Regulation of Oncogenic Targets by miR-99a-3p (Passenger Strand of miR-99a-Duplex) in Head and Neck Squamous Cell Carcinoma

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Clinical Human HNSCC Specimens and HNSCC Cell Lines
2.2. RNA Extraction and Quantitative Real-Time Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
2.3. Transfection of miRNAs, siRNAs, and Plasmid Vectors into HNSCC Cells
2.4. Functional Assays in HNSCC Cells (Cell Proliferation, Migration and Invasion Assays)
2.5. Measurement of miR-99a-3p Incorporated into the RISC
2.6. Identification of miR-99a-3p and miR-99a-5p Targets in HNSCC Cells
2.7. Plasmid Construction and Dual-Luciferase Reporter Assays
2.8. Clinical Data Analyses of miRNAs and Target Genes in HNSCC Specimens
2.9. Western Blotting and Immunohistochemistry
2.10. Statistical Analyses
3. Results
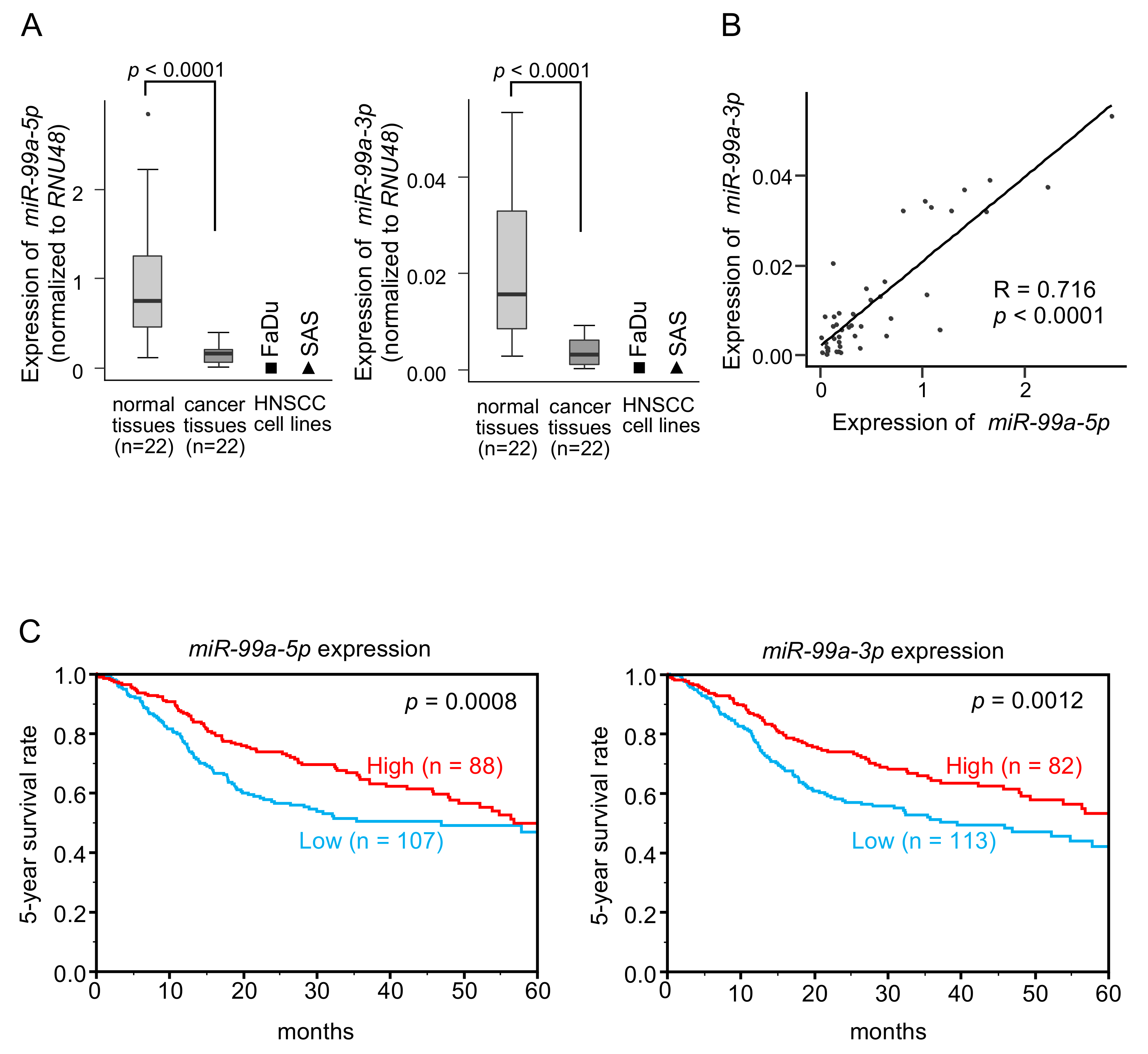
3.1. Downregulation and Clinical Significance of miR-99a-5p and miR-99a-3p in HNSCC Clinical Specimens
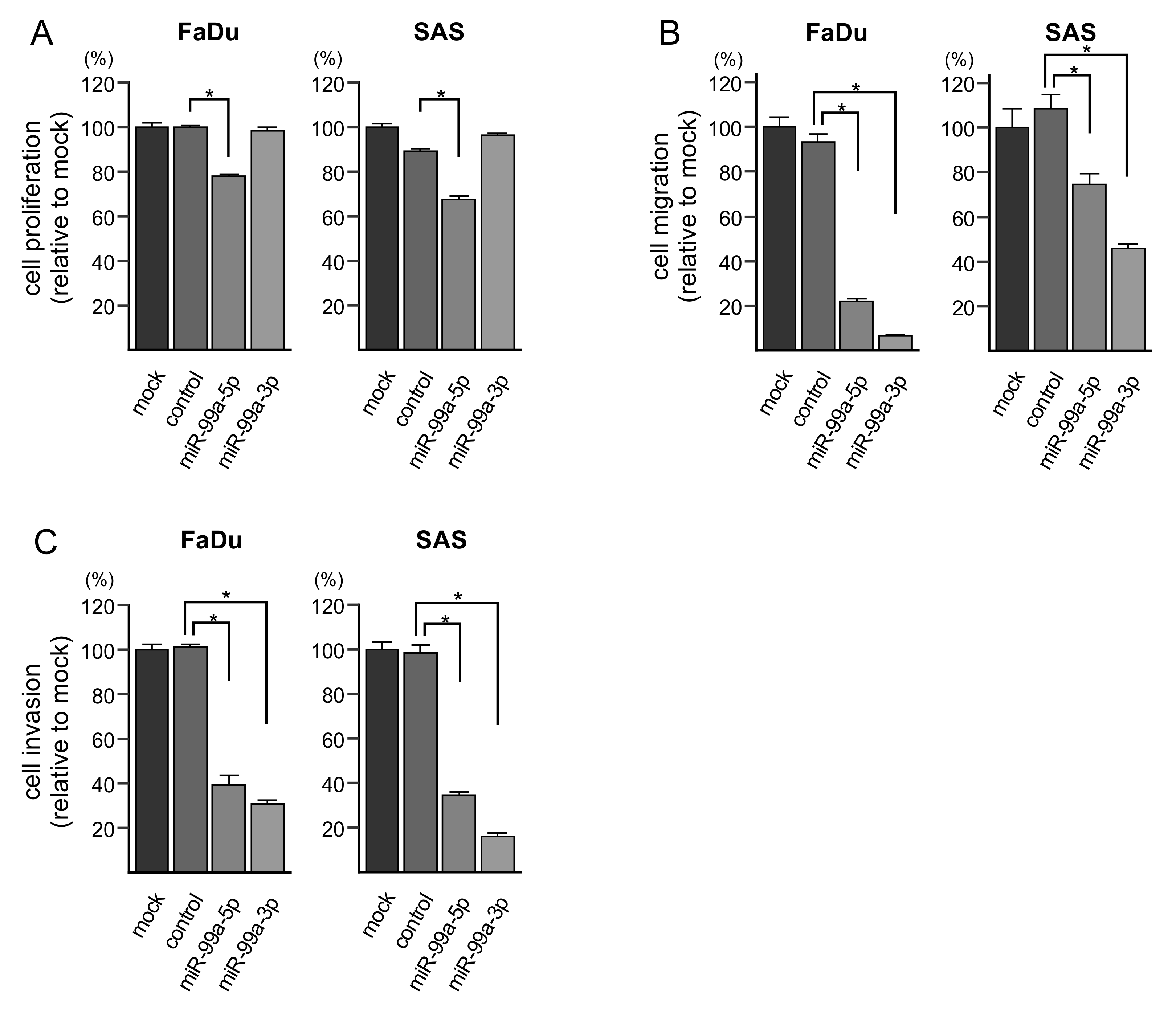
3.2. Ectopic Expression of miR-99a-5p and miR-99a-3p on Cell Proliferation, Migration and Invasion in HNSCC Cells
3.3. Incorporation of miR-99a-5p and miR-99a-3p into the RNA-Induced Silencing Complex (RISC) in HNSCC Cells
3.4. Screening of Molecular Targets Regulated by miR-99a-5p and miR-99a-3p in HNSCC Cells
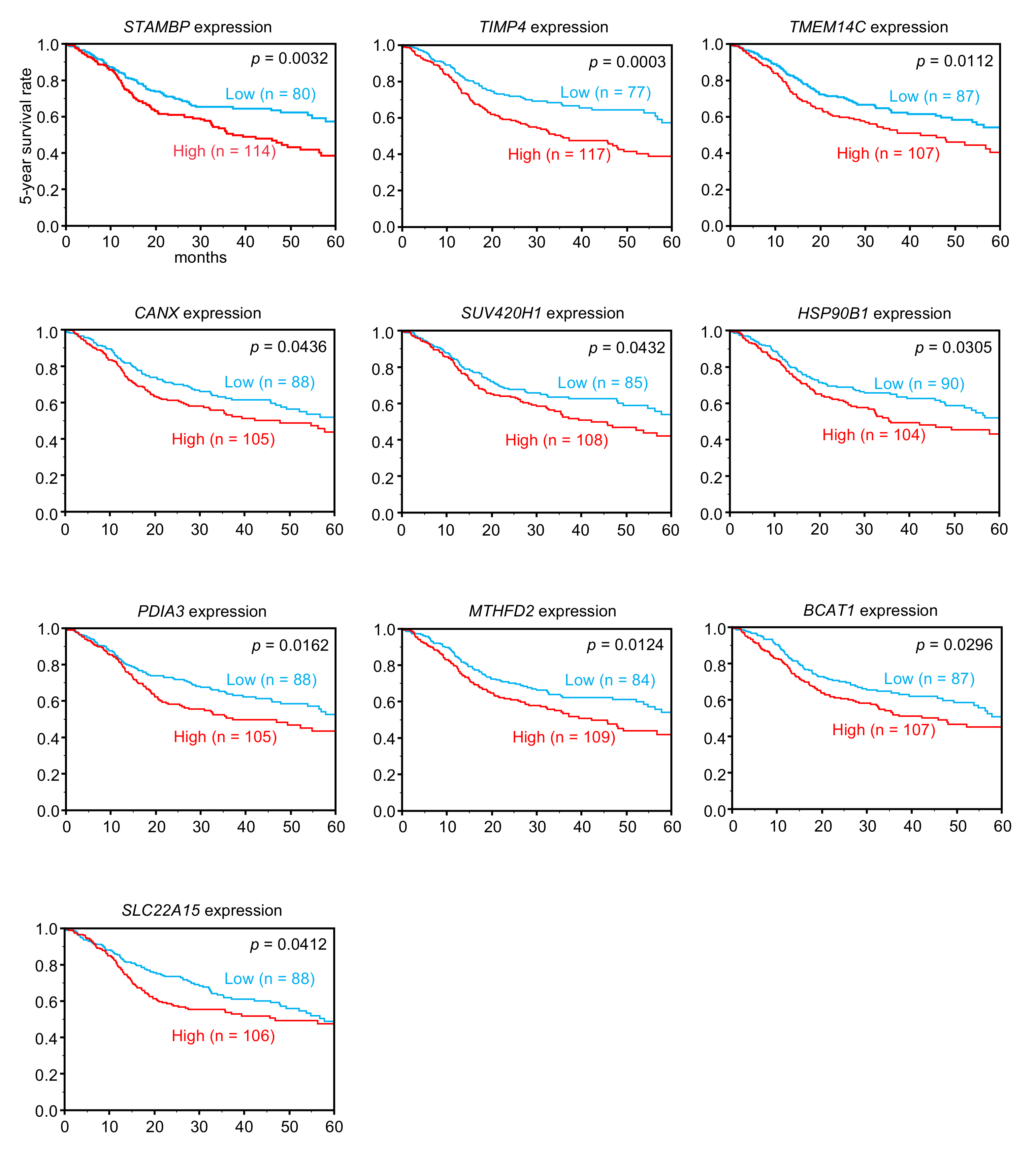
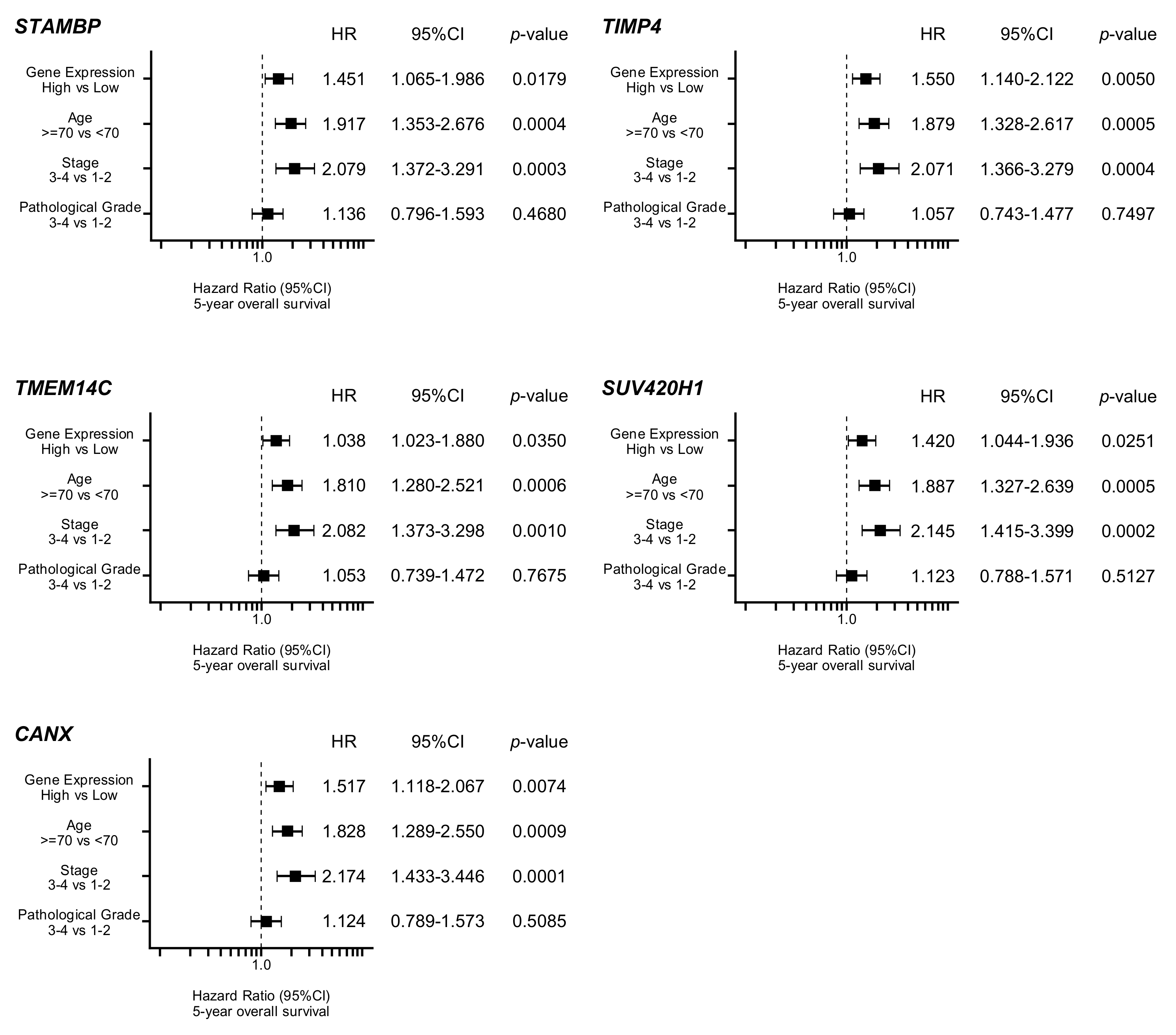
3.5. Clinical Significance of miR-99a-3p Targets in HNSCC Pathogenesis
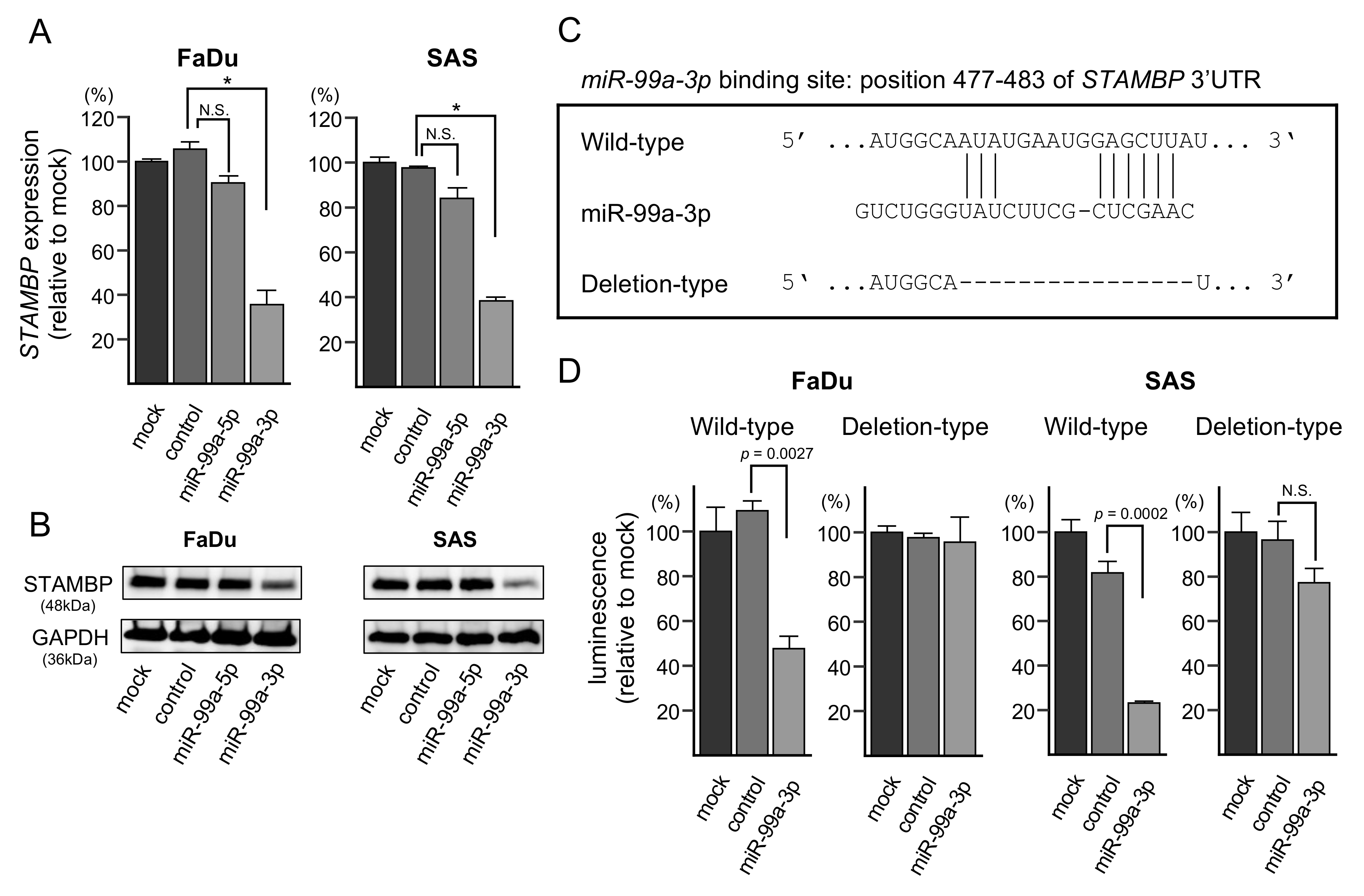
3.6. Direct Regulation of STAMBP by miR-99a-3p in HNSCC Cells
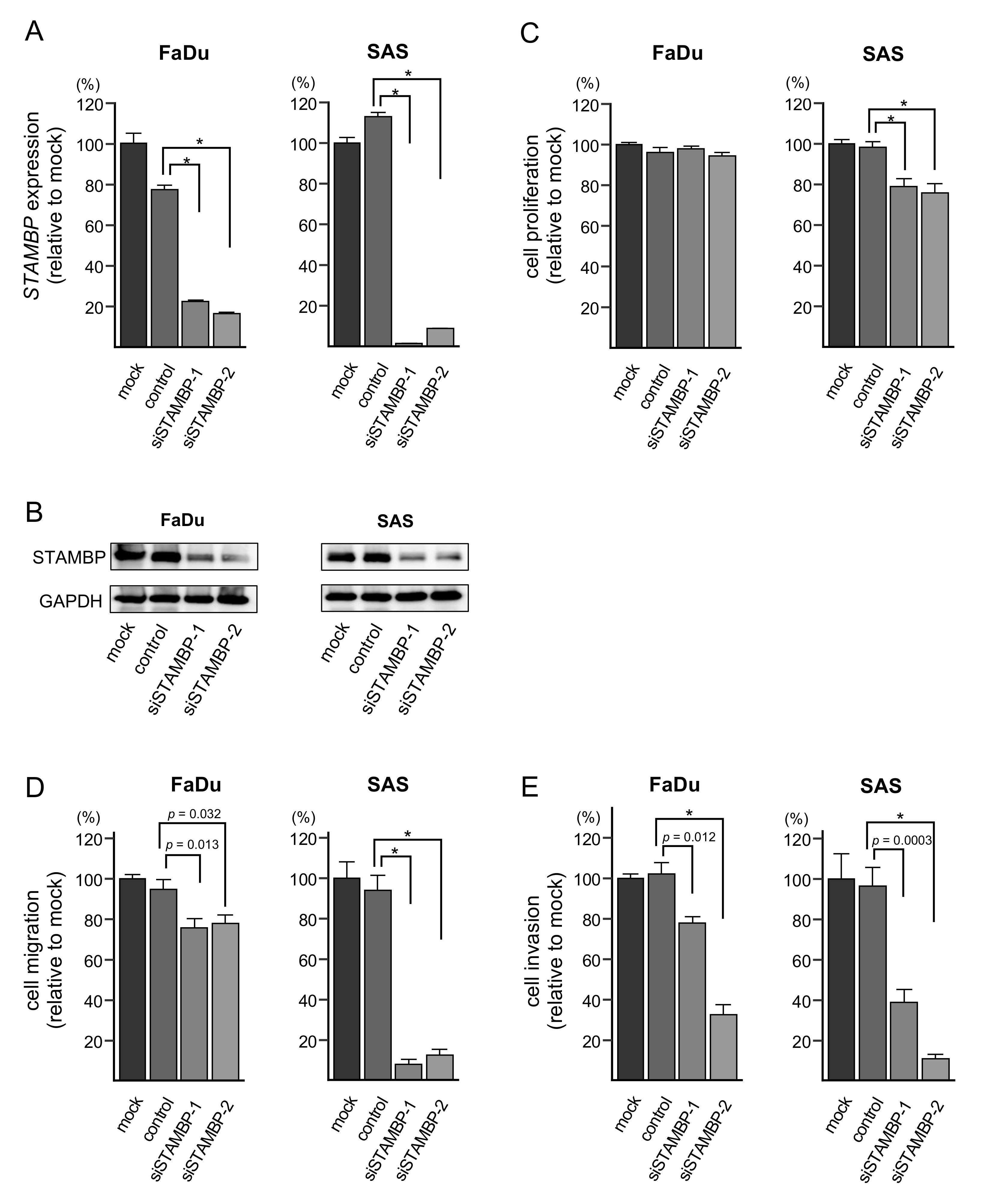
3.7. Effects of STAMBP Knockdown on Cell Proliferation, Migration, and Invasion in HNSCC Cells
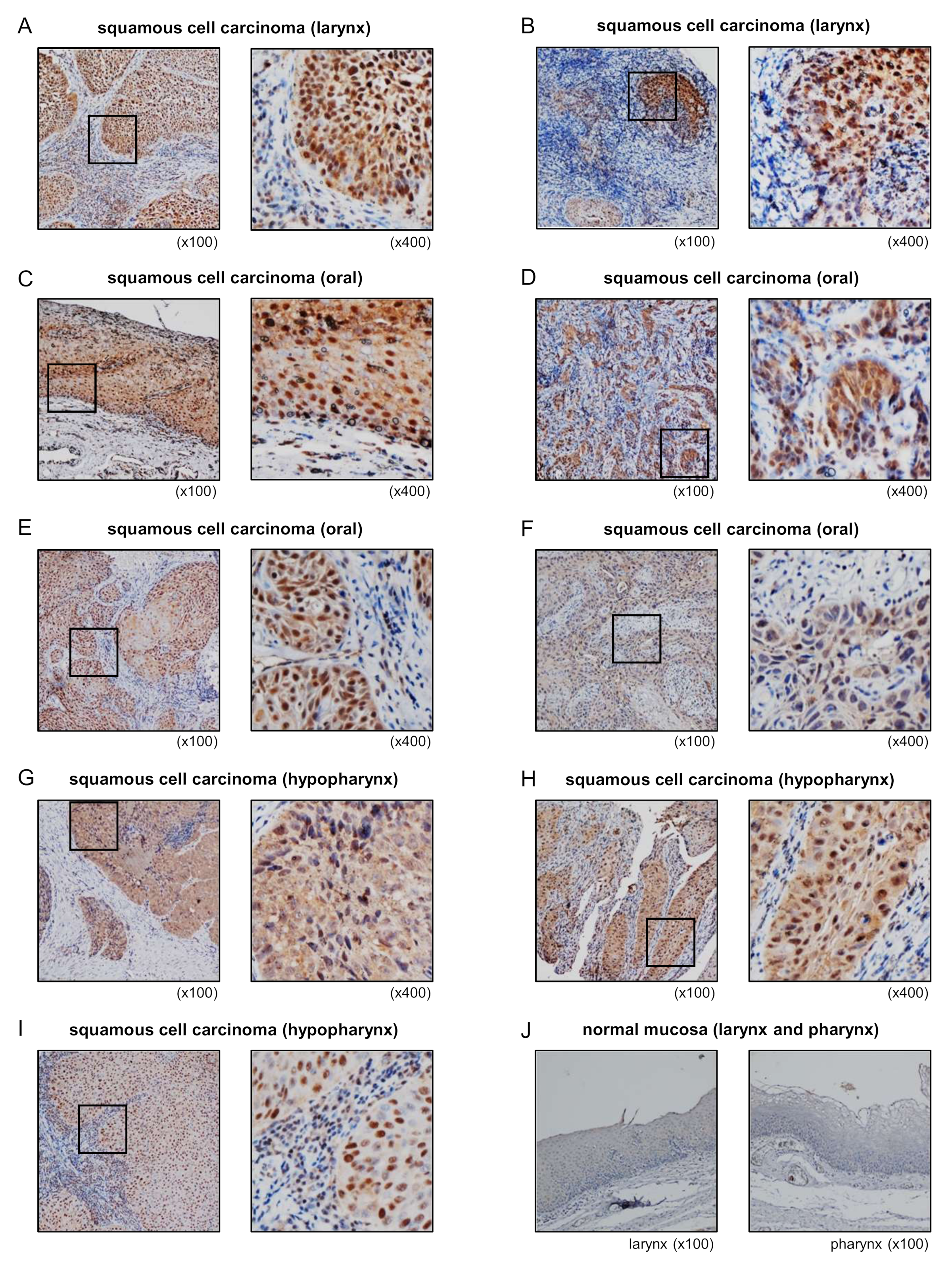
3.8. Overexpression of STAMBP in HNSCC Clinical Specimens
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. CA Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [PubMed]
- Leemans, C.R.; Braakhuis, B.J.; Brakenhoff, R.H. The molecular biology of head and neck cancer. Nat. Rev. Cancer 2011, 11, 9–22. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, G.; Kreimer, A.R.; Viscidi, R.; Pawlita, M.; Fakhry, C.; Koch, W.M.; Westra, W.H.; Gillison, M.L. Case-control study of human papillomavirus and oropharyngeal cancer. N. Engl. J. Med 2007, 356, 1944–1956. [Google Scholar] [CrossRef] [PubMed]
- Seiwert, T.Y.; Zuo, Z.; Keck, M.K.; Khattri, A.; Pedamallu, C.S.; Stricker, T.; Brown, C.; Pugh, T.J.; Stojanov, P.; Cho, J.; et al. Integrative and comparative genomic analysis of HPV-positive and HPV-negative head and neck squamous cell carcinomas. Clin. Cancer Res. 2015, 21, 632–641. [Google Scholar] [CrossRef] [PubMed]
- Bonner, J.A.; Harari, P.M.; Giralt, J.; Cohen, R.B.; Jones, C.U.; Sur, R.K.; Raben, D.; Baselga, J.; Spencer, S.A.; Zhu, J.; et al. Radiotherapy plus cetuximab for locoregionally advanced head and neck cancer: 5-year survival data from a phase 3 randomised trial, and relation between cetuximab-induced rash and survival. Lancet Oncol. 2010, 11, 21–28. [Google Scholar] [CrossRef]
- Ferris, R.L.; Blumenschein, G., Jr.; Fayette, J.; Guigay, J.; Colevas, A.D.; Licitra, L.; Harrington, K.; Kasper, S.; Vokes, E.E.; Even, C.; et al. Nivolumab for Recurrent Squamous-Cell Carcinoma of the Head and Neck. N. Engl. J. Med. 2016, 375, 1856–1867. [Google Scholar] [CrossRef]
- Argiris, A.; Harrington, K.J.; Tahara, M.; Schulten, J.; Chomette, P.; Ferreira Castro, A.; Licitra, L. Evidence-Based Treatment Options in Recurrent and/or Metastatic Squamous Cell Carcinoma of the Head and Neck. Front. Oncol. 2017, 7, 72. [Google Scholar] [CrossRef]
