Comparative Chromosome Mapping of Musk Ox and the X Chromosome among Some Bovidae Species

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Material and Methods
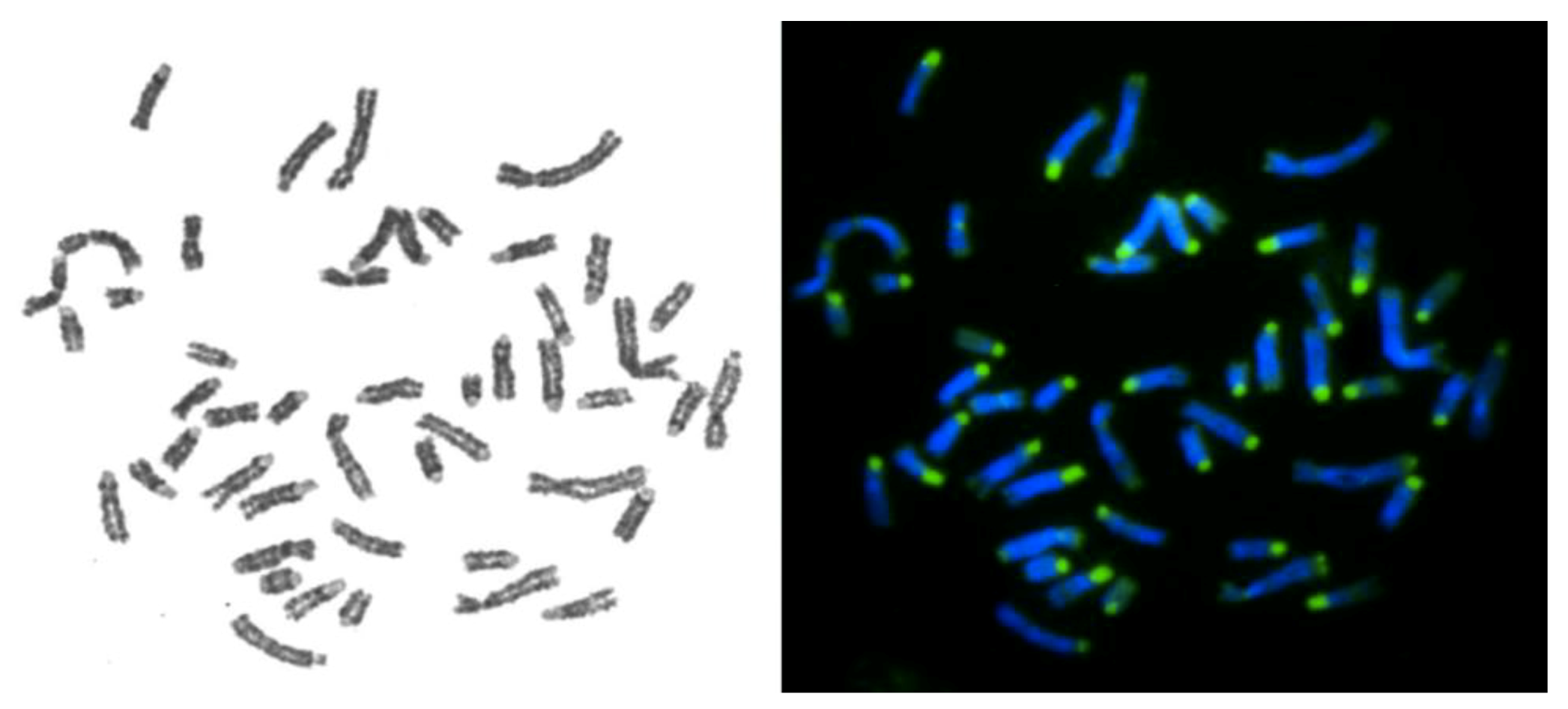
2.1. Species
2.2. Chromosome Preparation
2.3. FISH Probes
2.4. FISH Procedure
3. Results
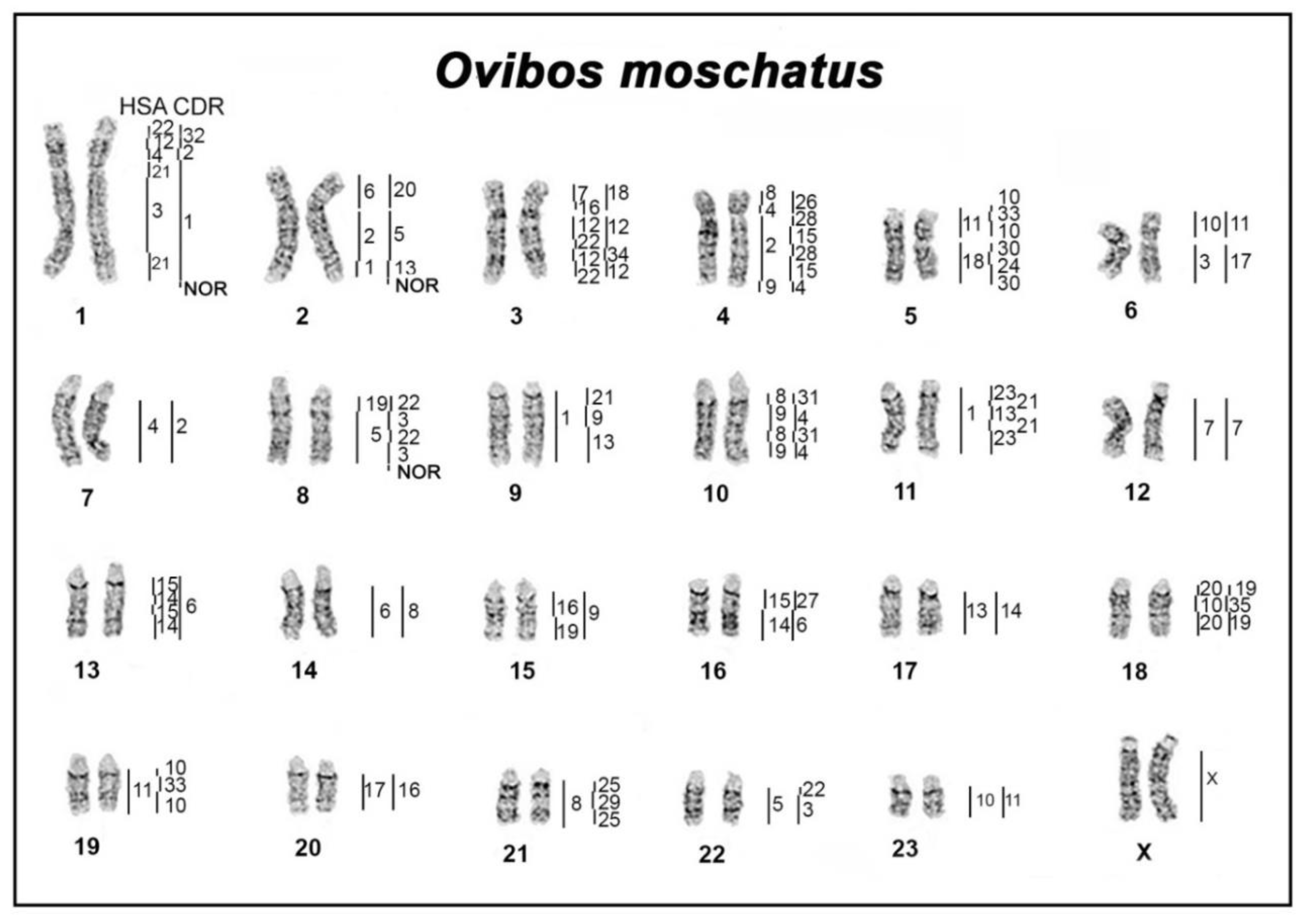
3.1. Comparative Chromosome Map of Musk Ox, Dromedary, and Human
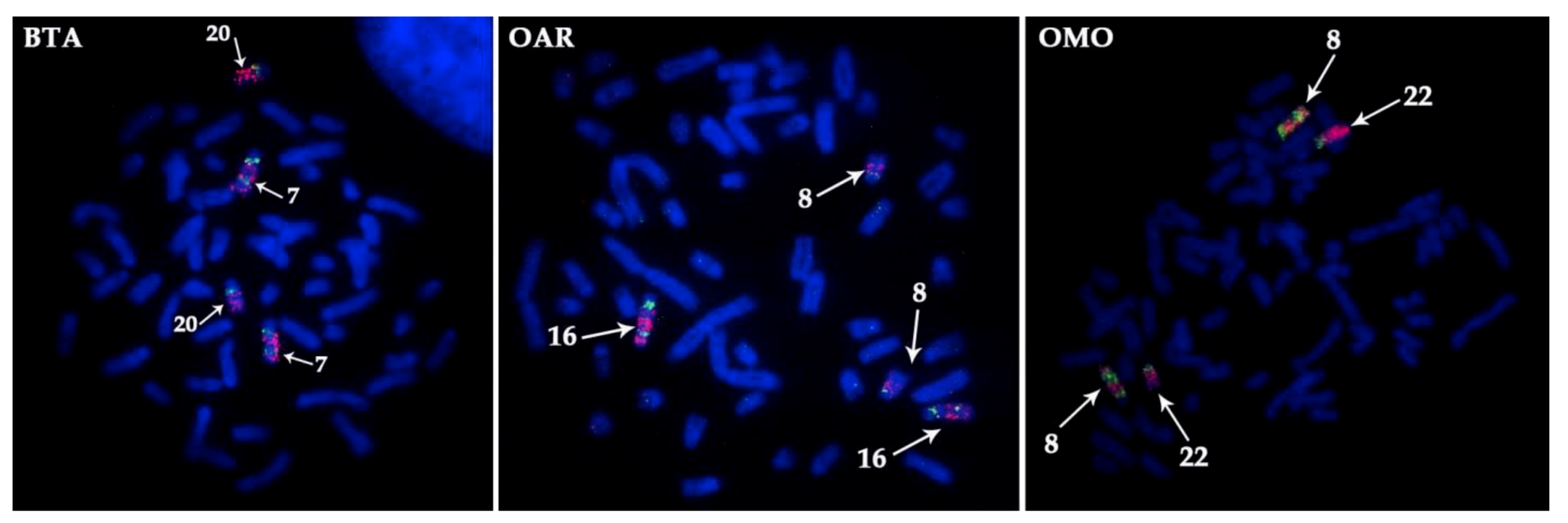
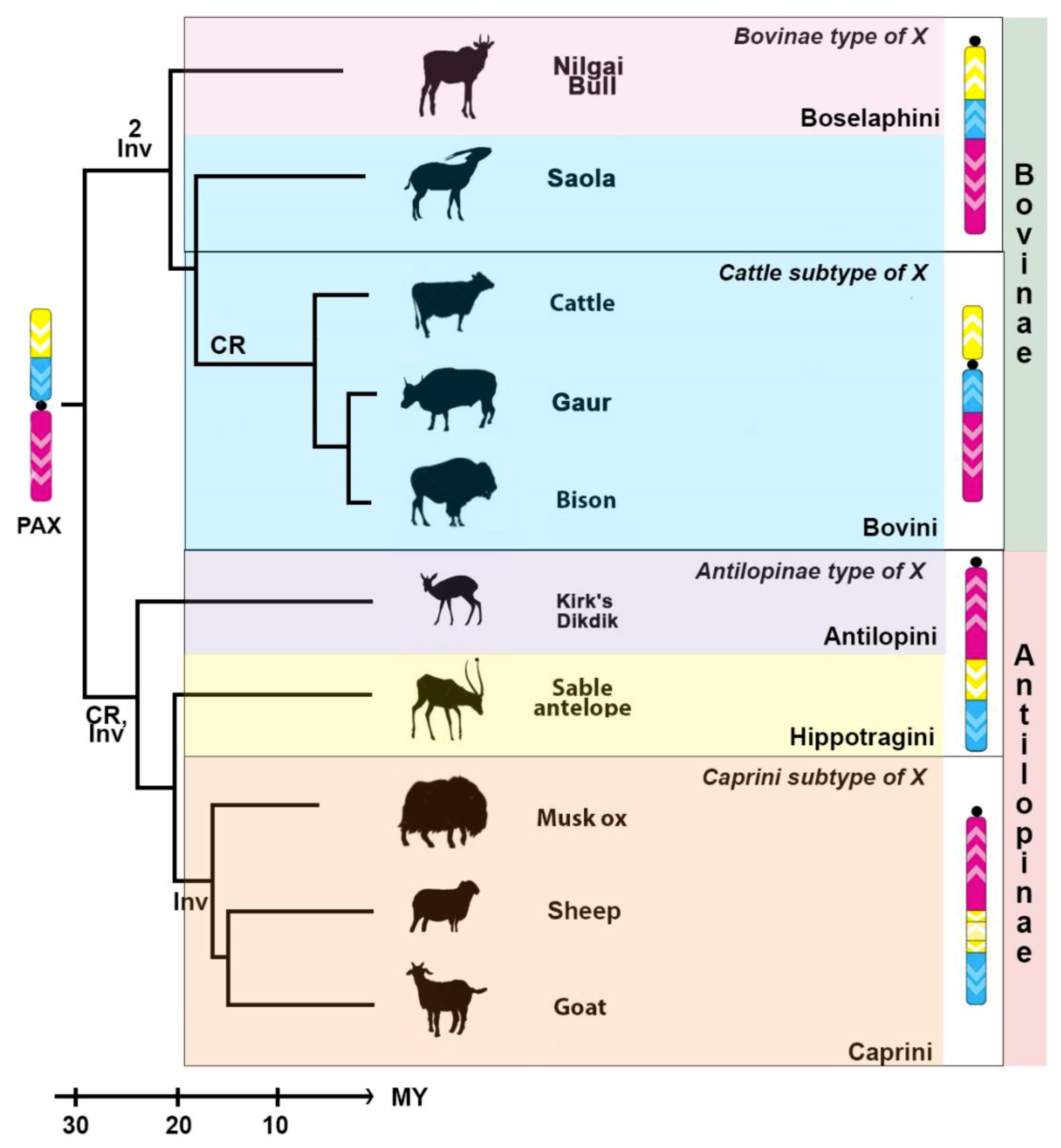
3.2. Mapping of the X Chromosome in Bovidae
4. Discussion
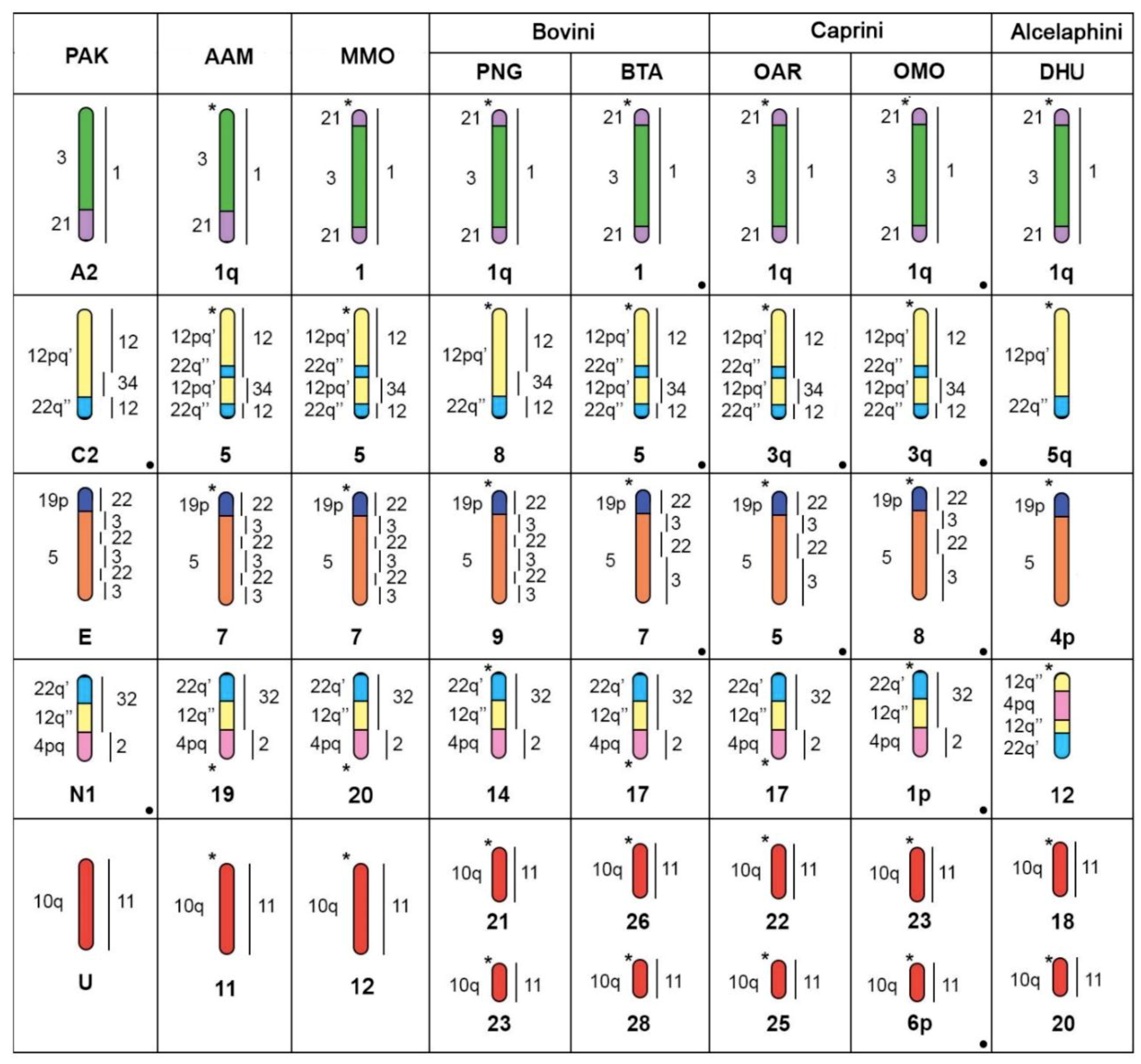
4.1. Evolution of Musk Ox and Bovid Karyotypes
4.2. Bovine X Chromosome Evolution
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wilson, D.E.; Reeder, D.M. Mammal Species of The World: A Taxonomic and Geographic Reference; JHU Press: Baltimore, MD, USA, 2005; Volume 1. [Google Scholar]
- Robinson, T.J.; Ropiquet, A. Examination of hemiplasy, homoplasy and phylogenetic discordance in chromosomal evolution of the Bovidae. Syst. Biol. 2011, 60, 439–450. [Google Scholar] [CrossRef]
- Zurano, J.P.; Magalhães, F.M.; Asato, A.E.; Silva, G.; Bidau, C.J.; Mesquita, D.O.; Costa, G.C. Cetartiodactyla: Updating a time-calibrated molecular phylogeny. Mol. Phylogenet. Evol. 2019, 133, 256–262. [Google Scholar] [CrossRef]
- Chi, J.X.; Huang, L.; Nie, W.; Wang, J.; Su, B.; Yang, F. Defining the orientation of the tandem fusions that occurred during the evolution of Indian muntjac chromosomes by BAC mapping. Chromosoma 2005, 114, 167–172. [Google Scholar] [CrossRef]
- Kulemzina, A.I.; Perelman, P.L.; Grafodatskaya, D.A.; Nguyen, T.T.; Thompson, M.; Roelke-Parker, M.E.; Graphodatsky, A.S. Comparative chromosome painting of pronghorn (Antilocapra americana) and saola (Pseudoryx nghetinhensis) karyotypes with human and dromedary camel probes. BMC Genet. 2014, 15, 68. [Google Scholar] [CrossRef]
- Balmus, G.; Trifonov, V.A.; Biltueva, L.S.; O’Brien, P.C.; Alkalaeva, E.S.; Fu, B.; Skidmore, J.A.; Allen, T.; Graphodatsky, A.S.; Yang, F.; et al. Cross-species chromosome painting among camel, cattle, pig and human: Further insights into the putative Cetartiodactyla ancestral karyotype. Chromosome Res. 2007, 15, 499–514. [Google Scholar] [CrossRef] [PubMed]
