Sex Determination Using RNA-Sequencing Analyses in Early Prenatal Pig Development


 , ,
, ,  , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals and Design
2.2. RNA Extraction and Library Preparation
2.3. Sequencing, Quality Control and Mapping
2.4. Sex Identification
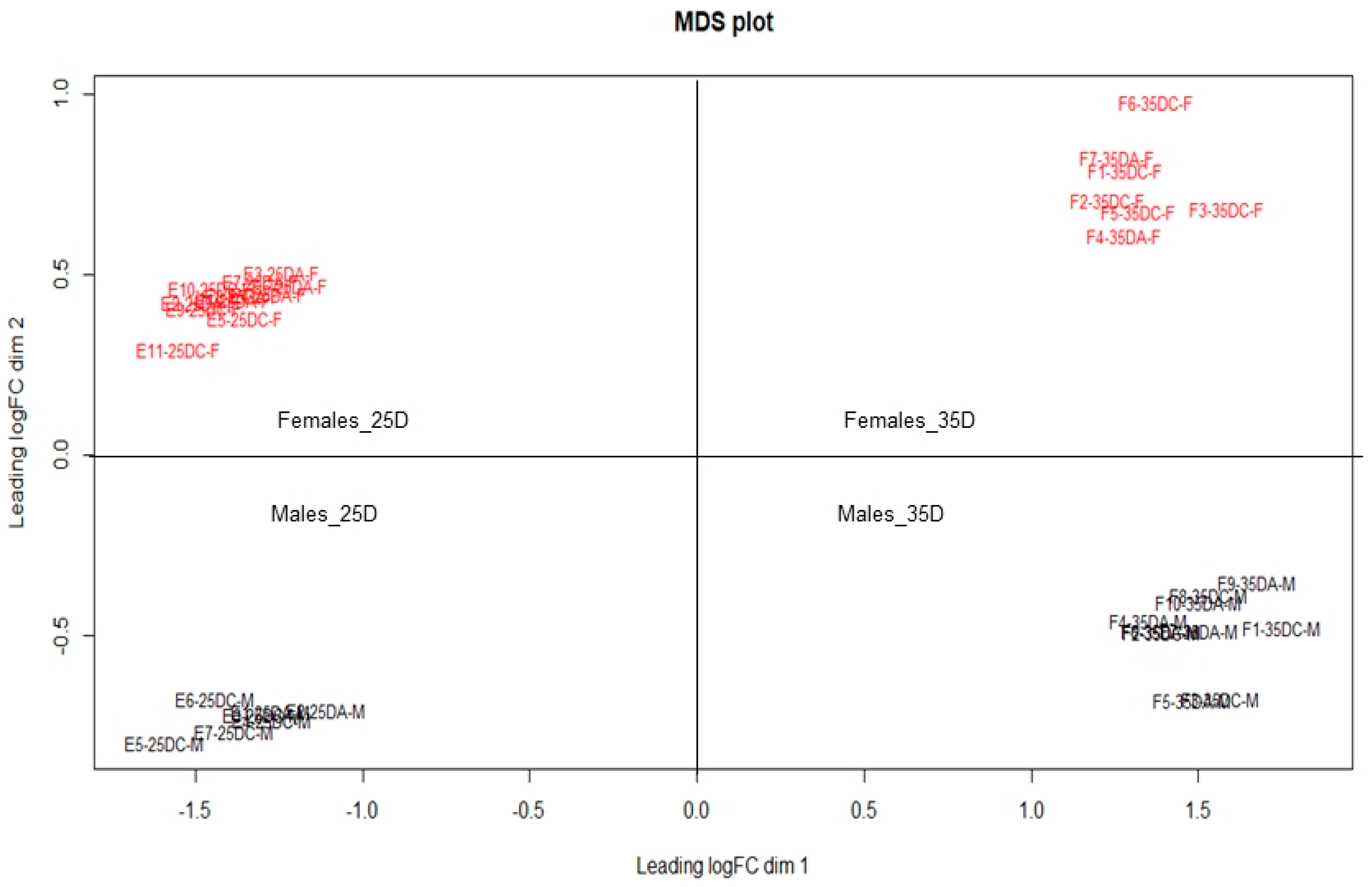
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Godfrey, K.M.; Barker, D.J.P. Fetal programming and adult health. Public Health Nutr. 2001, 4, 611–624. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Xia, H.; Yao, W.; Wang, T.; Li, J.; Piao, X.; Thacker, P.; Wu, G.; Wang, F. Effects of arginine supplementation during early gestation (day 1 to 30) on litter size and plasma metabolites in gilts and sows 1. J. Anim. Sci. 2015, 93, 5291–5303. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Bazer, F.W.; Cudd, T.A.; Meininger, C.J.; Spencer, T.E. Recent Advances in Nutritional Sciences Maternal Nutrition and Fetal Development. J. Nutr. 2004, 134, 2169–2172. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Heng, J.; Song, H.; Zhang, Y.; Lin, X.; Tian, M.; Chen, F.; Guan, W. Role of Maternal Dietary Protein and Amino Acids on Fetal Programming, Early Neonatal Development, and Lactation in Swine. Animals 2019, 9, 19. [Google Scholar] [CrossRef] [Green Version]
- Assis, K.; Botelho, D.; Marques, D.; Filardi, C.; Campos, D.; Saraiva, A.; Domingos, J.; Eliza, S.; Guimarães, F. Nutrition in fluence on sow reproductive performance and conceptuses development and survival: A review about L-arginine supplementation. Livest. Sci. 2019, 228, 97–103. [Google Scholar]
- Peñagaricano, F.; Wang, X.; Rosa, G.J.M.; Radunz, A.E.; Khatib, H. Maternal nutrition induces gene expression changes in fetal muscle and adipose tissues in sheep. BMC Genom. 2014, 15, 1034. [Google Scholar] [CrossRef] [Green Version]
- Verma, N.; Rettenmeier, A.W.; Schmitz-Spanke, S. Recent advances in the use of Sus scrofa (pig) as a model system for proteomic studies. Proteomics 2011, 11, 776–793. [Google Scholar] [CrossRef]
- Papatheodorou, I.; Fonseca, N.A.; Keays, M.; Tang, Y.A.; Barrera, E.; Bazant, W.; Burke, M.; Füllgrabe, A.; Fuentes, A.M.P.; George, N.; et al. Expression Atlas: Gene and protein expression across multiple studies and organisms. Nucleic Acids Res. 2018, 46, D246–D251. [Google Scholar] [CrossRef]
- Conesa, A.; Madrigal, P.; Tarazona, S.; Gomez-cabrero, D.; Cervera, A.; Mcpherson, A.; Szcze, W.; Gaffney, D.J.; Elo, L.L.; Zhang, X. A survey of best practices for RNA-seq data analysis. Genome Biol. 2016, 17, 13. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef]