- Hsieh, J.C.; Wang, H.M.; Wu, M.H.; Chang, K.P.; Chang, P.H.; Liao, C.T.; Liau, C.T. Review of emerging biomarkers in head and neck squamous cell carcinoma in the era of immunotherapy and targeted therapy. Head Neck 2019, 41, 19–45. [Google Scholar] [CrossRef]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef]
- Gebert, L.F.R.; MacRae, I.J. Regulation of microRNA function in animals. Nat. Rev. Mol. Cell Biol. 2019, 20, 21–37. [Google Scholar] [CrossRef]
- Nohata, N.; Hanazawa, T.; Kinoshita, T.; Okamoto, Y.; Seki, N. MicroRNAs function as tumor suppressors or oncogenes: aberrant expression of microRNAs in head and neck squamous cell carcinoma. Auris Nasus Larynx 2013, 40, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Gregory, R.I. MicroRNA biogenesis pathways in cancer. Nat. Rev. Cancer 2015, 15, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Koshizuka, K.; Hanazawa, T.; Arai, T.; Okato, A.; Kikkawa, N.; Seki, N. Involvement of aberrantly expressed microRNAs in the pathogenesis of head and neck squamous cell carcinoma. Cancer Metastasis Rev. 2017, 36, 525–545. [Google Scholar] [CrossRef] [PubMed]
- Rupaimoole, R.; Slack, F.J. MicroRNA therapeutics: towards a new era for the management of cancer and other diseases. Nat. Rev. Drug Discov. 2017, 16, 203–222. [Google Scholar] [CrossRef] [PubMed]
- Anfossi, S.; Babayan, A.; Pantel, K.; Calin, G.A. Clinical utility of circulating non-coding RNAs-an update. Nat. Rev. Clin. Oncol. 2018, 15, 541–563. [Google Scholar] [CrossRef] [PubMed]
- Kikkawa, N.; Hanazawa, T.; Fujimura, L.; Nohata, N.; Suzuki, H.; Chazono, H.; Sakurai, D.; Horiguchi, S.; Okamoto, Y.; Seki, N. miR-489 is a tumour-suppressive miRNA target PTPN11 in hypopharyngeal squamous cell carcinoma (HSCC). Br. J. Cancer 2010, 103, 877–884. [Google Scholar] [CrossRef] [PubMed]
- Nohata, N.; Hanazawa, T.; Kikkawa, N.; Sakurai, D.; Fujimura, L.; Chiyomaru, T.; Kawakami, K.; Yoshino, H.; Enokida, H.; Nakagawa, M.; et al. Tumour suppressive microRNA-874 regulates novel cancer networks in maxillary sinus squamous cell carcinoma. Br. J. Cancer 2011, 105, 833–841. [Google Scholar] [CrossRef]
- Fukumoto, I.; Kinoshita, T.; Hanazawa, T.; Kikkawa, N.; Chiyomaru, T.; Enokida, H.; Yamamoto, N.; Goto, Y.; Nishikawa, R.; Nakagawa, M.; et al. Identification of tumour suppressive microRNA-451a in hypopharyngeal squamous cell carcinoma based on microRNA expression signature. Br. J. Cancer 2014, 111, 386–394. [Google Scholar] [CrossRef]
- Fukumoto, I.; Hanazawa, T.; Kinoshita, T.; Kikkawa, N.; Koshizuka, K.; Goto, Y.; Nishikawa, R.; Chiyomaru, T.; Enokida, H.; Nakagawa, M.; et al. MicroRNA expression signature of oral squamous cell carcinoma: functional role of microRNA-26a/b in the modulation of novel cancer pathways. Br. J. Cancer 2015, 112, 891–900. [Google Scholar] [CrossRef]
- Koshizuka, K.; Nohata, N.; Hanazawa, T.; Kikkawa, N.; Arai, T.; Okato, A.; Fukumoto, I.; Katada, K.; Okamoto, Y.; Seki, N. Deep sequencing-based microRNA expression signatures in head and neck squamous cell carcinoma: Dual strands of pre-miR-150 as antitumor miRNAs. Oncotarget 2017, 8, 30288–30304. [Google Scholar] [CrossRef]
- Kinoshita, T.; Nohata, N.; Hanazawa, T.; Kikkawa, N.; Yamamoto, N.; Yoshino, H.; Itesako, T.; Enokida, H.; Nakagawa, M.; Okamoto, Y.; et al. Tumour-suppressive microRNA-29s inhibit cancer cell migration and invasion by targeting laminin-integrin signalling in head and neck squamous cell carcinoma. Br. J. Cancer 2013, 109, 2636–2645. [Google Scholar] [CrossRef] [PubMed]