- Iannuzzi, L.; Di Meo, G.P.; Perucatti, A.; Schibler, L.; Incarnato, D.; Cribiu, E.P. Comparative FISH mapping in river buffalo and sheep chromosomes: Assignment of forty autosomal type I loci from sixteen human chromosomes. Cytogenet. Genome Res. 2001, 94, 43–48. [Google Scholar] [CrossRef]
- Rubes, J.; Kubickova, S.; Pagacova, E.; Cernohorska, H.; Di Berardino, D.; Antoninova, M.; Vahala, J.; Robinson, T.J. Phylogenomic study of spiral-horned antelope by cross-species chromosome painting. Chromosome Res. 2008, 16, 935–947. [Google Scholar] [CrossRef] [PubMed]
- Cernohorska, H.; Kubickova, S.; Vahala, J.; Robinson, T.J.; Rubes, J. Cytotypes of Kirk’s dik-dik (Madoqua kirkii, Bovidae) show multiple tandem fusions. Cytogenet. Genome Res. 2011, 132, 255–263. [Google Scholar] [CrossRef]
- Iannuzzi, L.; Di Meo, G.P.; Perucatti, A.; Bardaro, T. ZOO-FISH and R-banding reveal extensive conservation of human chromosome regions in euchromatic regions of river buffalo chromosomes. Cytogenet. Genome Res. 1998, 82, 210–214. [Google Scholar] [CrossRef]
- Chaves, R.; Frönicke, L.; Guedes-Pinto, H.; Wienberg, J. Multidirectional chromosome painting between the Hirola antelope (Damaliscus hunteri, Alcelaphini, Bovidae), sheep and human. Chromosome Res. 2004, 12, 495–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buckland, R.A.; Evans, H.J. Cytogenetic aspects of phylogeny in the Bovidae. Cytogenet. Genome Res. 1978, 21, 42–63. [Google Scholar] [CrossRef] [PubMed]
- Pagacova, E.; Cernohorska, H.; Kubickova, S.; Vahala, J.; Rubes, J. Centric fusion polymorphism in captive animals of family Bovidae. Conserv. Genet. 2011, 12, 71–77. [Google Scholar] [CrossRef]
- Rubes, J.; Musilova, P.; Kopecna, O.; Kubickova, S.; Cernohorska, H.; Kulemsina, A.I. Comparative molecular cytogenetics in Cetartiodactyla. Cytogenet. Genome Res. 2012, 137, 194–207. [Google Scholar] [CrossRef]
- Proskuryakova, A.A.; Kulemzina, A.I.; Perelman, P.L.; Makunin, A.I.; Larkin, D.M.; Farré, M.; Kukekova, A.V.; Johnson, J.L.; Lemskaya, N.A.; Beklemisheva, V.R.; et al. X Chromosome Evolution in Cetartiodactyla. Genes 2017, 8, 216. [Google Scholar] [CrossRef]
