MicroRNAs Mediated Regulation of Expression of Nucleoside Analog Pathway Genes in Acute Myeloid Leukemia




Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell culture and Reagents
2.2. RNA Isolation
2.3. Gene Expression Analysis
2.4. MicroRNA Expression Analysis
2.5. Acute Myeloid Leukemia Patient Sample Data from The Cancer Genome Atlas
2.6. Electrophoretic Mobility Shift Assays
2.7. Bioinformatic Analysis
2.8. Statistical Analysis
3. Results
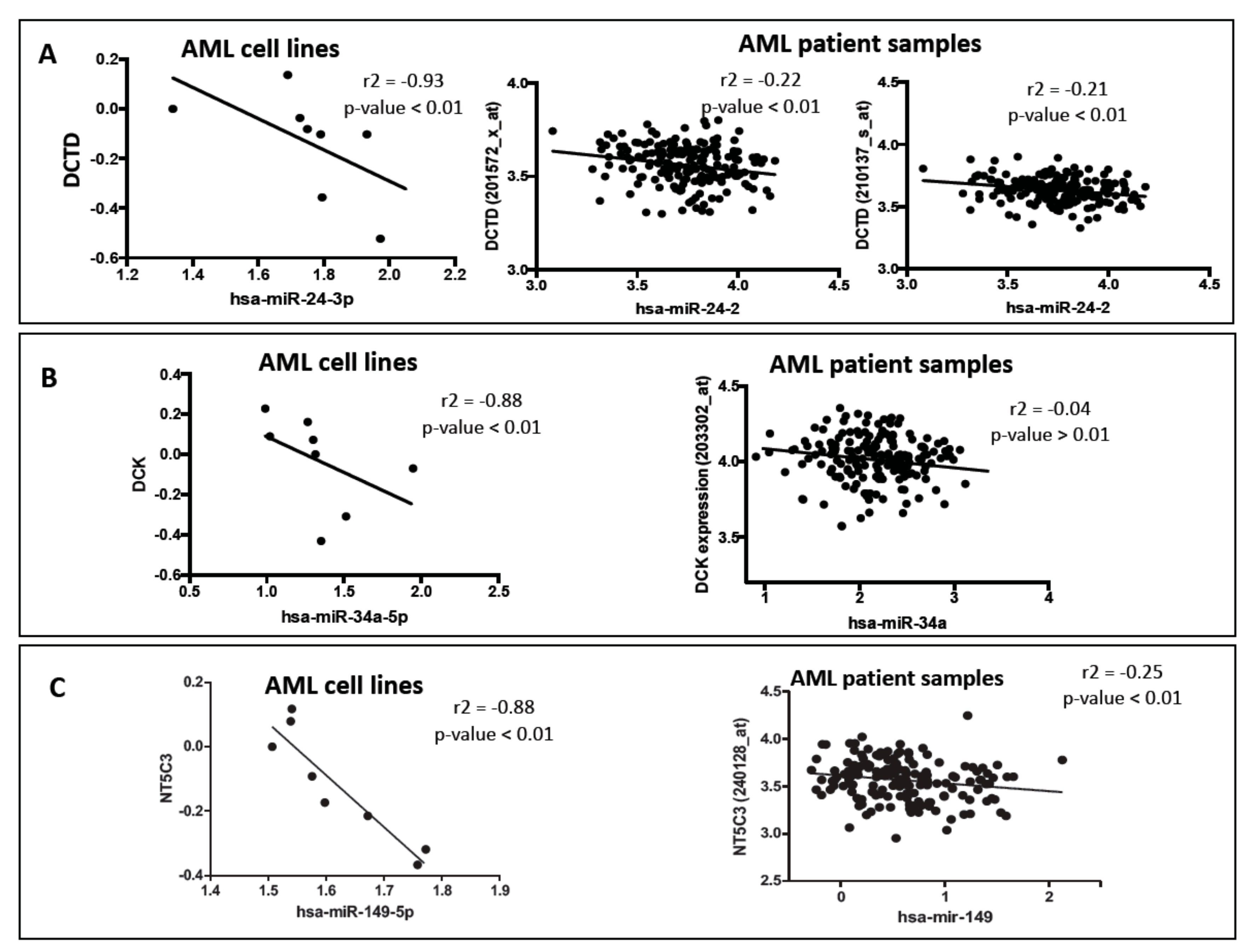
3.1. Effect of Micro RNA on the Expression of Nucleoside Analog Pathway Genes in Acute Myeloid Leukemia Cell Lines
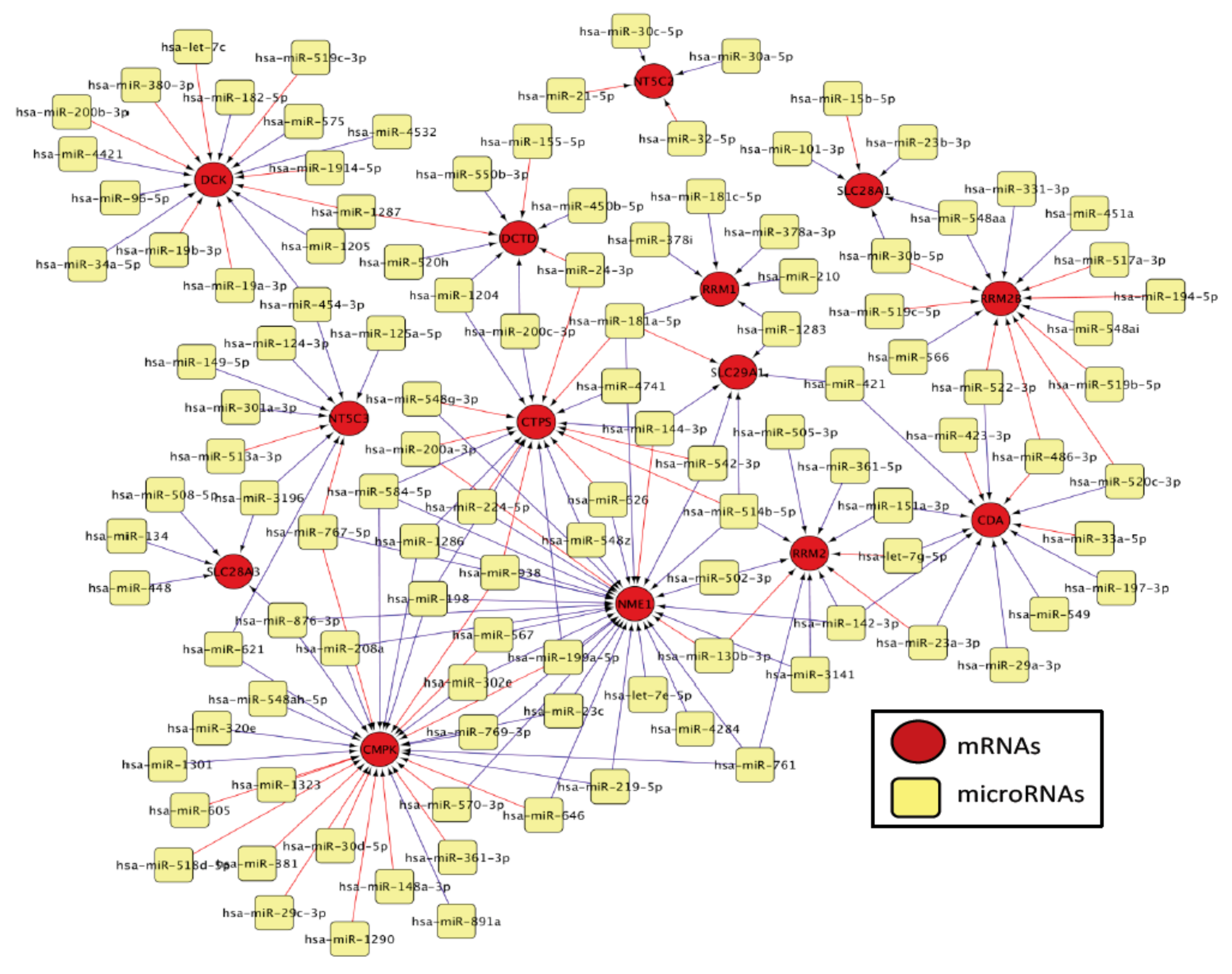
3.2. Bioinformatic Prediction of Binding of Micro RNAs and Messenger RNAs
3.3. Effect of Micro RNAs on the Expression of Nucleoside Analog Pathway Genes in Acute Myeloid Leukemia Patients
Validation of Binding Efficiencies between Messenger RNAs and Micro RNAs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Galmarini, C.M.; Clarke, M.L.; Jordheim, L.; Santos, C.L.; Cros, E.; Mackey, J.R.; Dumontet, C. Resistance to gemcitabine in a human follicular lymphoma cell line is due to partial deletion of the deoxycytidine kinase gene. BMC Pharmacol. 2004, 4, 8. [Google Scholar] [CrossRef] [PubMed]
- Galmarini, C.M.; Thomas, X.; Calvo, F.; Rousselot, P.; Rabilloud, M.; El Jaffari, A.; Cros, E.; Dumontet, C. In vivo mechanisms of resistance to cytarabine in acute myeloid leukaemia. Br. J. Haematol. 2002, 117, 860–868. [Google Scholar] [CrossRef] [PubMed]
- Lotfi, K.; Juliusson, G.; Albertioni, F. Pharmacological basis for cladribine resistance. Leuk. Lymphoma 2003, 44, 1705–1712. [Google Scholar] [CrossRef]
- Mansson, E.; Flordal, E.; Liliemark, J.; Spasokoukotskaja, T.; Elford, H.; Lagercrantz, S.; Eriksson, S.; Albertioni, F. Down-regulation of deoxycytidine kinase in human leukemic cell lines resistant to cladribine and clofarabine and increased ribonucleotide reductase activity contributes to fludarabine resistance. Biochem. Pharmacol. 2003, 65, 237–247. [Google Scholar] [CrossRef]
