Roles of Topoisomerases in Heterochromatin, Aging, and Diseases

Lab of Genetics and Genomics, National Institute on Aging, National Institute of Health, Baltimore, MD 21224, USA
*
Author to whom correspondence should be addressed.
Genes 2019, 10(11), 884; https://0-doi-org.brum.beds.ac.uk/10.3390/genes10110884
Submission received: 3 October 2019
/
Revised: 23 October 2019
/
Accepted: 26 October 2019
/
Published: 1 November 2019
(This article belongs to the Special Issue DNA Topoisomerases in Biology and Medicine)
{kind=link}
{kind=link}
{kind=link}
Abstract
:Heterochromatin is a transcriptionally repressive chromatin architecture that has a low abundance of genes but an enrichment of transposons. Defects in heterochromatin can cause the de-repression of genes and transposons, leading to deleterious physiological changes such as aging, cancer, and neurological disorders. While the roles of topoisomerases in many DNA-based processes have been investigated and reviewed, their roles in heterochromatin formation and function are only beginning to be understood. In this review, we discuss recent findings on how topoisomerases can promote heterochromatin organization and impact the transcription of genes and transposons. We will focus on two topoisomerases: Top2α, which catenates and decatenates double-stranded DNA, and Top3β, which can change the topology of not only DNA, but also RNA. Both enzymes are required for normal heterochromatin formation and function, as the inactivation of either protein by genetic mutations or chemical inhibitors can result in defective heterochromatin formation and the de-silencing of transposons. These defects may contribute to the shortened lifespan and neurological disorders observed in individuals carrying mutations of Top3β. We propose that topological stress may be generated in both DNA and RNA during heterochromatin formation and function, which depend on multiple topoisomerases to resolve.
1. Introduction
Topoisomerases are essential enzymes that can resolve topological problems generated during DNA and RNA metabolism. They have a unique ability to catalyze transient cleavage and passage of single or double-strand DNA, and then, reseal the broken ends after the passage. This ability allows them to relax supercoiled DNA and decatenate inter-tangled DNA produced during transcription, replication, recombination, and chromosome condensation and segregation [1]. Topoisomerases can be categorized into two main types: Type I and II, which can cleave and re-ligate single and double stranded DNA, respectively. Each type of topoisomerases can be further divided into A or B depending on the mechanism of the strand passage. Mammals have two Type IA topoisomerases, Top3α, Top3β; two Type IB topoisomerases, Top1 in nucleus and Top1mt in mitochondria; and two Type IIA topoisomerases: Top2α and Top2β. Type IB topoisomerases can cleave one strand of DNA, allow one broken strand to rotate round the unbroken strand to remove all supercoils, and then re-ligate the broken ends. Type IIA and IIB can catalyze ATP-dependent dsDNA passage reactions, enabling catenation and decatenation of DNA.
Topoisomerases are essential for viability of many organisms. For example, Top1 is essential in multicellular organisms, whereas Top2 is essential in all organisms. Specifically, mouse and Drosophila lacking either Top1 or Top2 (α and β) exhibits lethality in early developmental stage [2,3,4]. While Top3β is not an essential gene in these species, mouse lacking Top3β displays shortened lifespan, higher incidence of aneuploidy in germ cells, increased autoimmunity [5,6,7], and abnormal synapse formation [8]. Importantly, topoisomerase mutations have been associated with various human diseases, such as cancer, neurodegeneration, autism, and autoimmune diseases [9,10,11,12]. Moreover, drugs targeting topoisomerases have been widely used in treatment of cancers [13]. Thus, understanding the mechanism of these enzymes may allow for development of drugs in treatment of more human disorders.
In eukaryotes, DNA wraps around histone octamers to form nucleosomes, which are then assembled into higher-order chromatin structures. Formation of the chromatin allows efficient storage of genetic information, but also restricts the access of DNA to machinery of transcription, replication, and other processes. Topoisomerases often play a positive role to make the chromatin DNA accessible for these processes. For example, during transcription initiation, topoisomerases can facilitate disassembly of the nucleosome-DNA structures and reduce histone density to enable transcription machinery to access promoters [14,15]. In addition, Top1 and Top2 can regulate histone modifications to generate a conducive chromatin environment for binding of transcription factors and Pol II to promoters [16,17]. Moreover, Top1 and Top2α have been shown to work with chromatin remodeling enzymes during transcription and genome stabilization [14,18,19].
Chromatin can be classified into two main domains: euchromatin, which is transcriptionally active and enriched with genes; and heterochromatin, which is transcriptionally inactive, has low abundance of genes but high abundance of transposons [20,21]. The distinction between heterochromatin and euchromatin can be characterized by different histone modifications. Euchromatin exhibits active histone marks including acetylation of H3K36 and H3K4 methylation, while heterochromatin is associated with repressive histone marks such as heavily methylated H3K9 or global hypoacetylation [22]. Upon modification of histones, key proteins for heterochromatin formation, such as Heterochromatin Protein 1 (HP1) or Histone methyltransferase (HMT), are recruited and a compact chromatin structure is formed. There are two types of heterochromatin: facultative and constitutive heterochromatin. Both types are transcriptionally repressed and exhibit high nucleosome density. Constitutive heterochromatin is generally formed at pericentric region, which consists of repetitive tandem repeats with more static structure. Facultative heterochromatin can be temporally and spatially decondensed and condensed within euchromatin region. Contrary to the constitutive heterochromatin that is generally marked by H3K9me3, facultative heterochromatin often associates with Polycomb group protein mediated H3K27me3 [23].
2. Importance of Heterochromatin
2.1. Heterochromatin Is Critical for Transcriptional Silencing of Transposons
A main function of heterochromatin is protecting the underlying genome from being accessed and transcribed by transcriptional machinery. This is important because the major components of heterochromatin are transposable elements (TEs) and tandem repetitive sequences. TEs are mobile genetic elements that can jump and randomly insert within a genome. They are present in all living organisms, and are major constituents of eukaryotic genome (more than 40% of human genome, 37% of mouse genome, and more than 80% of maize genome) [27,28,29]. TE and repeat integration or excision can generate chromosomal reorganization and other genetic alterations [30]. They can produce deleterious effects by landing in essential genes, causing mutations or mis-regulation. Consequently, TEs have been shown to be a potent mutagen that can cause genomic instability, aging, and various diseases, including cancers and neurological disorders [31,32,33]. To suppress their harmful effects, TEs and repeats are subjected to transcriptional and post-transcriptional silencing mechanisms [34,35,36]. Heterochromatin has a crucial role in suppressing TE and repeat expression through diverse mechanisms.
2.2. Loss of Heterochromatin May Be a Cause of Aging and Premature Aging Syndromes
Aging is one of the many well-studied physiological processes that has been associated with heterochromatin loss. In the ‘heterochromatin loss model of aging’ [37], it was proposed that heterochromatin is established in the early developmental stage and maintained through life. This epigenetic heterochromatin structure must be regenerated each time DNA is replicated or repaired. DNA damage and cell division may be the major perturbing factors triggering heterochromatin loss. This could result in alteration of age-related gene expression at the periphery of the heterochromatin domains. Indeed, many studies have shown that increasing heterochromatin formation can extend lifespan. For example, over-expression of heterochromatin protein HP1 in Drosophila increases the longevity [38]. In human, the loss of heterochromatin underlies two well-characterized progeroid syndromes: Werner syndrome (WS) and Hutchinson-Gilford Progeria Syndrome (HGPS) [39,40,41,42], Notably, the HGPS model exhibits up-regulation of satellite III repeats, suggesting that de-silencing of repeats in loss of heterochromatin may contribute to aging [40].
2.3. Heterochromatin Loss May Increase Cancer Risks
Loss of heterochromatin has been shown to impair chromosome segregation, nucleosome compaction, and DNA repair in multiple cancer cells [43,44]. In several cancer progression models, loss of HP1 or H3K9 methylation in heterochromatin has been shown to correlate with cancer progression and tumorigenesis [43,45,46]. One potential mechanism is that the reduction in heterochromatin may trigger mutagenic retro-transposition of satellite DNA and transposons. Transposition of these mobile elements may result in inactivation of tumor suppressor genes or activation of oncogenes. For example, highly active retro-transposition of a mammalian retrotransposon LINE-1 has been identified in multiple types of cancers, suggesting that heterochromatin mediated silencing of LINE-1 is disrupted [47]. It has been proposed that the insertion of LINE-1 in specific loci can disrupt tumor suppressors or oncogenes, resulting in tumorigenesis [48].