- Farré, M.; Kim, J.; Proskuryakova, A.A.; Zhang, Y.; Kulemzina, A.I.; Li, Q.; Zhou, Y.; Xiong, Y.; Johnson, J.L.; Perelman, P.L. Evolution of gene regulation in ruminants differs between evolutionary breakpoint regions and homologous synteny blocks. Genome Res. 2019, 29, 576–589. [Google Scholar] [CrossRef] [Green Version]
- Slate, J.; Van Stijn, T.C.; Anderson, R.M.; McEwan, K.M.; Maqbool, N.J.; Mathias, H.C.; Bixley, M.J.; Stevens, D.R.; Molenaar, A.J.; Beever, J.E.; et al. A deer (subfamily Cervinae) genetic linkage map and the evolution of ruminant genomes. Genetics 2002, 160, 1587–1597. [Google Scholar] [PubMed]
- Gallagher, D.S., Jr.; Womack, J.E. Chromosome conservation in the Bovidae. J. Hered. 1992, 83, 287–298. [Google Scholar] [CrossRef]
- Ohno, S.; Beçak, W.; Beçak, M.L. X-autosome ratio and the behavior pattern of individual X-chromosomes in placental mammals. Chromosoma 1964, 15, 14–30. [Google Scholar] [CrossRef]
- Robinson, T.J.; Harrison, W.R.; Ponce de Leon, F.A.; Davis, S.K.; Elder, F.F.B. A molecular cytogenetic analysis of X chromosome repatterning in the Bovidae: Transpositions, inversions, and phylogenetic inference. Cytogenet. Genome Res. 1998, 80, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Iannuzzi, L.; King, W.A.; Di Berardino, D. Chromosome evolution in domestic bovids as revealed by chromosome banding and FISH-mapping techniques. Cytogenet. Genome Res. 2009, 126, 49–62. [Google Scholar] [CrossRef]
- Cernohorska, H.; Kubickova, S.; Vahala, J.; Rubes, J. Molecular insights into X; BTA5 chromosome rearrangements in the tribe Antilopini (Bovidae). Cytogenet. Genome Res. 2012, 136, 188–198. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, D.S.; Davis, S.K.; De Donato, M.; Burzlaff, J.D.; Womack, J.E.; Taylor, J.F.; Kumamoto, A.T. A karyotypic analysis of nilgai, Boselaphus tragocamelus (Artiodactyla: Bovidae). Chromosome Res. 1998, 6, 505–514. [Google Scholar] [CrossRef]
- Lent, P.C. Ovibos moschatus. Mamm. Species 1988, 302, 1–9. [Google Scholar] [CrossRef]