- Kent, O.A.; Mendell, J.T. A small piece in the cancer puzzle: MicroRNAs as tumor suppressors and oncogenes. Oncogene 2006, 25, 6188–6196. [Google Scholar] [CrossRef]
- Rieger, J.K.; Reutter, S.; Hofmann, U.; Schwab, M.; Zanger, U.M. Inflammation-associated microRNA-130b down-regulates cytochrome P450 activities and directly targets CYP2C9. Drug Metab. Dispos. 2015, 43, 884–888. [Google Scholar] [CrossRef]
- Pan, Y.Z.; Morris, M.E.; Yu, A.M. MicroRNA-328 negatively regulates the expression of breast cancer resistance protein (BCRP/ABCG2) in human cancer cells. Mol. Pharmacol. 2009, 75, 1374–1379. [Google Scholar] [CrossRef]
- Mishra, P.J.; Humeniuk, R.; Mishra, P.J.; Longo-Sorbello, G.S.; Banerjee, D.; Bertino, J.R. A miR-24 microRNA binding-site polymorphism in dihydrofolate reductase gene leads to methotrexate resistance. Proc. Natl. Acad. Sci. USA 2007, 104, 13513–13518. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Cancer Genome Atlas Research Network. Genomic and epigenomic landscapes of adult de novo acute myeloid leukemia. N. Engl. J. Med. 2013, 368, 2059–2074. [Google Scholar] [CrossRef] [PubMed]
- Kruger, J.; Rehmsmeier, M. RNAhybrid: microRNA target prediction easy, fast and flexible. Nucleic Acids Res. 2006, 34, W451–W454. [Google Scholar] [CrossRef] [PubMed]
- Kutmon, M.; Kelder, T.; Mandaviya, P.; Evelo, C.T.; Coort, S.L. CyTargetLinker: A cytoscape app to integrate regulatory interactions in network analysis. PLoS ONE 2013, 8, e82160. [Google Scholar] [CrossRef]
- Bhise, N.S.; Chauhan, L.; Shin, M.; Cao, X.; Pounds, S.; Lamba, V.; Lamba, J.K. MicroRNA-mRNA Pairs Associated with outcome in AML: From in vitro cell-based studies to AML patients. Front. Pharmacol. 2015, 6, 324. [Google Scholar] [CrossRef] [PubMed]