- Koshizuka, K.; Kikkawa, N.; Hanazawa, T.; Yamada, Y.; Okato, A.; Arai, T.; Katada, K.; Okamoto, Y.; Seki, N. Inhibition of integrin beta1-mediated oncogenic signalling by the antitumor microRNA-29 family in head and neck squamous cell carcinoma. Oncotarget 2018, 9, 3663–3676. [Google Scholar] [CrossRef] [PubMed]
- Koshizuka, K.; Hanazawa, T.; Kikkawa, N.; Arai, T.; Okato, A.; Kurozumi, A.; Kato, M.; Katada, K.; Okamoto, Y.; Seki, N. Regulation of ITGA3 by the anti-tumor miR-199 family inhibits cancer cell migration and invasion in head and neck cancer. Cancer Sci. 2017, 108, 1681–1692. [Google Scholar] [CrossRef] [PubMed]
- Fukumoto, I.; Kikkawa, N.; Matsushita, R.; Kato, M.; Kurozumi, A.; Nishikawa, R.; Goto, Y.; Koshizuka, K.; Hanazawa, T.; Enokida, H.; et al. Tumor-suppressive microRNAs (miR-26a/b, miR-29a/b/c and miR-218) concertedly suppressed metastasis-promoting LOXL2 in head and neck squamous cell carcinoma. J. Hum. Genet. 2016, 61, 109–118. [Google Scholar] [CrossRef]
- Chen, D.; Cabay, R.J.; Jin, Y.; Wang, A.; Lu, Y.; Shah-Khan, M.; Zhou, X. MicroRNA Deregulations in Head and Neck Squamous Cell Carcinomas. J. Oral. Maxillofac. Res. 2013, 4, e2. [Google Scholar] [CrossRef]
- Chen, Y.T.; Yao, J.N.; Qin, Y.T.; Hu, K.; Wu, F.; Fang, Y.Y. Biological role and clinical value of miR-99a-5p in head and neck squamous cell carcinoma (HNSCC): A bioinformatics-based study. FEBS Open Bio 2018, 8, 1280–1298. [Google Scholar] [CrossRef]
- Yamada, Y.; Koshizuka, K.; Hanazawa, T.; Kikkawa, N.; Okato, A.; Idichi, T.; Arai, T.; Sugawara, S.; Katada, K.; Okamoto, Y.; et al. Passenger strand of miR-145-3p acts as a tumor-suppressor by targeting MYO1B in head and neck squamous cell carcinoma. Int. J. Oncol. 2018, 52, 166–178. [Google Scholar] [CrossRef]
- Goto, Y.; Kurozumi, A.; Arai, T.; Nohata, N.; Kojima, S.; Okato, A.; Kato, M.; Yamazaki, K.; Ishida, Y.; Naya, Y.; et al. Impact of novel miR-145-3p regulatory networks on survival in patients with castration-resistant prostate cancer. Br. J. Cancer 2017, 117, 409–420. [Google Scholar] [CrossRef]
- Misono, S.; Seki, N.; Mizuno, K.; Yamada, Y.; Uchida, A.; Arai, T.; Kumamoto, T.; Sanada, H.; Suetsugu, T.; Inoue, H. Dual strands of the miR-145 duplex (miR-145-5p and miR-145-3p) regulate oncogenes in lung adenocarcinoma pathogenesis. J. Hum. Genet. 2018, 63, 1015–1028. [Google Scholar] [CrossRef]
- Misono, S.; Seki, N.; Mizuno, K.; Yamada, Y.; Uchida, A.; Sanada, H.; Moriya, S.; Kikkawa, N.; Kumamoto, T.; Suetsugu, T.; et al. Molecular Pathogenesis of Gene Regulation by the miR-150 Duplex: miR-150-3p Regulates TNS4 in Lung Adenocarcinoma. Cancers 2019, 11. [Google Scholar] [CrossRef]
- Yamada, Y.; Arai, T.; Kojima, S.; Sugawara, S.; Kato, M.; Okato, A.; Yamazaki, K.; Naya, Y.; Ichikawa, T.; Seki, N. Regulation of antitumor miR-144-5p targets oncogenes: Direct regulation of syndecan-3 and its clinical significance. Cancer Sci. 2018, 109, 2919–2936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uchida, A.; Seki, N.; Mizuno, K.; Misono, S.; Yamada, Y.; Kikkawa, N.; Sanada, H.; Kumamoto, T.; Suetsugu, T.; Inoue, H. Involvement of dual-strand of the miR-144 duplex and their targets in the pathogenesis of lung squamous cell carcinoma. Cancer Sci. 2019, 110, 420–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanada, H.; Seki, N.; Mizuno, K.; Misono, S.; Uchida, A.; Yamada, Y.; Moriya, S.; Kikkawa, N.; Machida, K.; Kumamoto, T.; et al. Involvement of Dual Strands of miR-143 (miR-143-5p and miR-143-3p) and Their Target Oncogenes in the Molecular Pathogenesis of Lung Adenocarcinoma. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimonosono, M.; Idichi, T.; Seki, N.; Yamada, Y.; Arai, T.; Arigami, T.; Sasaki, K.; Omoto, I.; Uchikado, Y.; Kita, Y.; et al. Molecular pathogenesis of esophageal squamous cell carcinoma: Identification of the antitumor effects of miR1453p on gene regulation. Int. J. Oncol. 2019, 54, 673–688. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Bo, Z.; Pang, G.; Qu, X.; Bao, W.; Yang, L.; Ma, Y. MiR-99a-5p regulates proliferation, migration and invasion abilities of human oral carcinoma cells by targeting NOX4. Neoplasma 2017, 64, 666–673. [Google Scholar] [CrossRef]
- Tsai, T.F.; Lin, J.F.; Chou, K.Y.; Lin, Y.C.; Chen, H.E.; Hwang, T.I. miR-99a-5p acts as tumor suppressor via targeting to mTOR and enhances RAD001-induced apoptosis in human urinary bladder urothelial carcinoma cells. OncoTargets Ther. 2018, 11, 239–252. [Google Scholar] [CrossRef] [Green Version]
- Yin, H.; Ma, J.; Chen, L.; Piao, S.; Zhang, Y.; Zhang, S.; Ma, H.; Li, Y.; Qu, Y.; Wang, X.; et al. MiR-99a Enhances the Radiation Sensitivity of Non-Small Cell Lung Cancer by Targeting mTOR. Cell Physiol. Biochem. 2018, 46, 471–481. [Google Scholar] [CrossRef]
- Liu, Y.; Li, B.; Yang, X.; Zhang, C. MiR-99a-5p inhibits bladder cancer cell proliferation by directly targeting mammalian target of rapamycin and predicts patient survival. J. Cell Biochem. 2019, 120, 19330–19337. [Google Scholar] [CrossRef]
- Tao, C.; Sun, H.; Sang, W.; Li, S. miRNA-99a inhibits cell invasion and migration in liver cancer by directly targeting HOXA1. Oncol. Lett. 2019, 17, 5108–5114. [Google Scholar] [CrossRef] [Green Version]
- Arai, T.; Okato, A.; Yamada, Y.; Sugawara, S.; Kurozumi, A.; Kojima, S.; Yamazaki, K.; Naya, Y.; Ichikawa, T.; Seki, N. Regulation of NCAPG by miR-99a-3p (passenger strand) inhibits cancer cell aggressiveness and is involved in CRPC. Cancer Med. 2018, 7, 1988–2002. [Google Scholar] [CrossRef]
- Osako, Y.; Yoshino, H.; Sakaguchi, T.; Sugita, S.; Yonemori, M.; Nakagawa, M.; Enokida, H. Potential tumorsuppressive role of microRNA99a3p in sunitinibresistant renal cell carcinoma cells through the regulation of RRM2. Int. J. Oncol. 2019, 54, 1759–1770. [Google Scholar] [CrossRef] [PubMed]
- Wei, G.G.; Guo, W.P.; Tang, Z.Y.; Li, S.H.; Wu, H.Y.; Zhang, L.C. Expression level and prospective mechanism of miRNA-99a-3p in head and neck squamous cell carcinoma based on miRNA-chip and miRNA-sequencing data in 1, 167 cases. Pathol. Res. Pr. 2019, 215, 963–976. [Google Scholar] [CrossRef] [PubMed]
- Boufraqech, M.; Zhang, L.; Nilubol, N.; Sadowski, S.M.; Kotian, S.; Quezado, M.; Kebebew, E. Lysyl Oxidase (LOX) Transcriptionally Regulates SNAI2 Expression and TIMP4 Secretion in Human Cancers. Clin. Cancer Res. 2016, 22, 4491–4504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, D.; Carberry, S.; Murphy, A.C.; Lindner, A.U.; Fay, J.; Hector, S.; McCawley, N.; Bacon, O.; Concannon, C.G.; Kay, E.W.; et al. Calnexin, an ER stress-induced protein, is a prognostic marker and potential therapeutic target in colorectal cancer. J. Transl. Med. 2016, 14, 196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, M.; Nagashio, R.; Jiang, S.X.; Saito, K.; Tsuchiya, B.; Ryuge, S.; Katono, K.; Nakashima, H.; Fukuda, E.; Goshima, N.; et al. Calnexin is a novel sero-diagnostic marker for lung cancer. Lung Cancer 2015, 90, 342–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vougiouklakis, T.; Sone, K.; Saloura, V.; Cho, H.S.; Suzuki, T.; Dohmae, N.; Alachkar, H.; Nakamura, Y.; Hamamoto, R. SUV420H1 enhances the phosphorylation and transcription of ERK1 in cancer cells. Oncotarget 2015, 6, 43162–43171. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Seth, A. An RNF11: Smurf2 complex mediates ubiquitination of the AMSH protein. Oncogene 2004, 23, 1801–1808. [Google Scholar] [CrossRef] [Green Version]