2.4. Heterochromatin and Neurological Disorders
Alteration in heterochromatin has been linked to several neurological disorders, including neurodegeneration-associated tauopathy (such as Alzheimer’s disease). Dysfunction of the hyperphosphorylated Tau protein is one of the main causes of tauopathy [49]. Drosophila tauopathy models, which transgenically express human tauR406W mutant protein, display neurodegenerative phenotype along with loss of heterochromatin, as shown by reduced H3K9 methylation and HP1a [50]. Moreover, disrupting heterochromatin structure enhances the neurodegeneration phenotype of flies that over-express tau, whereas enhancing heterochromatin formation reduces neurodegeneration, suggesting that heterochromatin loss has an important role in tau-induced neurodegeneration [50]. Indeed, several studies reported that brains of human Alzheimer’s disease patients exhibit loss of heterochromatin and de-silencing of genes [50,51]. Two other studies also show that a significant alteration of transposon expression in human and fly tauopathy brains, implicating that dysregulation of transposons in heterochromatin may cause neurotoxicity [52,53].
3. Multiple Topoisomerases Function in Heterochromatin
Heterochromatin is a stable but dynamic structure. Its formation and maintenance require numerous factors including: histone modification enzymes, chromatin assembly factors, heterochromatin associated structural proteins, chromatin remodeling enzymes [54,55], and several topoisomerases, as described below.
3.1. Top1
Top1 has been shown to be required for heterochromatin structure and histone modifications. An ultrastructural study showed that Top1 inhibitors, Camptothecin (CPT) and Rebeccamycin, can cause unpacking of heterochromatin in Trypanosoma cruzi [56]. Similarly, CPT treatment of human HCT116 cells induces decompression of heterochromatin with altered histone modifications [57]. There is also evidence that Top1 inactivation may disrupt a main function of heterochromatin, transcriptional silencing of transposons [58]. The mechanism of how Top1 inhibition leads to disruption of heterochromatin remains unclear. One study shows that depletion of Top1 in human HEK293 cells results in excessive RNA-DNA hybrid (R-loop) formation in heterochromatin domains, implicating a role of Top1 in regulating R-loop homeostasis in heterochromatin [59]. The R-loop is formed by nascent RNA entangled with template DNA during transcription; and it has been shown as a mediating structure during RNA-induced heterochromatin formation [60]. It is possible that Top1 inhibition may alter heterochromatin formation by disrupting R-loop homeostasis.
3.2. Top2
A fraction of Top2 has been detected in heterochromatin in Drosophila [61,62]. Subsequent studies revealed that Top2 is required for heterochromatin condensation, as Drosophila Top2 mutant cells have decondensed heterochromatin with increased aneuploidy and polyploidy [63]. In addition, Top2 depletion in Drosophila ovary results in poorly segregated heterochromatin, whereas euchromatin separation is normal during meiosis [64], suggesting that Top2 is specifically needed for segregation of heterochromatin region, which is resolved during cell cycle progression. Moreover, Top2 is needed for transcriptional silencing in heterochromatin, as displacement of Top2 from Satellite III in heterochromatin or chemical inhibition of Top2 activity can disrupt heterochromatic silencing of a reporter gene [65].
In studies from mammalian cell lines, Top2 is found to be required for chromosome condensation, segregation; and assembly of heterochromatin [66]. Silencing or chemical inhibition of Top2 induce chromatin reorganization and heterochromatin changes in mouse and human cell lines [67]. Moreover, Top2 inhibition by an inhibitor, etoposide, results in increased expression and mobility of repeats and transposons [68,69], consistent with the notion that Top2, like Top1, is also needed for heterochromatic silencing of transposons.
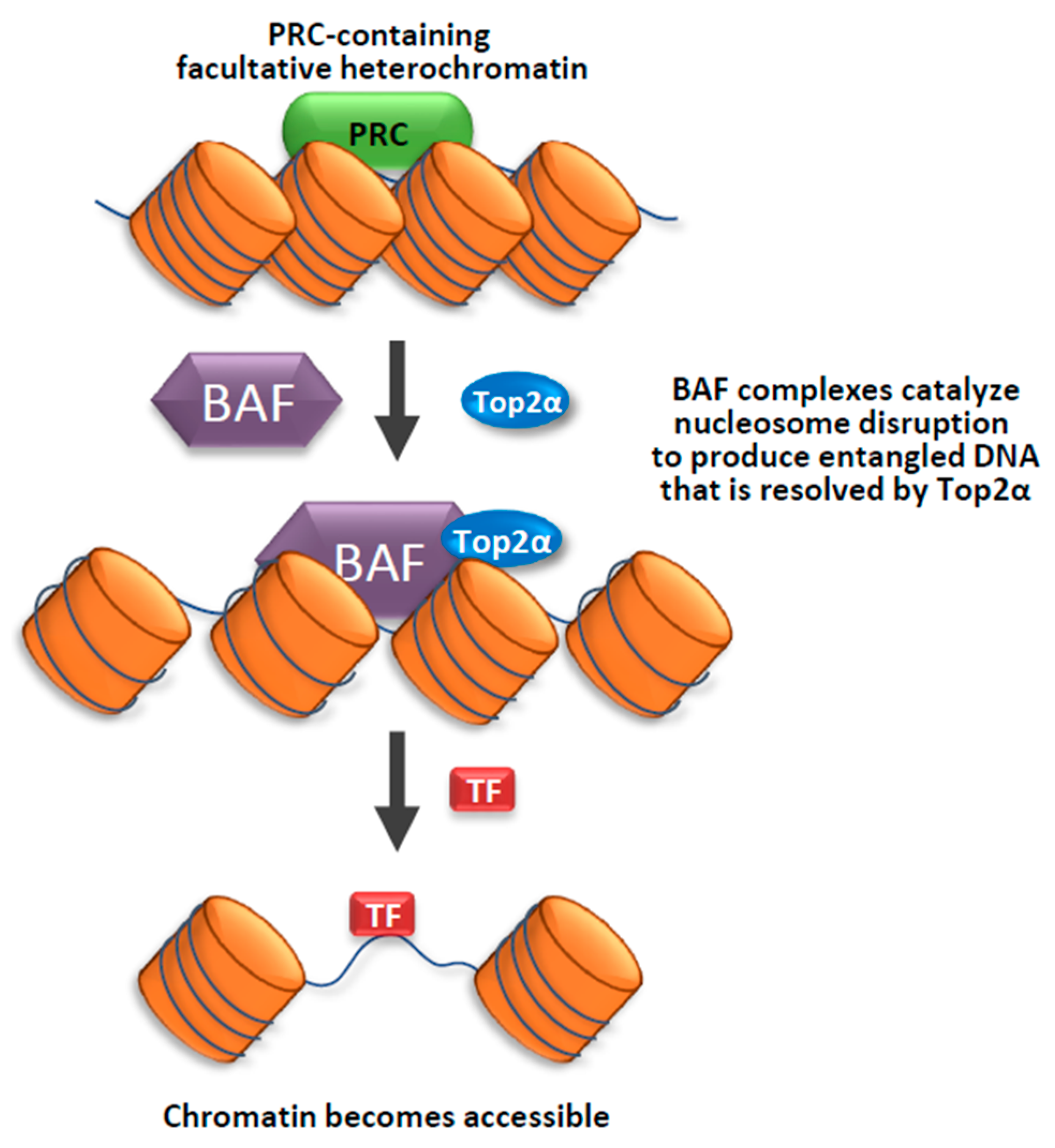
How does Top2 function in heterochromatin? Recent findings from the Crabtree lab suggest that Top2 can work with chromatin remodeling complexes, BAF, to make facultative heterochromatin more accessible to transcription factors [70] (Figure 1). The mammalian BAF complexes are ATP-dependent chromatin remodeling complexes that can alter chromatin structure to make the DNA more accessible to machinery of transcription, replication, repair, and other processes on DNA [71,72,73]. In an early study by Crabtree’s group, they found that BAF complexes can directly interact with Top2α to facilitates its access to approximately 12,000 sites across the genome [19]. They demonstrated that cells mutated in the BAF complex components are defective in decatenation of newly replicated sister chromatids, which is a phenotype of cells inhibited of Top2α activity. The findings that BAF complex is needed by Top2α to prevent DNA entanglement during mitosis provides an explanation for the observation that BAF subunits are mutated in a large fraction of human cancers [72,73]. In a later study, Crabtree’s group used an in vivo Chromatin indicator Assay (CiA), which can induce chromatin remodeling by recruiting a chromatin regulator to a designed locus upon Chemical Inducer of Proximity. They demonstrate that Top2α is required for an early process during BAF-mediated chromatin remodeling of facultative heterochromatin. They also found that this process is specifically dependent on Top2α, but not Top1, suggesting that the specific topological state of facultative heterochromatin is catenated, which can be resolved by Top2α; rather than supercoiled, which can be resolved by Top1. Their findings imply that decatenating DNA within facultative heterochromatin by Top2α can make the chromatin more accessible for chromatin remodelers as well as transcription factors [70] (Figure 1). The data also suggest that facultative heterochromatin and accessible chromatin likely have different DNA topologies, with the former being more catenated than the latter.
