 2000, 12, 2333–2344. [Google Scholar] [CrossRef]
- Ferraguti, F.; Corti, C.; Valerio, E.; Mion, S.; Xuereb, J. Activated astrocytes in areas of kainate-induced neuronal injury upregulate the expression of the metabotropic glutamate receptors 2/3 and 5. Exp. Brain Res. 2001, 137, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Loane, D.J.; Stoica, B.A.; Byrnes, K.R.; Jeong, W.; Faden, A.I. Activation of mGluR5 and inhibition of NADPH oxidase improves functional recovery after traumatic brain injury. J. Neurotrauma 2013, 30, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Loane, D.J.; Stoica, B.A.; Pajoohesh-Ganji, A.; Byrnes, K.R.; Faden, A.I. Activation of metabotropic glutamate receptor 5 modulates microglial reactivity and neurotoxicity by inhibiting NADPH oxidase. J. Biol. Chem. 2009, 284, 15629–15639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loane, D.J.; Stoica, B.A.; Tchantchou, F.; Kumar, A.; Barrett, J.P.; Akintola, T.; Xue, F.; Conn, P.J.; Faden, A.I. Novel mGluR5 positive allosteric modulator improves functional recovery, attenuates neurodegeneration, and alters microglial polarization after experimental traumatic brain injury. Neurotherapeutics 2014, 11, 857–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, F.; Stoica, B.A.; Hanscom, M.; Kabadi, S.V.; Faden, A.I. Positive allosteric modulators (PAMs) of metabotropic glutamate receptor 5 (mGluR5) attenuate microglial activation. CNS Neurol. Disord. Drug Targets 2014, 13, 558–566. [Google Scholar] [CrossRef]
- Ye, X.; Yu, L.; Zuo, D.; Zhang, L.; Zu, J.; Hu, J.; Tang, J.; Bao, L.; Cui, C.; Zhang, R.; et al. Activated mGluR5 protects BV2 cells against OGD/R induced cytotoxicity by modulating BDNF-TrkB pathway. Neurosci. Lett. 2017, 654, 70–79. [Google Scholar] [CrossRef]
- Piers, T.M.; Heales, S.J.; Pocock, J.M. Positive allosteric modulation of metabotropic glutamate receptor 5 down-regulates fibrinogen-activated microglia providing neuronal protection. Neurosci. Lett. 2011, 505, 140–145. [Google Scholar] [CrossRef]
- Qiu, J.L.; Zhu, W.L.; Lu, Y.J.; Bai, Z.F.; Liu, Z.G.; Zhao, P.; Sun, C.; Zhang, Y.B.; Li, H.; Liu, W. The selective mGluR5 agonist CHPG attenuates SO2-induced oxidative stress and inflammation through TSG-6/NF-kappaB pathway in BV2 microglial cells. Neurochem. Int. 2015, 85–86, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Chantong, B.; Kratschmar, D.V.; Lister, A.; Odermatt, A. Inhibition of metabotropic glutamate receptor 5 induces cellular stress through pertussis toxin-sensitive Gi-proteins in murine BV-2 microglia cells. J. Neuroinflamm. 2014, 11, 190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, M.H.; Ho, S.C.; Yeh, K.Y.; Pawlak, C.R.; Chang, H.M.; Ho, Y.J.; Lai, T.J.; Wu, F.Y. Blockade of metabotropic glutamate receptors inhibits cognition and neurodegeneration in an MPTP-induced Parkinson’s disease rat model. Pharmacol. Biochem. Behav. 2012, 102, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Pajares, M.; I Rojo, A.; Manda, G.; Bosca, L.; Cuadrado, A. Inflammation in Parkinson’s Disease: Mechanisms and Therapeutic Implications. Cells 2020, 9, 1687. [Google Scholar] [CrossRef]
- Sood, A.; Preeti, K.; Fernandes, V.; Khatri, D.K.; Singh, S.B. Glia: A major player in glutamate-GABA dysregulation-mediated neurodegeneration. J. Neurosci. Res. 2021, 99, 3148–3189. [Google Scholar] [CrossRef]