- Sanglard, L.P.; Nascimento, M.; Moriel, P.; Sommer, J.; Ashwell, M.; Poore, M.H.; Duarte, M.D.S.; Serão, N.V.L. Impact of energy restriction during late gestation on the muscle and blood transcriptome of beef calves after preconditioning. BMC Genom. 2018, 19, 702. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Zhang, J.; Ma, J.; Guo, Y.; Li, L.; Xiao, S.; Ren, J.; Yang, B.; Huang, L. Sexually dimorphic genetic architecture of complex traits in a large-scale F2 cross in pigs. Genet. Sel. Evol. 2014, 46, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanes, M.S.; Tsoi, S.C.M.; Dyck, M.K. Accurate and Phenol Free DNA Sexing of Day 30 Porcine Embryos by PCR. J. Vis. Exp. 2016, 108, e53301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miura, K.; Tomita, A.; Kanai, Y. Sex Determination and Differentiation in Mammals. In Reproductive and Developmental Strategies; Kazuya, K., Takeshi, K., Yasuhiro, I., Mariko, K., Eds.; Springer: Tokyo, Japan, 2018; pp. 407–433. ISBN 9784431566090. [Google Scholar]
- Goodfellow, P.N.; Darling, S.M. Genetics of sex determination in man and mouse. Development 1988, 102, 251–258. [Google Scholar] [PubMed]
- Pelliniemi, L.J. Ultrastructure of the early ovary and testis in pig embryos. Am. J. Anat. 1975, 144, 89–111. [Google Scholar] [CrossRef]
- Hyttel, P.; Sinowatz, F.; Vejlsted, M.; Betteridge, K. Essentials of Domestic Animal Embryology; Edwards, R., Rodenhuis, J., Betteridge, K., Eds.; Saunders: Philadelphia, PA, USA, 2010; ISBN 978-0-7020-2899-1. [Google Scholar]
- Tsoi, S.; Blanes, M.; Chen, T.Y.; Langendijk, P.; Athorn, R.; Foxcroft, G.; Dyck, M. Identification of differentially expressed genes in sexed pig embryos during post-hatching development in primiparous sows exposed to differing intermittent suckling and breeding strategies. Genom. Data 2016, 9, 30–34. [Google Scholar] [CrossRef] [Green Version]
- Bick, J.T.; Flöter, V.L.; Robinson, M.D.; Bauersachs, S.; Ulbrich, S.E. Small RNA-seq analysis of single porcine blastocysts revealed that maternal estradiol-17beta exposure does not affect miRNA isoform (isomiR) expression. BMC Genom. 2018, 19, 590. [Google Scholar] [CrossRef] [Green Version]
- Han, S.H.; Yang, B.C.; Ko, M.S.; Oh, H.S.; Lee, S.S. Length difference between equine ZFX and ZFY genes and its application for molecular sex determination. J. Assist. Reprod. Genet. 2010, 27, 725–728. [Google Scholar] [CrossRef] [Green Version]