- Biltueva, L.S.; Sharshov, A.A.; Graphodatsky, A.S. G-Banding Homologies in Musk Ox, Ovibos moschatus, and other Bovids. Hereditas 1995, 122, 185–187. [Google Scholar] [CrossRef]
- Pasitschniak-Arts, M.; Flood, P.F.; Schmutz, S.M.; Seidel, B. A Comparison of G-Band Patterns of The Muskox and Takin and Their Evolutionary Relationship to Sheep; Oxford University Press: Oxford, UK, 1994. [Google Scholar]
- Desaulniers, D.M.; King, W.A.; Rowell, J.E.; Flood, P.F. The banded chromosomes of the muskox (Ovibos moschatus). Can. J. Zool. 1989, 67, 1155–1158. [Google Scholar] [CrossRef]
- Huang, L.; Nie, W.; Wang, J.; Su, W.; Yang, F. Phylogenomic study of the subfamily Caprinae by cross-species chromosome painting with Chinese muntjac paints. Chromosome Res. 2005, 13, 389–399. [Google Scholar] [CrossRef]
- Kulemzina, A.I.; Yang, F.; Trifonov, V.A.; Ryder, O.A.; Ferguson-Smith, M.A.; Graphodatsky, A.S. Chromosome painting in Tragulidae facilitates the reconstruction of Ruminantia ancestral karyotype. Chromosome Res. 2011, 19, 531. [Google Scholar] [CrossRef]
- Kulemzina, A.I.; Trifonov, V.A.; Perelman, P.L.; Rubtsova, N.V.; Volobuev, V.; Ferguson-Smith, M.A.; Stanyon, R.; Yang, F.; Graphodatsky, A.S. Cross-species chromosome painting in Cetartiodactyla: Reconstructing the karyotype evolution in key phylogenetic lineages. Chromosome Res. 2009, 17, 419–436. [Google Scholar] [CrossRef]
- Yang, F.; O’Brien, P.C.M.; Milne, B.S.; Graphodatsky, A.S.; Solanky, N.; Trifonov, V.; Rens, W.; Sargan, D.; Ferguson-Smith, M.A. A complete comparative chromosome map for the dog, red fox, and human and its integration with canine genetic maps. Genomics 1999, 62, 189–202. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Graphodatsky, A.S. Animal probes and ZOO-FISH. In Fluorescence In Situ Hybridization (FISH); Springer: Berlin/Heidelberg, Germany, 2017; pp. 323–346. [Google Scholar]
- Seabright, M. A rapid banding technique for human chromosomes. The Lancet 1971, 2, 971–972. [Google Scholar] [CrossRef]