- McCullough, J.; Clague, M.J.; Urbe, S. AMSH is an endosome-associated ubiquitin isopeptidase. J. Cell Biol. 2004, 166, 487–492. [Google Scholar] [CrossRef] [Green Version]
- Agromayor, M.; Martin-Serrano, J. Interaction of AMSH with ESCRT-III and deubiquitination of endosomal cargo. J. Biol. Chem. 2006, 281, 23083–23091. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.M.; Boucrot, E.; Villen, J.; Affar el, B.; Gygi, S.P.; Gottlinger, H.G.; Kirchhausen, T. Targeting of AMSH to endosomes is required for epidermal growth factor receptor degradation. J. Biol. Chem. 2007, 282, 9805–9812. [Google Scholar] [CrossRef] [Green Version]
- Meijer, I.M.; van Rotterdam, W.; van Zoelen, E.J.; van Leeuwen, J.E. Recycling of EGFR and ErbB2 is associated with impaired Hrs tyrosine phosphorylation and decreased deubiquitination by AMSH. Cell Signal. 2012, 24, 1981–1988. [Google Scholar] [CrossRef] [PubMed]
- McDonell, L.M.; Mirzaa, G.M.; Alcantara, D.; Schwartzentruber, J.; Carter, M.T.; Lee, L.J.; Clericuzio, C.L.; Graham, J.M., Jr.; Morris-Rosendahl, D.J.; Polster, T.; et al. Mutations in STAMBP, encoding a deubiquitinating enzyme, cause microcephaly-capillary malformation syndrome. Nat. Genet. 2013, 45, 556–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwakami, Y.; Yokoyama, S.; Watanabe, K.; Hayakawa, Y. STAM-binding protein regulates melanoma metastasis through SLUG stabilization. Biochem. Biophys. Res. Commun. 2018, 507, 484–488. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Age | Sex | Location | T | N | M | Stage | Differentiation |
|---|---|---|---|---|---|---|---|---|
| 1 | 66 | M | hypopharynx | 2 | 2c | 0 | IVa | moderate |
| 2 | 66 | M | hypopharynx | 4a | 2c | 0 | IVa | well |
| 3 | 66 | M | hypopharynx | 4b | 2c | 0 | IVb | moderate |
| 4 | 76 | M | hypopharynx | 4a | 1 | 0 | IVa | well |
| 5 | 74 | M | hypopharynx | 4a | 2c | 0 | IVa | poor |
| 6 | 45 | M | hypopharynx | 4a | 2c | 0 | IVa | moderate |
| 7 | 75 | M | hypopharynx | 4a | 2c | 0 | IVa | well |
| 8 | 58 | M | hypopharynx | 4a | 0 | 0 | IVa | well |
| 9 | 69 | M | larynx | 3 | 0 | 0 | III | well |
| 10 | 70 | M | larynx | 4a | 1 | 0 | IVa | well-moderate |
| 11 | 84 | M | larynx | 4a | 0 | 0 | IVa | moderate |
| 12 | 50 | M | larynx | 4a | 2b | 0 | IVa | moderate |
| 13 | 82 | M | larynx | 4a | 0 | 0 | IVa | moderate |
| 14 | 85 | M | larynx | 3 | 2b | 0 | IVa | moderate |
| 15 | 66 | M | tongue | 2 | 0 | 0 | II | moderate |
| 16 | 73 | M | tongue | 3 | 1 | 0 | III | poor |
| 17 | 74 | M | tongue | 1 | 0 | 0 | I | well |
| 18 | 72 | M | tongue | 4a | 2b | 0 | IVa | moderate |
| 19 | 83 | M | oral floor | 2 | 1 | 0 | III | well |
| 20 | 68 | F | oral floor | 4a | 1 | 0 | IVa | well |
| 21 | 77 | M | oral floor | 2 | 2b | 0 | IVa | moderate |
| 22 | 69 | M | oropharynx | 1 | 0 | 0 | I | well |
| Entrez Gene ID | Gene Symbol | Gene Name | Total Sites | FaDu miR-99a-3p Transfectant FC (log2) | TCGA OncoLnc 5-Year OS p-Value |
|---|---|---|---|---|---|
| 10617 | STAMBP | STAM binding protein | 1 | −1.0548 | 0.0032 |
| 7079 | TIMP4 | TIMP metallopeptidase inhibitor 4 | 1 | −2.2976 | 0.0003 |