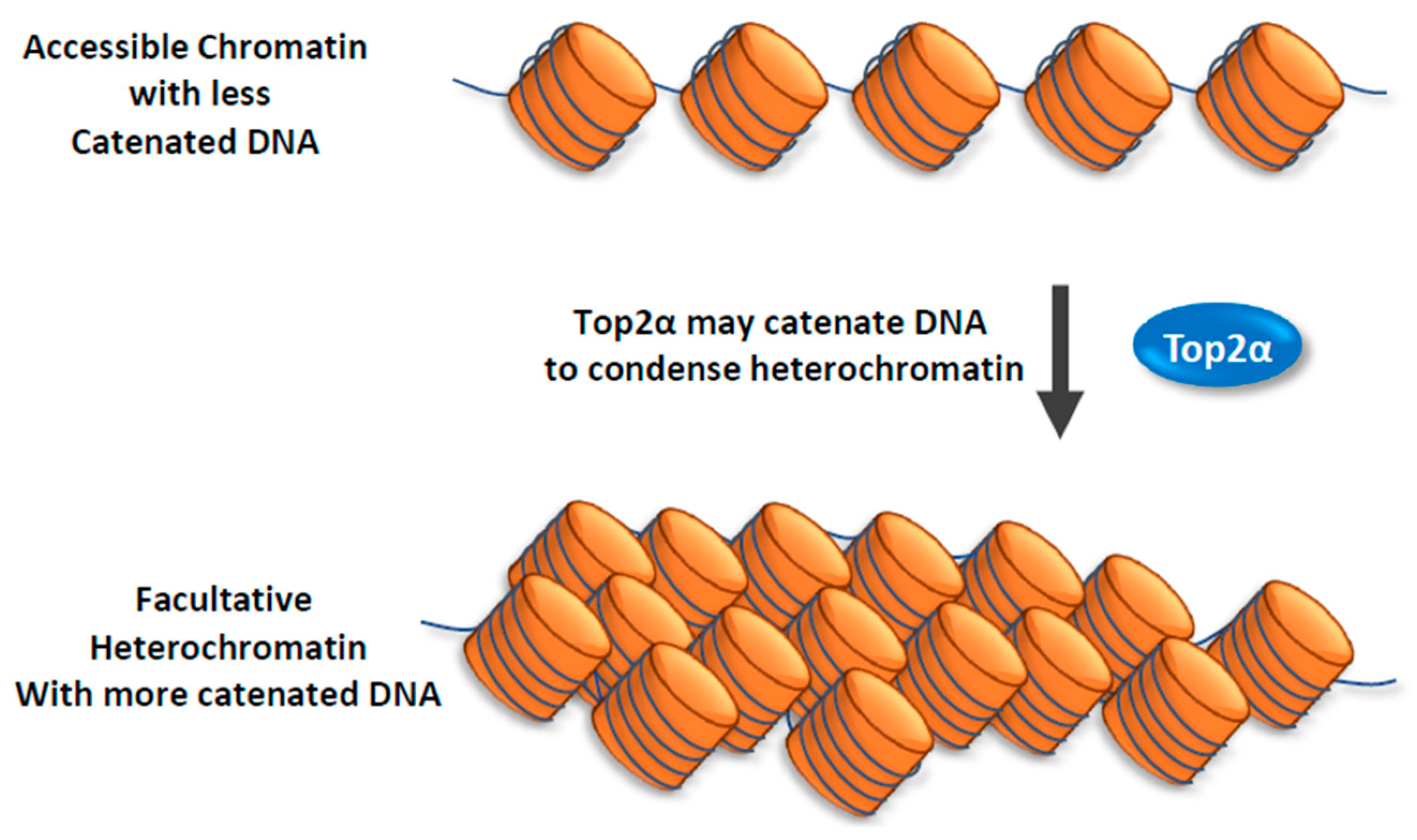
In the same study, Crabtree and colleagues discovered that Top2α has another function: it can facilitate reformation of facultative heterochromatin from accessible chromatin [70]. They found that this function of Top2α does not depend on BAF complex. Their findings suggest that Top2α may promote catenation of DNA during heterochromatin formation (Figure 2).
3.3. Top3β
3.3.1. Top3β Is a Dual-Activity Topoisomerase
Our group has recently discovered that Top3β is required for heterochromatin formation [74], as are other topoisomerases mentioned above. However, its mechanism appears to be different from others. This may not be a surprise because Top3β is the only dual activity topoisomerase in animals, and can bind and change topology of both DNA and RNA. Top3β is conserved from invertebrate to mammals, and is required for normal aging and genome stability in mice, as evidenced by findings that Top3β mutant mice exhibit reduced lifespan and fertility, chromosomal abnormality, and increased cell death [5,6,7]. Top3β contains an RNA-binding domain, possesses RNA topoisomerase activity in vitro, and is the major mRNA-binding topoisomerase in cells [8,75,76]. It should be mentioned that the RNA topoisomerase activity is conserved in type IA topoisomerases from all three domains of life: bacteria, archaea, and eukarya [75,76]; and most, but not all, type IA topoisomerases carry dual topoisomerase activity [76]. Therefore, Top3β and other type IA topoisomerases may act not only in processes of DNA, but also RNA.
3.3.2. Top3β Is Required for Heterochromatin Formation and Silencing of Transposons
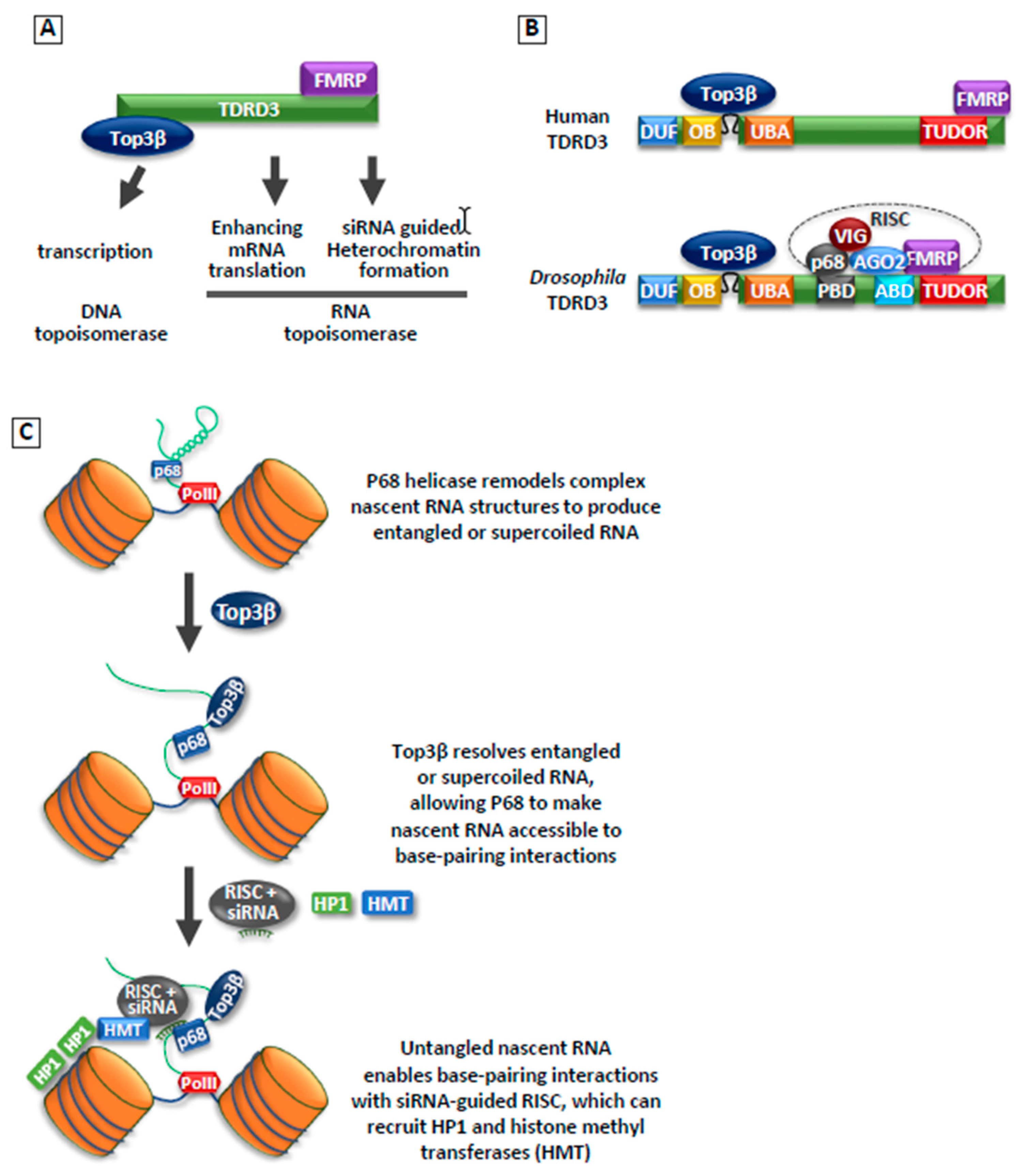
In 2018, our group reported that Top3β is required for heterochromatin formation and transcriptional silencing using Drosophila model system. We demonstrate that Top3β, along with its partner TDRD3, biochemically and genetically interact with the RNA-induced silencing complex (RISC), which consists of RNA helicase p68; RNAi effector Argonaut 2 (AGO2); and two RNA-binding proteins (RBPs), FMRP, and VIG; in this process [74] (Figure 3A,B). This interaction is likely important because the RISC complex has previously been shown to promote heterochromatin formation and silencing of transposons in Drosophila [77,78,79,80].