- Lemskaya, N.A.; Kulemzina, A.I.; Beklemisheva, V.R.; Biltueva, L.S.; Proskuryakova, A.A.; Perelman, P.L.; Graphodatsky, A.S. The combined method of heterogeneous heterochromatin detection (CDAG) in different mammalian species. Chromosoma 2018, 26, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Proskuryakova, A.; Kulemzina, A.; Perelman, P.; Serdukova, N.; Ryder, O.; Graphodatsky, A. The Case of X and Y Localization of Nucleolus Organizer Regions (NORs) in Tragulus javanicus (Cetartiodactyla, Mammalia). Genes 2018, 9, 312. [Google Scholar] [CrossRef] [PubMed]
- Ijdo, J.W.; Wells, R.A.; Baldini, A.; Reeders, S.T. Improved telomere detection using a telomere repeat probe (TTAGGG) n generated by PCR. Nucleic Acids Res. 1991, 19, 4780. [Google Scholar] [CrossRef]
- Telenius, H.; Ponder, B.A.; Tunnacliffe, A.; Pelmear, A.H.; Carter, N.P.; Ferguson-Smith, M.A.; Behmel, A.; Nordenskjöld, M.; Pfragner, R. Cytogenetic analysis by chromosome painting using DOP-PCR amplified flow-sorted chromosomes. Genes Chromosomes Cancer 1992, 4, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Berrios del Solar, M.S.; Manieu Seguel, C.; López Fenner, J.; Ayarza Ramírez, G.; Page, J.; González Vergara, M.; Manterola Zúñiga, M.; Fernández Donoso, R. Robertsonian chromosomes and the nuclear architecture of mouse meiotic prophase spermatocytes. Biol. Res. 2014, 47, 16. [Google Scholar] [CrossRef]
- De Gortari, M.J.; Freking, B.A.; Cuthbertson, R.P.; Kappes, S.M.; Keele, J.W.; Stone, R.T.; Leymaster, K.A.; Dodds, K.G.; Crawford, A.M.; Beattie, C.W. A second-generation linkage map of the sheep genome. Mamm. Genome 1998, 9, 204–209. [Google Scholar]
- Hassanin, A.; Delsuc, F.; Ropiquet, A.; Hammer, C.; van Vuuren, B.J.; Matthee, C.; Ruiz-Garcia, M.; Catzeflis, F.; Areskoug, V.; Nguyen, T.T.; et al. Pattern and timing of diversification of Cetartiodactyla (Mammalia, Laurasiatheria), as revealed by a comprehensive analysis of mitochondrial genomes. C. R. Biol. 2012, 335, 32–50. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Müller, S.; Just, R.; Ferguson-Smith, M.A.; Wienberg, J. Comparative chromosome painting in mammals: Human and the Indian muntjac (Muntiacus muntjak vaginalis). Genomics 1997, 39, 396–401. [Google Scholar] [CrossRef]
- Frohlich, J.; Kubickova, S.; Musilova, P.; Cernohorska, H.; Muskova, H.; Vodicka, R.; Rubes, J. Karyotype relationships among selected deer species and cattle revealed by bovine FISH probes. PLoS ONE 2017, 12, e0187559. [Google Scholar] [CrossRef]