- Wallace, J.A.; O’Connell, R.M. MicroRNAs and acute myeloid leukemia: Therapeutic implications and emerging concepts. Blood 2017, 130, 1290–1301. [Google Scholar] [CrossRef]
- Dumontet, C.; Fabianowska-Majewska, K.; Mantincic, D.; Callet Bauchu, E.; Tigaud, I.; Gandhi, V.; Lepoivre, M.; Peters, G.J.; Rolland, M.O.; Wyczechowska, D.; et al. Common resistance mechanisms to deoxynucleoside analogues in variants of the human erythroleukaemic line K562. Br. J. Haematol. 1999, 106, 78–85. [Google Scholar] [CrossRef]
- Bhalla, K.; Nayak, R.; Grant, S. Isolation and characterization of a deoxycytidine kinase-deficient human promyelocytic leukemic cell line highly resistant to 1-β-d-arabinofuranosylcytosine. Cancer Res. 1984, 44, 5029–5037. [Google Scholar]
- Verhoef, V.; Sarup, J.; Fridland, A. Identification of the mechanism of activation of 9-β-D-arabinofuranosyladenine in human lymphoid cells using mutants deficient in nucleoside kinases. Cancer Res. 1981, 41, 4478–4483. [Google Scholar]
- Galmarini, C.M.; Thomas, X.; Calvo, F.; Rousselot, P.; El Jafaari, A.; Cros, E.; Dumontet, C. Potential mechanisms of resistance to cytarabine in AML patients. Leuk. Res. 2002, 26, 621–629. [Google Scholar] [CrossRef]
- Garofalo, M.; Jeon, Y.J.; Nuovo, G.J.; Middleton, J.; Secchiero, P.; Joshi, P.; Alder, H.; Nazaryan, N.; di Leva, G. Correction: MiR-34a/c-dependent PDGFR-α/β downregulation inhibits tumorigenesis and enhances TRAIL-induced apoptosis in lung cancer. PLoS ONE 2015, 10, e0131729. [Google Scholar] [CrossRef]
- Hong, J.H.; Roh, K.S.; Suh, S.S.; Lee, S.; Sung, S.W.; Park, J.K.; Byun, J.H.; Kang, J.H. The expression of microRNA-34a is inversely correlated with c-MET and CDK6 and has a prognostic significance in lung adenocarcinoma patients. Tumour Biol. 2015, 36, 9327–9337. [Google Scholar] [CrossRef]