Indeed, Top3β mutant flies resemble those of RISC mutants in several regards, including suppression of position effect variegation (PEV), reduction of HP1 and H3K9 methylation in heterochromatin, and de-silencing of transposons. Moreover, Top3β exhibits a strong genetic interaction with siRNA biogenesis enzyme, Dcr-2; and RISC components: RNA helicase RM62 and RNAi effector, AGO2. These data suggest that Top3β may work with RISC to promote heterochromatic silencing, possibly by recruiting HP1 and other heterochromatin components. The fact that Top3β interacts with an RNA processing complex, RISC, suggests that it may solve RNA topological problems during heterochromatin function. Consistent with this notion, we showed that both RNA binding and catalytic activity of Top3β are required for Top3β-mediated heterochromatin formation and TE silencing. These findings provide a new mechanism for how topoisomerases may work: not only can they promote transcription activation by resolving supercoiled DNA, but also transcriptional repression by facilitating heterochromatin formation.
Several key questions remain unanswered regarding the mechanism of Top3β in heterochromatin formation. One question is whether the Top3β-TDRD3 complex acts on DNA or RNA. The current evidence implies that it may act on both. For example, ChIP-seq experiments revealed binding of Top3β to a small number of DNA loci within heterochromatin domains, and these sites correlate with those of a RISC component, AGO2. Some of these sites also overlap with those of HP1. These findings support a previously proposed mechanism of “nucleation and spreading” [35,81]: RISC and Top3β may bind a small number of loci to nucleate the initial assembly of heterochromatin, which may then recruit additional components to spread to other regions.
Conversely, there is also evidence that Top3β may act on RNA. In particular, Top3β can physically and functionally interact with the RNA processing complex, which includes an RNA helicase, p68. The interaction between a Type1A topoisomerase and a helicase has been observed for Top3α (which is the paralogue of Top3β) and BLM DNA helicase. It was shown that Top3α and BLM cooperate to resolve complex DNA structures, such as double-Holliday junctions [82,83]. We therefore postulated a potential mechanism of Top3β on RNA, based on the mechanism of Top3α and BLM. Top3β may work with p68 to make the nascent RNAs transcribed from heterochromatin more accessible to siRNA-guided RISC complex (Figure 3C). Possibly, p68 helicase may unwind the secondary and other higher order structures in the nascent RNAs, which may produce supercoiled or tangled RNA structures. Top3β may resolve these structures, allowing nascent RNA to be accessible to base-pairing interactions during siRNA-guided heterochromatin assembly.
It should be mentioned that mammalian Top3β-TDRD3 complex differs from its Drosophila counterpart in that it lacks stable association with AGO2, as evidenced by the absence of AGO2 in Top3β or TDRD3 immunoprecipitation from human cancer cell lines [8]. This result is consistent with the findings that mammalian TDRD3 lacks a recognizable AGO2-binding domain, which was detected in Drosophila TDRD3 [74]. These data argue that the mechanism by which Top3β-TDRD3 complex works with RISC complex could be different in mammals versus flies. Notably, we and others have consistently observed that TDRD3 interacts with a RISC component, FMRP [8,84,85]. FMRP has been reported to interact with AGO2 [86], and to regulate AGO2 binding to its target mRNAs in human cancer cell lines [87,88,89]. Thus, one possibility is that mammalian Top3β-TDRD3 may interact indirectly with AGO2, and this interaction is mediated by FMRP. It remains to be determined if Top3β-TDRD3 functionally works with AGO2 in promoting heterochromatin formation and silencing of transposons in mammals.
4. Importance of Topoisomerase Function in Heterochromatin
Because topoisomerases can participate in a variety of processes on DNA and RNA, one question is whether their function in heterochromatin is relevant. In the case of Top3β, its function in heterochromatin could be very important, because it may account for several deleterious phenotypes observed in humans and mice with Top3β mutations. For example, Top3β-KO mice display reduced lifespan [6]. It is known that loss of heterochromatin and de-silencing of transposons could be a driver for aging [20,90]. Thus, the impaired function of Top3β in heterochromatin and transposon silencing may contribute to the aging phenotype in the knockout mice. In human, Top3β mutations have been linked to neurological and mental disorders, including: schizophrenia, autism, epilepsy, intellectual disability, and cognitive impairment [11,91,92,93,94], suggesting that Top3β mutations may cause neurological defects. Similarly, mouse and Drosophila with Top3β mutations also show abnormal synapse formation [8]. Heterochromatin and silencing of TEs have been found to be important in neurodevelopment. Dysregulation of heterochromatin can impair normal gene expression during neuronal development, leading to neurological diseases in humans [95]. Moreover, activation of TEs has been observed is in a variety of age-associated neurogenerative diseases [96,97,98], including Alzheimer’s disease [52,53]. It is therefore possible that the defective function of Top3β in heterochromatin may contribute to the neurological and mental disorders.
It should be mentioned that de novo mutation in BAF complexes has been linked to neurological disorders, including autism and schizophrenia [99,100,101,102]. Because BAF complexes can cooperate with Top2α to promote access of facultative heterochromatin to transcription, it is possible that defective heterochromatin function may contribute to these diseases in patients with BAF mutations.
5. Conclusive Remarks
Increasing evidence shows that defective heterochromatin and dysregulation of TEs may be a cause for aging, cancer, and neurological disorders. Several topoisomerases, Top1, Top2, and Top3β, are all important for heterochromatin formation and silencing of transposons. These topoisomerases have distinct properties and can solve different topological problems—Top1 can relax DNA supercoils by swiveling; Top2 can catenate or decatenate dsDNA; and Top3β can catalyze strand passage reactions for both DNA and RNA. These data suggest that there may exist different topological problems in heterochromatin, which require multiple topoisomerases to resolve. The heterochromatin function of Top3β could be particularly important because its mutations in mouse can cause shortened life, and its mutations in human have been linked to neurological disorders.
There are many unanswered questions regarding mechanisms of topoisomerases. Are there both DNA and RNA topological problems during heterochromatin formation? If so, what types of structures? Entangled dsDNA, RNA/DNA hybrid, entangled RNA, or supercoiled DNA or RNA? It seems that all topoisomerases can work with ATP-dependent remodelers (BAF, FACT, and p68 helicase). It would be important to develop in vitro and in vivo systems to investigate the underlying mechanism. Do different topoisomerases act in common or different pathways to promote heterochromatin formation and silencing of transposons? Genetic studies in model systems or cell lines can be used to address this issue.
Author Contributions
Writing-original draft preparation, S.K.L.; writing-review and editing, W.W.