- Pisanu, A.; Boi, L.; Mulas, G.; Spiga, S.; Fenu, S.; Carta, A.R. Neuroinflammation in L-DOPA-induced dyskinesia: Beyond the immune function. J. Neural Transm. 2018, 125, 1287–1297. [Google Scholar] [CrossRef]
- Hurley, M.J.; Jackson, M.J.; Smith, L.A.; Rose, S.; Jenner, P. Proteomic analysis of striatum from MPTP-treated marmosets (Callithrix jacchus) with L-DOPA-induced dyskinesia of differing severity. J. Mol. Neurosci. 2014, 52, 302–312. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Q.J.; Wang, H.S.; Wang, T.; Liu, J. Genome-wide microarray analysis identifies a potential role for striatal retrograde endocannabinoid signaling in the pathogenesis of experimental L-DOPA-induced dyskinesia. Synapse 2014, 68, 332–343. [Google Scholar] [CrossRef]
- Doorn, K.J.; Moors, T.; Drukarch, B.; van de Berg, W.; Lucassen, P.J.; van Dam, A.M. Microglial phenotypes and toll-like receptor 2 in the substantia nigra and hippocampus of incidental Lewy body disease cases and Parkinson’s disease patients. Acta Neuropathol. Commun. 2014, 2, 90. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Guajardo, V.; Febbraro, F.; Kirik, D.; Romero-Ramos, M. Microglia acquire distinct activation profiles depending on the degree of alpha-synuclein neuropathology in a rAAV based model of Parkinson’s disease. PLoS ONE 2010, 5, e8784. [Google Scholar] [CrossRef] [PubMed]
- Theodore, S.; Cao, S.; McLean, P.J.; Standaert, D.G. Targeted overexpression of human alpha-synuclein triggers microglial activation and an adaptive immune response in a mouse model of Parkinson disease. J. Neuropathol. Exp. Neurol. 2008, 67, 1149–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, R.F.; Bowden, D.M. Primate Brain Maps: Structure of the Macaque Brain; Elsevier Science: Amsterdam, NL, USA, 2000. [Google Scholar]
- Belic, J.J.; Halje, P.; Richter, U.; Petersson, P.; Hellgren Kotaleski, J. Untangling Cortico-Striatal Connectivity and Cross-Frequency Coupling in L-DOPA-Induced Dyskinesia. Front. Syst. Neurosci. 2016, 10, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.Z.; Rothwell, J.C.; Lu, C.S.; Chuang, W.L.; Chen, R.S. Abnormal bidirectional plasticity-like effects in Parkinson’s disease. Brain 2011, 134, 2312–2320. [Google Scholar] [CrossRef] [Green Version]
- Kuter, K.; Olech, L.; Glowacka, U.; Paleczna, M. Astrocyte support is important for the compensatory potential of the nigrostriatal system neurons during early neurodegeneration. J. Neurochem. 2019, 148, 63–79. [Google Scholar] [CrossRef] [Green Version]
- Emmer, K.L.; Waxman, E.A.; Covy, J.P.; Giasson, B.I. E46K human alpha-synuclein transgenic mice develop Lewy-like and tau pathology associated with age-dependent, detrimental motor impairment. J. Biol. Chem. 2011, 286, 35104–35118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Isla, T.; Irizarry, M.C.; Mariash, A.; Cheung, B.; Soto, O.; Schrump, S.; Sondel, J.; Kotilinek, L.; Day, J.; Schwarzschild, M.A.; et al. Motor dysfunction and gliosis with preserved dopaminergic markers in human alpha-synuclein A30P transgenic mice. Neurobiol. Aging 2003, 24, 245–258. [Google Scholar] [CrossRef]