| 51522 | TMEM14C | transmembrane protein 14C | 1 | −3.0740 | 0.0112 |
| 51111 | SUV420H1 (KMT5B) | suppressor of variegation 4-20 homolog 1 | 1 | −1.0238 | 0.0432 |
| 821 | CANX | calnexin | 1 | −2.3590 | 0.0436 |
| 10797 | MTHFD2 | methylenetetrahydrofolate dehydrogenase (NADP+ dependent) 2,methenyltetrahydrofolate cyclohydrolase | 1 | −1.6298 | 0.0124 |
| 2923 | PDIA3 | protein disulfide isomerase family A, member 3 | 1 | −1.3332 | 0.0162 |
| 586 | BCAT1 | branched chain amino-acid transaminase 1, cytosolic | 3 | −2.0236 | 0.0296 |
| 7184 | HSP90B1 | heat shock protein 90kDa beta (Grp94), member 1 | 1 | −2.3549 | 0.0305 |
| 55356 | SLC22A15 | solute carrier family 22, member 15 | 1 | −1.9007 | 0.0412 |
| 23786 | BCL2L13 | BCL2-like 13 (apoptosis facilitator) | 1 | −1.2622 | 0.0604 |
| 29967 | LRP12 | low density lipoprotein receptor-related protein 12 | 1 | −1.0258 | 0.0897 |
| 112752 | IFT43 | intraflagellar transport 43 | 1 | −1.4595 | 0.0922 |
| 23516 | SLC39A14 | solute carrier family 39 (zinc transporter), member 14 | 1 | −1.6435 | 0.1124 |
| 55255 | WDR41 | WD repeat domain 41 | 1 | −1.2410 | 0.1145 |
| 56886 | UGGT1 | UDP-glucose glycoprotein glucosyltransferase 1 | 1 | −1.0232 | 0.1228 |
| 6137 | RPL13 | ribosomal protein L13 | 1 | −1.6427 | 0.1408 |
| 114971 | PTPMT1 | protein tyrosine phosphatase, mitochondrial 1 | 1 | −1.2094 | 0.1545 |
| 27 | ABL2 | ABL proto-oncogene 2, non-receptor tyrosine kinase | 1 | −1.8956 | 0.1549 |
| 114818 | KLHL29 | kelch-like family member 29 | 1 | −1.4701 | 0.1551 |
| 2512 | FTL | ferritin, light polypeptide | 1 | −1.2722 | 0.1809 |
| 84803 | AGPAT9 (GPAT3) | 1-acylglycerol-3-phosphate O-acyltransferase 9 | 1 | −1.8971 | 0.2008 |
| 23271 | CAMSAP2 | calmodulin regulated spectrin-associated protein family, member 2 | 1 | −1.0735 | 0.3277 |
| 122953 | JDP2 | Jun dimerization protein 2 | 1 | −2.2594 | 0.3311 |
| 219902 | TMEM136 | transmembrane protein 136 | 1 | −1.4956 | 0.3455 |
| 440026 | TMEM41B | transmembrane protein 41B | 1 | −2.3021 | 0.3843 |
| 54629 | FAM63B | family with sequence similarity 63, member B | 1 | −1.2972 | 0.3940 |
| 182 | JAG1 | jagged 1 | 1 | −1.0082 | 0.3971 |
| 2121 | EVC | Ellis van Creveld syndrome | 1 | −1.7233 | 0.4006 |
| 490 | ATP2B1 | ATPase, Ca++ transporting, plasma membrane 1 | 1 | −2.0436 | 0.5557 |
| 9208 | LRRFIP1 | leucine rich repeat (in FLII) interacting protein 1 | 1 | −1.1882 | 0.6483 |
| 50848 | F11R | F11 receptor | 1 | −1.0283 | 0.7312 |
| 79152 | FA2H | fatty acid 2-hydroxylase | 1 | −1.7458 | 0.0877 |
| 23049 | SMG1 | SMG1 phosphatidylinositol 3-kinase-related kinase | 1 | −1.1426 | 0.1267 |
| 5337 | PLD1 | phospholipase D1, phosphatidylcholine-specific | 1 | −1.0968 | 0.1538 |
| 5935 | RBM3 | RNA binding motif (RNP1, RRM) protein 3 | 1 | −1.3755 | 0.1554 |
| 135398 | C6orf141 | chromosome 6 open reading frame 141 | 1 | −1.4022 | 0.1594 |
| 5251 | PHEX | phosphate regulating endopeptidase homolog, X-linked | 1 | −1.0845 | 0.1616 |
| 201229 | LYRM9 | LYR motif containing 9 | 1 | −1.8318 | 0.1709 |
| 6095 | RORA | RAR-related orphan receptor A | 1 | −1.6688 | 0.1809 |
| 85439 | STON2 | stonin 2 | 2 | −1.0335 | 0.2107 |
| 114781 | BTBD9 | BTB (POZ) domain containing 9 | 1 | −1.4022 | 0.2145 |
| 144348 | ZNF664 | zinc finger protein 664 | 1 | −1.1221 | 0.2262 |
| 27125 | AFF4 | AF4/FMR2 family, member 4 | 1 | −1.3648 | 0.2620 |
| 152007 | GLIPR2 | GLI pathogenesis-related 2 | 1 | −1.8377 | 0.2882 |
| 688 | KLF5 | Kruppel-like factor 5 (intestinal) | 1 | −1.0461 | 0.3321 |