Funding
This work is supported by the Intramural Research Program of the National Institute on Aging (Z01 AG000657-08), National Institutes of Health.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wang, J.C. Cellular roles of DNA topoisomerases: A molecular perspective. Nat. Rev. Mol. Cell Biol. 2002, 3, 430–440. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.X.; Chen, A.D.; Gettel, N.J.; Hsieh, T.S. Essential functions of DNA topoisomerase I in Drosophila melanogaster. Dev. Biol. 2000, 222, 27–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morham, S.G.; Kluckman, K.D.; Voulomanos, N.; Smithies, O. Targeted disruption of the mouse topoisomerase I gene by camptothecin selection. Mol. Cell Biol. 1996, 16, 6804–6809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hohl, A.M.; Thompson, M.; Soshnev, A.A.; Wu, J.; Morris, J.; Hsieh, T.S.; Wu, C.T.; Geyer, P.K. Restoration of topoisomerase 2 function by complementation of defective monomers in Drosophila. Genetics 2012, 192, 843–856. [Google Scholar] [CrossRef] [PubMed]
- Kwan, K.Y.; Moens, P.B.; Wang, J.C. Infertility and aneuploidy in mice lacking a type IA DNA topoisomerase III β. Proc. Natl. Acad. Sci. USA 2003, 100, 2526–2531. [Google Scholar] [CrossRef] [PubMed]
- Kwan, K.Y.; Wang, J.C. Mice lacking DNA topoisomerase IIIβ develop to maturity but show a reduced mean lifespan. Proc. Natl. Acad. Sci. USA 2001, 98, 5717–5721. [Google Scholar] [CrossRef] [PubMed]
- Kwan, K.Y.; Greenwald, R.J.; Mohanty, S.; Sharpe, A.H.; Shaw, A.C.; Wang, J.C. Development of autoimmunity in mice lacking DNA topoisomerase 3β. Proc. Natl. Acad. Sci. USA 2007, 104, 9242–9247. [Google Scholar] [CrossRef]
- Xu, D.; Shen, W.; Guo, R.; Xue, Y.; Peng, W.; Sima, J.; Yang, J.; Sharov, A.; Srikantan, S.; Yang, J.; et al. Top3β is an RNA topoisomerase that works with fragile X syndrome protein to promote synapse formation. Nat. Neurosci. 2013, 16, 1238–1247. [Google Scholar] [CrossRef]
- Li, M.; Liu, Y. Topoisomerase I in Human Disease Pathogenesis and Treatments. Genomics Proteomics Bioinform. 2016, 14, 166–171. [Google Scholar] [CrossRef] [Green Version]
- Lam, C.W.; Yeung, W.L.; Law, C.Y. Global developmental delay and intellectual disability associated with a de novo TOP2B mutation. Clin Chim Acta 2017, 469, 63–68. [Google Scholar] [CrossRef]
- Stoll, G.; Pietilainen, O.P.; Linder, B.; Suvisaari, J.; Brosi, C.; Hennah, W.; Leppa, V.; Torniainen, M.; Ripatti, S.; Ala-Mello, S.; et al. Deletion of TOP3β, a component of FMRP-containing mRNPs, contributes to neurodevelopmental disorders. Nat. Neurosci. 2013, 16, 1228–1237. [Google Scholar] [CrossRef] [PubMed]
- Pommier, Y.; Sun, Y.; Huang, S.N.; Nitiss, J.L. Roles of eukaryotic topoisomerases in transcription, replication and genomic stability. Nat. Rev. Mol. Cell Biol. 2016, 17, 703–721. [Google Scholar] [CrossRef] [PubMed]
- Pommier, Y. Drugging topoisomerases: Lessons and challenges. ACS Chem. Biol. 2013, 8, 82–95. [Google Scholar] [CrossRef] [PubMed]
- Durand-Dubief, M.; Persson, J.; Norman, U.; Hartsuiker, E.; Ekwall, K. Topoisomerase I regulates open chromatin and controls gene expression in vivo. EMBO J. 2010, 29, 2126–2134. [Google Scholar] [CrossRef]
- Sperling, A.S.; Jeong, K.S.; Kitada, T.; Grunstein, M. Topoisomerase II binds nucleosome-free DNA and acts redundantly with topoisomerase I to enhance recruitment of RNA Pol II in budding yeast. Proc. Natl. Acad. Sci. USA 2011, 108, 12693–12698. [Google Scholar] [CrossRef] [Green Version]
- Baranello, L.; Wojtowicz, D.; Cui, K.; Devaiah, B.N.; Chung, H.J.; Chan-Salis, K.Y.; Guha, R.; Wilson, K.; Zhang, X.; Zhang, H.; et al. RNA Polymerase II Regulates Topoisomerase 1 Activity to Favor Efficient Transcription. Cell 2016, 165, 357–371. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, V.K.; Burger, L.; Nikoletopoulou, V.; Deogracias, R.; Thakurela, S.; Wirbelauer, C.; Kaut, J.; Terranova, R.; Hoerner, L.; Mielke, C.; et al. Target genes of Topoisomerase IIβ regulate neuronal survival and are defined by their chromatin state. Proc. Natl. Acad. Sci. USA 2012, 109, E934–E943. [Google Scholar] [CrossRef]
- Husain, A.; Begum, N.A.; Taniguchi, T.; Taniguchi, H.; Kobayashi, M.; Honjo, T. Chromatin remodeller SMARCA4 recruits topoisomerase 1 and suppresses transcription-associated genomic instability. Nat. Commun. 2016, 7, 10549. [Google Scholar] [CrossRef] [Green Version]
- Dykhuizen, E.C.; Hargreaves, D.C.; Miller, E.L.; Cui, K.; Korshunov, A.; Kool, M.; Pfister, S.; Cho, Y.J.; Zhao, K.; Crabtree, G.R. BAF complexes facilitate decatenation of DNA by topoisomerase IIalpha. Nature 2013, 497, 624–627. [Google Scholar] [CrossRef]
- Wood, J.G.; Helfand, S.L. Chromatin structure and transposable elements in organismal aging. Front. Genet. 2013, 4, 274. [Google Scholar] [CrossRef] [Green Version]
- Huisinga, K.L.; Brower-Toland, B.; Elgin, S.C. The contradictory definitions of heterochromatin: Transcription and silencing. Chromosoma 2006, 115, 110–122. [Google Scholar] [CrossRef] [PubMed]
- Jenuwein, T.; Allis, C.D. Translating the histone code. Science 2001, 293, 1074–1080. [Google Scholar] [CrossRef] [PubMed]
- Trojer, P.; Reinberg, D. Facultative heterochromatin: Is there a distinctive molecular signature? Mol. Cell 2007, 28, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Baranello, L.; Kouzine, F.; Levens, D. DNA topoisomerases beyond the standard role. Transcription 2013, 4, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Madabhushi, R. The Roles of DNA Topoisomerase IIβ in Transcription. Int. J. Mol. Sci. 2018, 19, 1917. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.H.; Chan, N.L.; Hsieh, T.S. New mechanistic and functional insights into DNA topoisomerases. Annu. Rev. Biochem. 2013, 82, 139–170. [Google Scholar] [CrossRef]
- SanMiguel, P.; Tikhonov, A.; Jin, Y.K.; Motchoulskaia, N.; Zakharov, D.; Melake-Berhan, A.; Springer, P.S.; Edwards, K.J.; Lee, M.; Avramova, Z.; et al. Nested retrotransposons in the intergenic regions of the maize genome. Science 1996, 274, 765–768. [Google Scholar] [CrossRef]
- Lander, E.S.; Linton, L.M.; Birren, B.; Nusbaum, C.; Zody, M.C.; Baldwin, J.; Devon, K.; Dewar, K.; Doyle, M.; FitzHugh, W.; et al. Initial sequencing and analysis of the human genome. Nature 2001, 409, 860–921. [Google Scholar] [Green Version]
- Mouse Genome Sequencing, C.; Waterston, R.H.; Lindblad-Toh, K.; Birney, E.; Rogers, J.; Abril, J.F.; Agarwal, P.; Agarwala, R.; Ainscough, R.; Alexandersson, M.; et al. Initial sequencing and comparative analysis of the mouse genome. Nature 2002, 420, 520–562. [Google Scholar] [CrossRef]
- Munoz-Lopez, M.; Garcia-Perez, J.L. DNA transposons: Nature and applications in genomics. Curr. Genomics 2010, 11, 115–128. [Google Scholar] [CrossRef]
- Li, W.; Jin, Y.; Prazak, L.; Hammell, M.; Dubnau, J. Transposable elements in TDP-43-mediated neurodegenerative disorders. PLoS ONE 2012, 7, e44099. [Google Scholar] [CrossRef] [PubMed]
- Batzer, M.A.; Deininger, P.L. Alu repeats and human genomic diversity. Nat. Rev. Genet. 2002, 3, 370–379. [Google Scholar] [CrossRef] [PubMed]