- Kuter, K.; Olech, L.; Glowacka, U. Prolonged Dysfunction of Astrocytes and Activation of Microglia Accelerate Degeneration of Dopaminergic Neurons in the Rat Substantia Nigra and Block Compensation of Early Motor Dysfunction Induced by 6-OHDA. Mol. Neurobiol. 2018, 55, 3049–3066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.K.; Stirling, W.; Xu, Y.; Xu, X.; Qui, D.; Mandir, A.S.; Dawson, T.M.; Copeland, N.G.; Jenkins, N.A.; Price, D.L. Human alpha-synuclein-harboring familial Parkinson’s disease-linked Ala-53 -> Thr mutation causes neurodegenerative disease with alpha-synuclein aggregation in transgenic mice. Proc. Natl. Acad. Sci. USA 2002, 99, 8968–8973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, R.M.; Kiser, G.L.; Kaysser-Kranich, T.; Casaceli, C.; Colla, E.; Lee, M.K.; Palaniappan, C.; Federoff, H.J. Wild-type and mutant alpha-synuclein induce a multi-component gene expression profile consistent with shared pathophysiology in different transgenic mouse models of PD. Exp. Neurol. 2007, 204, 421–432. [Google Scholar] [CrossRef]
- Pisanu, A.; Lecca, D.; Mulas, G.; Wardas, J.; Simbula, G.; Spiga, S.; Carta, A.R. Dynamic changes in pro- and anti-inflammatory cytokines in microglia after PPAR-gamma agonist neuroprotective treatment in the MPTPp mouse model of progressive Parkinson’s disease. Neurobiol. Dis. 2014, 71, 280–291. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Federoff, H.J.; Maguire-Zeiss, K.A. Mutant alpha-synuclein overexpression mediates early proinflammatory activity. Neurotox. Res. 2009, 16, 238–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson, M.B.; Richter, F.; Lee, S.K.; Gabby, L.; Wu, J.; Masliah, E.; Effros, R.B.; Chesselet, M.F. Regionally-specific microglial activation in young mice over-expressing human wildtype alpha-synuclein. Exp. Neurol. 2012, 237, 318–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernaus, A.; Blanco, S.; Sevilla, A. Glia Crosstalk in Neuroinflammatory Diseases. Front. Cell. Neurosci. 2020, 14, 209. [Google Scholar] [CrossRef]
- Liddelow, S.A.; Guttenplan, K.A.; Clarke, L.E.; Bennett, F.C.; Bohlen, C.J.; Schirmer, L.; Bennett, M.L.; Munch, A.E.; Chung, W.S.; Peterson, T.C.; et al. Neurotoxic reactive astrocytes are induced by activated microglia. Nature 2017, 541, 481–487. [Google Scholar] [CrossRef]
- Joshi, A.U.; Minhas, P.S.; Liddelow, S.A.; Haileselassie, B.; Andreasson, K.I.; Dorn, G.W., 2nd; Mochly-Rosen, D. Fragmented mitochondria released from microglia trigger A1 astrocytic response and propagate inflammatory neurodegeneration. Nat. Neurosci. 2019, 22, 1635–1648. [Google Scholar] [CrossRef]
- L’Episcopo, F.; Tirolo, C.; Serapide, M.F.; Caniglia, S.; Testa, N.; Leggio, L.; Vivarelli, S.; Iraci, N.; Pluchino, S.; Marchetti, B. Microglia Polarization, Gene-Environment Interactions and Wnt/beta-Catenin Signaling: Emerging Roles of Glia-Neuron and Glia-Stem/Neuroprogenitor Crosstalk for Dopaminergic Neurorestoration in Aged Parkinsonian Brain. Front. Aging Neurosci. 2018, 10, 12. [Google Scholar] [CrossRef] [Green Version]
- Escartin, C.; Galea, E.; Lakatos, A.; O’Callaghan, J.P.; Petzold, G.C.; Serrano-Pozo, A.; Steinhauser, C.; Volterra, A.; Carmignoto, G.; Agarwal, A.; et al. Reactive astrocyte nomenclature, definitions, and future directions. Nat. Neurosci. 2021, 24, 312–325. [Google Scholar] [CrossRef]
- Marchetti, B.; L’Episcopo, F.; Morale, M.C.; Tirolo, C.; Testa, N.; Caniglia, S.; Serapide, M.F.; Pluchino, S. Uncovering novel actors in astrocyte-neuron crosstalk in Parkinson’s disease: The Wnt/beta-catenin signaling cascade as the common final pathway for neuroprotection and self-repair. Eur. J. Neurosci. 2013, 37, 1550–1563. [Google Scholar] [CrossRef] [Green Version]
- Bortolanza, M.; Cavalcanti-Kiwiatkoski, R.; Padovan-Neto, F.E.; da-Silva, C.A.; Mitkovski, M.; Raisman-Vozari, R.; Del-Bel, E. Glial activation is associated with l-DOPA induced dyskinesia and blocked by a nitric oxide synthase inhibitor in a rat model of Parkinson’s disease. Neurobiol. Dis. 2015, 73, 377–387. [Google Scholar] [CrossRef]