| 27250 | PDCD4 | programmed cell death 4 (neoplastic transformation inhibitor) | 1 | −1.3298 | 0.3380 |
| 440295 | GOLGA6L9 | golgin A6 family-like 9 | 2 | −1.5553 | 0.3385 |
| 55175 | KLHL11 | kelch-like family member 11 | 1 | −1.1230 | 0.3808 |
| 85015 | USP45 | ubiquitin specific peptidase 45 | 1 | −1.0026 | 0.3813 |
| 27109 | ATP5S | ATP synthase, H+ transporting, mitochondrial Fo complex, subunit s (factor B) | 1 | −1.0441 | 0.3939 |
| 10802 | SEC24A | SEC24 family member A | 1 | −1.1434 | 0.4081 |
| 2639 | GCDH | glutaryl-CoA dehydrogenase | 1 | −1.2904 | 0.4110 |
| 843 | CASP10 | caspase 10, apoptosis-related cysteine peptidase | 1 | −1.1677 | 0.4282 |
| 8774 | NAPG | N-ethylmaleimide-sensitive factor attachment protein, gamma | 1 | −1.1850 | 0.4452 |
| 54462 | CCSER2 | coiled-coil serine-rich protein 2 | 1 | −1.3840 | 0.4566 |
| 9848 | MFAP3L | microfibrillar-associated protein 3-like | 1 | −1.1114 | 0.4710 |
| 64764 | CREB3L2 | cAMP responsive element binding protein 3-like 2 | 1 | −2.2962 | 0.4754 |
| 334 | APLP2 | amyloid beta (A4) precursor-like protein 2 | 1 | −1.3719 | 0.5068 |
| 5163 | PDK1 | pyruvate dehydrogenase kinase, isozyme 1 | 1 | −1.1852 | 0.5112 |
| 10124 | ARL4A | ADP-ribosylation factor-like 4A | 1 | −1.4088 | 0.5161 |
| 145781 | GCOM1 | GRINL1A complex locus 1 | 1 | −1.0388 | 0.5289 |
| 3987 | LIMS1 | LIM and senescent cell antigen-like domains 1 | 1 | −1.0346 | 0.5454 |
| 57498 | KIDINS220 | kinase D-interacting substrate, 220kDa | 1 | −1.4304 | 0.5664 |
| 285636 | C5orf51 | chromosome 5 open reading frame 51 | 1 | −1.0631 | 0.5666 |
| 9761 | MLEC | malectin | 1 | −1.6201 | 0.5725 |
| 54477 | PLEKHA5 | pleckstrin homology domain containing, family A member 5 | 1 | −1.3145 | 0.5846 |
| 10221 | TRIB1 | tribbles pseudokinase 1 | 1 | −1.7224 | 0.5958 |
| 54014 | BRWD1 | bromodomain and WD repeat domain containing 1 | 1 | −1.0694 | 0.6033 |
| 390 | RND3 | Rho family GTPase 3 | 1 | −1.0681 | 0.6139 |
| 55823 | VPS11 | vacuolar protein sorting 11 homolog (S. cerevisiae) | 1 | −1.0569 | 0.6269 |
| 8444 | DYRK3 | dual-specificity tyrosine-(Y)-phosphorylation regulated kinase 3 | 1 | −1.9227 | 0.6459 |
| 1978 | EIF4EBP1 | eukaryotic translation initiation factor 4E binding protein 1 | 1 | −1.2832 | 0.6529 |
| 8874 | ARHGEF7 | Rho guanine nucleotide exchange factor (GEF) 7 | 1 | −1.2069 | 0.6717 |
| 309 | ANXA6 | annexin A6 | 1 | −1.9168 | 0.6821 |
| 5784 | PTPN14 | protein tyrosine phosphatase, non-receptor type 14 | 2 | −1.2144 | 0.6888 |
| 100534599 | ISY1-RAB43 | ISY1-RAB43 readthrough | 1 | −2.4556 | 0.6928 |
| 54431 | DNAJC10 | DnaJ (Hsp40) homolog, subfamily C, member 10 | 2 | −1.6506 | 0.7051 |
| 63874 | ABHD4 | abhydrolase domain containing 4 | 1 | −1.6850 | 0.7071 |
| 196 | AHR | aryl hydrocarbon receptor | 1 | −1.1288 | 0.7219 |
| 63897 | HEATR6 | HEAT repeat containing 6 | 1 | −1.0711 | 0.7291 |
| 10961 | ERP29 | endoplasmic reticulum protein 29 | 1 | −1.0578 | 0.7355 |
| 126626 | GABPB2 | GA binding protein transcription factor, beta subunit 2 | 2 | −1.1151 | 0.7488 |
| 79794 | C12orf49 | chromosome 12 open reading frame 49 | 1 | −1.7522 | 0.7864 |
| 5965 | RECQL | RecQ helicase-like | 3 | −1.2203 | 0.7885 |
| 64651 | CSRNP1 | cysteine-serine-rich nuclear protein 1 | 1 | −2.1079 | 0.8036 |
| 81558 | FAM117A | family with sequence similarity 117, member A | 1 | −2.0458 | 0.8054 |
| 7706 | TRIM25 | tripartite motif containing 25 | 2 | −1.2522 | 0.8360 |
| 55339 | WDR33 | WD repeat domain 33 | 1 | −1.5940 | 0.8369 |
| 10097 | ACTR2 | ARP2 actin-related protein 2 homolog (yeast) | 1 | −1.0537 | 0.8586 |