- Solyom, S.; Ewing, A.D.; Rahrmann, E.P.; Doucet, T.; Nelson, H.H.; Burns, M.B.; Harris, R.S.; Sigmon, D.F.; Casella, A.; Erlanger, B.; et al. Extensive somatic L1 retrotransposition in colorectal tumors. Genome Res. 2012, 22, 2328–2338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volpe, T.A.; Kidner, C.; Hall, I.M.; Teng, G.; Grewal, S.I.; Martienssen, R.A. Regulation of heterochromatic silencing and histone H3 lysine-9 methylation by RNAi. Science 2002, 297, 1833–1837. [Google Scholar] [CrossRef]
- Hall, I.M.; Shankaranarayana, G.D.; Noma, K.; Ayoub, N.; Cohen, A.; Grewal, S.I. Establishment and maintenance of a heterochromatin domain. Science 2002, 297, 2232–2237. [Google Scholar] [CrossRef]
- Allshire, R. Molecular biology. RNAi and heterochromatin—A hushed-up affair. Science 2002, 297, 1818–1819. [Google Scholar] [CrossRef]
- Villeponteau, B. The heterochromatin loss model of aging. Exp Gerontol 1997, 32, 383–394. [Google Scholar] [CrossRef]
- Larson, K.; Yan, S.J.; Tsurumi, A.; Liu, J.; Zhou, J.; Gaur, K.; Guo, D.; Eickbush, T.H.; Li, W.X. Heterochromatin formation promotes longevity and represses ribosomal RNA synthesis. PLoS Genet. 2012, 8, e1002473. [Google Scholar] [CrossRef]
- Zhang, W.; Li, J.; Suzuki, K.; Qu, J.; Wang, P.; Zhou, J.; Liu, X.; Ren, R.; Xu, X.; Ocampo, A.; et al. Aging stem cells. A Werner syndrome stem cell model unveils heterochromatin alterations as a driver of human aging. Science 2015, 348, 1160–1163. [Google Scholar] [CrossRef]
- Shumaker, D.K.; Dechat, T.; Kohlmaier, A.; Adam, S.A.; Bozovsky, M.R.; Erdos, M.R.; Eriksson, M.; Goldman, A.E.; Khuon, S.; Collins, F.S.; et al. Mutant nuclear lamin A leads to progressive alterations of epigenetic control in premature aging. Proc. Natl. Acad. Sci. USA 2006, 103, 8703–8708. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Jia, S.T.; Jia, S. New Insights into the Regulation of Heterochromatin. Trends Genet 2016, 32, 284–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kudlow, B.A.; Kennedy, B.K.; Monnat, R.J., Jr. Werner and Hutchinson-Gilford progeria syndromes: Mechanistic basis of human progeroid diseases. Nat. Rev. Mol. Cell Biol. 2007, 8, 394–404. [Google Scholar] [CrossRef] [PubMed]
- Feinberg, A.P.; Koldobskiy, M.A.; Gondor, A. Epigenetic modulators, modifiers and mediators in cancer aetiology and progression. Nat. Rev. Genet. 2016, 17, 284–299. [Google Scholar] [CrossRef] [PubMed]
- Janssen, A.; Colmenares, S.U.; Karpen, G.H. Heterochromatin: Guardian of the Genome. Annu. Rev. Cell Dev. Biol. 2018, 34, 265–288. [Google Scholar] [CrossRef] [Green Version]
- Dialynas, G.K.; Vitalini, M.W.; Wallrath, L.L. Linking Heterochromatin Protein 1 (HP1) to cancer progression. Mutat. Res. 2008, 647, 13–20. [Google Scholar] [CrossRef] [Green Version]
- Rondinelli, B.; Rosano, D.; Antonini, E.; Frenquelli, M.; Montanini, L.; Huang, D.; Segalla, S.; Yoshihara, K.; Amin, S.B.; Lazarevic, D.; et al. Histone demethylase JARID1C inactivation triggers genomic instability in sporadic renal cancer. J. Clin. Investig. 2016, 126, 4387. [Google Scholar] [CrossRef]
- Burns, K.H. Transposable elements in cancer. Nat. Rev. Cancer 2017, 17, 415–424. [Google Scholar] [CrossRef]
- Scott, E.C.; Gardner, E.J.; Masood, A.; Chuang, N.T.; Vertino, P.M.; Devine, S.E. A hot L1 retrotransposon evades somatic repression and initiates human colorectal cancer. Genome Res. 2016, 26, 745–755. [Google Scholar] [CrossRef] [Green Version]
- Tenreiro, S.; Eckermann, K.; Outeiro, T.F. Protein phosphorylation in neurodegeneration: Friend or foe? Front. Mol. Neurosci. 2014, 7, 42. [Google Scholar] [CrossRef]
- Frost, B.; Hemberg, M.; Lewis, J.; Feany, M.B. Tau promotes neurodegeneration through global chromatin relaxation. Nat. Neurosci. 2014, 17, 357–366. [Google Scholar] [CrossRef] [Green Version]
- Mansuroglu, Z.; Benhelli-Mokrani, H.; Marcato, V.; Sultan, A.; Violet, M.; Chauderlier, A.; Delattre, L.; Loyens, A.; Talahari, S.; Begard, S.; et al. Loss of Tau protein affects the structure, transcription and repair of neuronal pericentromeric heterochromatin. Sci. Rep. 2016, 6, 33047. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Samimi, H.; Gamez, M.; Zare, H.; Frost, B. Pathogenic tau-induced piRNA depletion promotes neuronal death through transposable element dysregulation in neurodegenerative tauopathies. Nat. Neurosci. 2018, 21, 1038–1048. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Jeong, H.H.; Hsieh, Y.C.; Klein, H.U.; Bennett, D.A.; De Jager, P.L.; Liu, Z.; Shulman, J.M. Tau Activates Transposable Elements in Alzheimer’s Disease. Cell Rep. 2018, 23, 2874–2880. [Google Scholar] [CrossRef] [PubMed]
- Cheutin, T.; McNairn, A.J.; Jenuwein, T.; Gilbert, D.M.; Singh, P.B.; Misteli, T. Maintenance of stable heterochromatin domains by dynamic HP1 binding. Science 2003, 299, 721–725. [Google Scholar] [CrossRef] [PubMed]
- Saksouk, N.; Simboeck, E.; Dejardin, J. Constitutive heterochromatin formation and transcription in mammals. Epigenet. Chromatin 2015, 8, 3. [Google Scholar] [CrossRef] [PubMed]
- Zuma, A.A.; Cavalcanti, D.P.; Maia, M.C.; de Souza, W.; Motta, M.C. Effect of topoisomerase inhibitors and DNA-binding drugs on the cell proliferation and ultrastructure of Trypanosoma cruzi. Int. J. Antimicrob. Agents 2011, 37, 449–456. [Google Scholar] [CrossRef] [Green Version]
- Baranello, L.; Bertozzi, D.; Fogli, M.V.; Pommier, Y.; Capranico, G. DNA topoisomerase I inhibition by camptothecin induces escape of RNA polymerase II from promoter-proximal pause site, antisense transcription and histone acetylation at the human HIF-1alpha gene locus. Nucleic Acids Res. 2010, 38, 159–171. [Google Scholar] [CrossRef]
- Dinh, T.T.; Gao, L.; Liu, X.; Li, D.; Li, S.; Zhao, Y.; O’Leary, M.; Le, B.; Schmitz, R.J.; Manavella, P.A.; et al. Correction: DNA Topoisomerase 1alpha Promotes Transcriptional Silencing of Transposable Elements through DNA Methylation and Histone Lysine 9 Dimethylation in Arabidopsis. PLoS Genet. 2015, 11, e1005452. [Google Scholar] [CrossRef]
- Manzo, S.G.; Hartono, S.R.; Sanz, L.A.; Marinello, J.; De Biasi, S.; Cossarizza, A.; Capranico, G.; Chedin, F. DNA Topoisomerase I differentially modulates R-loops across the human genome. Genome Biol. 2018, 19, 100. [Google Scholar] [CrossRef]
- Nakama, M.; Kawakami, K.; Kajitani, T.; Urano, T.; Murakami, Y. DNA-RNA hybrid formation mediates RNAi-directed heterochromatin formation. Genes Cells 2012, 17, 218–233. [Google Scholar] [CrossRef] [Green Version]
- Kas, E.; Laemmli, U.K. In vivo topoisomerase II cleavage of the Drosophila histone and satellite III repeats: DNA sequence and structural characteristics. EMBO J. 1992, 11, 705–716. [Google Scholar] [CrossRef] [PubMed]
- Swedlow, J.R.; Sedat, J.W.; Agard, D.A. Multiple chromosomal populations of topoisomerase II detected in vivo by time-lapse, three-dimensional wide-field microscopy. Cell 1993, 73, 97–108. [Google Scholar] [CrossRef]
- Mengoli, V.; Bucciarelli, E.; Lattao, R.; Piergentili, R.; Gatti, M.; Bonaccorsi, S. The analysis of mutant alleles of different strength reveals multiple functions of topoisomerase 2 in regulation of Drosophila chromosome structure. PLoS Genet. 2014, 10, e1004739. [Google Scholar] [CrossRef] [PubMed]
- Hughes, S.E.; Hawley, R.S. Topoisomerase II is required for the proper separation of heterochromatic regions during Drosophila melanogaster female meiosis. PLoS Genet. 2014, 10, e1004650. [Google Scholar] [CrossRef] [PubMed]