- Mulas, G.; Espa, E.; Fenu, S.; Spiga, S.; Cossu, G.; Pillai, E.; Carboni, E.; Simbula, G.; Jadzic, D.; Angius, F.; et al. Differential induction of dyskinesia and neuroinflammation by pulsatile versus continuous l-DOPA delivery in the 6-OHDA model of Parkinson’s disease. Exp. Neurol. 2016, 286, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Garcia, G.; Palafox-Sanchez, V.; Limon, I.D. Nitrosative and cognitive effects of chronic L-DOPA administration in rats with intra-nigral 6-OHDA lesion. Neuroscience 2015, 290, 492–508. [Google Scholar] [CrossRef] [PubMed]
- Padovan-Neto, F.E.; Cavalcanti-Kiwiatkoviski, R.; Carolino, R.O.; Anselmo-Franci, J.; Del Bel, E. Effects of prolonged neuronal nitric oxide synthase inhibition on the development and expression of L-DOPA-induced dyskinesia in 6-OHDA-lesioned rats. Neuropharmacology 2015, 89, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Padovan-Neto, F.E.; Echeverry, M.B.; Tumas, V.; Del-Bel, E.A. Nitric oxide synthase inhibition attenuates L-DOPA-induced dyskinesias in a rodent model of Parkinson’s disease. Neuroscience 2009, 159, 927–935. [Google Scholar] [CrossRef] [PubMed]
- Vermeiren, C.; Najimi, M.; Vanhoutte, N.; Tilleux, S.; de Hemptinne, I.; Maloteaux, J.M.; Hermans, E. Acute up-regulation of glutamate uptake mediated by mGluR5a in reactive astrocytes. J. Neurochem. 2005, 94, 405–416. [Google Scholar] [CrossRef] [PubMed]
- Aronica, E.; Gorter, J.A.; Ijlst-Keizers, H.; Rozemuller, A.J.; Yankaya, B.; Leenstra, S.; Troost, D. Expression and functional role of mGluR3 and mGluR5 in human astrocytes and glioma cells: Opposite regulation of glutamate transporter proteins. Eur. J. Neurosci. 2003, 17, 2106–2118. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.; Silverstein, P.S.; Singh, D.P.; Kumar, A. Involvement of metabotropic glutamate receptor 5, AKT/PI3K signaling and NF-kappaB pathway in methamphetamine-mediated increase in IL-6 and IL-8 expression in astrocytes. J. Neuroinflamm. 2012, 9, 52. [Google Scholar] [CrossRef] [Green Version]
- Barcia, C.; Ros, C.M.; Ros-Bernal, F.; Gomez, A.; Annese, V.; Carrillo-de Sauvage, M.A.; Yuste, J.E.; Campuzano, C.M.; de Pablos, V.; Fernandez-Villalba, E.; et al. Persistent phagocytic characteristics of microglia in the substantia nigra of long-term Parkinsonian macaques. J. Neuroimmunol. 2013, 261, 60–66. [Google Scholar] [CrossRef]
- Lecours, C.; St-Pierre, M.K.; Picard, K.; Bordeleau, M.; Bourque, M.; Awogbindin, I.O.; Benadjal, A.; Ibanez, F.G.; Gagnon, D.; Cantin, L.; et al. Levodopa partially rescues microglial numerical, morphological, and phagolysosomal alterations in a monkey model of Parkinson’s disease. Brain. Behav. Immun. 2020, 90, 81–96. [Google Scholar] [CrossRef]
- Hurley, S.D.; O’Banion, M.K.; Song, D.D.; Arana, F.S.; Olschowka, J.A.; Haber, S.N. Microglial response is poorly correlated with neurodegeneration following chronic, low-dose MPTP administration in monkeys. Exp. Neurol. 2003, 184, 659–668. [Google Scholar] [CrossRef]