| 23348 | DOCK9 | dedicator of cytokinesis 9 | 1 | −1.2887 | 0.8621 |
| 10079 | ATP9A | ATPase, class II, type 9A | 1 | −2.2117 | 0.8625 |
| 9497 | SLC4A7 | solute carrier family 4, sodium bicarbonate cotransporter, member 7 | 1 | −1.1702 | 0.9121 |
| 54832 | VPS13C | vacuolar protein sorting 13 homolog C (S. cerevisiae) | 2 | −1.1051 | 0.9176 |
| 23433 | RHOQ | ras homolog family member Q | 1 | −1.6436 | 0.9319 |
| 55727 | BTBD7 | BTB (POZ) domain containing 7 | 1 | −1.2974 | 0.9480 |
| 11260 | XPOT | exportin, tRNA | 1 | −1.4398 | 0.9544 |
| 1362 | CPD | carboxypeptidase D | 2 | −1.3036 | 0.9645 |
| 151887 | CCDC80 | coiled-coil domain containing 80 | 2 | −2.1921 | 0.9667 |
| 116496 | FAM129A | family with sequence similarity 129, member A | 1 | −1.8099 | 0.9903 |
| 9709 | HERPUD1 | homocysteine-inducible, endoplasmic reticulum stress-inducible, ubiquitin-like domain member 1 | 1 | −3.3737 | 0.0854* |
| 284723 | SLC25A34 | solute carrier family 25, member 34 | 1 | −1.5051 | 0.0757* |
| 83641 | FAM107B | family with sequence similarity 107, member B | 1 | −2.2173 | 0.0749* |
| 60412 | EXOC4 | exocyst complex component 4 | 1 | −1.0957 | 0.0691* |
| 6700 | SPRR2A | small proline-rich protein 2A | 1 | −2.1995 | 0.0568* |
| 10365 | KLF2 | Kruppel-like factor 2 | 1 | −1.3378 | 0.0386* |
| 3572 | IL6ST | interleukin 6 signal transducer | 1 | −1.3786 | 0.0330* |
| 10551 | AGR2 | anterior gradient 2 | 1 | −3.7365 | 0.0134* |
| 9663 | LPIN2 | lipin 2 | 1 | −2.6332 | 0.0033* |
| 155435 | RBM33 | RNA binding motif protein 33 | 1 | −1.3082 | 0.0029* |
| 54855 | FAM46C | family with sequence similarity 46, member C | 2 | −1.0118 | 0.0028* |
| 728661 | SLC35E2B | solute carrier family 35, member E2B | 1 | −1.0811 | 0.0004* |
| 23591 | FAM215A | family with sequence similarity 215, member A (non-protein coding) | 1 | −1.4327 | N/A |
| 643707 | GOLGA6L4 | golgin A6 family-like 4 | 2 | −1.3773 | N/A |
| Age | Sex | Location | T | N | M | Stage | Differentiation | |
|---|---|---|---|---|---|---|---|---|
| A | 80 | M | larynx | 3 | 2c | 0 | IVa | moderate |
| B | 73 | M | larynx | 3 | 0 | 0 | III | poor |
| C | 77 | M | oral | 2 | 2b | 0 | Iva | moderate |
| D | 42 | F | oral | 4a | 0 | 0 | IVa | poor |
| E | 51 | M | oral | 2 | 0 | 0 | II | well |
| F | 52 | F | oral | 4a | 2c | 1 | Ivc | well |
| G | 72 | M | hypopharynx | 2 | 0 | 0 | II | moderate |
| H | 64 | M | hypopharynx | 2 | 2b | 0 | IVa | well |
| I | 70 | M | hypopharynx | 2 | 2b | 0 | Iva | well |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okada, R.; Koshizuka, K.; Yamada, Y.; Moriya, S.; Kikkawa, N.; Kinoshita, T.; Hanazawa, T.; Seki, N. Regulation of Oncogenic Targets by miR-99a-3p (Passenger Strand of miR-99a-Duplex) in Head and Neck Squamous Cell Carcinoma. Cells 2019, 8, 1535. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8121535
Okada R, Koshizuka K, Yamada Y, Moriya S, Kikkawa N, Kinoshita T, Hanazawa T, Seki N. Regulation of Oncogenic Targets by miR-99a-3p (Passenger Strand of miR-99a-Duplex) in Head and Neck Squamous Cell Carcinoma. Cells. 2019; 8(12):1535. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8121535
Chicago/Turabian StyleOkada, Reona, Keiichi Koshizuka, Yasutaka Yamada, Shogo Moriya, Naoko Kikkawa, Takashi Kinoshita, Toyoyuki Hanazawa, and Naohiko Seki. 2019. "Regulation of Oncogenic Targets by miR-99a-3p (Passenger Strand of miR-99a-Duplex) in Head and Neck Squamous Cell Carcinoma" Cells 8, no. 12: 1535. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8121535