- Kulemzina, A.I.; Proskuryakova, A.A.; Beklemisheva, V.R.; Lemskaya, N.A.; Perelman, P.L.; Graphodatsky, A.S. Comparative Chromosome Map and Heterochromatin Features of the Gray Whale Karyotype (Cetacea). Cytogenet. Genome Res. 2016, 148, 25–34. [Google Scholar] [CrossRef]
- Iannuzzi, L.; Di Meo, G.P.; Perucatti, A.; Incarnato, D.; Schibler, L.; Cribiu, E.P. Comparative FISH mapping of bovid X chromosomes reveals homologies and divergences between the subfamilies Bovinae and Caprinae. Cytogenet. Genome Res. 2000, 89, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Vozdova, M.; Ruiz-Herrera, A.; Fernandez, J.; Cernohorska, H.; Frohlich, J.; Sebestova, H.; Kubickova, S.; Rubes, J. Meiotic behaviour of evolutionary sex-autosome translocations in Bovidae. Chromosome Res. 2016, 24, 325–338. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, S.J.; Menninger, J.C.; Nash, W.G. Atlas of Mammalian Chromosomes; John Wiley & Sons: Hoboken, NJ, USA, 2006. [Google Scholar]
- Rieseberg, L.H. Chromosomal rearrangements and speciation. Trends Ecol. Evol. 2001, 16, 351–358. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scientific Name, Abbreviation | Common Name | Subfamily | Diploid Number | Sample/Cell Line Source/Acknowledgment |
|---|---|---|---|---|
| Ovibos moschatus, OMO | Musk ox | Antilopinae (Caprini) | 48, XX | Allaikhovsky District, Sakha Republic, Yakutia, Russia. IMCB SB RAS, Novosibirsk |
| Ovis aries musimon, OAR | Sheep | Antilopinae (Caprini) | 54, XX | Melody Roelke and June Bellizzi, Catoctin Wildlife Preserve and Zoo, Maryland, USA; Laboratory of Genomic Diversity, NCI-Frederick, MD, USA |
| Madoqua kirkii, MKI | Kirk’s Dikdik | Antilopinae (Antilopini) | 48, XY | Mitchell Bush, Conservation and Research Center, National Zoological Park, Virginia, USA; Laboratory of Genomic Diversity, NCI-Frederick, MD, USA |
| Bos taurus, BTA | Cattle | Bovinae (Bovini) | 60, XX | IMCB SB RAS, Novosibirsk. |
| Bos gaurus, BGA | Gaur | Bovinae (Bovini) | 58, XX | Doug Armstrong, Henry Doorly Zoo, Omaha, NE, USA; Laboratory of Genomic Diversity, NCI-Frederick, MD, USA |