- Carta, A.R.; Mulas, G.; Bortolanza, M.; Duarte, T.; Pillai, E.; Fisone, G.; Vozari, R.R.; Del-Bel, E. l-DOPA-induced dyskinesia and neuroinflammation: Do microglia and astrocytes play a role? Eur. J. Neurosci. 2017, 45, 73–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barcia, C.; Ros, C.M.; Annese, V.; Carrillo-de Sauvage, M.A.; Ros-Bernal, F.; Gomez, A.; Yuste, J.E.; Campuzano, C.M.; de Pablos, V.; Fernandez-Villalba, E.; et al. ROCK/Cdc42-mediated microglial motility and gliapse formation lead to phagocytosis of degenerating dopaminergic neurons in vivo. Sci. Rep. 2012, 2, 809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, D.; Xu, J.; Wang, J.; Tong, J.; Bai, X.; Li, H.; Wang, Z.; Huang, Y.; Wu, Y.; Yu, M.; et al. Dynamic Changes in the Nigrostriatal Pathway in the MPTP Mouse Model of Parkinson’s Disease. Parkinsons Dis. 2017, 2017, 9349487. [Google Scholar] [CrossRef]
- De Miranda, B.R.; Fazzari, M.; Rocha, E.M.; Castro, S.; Greenamyre, J.T. Sex Differences in Rotenone Sensitivity Reflect the Male-to-Female Ratio in Human Parkinson’s Disease Incidence. Toxicol. Sci. 2019, 170, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Siani, F.; Greco, R.; Levandis, G.; Ghezzi, C.; Daviddi, F.; Demartini, C.; Vegeto, E.; Fuzzati-Armentero, M.T.; Blandini, F. Influence of Estrogen Modulation on Glia Activation in a Murine Model of Parkinson’s Disease. Front. Neurosci. 2017, 11, 306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Croisier, E.; Moran, L.B.; Dexter, D.T.; Pearce, R.K.; Graeber, M.B. Microglial inflammation in the parkinsonian substantia nigra: Relationship to alpha-synuclein deposition. J. Neuroinflamm. 2005, 2, 14. [Google Scholar] [CrossRef] [Green Version]
- Imamura, K.; Hishikawa, N.; Sawada, M.; Nagatsu, T.; Yoshida, M.; Hashizume, Y. Distribution of major histocompatibility complex class II-positive microglia and cytokine profile of Parkinson’s disease brains. Acta Neuropathol. 2003, 106, 518–526. [Google Scholar] [CrossRef]
- Langston, J.W.; Forno, L.S.; Tetrud, J.; Reeves, A.G.; Kaplan, J.A.; Karluk, D. Evidence of active nerve cell degeneration in the substantia nigra of humans years after 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine exposure. Ann. Neurol. 1999, 46, 598–605. [Google Scholar] [CrossRef]
- McGeer, P.L.; McGeer, E.G. Glial reactions in Parkinson’s disease. Mov. Disord. 2008, 23, 474–483. [Google Scholar] [CrossRef]
- Hunot, S.; Boissiere, F.; Faucheux, B.; Brugg, B.; Mouatt-Prigent, A.; Agid, Y.; Hirsch, E.C. Nitric oxide synthase and neuronal vulnerability in Parkinson’s disease. Neuroscience 1996, 72, 355–363. [Google Scholar] [CrossRef]
- Hunot, S.; Dugas, N.; Faucheux, B.; Hartmann, A.; Tardieu, M.; Debre, P.; Agid, Y.; Dugas, B.; Hirsch, E.C. FcepsilonRII/CD23 is expressed in Parkinson’s disease and induces, in vitro, production of nitric oxide and tumor necrosis factor-alpha in glial cells. J. Neurosci. 1999, 19, 3440–3447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knott, C.; Stern, G.; Wilkin, G.P. Inflammatory regulators in Parkinson’s disease: iNOS, lipocortin-1, and cyclooxygenases-1 and -2. Mol. Cell. Neurosci. 2000, 16, 724–739. [Google Scholar] [CrossRef] [PubMed]
- Mogi, M.; Harada, M.; Kondo, T.; Riederer, P.; Inagaki, H.; Minami, M.; Nagatsu, T. Interleukin-1 beta, interleukin-6, epidermal growth factor and transforming growth factor-alpha are elevated in the brain from parkinsonian patients. Neurosci. Lett. 1994, 180, 147–150. [Google Scholar] [CrossRef]
- Mogi, M.; Harada, M.; Riederer, P.; Narabayashi, H.; Fujita, K.; Nagatsu, T. Tumor necrosis factor-alpha (TNF-alpha) increases both in the brain and in the cerebrospinal fluid from parkinsonian patients. Neurosci. Lett. 1994, 165, 208–210. [Google Scholar] [CrossRef]