- Petropoulos, S.; Edsgärd, D.; Reinius, B.; Deng, Q.; Panula, S.P.; Codeluppi, S.; Reyes, A.P.; Linnarsson, S.; Sandberg, R.; Lanner, F. Single-Cell RNA-Seq Reveals Lineage and X Chromosome Dynamics in Human Preimplantation Embryos. Cell 2016, 165, 1012–1026. [Google Scholar] [CrossRef] [Green Version]
- Costa, K.A.; Saraiva, A.; Guimaraes, J.D.; Marques, D.B.D.; Machado-neves, M.; Barbosa, L.M.R.; Villadiego, F.A.C.; Veroneze, R.; de Oliveira, L.F.; Garcia, I.S.; et al. Dietary L-arginine supplementation during early gestation of gilts affects conceptuses development. Theriogenology 2019, 140, 62–71. [Google Scholar] [CrossRef]
- Nie, H.; Wang, Y.; Su, Y.; Hua, J. Exploration of miRNAs and target genes of cytoplasmic male sterility line in cotton during flower bud development. Funct. Integr. Genom. 2018, 18, 457–476. [Google Scholar] [CrossRef] [PubMed]
- BAQCOM: Bioinformatics Analysis for Quality Control and Mapping. Available online: https://github.com/hanielcedraz/BAQCOM (accessed on 15 April 2019).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; Mccarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Koopman, P.; Gubbay, J.; Vivian, N.; Goodfellow, P.; Lovell-Badge, R. Male development of chromosomally female mice transgenic for Sry. Nature 1991, 351, 117–121. [Google Scholar] [CrossRef]
- Krausz, C.; Casamonti, E. Spermatogenic failure and the Y chromosome. Hum. Genet. 2017, 136, 637–655. [Google Scholar] [CrossRef]
- Kashimada, K.; Koopman, P. Sry: The master switch in mammalian sex determination. Development 2010, 137, 3921–3930. [Google Scholar] [CrossRef] [Green Version]
- Rauschendorf, M.A.; Zimmer, J.; Ohnmacht, C.; Vogt, P.H. DDX3X, the X homologue of AZFa gene DDX3Y, expresses a complex pattern of transcript variants only in the male germ line. Mol. Hum. Reprod. 2014, 20, 1208–1222. [Google Scholar] [CrossRef]
- Peneder, P.; Wallner, B.; Vogl, C. Exchange of genetic information between therian X and Y chromosome gametologs in old evolutionary strata. Ecol. Evol. 2017, 7, 8478–8487. [Google Scholar] [CrossRef]
- Fan, H.; Hu, Y.; Shan, L.; Yu, L.; Wang, B.; Li, M.; Wu, Q.; Wei, F. Synteny search identifies Carnivore Y chromosome for evolution of male specific genes. Integr. Zool. 2018, 14, 224–234. [Google Scholar] [CrossRef] [PubMed]
- Maan, A.A.; Eales, J.; Akbarov, A.; Rowland, J.; Xu, X.; Jobling, M.A.; Charchar, F.J.; Tomaszewski, M. The y chromosome: A blueprint for men’s health? Eur. J. Hum. Genet. 2017, 25, 1181–1188. [Google Scholar] [CrossRef] [PubMed]
- Hacker, A.; Capel, B.; Goodfellow, P.; Lovell-Badge, R. Expression of Sry, the mouse sex determining gene. Development 1995, 121, 1603–1614. [Google Scholar] [PubMed]