| Pseudoryx nghetinhensis, PNG | Saola | Bovinae (Bovini) | 50, XX | [5] |
| Boselaphus tragocamelus, BTR | Nilgai bull | Bovinae (Boselaphini) | 44, X+14, X+14 | Melody Roelke and June Bellizzi, Catoctin Wildlife Preserve and Zoo, Maryland, USA; Laboratory of Genomic Diversity, NCI-Frederick, MD, USA |
| OMO | HSA | CDR | BTA | PAK |
|---|---|---|---|---|
| 1p | 22q’/12q”/4pq | 32/2 | 17 | N1 |
| 1q | 21/3/21 | 1 | 1 | A2 |
| 2p | 6p | 20 | 23 | R |
| 2q | 2q”/1 | 5/13 | 2 | B2 |
| 3p | 16p/7 | 18 | 25 | T |
| 3q | 12pq’/22q”/12pq’/22q” | 12/34/12 | 5 | C2 |
| 4p | 4/8p” | 26 | 27 | V |
| 4q | 2pq/9 | 28/15/28/15/4 | 11 | C1 |
| 5p | 11 | 10/33/10 | 29 | W |
| 5q | 18 | 30/24/30 | 24 | S |
| 6p | 10q | 11 | 28 | U |
| 6q | 3 | 17 | 22 | Q |
| 7 | 4pq | 2 | 6 | F |
| 8 | 19p/5 | 22/3/22/3 | 7 | E |
| 9 | 1 | 21/9/13 | 3 | A1 |
| 10 | 8p’/9/8p’/9 | 31/4/31/4 | 8 | B1 |
| 11 | 1 | 23/21/13/21/23 | 16 | K |
| 12 | 7 | 7 | 4 | D |
| 13 | 15/14/15/14 | 6 | 10 | G |
| 14 | 6q | 8 | 9 | H1 |
| 15 | 16q/19p | 9 | 18 | M |
| 16 | 15/14 | 27/6 | 21 | P |
| 17 | 13 | 14 | 12 | I |
| 18 | 20/10p/20 | 19/35/19 | 13 | J |
| 19 | 11 | 10/33/10 | 15 | L |
| 20 | 17 | 16 | 19 | N2 |
| 21 | 8q | 25/29/25 | 14 | H2 |
| 22 | 5 | 22/3 | 20 | O |
| 23 | 10q | 11 | 26 | U |
| X | X | X | X | X |
| Syntenic Block | X BAC’s Order in Bovinae Subfamily | X BAC’s Order in Antilopinae Subfamily | ||
|---|---|---|---|---|
| In Most Bovinae | In Saola | Caprini Tribe | Hippotragini and Antilopini Tribe | |
| X syntenic block 1 (XSB1) | CH240-514O22 | CH240-514O22 | CH240-66H2 | CH240-66H2 |
| CH240-287O21 | CH240-287O21 | CH240-155A13 | CH240-155A13 | |
| CH240-128C9 | CH240-128C9 CH240-106A3 | CH240-90L14 | CH240-90L14 | |
| CH240-106A3 | CH240-373L23 | CH240-373L23 | ||
| CH240-229I15 | CH240-229I15 | CH240-62M10 | CH240-62M10 | |
| CH240-103E10 | CH240-103E10 | CH240-122P17 | CH240-122P17 | |
| CH240-386M8 | CH240-386M8 | CH240-252G15 | CH240-252G15 | |
| X syntenic block 2 (XSB2) | CH240-108D16 | CH240-108D16 | CH240-375C5 | CH240-375C5 |
| CH240-54D24 | CH240-54D24 | CH240-130I15 | CH240-130I15 | |
| CH240-93K24 | CH240-93K24 | CH240-118P13 | CH240-118P13 | |