- Gerhard, A.; Pavese, N.; Hotton, G.; Turkheimer, F.; Es, M.; Hammers, A.; Eggert, K.; Oertel, W.; Banati, R.B.; Brooks, D.J. In vivo imaging of microglial activation with [11C](R)-PK11195 PET in idiopathic Parkinson’s disease. Neurobiol. Dis. 2006, 21, 404–412. [Google Scholar] [CrossRef] [PubMed]
- Ouchi, Y.; Yoshikawa, E.; Sekine, Y.; Futatsubashi, M.; Kanno, T.; Ogusu, T.; Torizuka, T. Microglial activation and dopamine terminal loss in early Parkinson’s disease. Ann. Neurol. 2005, 57, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Janda, E.; Boi, L.; Carta, A.R. Microglial Phagocytosis and Its Regulation: A Therapeutic Target in Parkinson’s Disease? Front. Mol. Neurosci. 2018, 11, 144. [Google Scholar] [CrossRef] [Green Version]
- Tremblay, M.E.; Cookson, M.R.; Civiero, L. Glial phagocytic clearance in Parkinson’s disease. Mol. Neurodegener. 2019, 14, 16. [Google Scholar] [CrossRef] [Green Version]
- Barnum, C.J.; Eskow, K.L.; Dupre, K.; Blandino, P., Jr.; Deak, T.; Bishop, C. Exogenous corticosterone reduces L-DOPA-induced dyskinesia in the hemi-parkinsonian rat: Role for interleukin-1beta. Neuroscience 2008, 156, 30–41. [Google Scholar] [CrossRef] [Green Version]
- Boi, L.; Pisanu, A.; Greig, N.H.; Scerba, M.T.; Tweedie, D.; Mulas, G.; Fenu, S.; Carboni, E.; Spiga, S.; Carta, A.R. Immunomodulatory drugs alleviate l-dopa-induced dyskinesia in a rat model of Parkinson’s disease. Mov. Disord. 2019, 34, 1818–1830. [Google Scholar] [CrossRef]
- Bortolanza, M.; Padovan-Neto, F.E.; Cavalcanti-Kiwiatkoski, R.; Dos Santos-Pereira, M.; Mitkovski, M.; Raisman-Vozari, R.; Del-Bel, E. Are cyclooxygenase-2 and nitric oxide involved in the dyskinesia of Parkinson’s disease induced by L-DOPA? Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370. [Google Scholar] [CrossRef] [Green Version]
- Carta, A.R.; Frau, L.; Pisanu, A.; Wardas, J.; Spiga, S.; Carboni, E. Rosiglitazone decreases peroxisome proliferator receptor-gamma levels in microglia and inhibits TNF-alpha production: New evidences on neuroprotection in a progressive Parkinson’s disease model. Neuroscience 2011, 194, 250–261. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos Pereira, M.; Abreu, G.H.D.; Rocca, J.; Hamadat, S.; Raisman-Vozari, R.; Michel, P.P.; Del Bel, E. Contributive Role of TNF-alpha to L-DOPA-Induced Dyskinesia in a Unilateral 6-OHDA Lesion Model of Parkinson’s Disease. Front. Pharmacol. 2020, 11, 617085. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, D.P.; Bortolanza, M.; Del Bel, E.A. Interferon-gamma Involvement in the Neuroinflammation Associated with Parkinson’s Disease and L-DOPA-Induced Dyskinesia. Neurotox. Res. 2021, 39, 705–719. [Google Scholar] [CrossRef]
- Gelders, G.; Baekelandt, V.; Van der Perren, A. Linking Neuroinflammation and Neurodegeneration in Parkinson’s Disease. J. Immunol. Res. 2018, 2018, 4784268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.H.; Lee, H.W.; Hwang, J.; Kim, J.; Lee, M.J.; Han, H.S.; Lee, W.H.; Suk, K. Microglia-inhibiting activity of Parkinson’s disease drug amantadine. Neurobiol. Aging 2012, 33, 2145–2159. [Google Scholar] [CrossRef] [PubMed]
- Martinez, A.A.; Morgese, M.G.; Pisanu, A.; Macheda, T.; Paquette, M.A.; Seillier, A.; Cassano, T.; Carta, A.R.; Giuffrida, A. Activation of PPAR gamma receptors reduces levodopa-induced dyskinesias in 6-OHDA-lesioned rats. Neurobiol. Dis. 2015, 74, 295–304. [Google Scholar] [CrossRef] [Green Version]