- Daneau, I.; Ethier, J.-F.; Lussier, J.G.; Silversides, D.W. Porcine SRY Gene Locus and Genital Ridge Expression. Biol. Reprod. 2005, 55, 47–53. [Google Scholar] [CrossRef] [Green Version]
- Akimoto, C.; Kitagawa, H.; Matsumoto, T.; Kato, S. Spermatogenesis-specific association of SMCY and MSH5. Genes Cells 2008, 13, 623–633. [Google Scholar] [CrossRef]
- Bellott, D.W.; Hughes, J.F.; Skaletsky, H.; Brown, L.G.; Pyntikova, T.; Cho, T.; Koutseva, N.; Zaghlul, S.; Graves, T.; Rock, S.; et al. Mammalian Y chromosomes retain widely expressed dosage-sensitive regulators. Nature 2014, 508, 494–499. [Google Scholar] [CrossRef]
- Rasmussen, P.B.; Staller, P. The KDM5 family of histone demethylases as targets in oncology drug discovery. Epigenomics 2014, 6, 277–286. [Google Scholar] [CrossRef]
- Nakasuji, T.; Ogonuki, N.; Chiba, T.; Kato, T.; Shiozawa, K.; Yamatoya, K.; Tanaka, H.; Kondo, T.; Miyado, K.; Miyasaka, N.; et al. Complementary Critical Functions of Zfy1 and Zfy2 in Mouse Spermatogenesis and Reproduction. PLoS Genet. 2017, 13, e1006578. [Google Scholar] [CrossRef]
- Kleiman, S.E.; Yogev, L.; Hauser, R.; Botchan, A.; Maymon, B.B.S.; Paz, G.; Yavetz, H. Expression profile of AZF genes in testicular biopsies of azoospermic men. Hum. Reprod. 2007, 22, 151–158. [Google Scholar] [CrossRef] [Green Version]
- Mazeyrat, S.; Saut, N.; Grigoriev, V.; Mahadevaiah, S.K.; Ojarikre, O.A.; Rattigan, Á.; Bishop, C.; Eicher, E.M.; Mitchell, M.J.; Burgoyne, P.S. A Y-encoded subunit of the translation initiation factor Eif2 is essential for mouse spermatogenesis. Nat. Genet. 2002, 29, 49–53. [Google Scholar] [CrossRef]
- Mitchell, M.J.; Metzler-Guillemain, C.; Toure, A.; Coutton, C.; Arnoult, C.; Ray, P.F. Single gene defects leading to sperm quantitative anomalies. Clin. Genet. 2017, 91, 208–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotov, A.A.; Akulenko, N.V.; Kibanov, M.V.; Olenina, L.V. DEAD-Box RNA Helicases in Animal Gametogenesis. Mol. Biol. 2014, 48, 16–28. [Google Scholar] [CrossRef]
- Webster, T.H.; Couse, M.; Grande, B.M.; Karlins, E.; Phung, T.N.; Richmond, P.A.; Whitford, W.; Wilson, M.A. Identifying, understanding, and correcting technical artifacts on the sex chromosomes in next-generation sequencing data. GigaScience 2019, 8, giz074. [Google Scholar] [CrossRef] [Green Version]
- Ballouz, S.; Dobin, A.; Gingeras, T.R.; Gillis, J. The fractured landscape of RNA-seq alignment: The default in our STARs. Nucleic Acids Res. 2018, 46, 5125–5138. [Google Scholar] [CrossRef]