- Blattes, R.; Monod, C.; Susbielle, G.; Cuvier, O.; Wu, J.H.; Hsieh, T.S.; Laemmli, U.K.; Kas, E. Displacement of D1, HP1 and topoisomerase II from satellite heterochromatin by a specific polyamide. EMBO J. 2006, 25, 2397–2408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adachi, Y.; Luke, M.; Laemmli, U.K. Chromosome assembly in vitro: Topoisomerase II is required for condensation. Cell 1991, 64, 137–148. [Google Scholar] [CrossRef]
- Jiao, W.; Lin, H.M.; Timmons, J.; Nagaich, A.K.; Ng, S.W.; Misteli, T.; Rane, S.G. E2F-dependent repression of topoisomerase II regulates heterochromatin formation and apoptosis in cells with melanoma-prone mutation. Cancer Res. 2005, 65, 4067–4077. [Google Scholar] [CrossRef]
- Yang, X.; Yu, Y.; Jiang, L.; Lin, X.; Zhang, C.; Ou, X.; Osabe, K.; Liu, B. Changes in DNA methylation and transgenerational mobilization of a transposable element (mPing) by the topoisomerase II inhibitor, etoposide, in rice. BMC Plant Biol. 2012, 12, 48. [Google Scholar] [CrossRef]
- Hagan, C.R.; Sheffield, R.F.; Rudin, C.M. Human Alu element retrotransposition induced by genotoxic stress. Nat. Genet. 2003, 35, 219–220. [Google Scholar] [CrossRef]
- Miller, E.L.; Hargreaves, D.C.; Kadoch, C.; Chang, C.Y.; Calarco, J.P.; Hodges, C.; Buenrostro, J.D.; Cui, K.; Greenleaf, W.J.; Zhao, K.; et al. TOP2 synergizes with BAF chromatin remodeling for both resolution and formation of facultative heterochromatin. Nat. Struct. Mol. Biol. 2017, 24, 344–352. [Google Scholar] [CrossRef]
- Clapier, C.R.; Cairns, B.R. The biology of chromatin remodeling complexes. Annu. Rev. Biochem. 2009, 78, 273–304. [Google Scholar] [CrossRef] [PubMed]
- Kadoch, C.; Hargreaves, D.C.; Hodges, C.; Elias, L.; Ho, L.; Ranish, J.; Crabtree, G.R. Proteomic and bioinformatic analysis of mammalian SWI/SNF complexes identifies extensive roles in human malignancy. Nat. Genet. 2013, 45, 592–601. [Google Scholar] [CrossRef] [PubMed]
- Kadoch, C.; Williams, R.T.; Calarco, J.P.; Miller, E.L.; Weber, C.M.; Braun, S.M.; Pulice, J.L.; Chory, E.J.; Crabtree, G.R. Dynamics of BAF-Polycomb complex opposition on heterochromatin in normal and oncogenic states. Nat. Genet. 2017, 49, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.K.; Xue, Y.; Shen, W.; Zhang, Y.; Joo, Y.; Ahmad, M.; Chinen, M.; Ding, Y.; Ku, W.L.; De, S.; et al. Topoisomerase 3β interacts with RNAi machinery to promote heterochromatin formation and transcriptional silencing in Drosophila. Nat. Commun. 2018, 9, 4946. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.; Shen, W.; Li, W.; Xue, Y.; Zou, S.; Xu, D.; Wang, W. Topoisomerase 3β is the major topoisomerase for mRNAs and linked to neurodevelopment and mental dysfunction. Nucleic Acids Res. 2017, 45, 2704–2713. [Google Scholar] [PubMed]
- Ahmad, M.; Xue, Y.; Lee, S.K.; Martindale, J.L.; Shen, W.; Li, W.; Zou, S.; Ciaramella, M.; Debat, H.; Nadal, M.; et al. RNA topoisomerase is prevalent in all domains of life and associates with polyribosomes in animals. Nucleic Acids Res. 2016, 44, 6335–6349. [Google Scholar] [CrossRef] [Green Version]
- Fagegaltier, D.; Bouge, A.L.; Berry, B.; Poisot, E.; Sismeiro, O.; Coppee, J.Y.; Theodore, L.; Voinnet, O.; Antoniewski, C. The endogenous siRNA pathway is involved in heterochromatin formation in Drosophila. Proc. Natl. Acad. Sci. USA 2009, 106, 21258–21263. [Google Scholar] [CrossRef] [Green Version]
- Csink, A.K.; Linsk, R.; Birchler, J.A. The Lighten up (Lip) gene of Drosophila melanogaster, a modifier of retroelement expression, position effect variegation and white locus insertion alleles. Genetics 1994, 138, 153–163. [Google Scholar]
- Gracheva, E.; Dus, M.; Elgin, S.C. Drosophila RISC component VIG and its homolog Vig2 impact heterochromatin formation. PLoS ONE 2009, 4, e6182. [Google Scholar] [CrossRef]
- Pal-Bhadra, M.; Leibovitch, B.A.; Gandhi, S.G.; Chikka, M.R.; Bhadra, U.; Birchler, J.A.; Elgin, S.C. Heterochromatic silencing and HP1 localization in Drosophila are dependent on the RNAi machinery. Science 2004, 303, 669–672. [Google Scholar] [CrossRef]
- Allshire, R.C.; Madhani, H.D. Ten principles of heterochromatin formation and function. Nat. Rev. Mol. Cell Biol. 2018, 19, 229–244. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Siaw, G.E.; Willcox, S.; Griffith, J.D.; Hsieh, T.S. Synthesis and dissolution of hemicatenanes by type IA DNA topoisomerases. Proc. Natl. Acad. Sci. USA 2013, 110, E3587–E3594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.; Hickson, I.D. The Bloom’s syndrome helicase suppresses crossing over during homologous recombination. Nature 2003, 426, 870–874. [Google Scholar] [CrossRef] [PubMed]
- Goulet, I.; Boisvenue, S.; Mokas, S.; Mazroui, R.; Cote, J. TDRD3, a novel Tudor domain-containing protein, localizes to cytoplasmic stress granules. Hum. Mol. Genet. 2008, 17, 3055–3074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linder, B.; Plottner, O.; Kroiss, M.; Hartmann, E.; Laggerbauer, B.; Meister, G.; Keidel, E.; Fischer, U. Tdrd3 is a novel stress granule-associated protein interacting with the Fragile-X syndrome protein FMRP. Hum. Mol. Genet. 2008, 17, 3236–3246. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Tang, W.; Zhang, L.R.; Zhang, C.Y. FMRP regulates miR196a-mediated repression of HOXB8 via interaction with the AGO2 MID domain. Mol. Biosyst. 2014, 10, 1757–1764. [Google Scholar] [CrossRef]
- Lee, E.K.; Kim, H.H.; Kuwano, Y.; Abdelmohsen, K.; Srikantan, S.; Subaran, S.S.; Gleichmann, M.; Mughal, M.R.; Martindale, J.L.; Yang, X.; et al. hnRNP C promotes APP translation by competing with FMRP for APP mRNA recruitment to P bodies. Nat. Struct. Mol. Biol. 2010, 17, 732–739. [Google Scholar] [CrossRef] [Green Version]
- Kenny, P.J.; Zhou, H.; Kim, M.; Skariah, G.; Khetani, R.S.; Drnevich, J.; Arcila, M.L.; Kosik, K.S.; Ceman, S. MOV10 and FMRP regulate AGO2 association with microRNA recognition elements. Cell Rep. 2014, 9, 1729–1741. [Google Scholar] [CrossRef]
- Skariah, G.; Seimetz, J.; Norsworthy, M.; Lannom, M.C.; Kenny, P.J.; Elrakhawy, M.; Forsthoefel, C.; Drnevich, J.; Kalsotra, A.; Ceman, S. Mov10 suppresses retroelements and regulates neuronal development and function in the developing brain. BMC Biol. 2017, 15, 54. [Google Scholar] [CrossRef]
- Orr, W.C. Tightening the connection between transposable element mobilization and aging. Proc. Natl. Acad. Sci. USA 2016, 113, 11069–11070. [Google Scholar] [CrossRef] [Green Version]
- Xu, B.; Ionita-Laza, I.; Roos, J.L.; Boone, B.; Woodrick, S.; Sun, Y.; Levy, S.; Gogos, J.A.; Karayiorgou, M. De novo gene mutations highlight patterns of genetic and neural complexity in schizophrenia. Nat. Genet. 2012, 44, 1365–1369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iossifov, I.; Ronemus, M.; Levy, D.; Wang, Z.; Hakker, I.; Rosenbaum, J.; Yamrom, B.; Lee, Y.H.; Narzisi, G.; Leotta, A.; et al. De novo gene disruptions in children on the autistic spectrum. Neuron 2012, 74, 285–299. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, C.S.; Genovese, A.; Butler, M.G. Deletion of TOP3B Is Associated with Cognitive Impairment and Facial Dysmorphism. Cytogenet. Genome Res. 2016, 150, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Daghsni, M.; Lahbib, S.; Fradj, M.; Sayeb, M.; Kelmemi, W.; Kraoua, L.; Kchaou, M.; Maazoul, F.; Echebbi, S.; Ben Ali, N.; et al. TOP3B: A Novel Candidate Gene in Juvenile Myoclonic Epilepsy? Cytogenet. Genome Res. 2018, 154, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Hahn, M.; Dambacher, S.; Schotta, G. Heterochromatin dysregulation in human diseases. J. Appl. Physiol. 2010, 109, 232–242. [Google Scholar] [CrossRef] [Green Version]