- Ossola, B.; Schendzielorz, N.; Chen, S.H.; Bird, G.S.; Tuominen, R.K.; Mannisto, P.T.; Hong, J.S. Amantadine protects dopamine neurons by a dual action: Reducing activation of microglia and inducing expression of GDNF in astroglia [corrected]. Neuropharmacology 2011, 61, 574–582. [Google Scholar] [CrossRef] [Green Version]
- Hardman, C.D.; Halliday, G.M. The external globus pallidus in patients with Parkinson’s disease and progressive supranuclear palsy. Mov. Disord. 1999, 14, 626–633. [Google Scholar] [CrossRef]
- Hardman, C.D.; Halliday, G.M. The internal globus pallidus is affected in progressive supranuclear palsy and Parkinson’s disease. Exp. Neurol. 1999, 158, 135–142. [Google Scholar] [CrossRef]
- Aono, H.; Choudhury, M.E.; Higaki, H.; Miyanishi, K.; Kigami, Y.; Fujita, K.; Akiyama, J.I.; Takahashi, H.; Yano, H.; Kubo, M.; et al. Microglia may compensate for dopaminergic neuron loss in experimental Parkinsonism through selective elimination of glutamatergic synapses from the subthalamic nucleus. Glia 2017, 65, 1833–1847. [Google Scholar] [CrossRef] [PubMed]
- Cenci, M.A.; Jorntell, H.; Petersson, P. On the neuronal circuitry mediating L-DOPA-induced dyskinesia. J. Neural Transm. 2018, 125, 1157–1169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battaglia, G.; Fornai, F.; Busceti, C.L.; Aloisi, G.; Cerrito, F.; De Blasi, A.; Melchiorri, D.; Nicoletti, F. Selective blockade of mGlu5 metabotropic glutamate receptors is protective against methamphetamine neurotoxicity. J. Neurosci. 2002, 22, 2135–2141. [Google Scholar] [CrossRef] [PubMed]
- Black, Y.D.; Xiao, D.; Pellegrino, D.; Kachroo, A.; Brownell, A.L.; Schwarzschild, M.A. Protective effect of metabotropic glutamate mGluR5 receptor elimination in a 6-hydroxydopamine model of Parkinson’s disease. Neurosci. Lett. 2010, 486, 161–165. [Google Scholar] [CrossRef] [Green Version]
- Bruno, V.; Caraci, F.; Copani, A.; Matrisciano, F.; Nicoletti, F.; Battaglia, G. The impact of metabotropic glutamate receptors into active neurodegenerative processes: A ‘dark side’ in the development of new symptomatic treatments for neurologic and psychiatric disorders. Neuropharmacology 2017, 115, 180–192. [Google Scholar] [CrossRef]
- Fuzzati-Armentero, M.T.; Cerri, S.; Levandis, G.; Ambrosi, G.; Montepeloso, E.; Antoninetti, G.; Blandini, F.; Baqi, Y.; Muller, C.E.; Volpini, R.; et al. Dual target strategy: Combining distinct non-dopaminergic treatments reduces neuronal cell loss and synergistically modulates L-DOPA-induced rotational behavior in a rodent model of Parkinson’s disease. J. Neurochem. 2015, 134, 740–747. [Google Scholar] [CrossRef]
- Xia, N.; Zhang, Q.; Wang, S.T.; Gu, L.; Yang, H.M.; Liu, L.; Bakshi, R.; Yang, H.; Zhang, H. Blockade of metabotropic glutamate receptor 5 protects against DNA damage in a rotenone-induced Parkinson’s disease model. Free Radic. Biol. Med. 2015, 89, 567–580. [Google Scholar] [CrossRef]
- Zhang, Y.N.; Fan, J.K.; Gu, L.; Yang, H.M.; Zhan, S.Q.; Zhang, H. Metabotropic glutamate receptor 5 inhibits alpha-synuclein-induced microglia inflammation to protect from neurotoxicity in Parkinson’s disease. J. Neuroinflamm. 2021, 18, 23. [Google Scholar] [CrossRef]
- Ambrosi, G.; Armentero, M.T.; Levandis, G.; Bramanti, P.; Nappi, G.; Blandini, F. Effects of early and delayed treatment with an mGluR5 antagonist on motor impairment, nigrostriatal damage and neuroinflammation in a rodent model of Parkinson’s disease. Brain Res. Bull. 2010, 82, 29–38. [Google Scholar] [CrossRef]