- Skinner, B.M.; Sargent, C.A.; Churcher, C.; Hunt, T.; Herrero, J.; Loveland, J.E.; Dunn, M.; Louzada, S.; Fu, B.; Chow, W.; et al. The pig X and Y chromosomes: Structure, sequence and evolution. Genome Res. 2016, 26, 130–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Conceptuses Sample_ID 1 | Counts Per Million (CPM) | ΣCPMchrY 3 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| DDX3Y 2 | KDM5D | ZFY | EIF2S3Y | EIF1AY | LOC110255320 | LOC110257894 | LOC396706 | LOC100625207 | LOC110255257 | ||
| E1-25DA-M | 151.46 | 7.52 | 24.81 | 162.23 | 40.64 | 12.04 | 9.71 | 1.53 | 61.73 | 15.17 | 486.84 |
| E2-25DA-M | 166.54 | 7.64 | 31.68 | 197.73 | 45.16 | 12.30 | 11.81 | 1.95 | 61.00 | 17.72 | 553.53 |
| E3-25DC-M | 167.21 | 9.56 | 28.67 | 211.97 | 45.86 | 13.23 | 12.79 | 2.21 | 66.52 | 15.07 | 573.09 |
| E4-25DC-M | 170.76 | 10.15 | 29.78 | 189.79 | 46.39 | 11.43 | 12.41 | 2.11 | 71.81 | 17.90 | 562.53 |
| E5-25DC-M | 149.28 | 6.56 | 24.94 | 180.78 | 36.55 | 13.47 | 10.47 | 1.43 | 55.29 | 16.46 | 495.23 |
| E6-25DC-M | 156.64 | 9.19 | 30.92 | 192.23 | 40.76 | 12.14 | 10.99 | 2.30 | 68.56 | 16.24 | 539.97 |
| E7-25DC-M | 166.10 | 9.43 | 33.57 | 192.23 | 44.92 | 12.31 | 11.43 | 1.84 | 71.78 | 17.03 | 560.64 |
| F1-35DC-M | 193.41 | 5.35 | 26.07 | 130.67 | 44.65 | 9.79 | 6.25 | 1.23 | 52.46 | 13.16 | 483.04 |
| F2-35DC-M | 167.28 | 7.11 | 28.43 | 135.38 | 39.84 | 10.25 | 7.19 | 1.41 | 56.20 | 15.04 | 468.13 |
| F3-35DC-M | 168.09 | 7.05 | 31.32 | 141.53 | 37.80 | 10.96 | 7.26 | 1.78 | 58.09 | 13.03 | 476.91 |
| F4-35DA-M | 157.51 | 8.17 | 26.77 | 119.45 | 33.45 | 9.30 | 6.46 | 2.06 | 51.27 | 12.85 | 427.29 |
| F5-35DA-M | 200.33 | 8.04 | 24.02 | 126.13 | 45.09 | 12.20 | 6.56 | 2.13 | 45.83 | 12.20 | 482.53 |
| F6-35DA-M | 187.01 | 6.47 | 24.02 | 128.61 | 41.58 | 10.35 | 8.01 | 2.02 | 45.54 | 13.43 | 467.04 |
| F7-35DA-M | 180.78 | 7.06 | 29.27 | 133.42 | 41.14 | 11.74 | 6.57 | 2.10 | 50.78 | 13.20 | 476.06 |
| F8-35DC-M | 178.15 | 10.68 | 33.04 | 155.57 | 43.49 | 11.29 | 9.23 | 2.06 | 62.94 | 17.32 | 523.77 |
| F9-35DA-M | 143.33 | 10.28 | 27.83 | 135.33 | 31.42 | 12.33 | 8.65 | 1.55 | 53.79 | 12.73 | 437.24 |
| F10-35DA-M | 173.51 | 10.40 | 26.69 | 128.32 | 36.71 | 12.62 | 6.88 | 1.91 | 58.19 | 15.37 | 470.60 |
| Conceptuses Sample_ID 1 | Counts Per Million (CPM) | ΣCPMchrY 3 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| DDX3Y2 | KDM5D | ZFY | EIF2S3Y | EIF1AY | LOC110255320 | LOC110257894 | LOC396706 | LOC100625207 | LOC110255257 | ||
| E1-25DA-F | 0.00 | 0.07 | 0.00 | 0.00 | 0.07 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.14 |
| E2-25DA-F | 0.00 | 0.00 | 0.00 | 0.00 | 0.07 | 0.00 | 0.00 | 0.00 | 0.00 | 0.07 | 0.14 |