- Lu, G.; Sun, Y.; An, S.; Xin, S.; Ren, X.; Zhang, D.; Wu, P.; Liao, W.; Ding, Y.; Liang, L. MicroRNA-34a targets FMNL2 and E2F5 and suppresses the progression of colorectal cancer. Exp. Mol. Pathol. 2015, 99, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Qiao, P.; Li, G.; Bi, W.; Yang, L.; Yao, L.; Wu, D. microRNA-34a inhibits epithelial mesenchymal transition in human cholangiocarcinoma by targeting Smad4 through transforming growth factor-beta/Smad pathway. BMC Cancer 2015, 15, 469. [Google Scholar] [CrossRef]
- Wang, X.; Li, J.; Dong, K.; Lin, F.; Long, M.; Ouyang, Y.; Wei, J.; Chen, X.; Weng, Y.; He, T.; et al. Tumor suppressor miR-34a targets PD-L1 and functions as a potential immunotherapeutic target in acute myeloid leukemia. Cell. Signal. 2015, 27, 443–452. [Google Scholar] [CrossRef]
- Capizzi, R.L.; White, J.C.; Powell, B.L.; Perrino, F. Effect of dose on the pharmacokinetic and pharmacodynamic effects of cytarabine. Semin. Hematol. 1991, 28 (Suppl. 4), 54–69. [Google Scholar] [PubMed]
- Fridland, A.; Verhoef, V. Mechanism for ara-CTP catabolism in human leukemic cells and effect of deaminase inhibitors on this process. Semin. Oncol. 1987, 14 (Suppl. 1), 262–268. [Google Scholar] [PubMed]
- Liliemark, J.O.; Plunkett, W. Regulation of 1-β-D-arabinofuranosylcytosine 5′-triphosphate accumulation in human leukemia cells by deoxycytidine 5′-triphosphate. Cancer Res. 1986, 46, 1079–1083. [Google Scholar]
- Gilbert, J.A.; Salavaggione, O.E.; Ji, Y.; Pelleymounter, L.L.; Eckloff, B.W.; Wieben, E.D.; Ames, M.M.; Weinshilboum, R.M. Gemcitabine pharmacogenomics: Cytidine deaminase and deoxycytidylate deaminase gene resequencing and functional genomics. Clin. Cancer Res. 2006, 12, 1794–1803. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Chen, Z.; Xue, F.; Chen, W.; Ma, R.; Cheng, S.; Cui, P. MicroRNA-24 inhibits growth, induces apoptosis, and reverses radioresistance in laryngeal squamous cell carcinoma by targeting X-linked inhibitor of apoptosis protein. Cancer Cell Int. 2015, 15, 61. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.; Wang, J.; Song, Y.; Zhao, S.; Liu, H.; Tang, D.; Pan, B.; Zhao, H.; Zhang, Q. miRNA-24-3p promotes cell proliferation and inhibits apoptosis in human breast cancer by targeting p27Kip1. Oncol. Rep. 2015, 34, 995–1002. [Google Scholar] [CrossRef] [PubMed]