- Holmer, H.K.; Keyghobadi, M.; Moore, C.; Meshul, C.K. l-dopa-induced reversal in striatal glutamate following partial depletion of nigrostriatal dopamine with 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine. Neuroscience 2005, 136, 333–341. [Google Scholar] [CrossRef]
- Morin, N.; Morissette, M.; Gregoire, L.; Di Paolo, T. mGlu5, dopamine D2 and adenosine A2A receptors in L-DOPA-induced dyskinesias. Curr. Neuropharmacol. 2015, 14, 481–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| INTACT (Control) | MPTP | MPTP + L-Dopa | MPTP + L-Dopa + MPEP | |
|---|---|---|---|---|
| Animal data | ||||
| Number of monkeys | 4 | 4 | 5 | 5 |
| Age (year) | 6.2 ± 0.6 | 6.1 ± 0.2 | 7.1 ± 0.3 | 6.0 ± 0.4 |
| Weight (kg) | 3.5 ± 0.1 | 3.6 ± 0.2 | 3.7 ± 0.3 | 3.6 ± 0.1 |
| Parkinsonian score a | - | 9.5 ± 1.2 | 10.3 ± 0.9 | 9.8 ± 0.8 |
| Mean dyskinesias scores (total period) | - | - | 3.3 ± 0.9 | 0.7 ± 0.3 † |
| Survival time post MPTP (months) | - | 3.5 ± 0.9 | 6.6 ± 1.1 | 6.2 ± 1.1 |
| Biochemical data | ||||
| Caudate nucleus post-commissural | ||||
| DA content (ng/mg protein) | 112.06 ± 7.12 | 0.35 ± 0.08 **** | 0.96 ± 0.22 **** | 0.93 ± 0.30 **** |
| 5-HT content (ng/mg protein) | 14.43 ± 0.31 | 9.86 ± 1.44 | 10.43 ± 1.3 | 13.48 ± 1.75 |
| Putamen post-commissural | ||||
| DA content (ng/mg protein) | 94.27 ± 3.94 | 0.79 ± 0.18 **** | 2.13 ± 0.52 **** | 1.33 ± 0.42 **** |
| 5-HT content (ng/mg protein) | 7.70 ± 0.49 | 7.21 ± 1.10 | 6.87 ± 0.75 | 6.91 ± 0.48 |
| Brain Region | Treatment | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MPTP | MPTP + L-Dopa | MPTP + L-Dopa + MPEP | Correlation LID vs | ||||||||||||
| GFAP | Iba1 | CD68 | GFAP | Iba1 | CD68 | GFAP | Iba1 | CD68 | GFAP | Iba1 | CD68 | ||||
| Caudate nucleus Pre-commissural Post-commissural | +40 +71 | −45 −22 | −1 +22 | +84 +73 | −31 +8 | +1 +13 | +86 +56 | −48 −5 | −3 0 | no no | positive no | no no | |||
| Putamen Pre-commissural Post-commissural | +58 +86 | −12 −21 | +14 +22 | +117 +205 | +23 +5 | +49 +37 | +71 +105 | −9 +3 | +26 +23 | no no | positive no | positive no | |||
| Globus pallidus GPe GPi | +9 +8 | +11 −3 | +8 +5 | +24 +26 | +61 +55 | +13 +57 | +24 +12 | +29 +24 | +10 +22 | no no | positive positive | no no | |||
| Substantia nigra | +20 | +10 | +35 | +25 | +37 | +41 | +22 | +22 | +13 | no | positive | no | |||
| Subthalamic nucleus | +18 | +20 | ND | +76 | +33 | ND | +45 | +5 | ND | no | no | ND | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morissette, M.; Bourque, M.; Tremblay, M.-È.; Di Paolo, T. Prevention of L-Dopa-Induced Dyskinesias by MPEP Blockade of Metabotropic Glutamate Receptor 5 Is Associated with Reduced Inflammation in the Brain of Parkinsonian Monkeys. Cells 2022, 11, 691. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11040691
Morissette M, Bourque M, Tremblay M-È, Di Paolo T. Prevention of L-Dopa-Induced Dyskinesias by MPEP Blockade of Metabotropic Glutamate Receptor 5 Is Associated with Reduced Inflammation in the Brain of Parkinsonian Monkeys. Cells. 2022; 11(4):691. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11040691
Chicago/Turabian StyleMorissette, Marc, Mélanie Bourque, Marie-Ève Tremblay, and Thérèse Di Paolo. 2022. "Prevention of L-Dopa-Induced Dyskinesias by MPEP Blockade of Metabotropic Glutamate Receptor 5 Is Associated with Reduced Inflammation in the Brain of Parkinsonian Monkeys" Cells 11, no. 4: 691. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11040691