| E3-25DC-F | 0.00 | 0.00 | 0.07 | 0.07 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.14 |
| E4-25DC-F | 0.28 | 0.00 | 0.07 | 0.07 | 0.07 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.49 |
| E5-25DC-F | 0.00 | 0.00 | 0.00 | 0.15 | 0.00 | 0.00 | 0.00 | 0.00 | 0.07 | 0.15 | 0.37 |
| E6-25DC-F | 0.00 | 0.00 | 0.14 | 0.00 | 0.07 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.21 |
| E7-25DC-F | 0.00 | 0.00 | 0.15 | 0.08 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.23 |
| E8-25DC-F | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| E9-25DC-F | 0.00 | 0.00 | 0.00 | 0.00 | 0.07 | 0.00 | 0.00 | 0.00 | 0.00 | 0.07 | 0.14 |
| E10-25DC-F | 0.00 | 0.00 | 0.08 | 0.08 | 0.08 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.24 |
| E11-25DA-F | 0.13 | 0.00 | 0.07 | 0.13 | 0.07 | 0.00 | 0.00 | 0.00 | 0.20 | 0.00 | 0.60 |
| F1-35DA-F | 0.08 | 0.00 | 0.08 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.08 | 0.24 |
| F2-35DA-F | 0.00 | 0.08 | 0.23 | 0.00 | 0.08 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.39 |
| F3-35DA-F | 0.16 | 0.00 | 0.00 | 0.25 | 0.08 | 0.00 | 0.00 | 0.00 | 0.08 | 0.00 | 0.57 |
| F4-35DC-F | 0.00 | 0.00 | 0.07 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.07 |
| F5-35DA-F | 0.71 | 0.00 | 0.32 | 0.48 | 0.08 | 0.00 | 0.00 | 0.00 | 0.16 | 0.00 | 1.75 |
| F6-35DA-F | 0.22 | 0.07 | 0.14 | 0.29 | 0.07 | 0.00 | 0.00 | 0.00 | 0.22 | 0.00 | 1.01 |
| F7-35DA-F | 0.00 | 0.00 | 0.07 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.07 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teixeira, S.A.; Ibelli, A.M.G.; Cantão, M.E.; Oliveira, H.C.d.; Ledur, M.C.; Peixoto, J.d.O.; Marques, D.B.D.; Costa, K.A.; Coutinho, L.L.; Guimarães, S.E.F. Sex Determination Using RNA-Sequencing Analyses in Early Prenatal Pig Development. Genes 2019, 10, 1010. https://0-doi-org.brum.beds.ac.uk/10.3390/genes10121010
Teixeira SA, Ibelli AMG, Cantão ME, Oliveira HCd, Ledur MC, Peixoto JdO, Marques DBD, Costa KA, Coutinho LL, Guimarães SEF. Sex Determination Using RNA-Sequencing Analyses in Early Prenatal Pig Development. Genes. 2019; 10(12):1010. https://0-doi-org.brum.beds.ac.uk/10.3390/genes10121010
Chicago/Turabian StyleTeixeira, Susana A., Adriana M. G. Ibelli, Maurício E. Cantão, Haniel C. de Oliveira, Mônica C. Ledur, Jane de O. Peixoto, Daniele B. D. Marques, Karine A. Costa, Luiz. L. Coutinho, and Simone E. F. Guimarães. 2019. "Sex Determination Using RNA-Sequencing Analyses in Early Prenatal Pig Development" Genes 10, no. 12: 1010. https://0-doi-org.brum.beds.ac.uk/10.3390/genes10121010