- Manvati, S.; Mangalhara, K.C.; Kalaiarasan, P.; Srivastava, N.; Bamezai, R.N. miR-24-2 regulates genes in survival pathway and demonstrates potential in reducing cellular viability in combination with docetaxel. Gene 2015, 567, 217–224. [Google Scholar] [CrossRef]
- Zhao, G.; Liu, L.; Zhao, T.; Jin, S.; Jiang, S.; Cao, S.; Han, J.; Xin, Y.; Dong, Q.; Liu, X.; et al. Upregulation of miR-24 promotes cell proliferation by targeting NAIF1 in non-small cell lung cancer. Tumour Biol. 2015, 36, 3693–3701. [Google Scholar] [CrossRef] [PubMed]
- Organista-Nava, J.; Gomez-Gomez, Y.; Illades-Aguiar, B.; del Carmen Alarcon-Romero, L.; Saavedra-Herrera, M.V.; Rivera-Ramirez, A.B.; Garzón-Barrientos, V.H.; Leyva-Vázquez, M.A. High miR-24 expression is associated with risk of relapse and poor survival in acute leukemia. Oncol. Rep. 2015, 33, 1639–1649. [Google Scholar] [CrossRef]
- Gao, Y.; Liu, Y.; Du, L.; Li, J.; Qu, A.; Zhang, X.; Wang, L.; Wang, C. Down-regulation of miR-24-3p in colorectal cancer is associated with malignant behavior. Med. Oncol. 2015, 32, 362. [Google Scholar] [CrossRef]
- Pan, B.; Chen, Y.; Song, H.; Xu, Y.; Wang, R.; Chen, L. Mir-24-3p downregulation contributes to VP16-DDP resistance in small-cell lung cancer by targeting ATG4A. Oncotarget 2015, 6, 317–331. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathway Genes | MiRNAs | Spearman r | p Value | Minimum Free Energy (mfe) mRNA-miRNA Pair (kcal/mol) |
|---|---|---|---|---|
| DCTD | hsa-miR-24-3p | −0.9341 | 0.0011 | −27.2 |
| DCK | hsa-miR-96-5p | −0.9048 | 0.0046 | −25.6 |
| DCK | hsa-miR-34a-5p | −0.881 | 0.0072 | −24.3 |
| NT5C3 | hsa-miR-149-5p | −0.881 | 0.0072 | −21.4 |