| CH240-108D16 | ||||
| CH240-122N13 | CH240-122N13 | CH240-25P8 | CH240-25P8 | |
| CH240-195J23 | CH240-195J23 | CH240-14O10 | CH240-14O10 | |
| CH240-316D2 | CH240-316D2 | CH240-214A3 | CH240-214A3 | |
| X syntenic block 3 (XSB3) | CH240-214A3 | CH240-214A3 | CH240-386M8 | CH240-386M8 |
| CH240-14O10 | CH240-14O10 | CH240-103E10 | CH240-103E10 | |
| CH240-25P8 | CH240-25P8 | CH240-128C9 | CH240-229I15 | |
| CH240-118P13 | CH240-118P13 | CH240-106A3 | CH240-106A3 | |
| CH240-130I15 | CH240-130I15 | CH240-229I15 | CH240-128C9 | |
| CH240-375C5 | CH240-375C5 | CH240-287O21 | CH240-287O21 | |
| CH240-252G15 | CH240-252G15 CH240-122P17 | CH240-514O22 | CH240-514O22 | |
| CH240-122P17 | CH240-316D2 | CH240-316D2 | ||
| CH240-62M10 | CH240-62M10 | CH240-195J23 | CH240-195J23 | |
| CH240-373L23 | CH240-373L23 | CH240-122N13 | CH240-122N13 | |
| CH240-90L14 | CH240-90L14 | CH240-93K2 | CH240-93K2 | |
| CH240-155A13 | CH240-155A13 | CH240-54D24 | CH240-54D24 | |
| CH240-66H2 | CH240-66H2 | CH240-108D16 | CH240-108D16 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Proskuryakova, A.A.; Kulemzina, A.I.; Perelman, P.L.; Yudkin, D.V.; Lemskaya, N.A.; Okhlopkov, I.M.; Kirillin, E.V.; Farré, M.; Larkin, D.M.; Roelke-Parker, M.E.; et al. Comparative Chromosome Mapping of Musk Ox and the X Chromosome among Some Bovidae Species. Genes 2019, 10, 857. https://0-doi-org.brum.beds.ac.uk/10.3390/genes10110857
Proskuryakova AA, Kulemzina AI, Perelman PL, Yudkin DV, Lemskaya NA, Okhlopkov IM, Kirillin EV, Farré M, Larkin DM, Roelke-Parker ME, et al. Comparative Chromosome Mapping of Musk Ox and the X Chromosome among Some Bovidae Species. Genes. 2019; 10(11):857. https://0-doi-org.brum.beds.ac.uk/10.3390/genes10110857
Chicago/Turabian StyleProskuryakova, Anastasia A., Anastasia I. Kulemzina, Polina L. Perelman, Dmitry V. Yudkin, Natalya A. Lemskaya, Innokentii M. Okhlopkov, Egor V. Kirillin, Marta Farré, Denis M. Larkin, Melody E. Roelke-Parker, and et al. 2019. "Comparative Chromosome Mapping of Musk Ox and the X Chromosome among Some Bovidae Species" Genes 10, no. 11: 857. https://0-doi-org.brum.beds.ac.uk/10.3390/genes10110857