- Muotri, A.R.; Marchetto, M.C.; Coufal, N.G.; Oefner, R.; Yeo, G.; Nakashima, K.; Gage, F.H. L1 retrotransposition in neurons is modulated by MeCP2. Nature 2010, 468, 443–446. [Google Scholar] [CrossRef]
- Douville, R.; Liu, J.; Rothstein, J.; Nath, A. Identification of active loci of a human endogenous retrovirus in neurons of patients with amyotrophic lateral sclerosis. Ann. Neurol. 2011, 69, 141–151. [Google Scholar] [CrossRef]
- Li, W.; Prazak, L.; Chatterjee, N.; Gruninger, S.; Krug, L.; Theodorou, D.; Dubnau, J. Activation of transposable elements during aging and neuronal decline in Drosophila. Nat. Neurosci. 2013, 16, 529–531. [Google Scholar] [CrossRef] [Green Version]
- Neale, B.M.; Kou, Y.; Liu, L.; Ma’ayan, A.; Samocha, K.E.; Sabo, A.; Lin, C.F.; Stevens, C.; Wang, L.S.; Makarov, V.; et al. Patterns and rates of exonic de novo mutations in autism spectrum disorders. Nature 2012, 485, 242–245. [Google Scholar] [CrossRef]
- O’Roak, B.J.; Vives, L.; Girirajan, S.; Karakoc, E.; Krumm, N.; Coe, B.P.; Levy, R.; Ko, A.; Lee, C.; Smith, J.D.; et al. Sporadic autism exomes reveal a highly interconnected protein network of de novo mutations. Nature 2012, 485, 246–250. [Google Scholar] [CrossRef] [Green Version]
- Koga, M.; Ishiguro, H.; Yazaki, S.; Horiuchi, Y.; Arai, M.; Niizato, K.; Iritani, S.; Itokawa, M.; Inada, T.; Iwata, N.; et al. Involvement of SMARCA2/BRM in the SWI/SNF chromatin-remodeling complex in schizophrenia. Hum. Mol. Genet. 2009, 18, 2483–2494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loe-Mie, Y.; Lepagnol-Bestel, A.M.; Maussion, G.; Doron-Faigenboim, A.; Imbeaud, S.; Delacroix, H.; Aggerbeck, L.; Pupko, T.; Gorwood, P.; Simonneau, M.; et al. SMARCA2 and other genome-wide supported schizophrenia-associated genes: Regulation by REST/NRSF, network organization and primate-specific evolution. Hum. Mol. Genet. 2010, 19, 2841–2857. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Top2α and the BAF complex cooperate to facilitate access to facultative heterochromatin. A model illustrating how Top2α and the BAF complex may work together to facilitate access to facultative heterochromatin. The BAF complex uses its ATP-dependent chromatin remodeling activity to disrupt the Polycomb group (PcG) complex containing facultative heterochromatin. Top2α is recruited in the early stage of chromatin remodeling to resolve the topological stress produced by the BAF complex, possibly using its DNA decatenation activity. This model is based on findings in the paper of Miller et al. (Nat. Struct. Mol. Biol., 2017).
Figure 1.
Top2α and the BAF complex cooperate to facilitate access to facultative heterochromatin. A model illustrating how Top2α and the BAF complex may work together to facilitate access to facultative heterochromatin. The BAF complex uses its ATP-dependent chromatin remodeling activity to disrupt the Polycomb group (PcG) complex containing facultative heterochromatin. Top2α is recruited in the early stage of chromatin remodeling to resolve the topological stress produced by the BAF complex, possibly using its DNA decatenation activity. This model is based on findings in the paper of Miller et al. (Nat. Struct. Mol. Biol., 2017).

Figure 2.
Top2α promotes the formation of facultative heterochromatin. A model illustrating how Top2α may facilitate the formation of facultative heterochromatin. Facultative heterochromatin may consist of DNA that is more catenated than that in the accessible chromatin. Top2α may catalyze DNA catenation during the formation of facultative heterochromatin from accessible chromatin. This model is based on the findings in a previous paper (Miller et al., Nat. Struct. Mol. Biol., 2017).
Figure 2.
Top2α promotes the formation of facultative heterochromatin. A model illustrating how Top2α may facilitate the formation of facultative heterochromatin. Facultative heterochromatin may consist of DNA that is more catenated than that in the accessible chromatin. Top2α may catalyze DNA catenation during the formation of facultative heterochromatin from accessible chromatin. This model is based on the findings in a previous paper (Miller et al., Nat. Struct. Mol. Biol., 2017).

Figure 3.
Top3β is a dual-activity topoisomerase that can act on both DNA and RNA, including RNAi-guided heterochromatin formation. (A) A cartoon illustrating that Top3β forms a complex with TDRD3, which interacts with FMRP. This complex can stimulate DNA transcription, mRNA translation, and siRNA-guided heterochromatin formation. (B) A cartoon illustrates human and Drosophila Top3β–TDRD3 complexes. In the latter species, Top3β–TDRD3 stably associates with the RNA-induced silencing complex (RISC) complex containing Argonaut 2 (AGO2), p68, FMRP, and VIG. It remains to be determined if mammalian Top3b–TDRD3 can also interact with RISC. (C) A model illustrates how Top3β may function during siRNA-guided heterochromatin formation. The model postulates that the unwinding of nascent transcript by p68 may generate entangled or supercoiled RNA that requires topoisomerase activity to resolve. Unwound RNAs by Top3β and p68 become more accessible for targeting by RISC to silence transposable elements (TE) as well as recruiting heterochromatin factors histone methyltransferase (HMT) and heterochromatin protein 1 (HP1).
Figure 3.
Top3β is a dual-activity topoisomerase that can act on both DNA and RNA, including RNAi-guided heterochromatin formation. (A) A cartoon illustrating that Top3β forms a complex with TDRD3, which interacts with FMRP. This complex can stimulate DNA transcription, mRNA translation, and siRNA-guided heterochromatin formation. (B) A cartoon illustrates human and Drosophila Top3β–TDRD3 complexes. In the latter species, Top3β–TDRD3 stably associates with the RNA-induced silencing complex (RISC) complex containing Argonaut 2 (AGO2), p68, FMRP, and VIG. It remains to be determined if mammalian Top3b–TDRD3 can also interact with RISC. (C) A model illustrates how Top3β may function during siRNA-guided heterochromatin formation. The model postulates that the unwinding of nascent transcript by p68 may generate entangled or supercoiled RNA that requires topoisomerase activity to resolve. Unwound RNAs by Top3β and p68 become more accessible for targeting by RISC to silence transposable elements (TE) as well as recruiting heterochromatin factors histone methyltransferase (HMT) and heterochromatin protein 1 (HP1).

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lee, S.K.; Wang, W. Roles of Topoisomerases in Heterochromatin, Aging, and Diseases. Genes 2019, 10, 884. https://0-doi-org.brum.beds.ac.uk/10.3390/genes10110884
AMA Style
Lee SK, Wang W. Roles of Topoisomerases in Heterochromatin, Aging, and Diseases. Genes. 2019; 10(11):884. https://0-doi-org.brum.beds.ac.uk/10.3390/genes10110884
Chicago/Turabian StyleLee, Seung Kyu, and Weidong Wang. 2019. "Roles of Topoisomerases in Heterochromatin, Aging, and Diseases" Genes 10, no. 11: 884. https://0-doi-org.brum.beds.ac.uk/10.3390/genes10110884
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.