| RRM2 | hsa-miR-151a-3p | −0.9524 | 0.0011 | −26.4 |
| RRM2B | hsa-miR-194-5p | −0.881 | 0.0072 | −21.6 |
| CMPK | hsa-miR-1301 | −0.9762 | 0.0004 | −23.6 |
| CMPK | hsa-miR-320e | −0.9524 | 0.0011 | −20.1 |
| CMPK | hsa-miR-1323 | −0.9286 | 0.0022 | −22.3 |
| CMPK | hsa-miR-584-5p | −0.9286 | 0.0022 | −23.7 |
| CMPK | hsa-miR-381 | −0.881 | 0.0072 | −25.8 |
| CMPK | hsa-miR-507 | −0.881 | 0.0072 | −19.1 |
| CMPK | hsa-miR-605 | −0.881 | 0.0072 | −23.1 |
| CMPK | hsa-miR-762 | −0.881 | 0.0072 | −29.2 |
| CMPK | hsa-miR-769-3p | −0.881 | 0.0072 | −27.6 |
| CMPK | hsa-miR-891a | −0.881 | 0.0072 | −22.9 |
| NME1 | hsa-miR-514b-5p | −0.9286 | 0.0022 | −22 |
| NME1 | hsa-miR-542-3p | −0.9286 | 0.0022 | −19.3 |
| NME1 | hsa-miR-570-3p | −0.9048 | 0.0046 | −22.2 |
| NME1 | hsa-miR-646 | −0.9048 | 0.0046 | −25.9 |
| NME1 | hsa-miR-224-5p | −0.881 | 0.0072 | −17.9 |
| NME1 | hsa-miR-761 | −0.881 | 0.0072 | −24.3 |
| NME1 | hsa-miR-767-5p | −0.881 | 0.0072 | −28.8 |
| SLC28A1 | hsa-miR-548aa | −0.9643 | 0.0028 | −16.3 |
| SLC28A3 | hsa-miR-448 | −0.9643 | 0.0028 | −23.8 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhise, N.S.; Elsayed, A.H.; Cao, X.; Pounds, S.; Lamba, J.K. MicroRNAs Mediated Regulation of Expression of Nucleoside Analog Pathway Genes in Acute Myeloid Leukemia. Genes 2019, 10, 319. https://0-doi-org.brum.beds.ac.uk/10.3390/genes10040319
Bhise NS, Elsayed AH, Cao X, Pounds S, Lamba JK. MicroRNAs Mediated Regulation of Expression of Nucleoside Analog Pathway Genes in Acute Myeloid Leukemia. Genes. 2019; 10(4):319. https://0-doi-org.brum.beds.ac.uk/10.3390/genes10040319
Chicago/Turabian StyleBhise, Neha S., Abdelrahman H. Elsayed, Xueyuan Cao, Stanley Pounds, and Jatinder K. Lamba. 2019. "MicroRNAs Mediated Regulation of Expression of Nucleoside Analog Pathway Genes in Acute Myeloid Leukemia" Genes 10, no. 4: 319. https://0-doi-org.brum.beds.ac.uk/10.3390/genes10040319