Selection of Suitable Reference Genes for RT-qPCR Gene Expression Analysis in Siberian Wild Rye (Elymus sibiricus) under Different Experimental Conditions

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Treatments and Tissue Collection
2.3. RNA Extraction
2.4. First-Strand cDNA Synthesis
2.5. Selection of Candidate Reference Genes and Primer Design
2.6. RT-qPCR
2.7. Data Analysis
3. Results
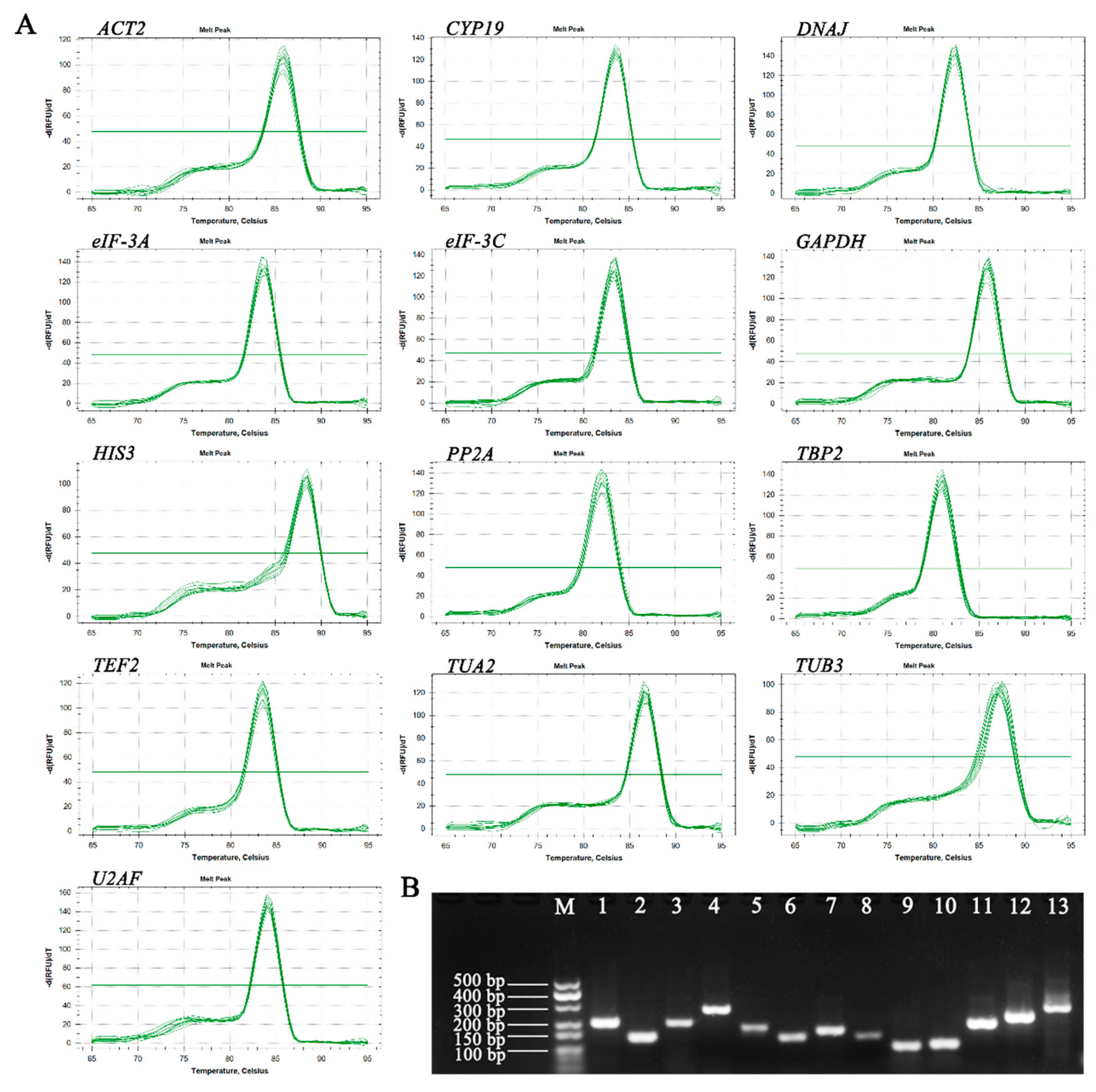
3.1. Verification of Primer Specificity and PCR Amplification Efficiency
3.2. Cq Data Collection and Variations in Reference Genes
3.3. Expression Stability Analysis of the Candidate Reference Genes
3.3.1. GeNorm Analysis
3.3.2. NormFinder Analysis
3.3.3. BestKeeper Analysis
3.3.4. Delta Ct Analysis
3.3.5. RefFinder Analysis
3.4. Validation of the Stability of Reference Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Xie, W.G.; Zhang, J.C.; Zhao, X.H.; Zhang, J.Q.; Wang, Y.R. Siberian wild rye (Elymus sibiricus L.): Genetic diversity of germplasm determined using DNA fingerprinting and SCoT markers. Biochem. Syst. Ecol. 2015, 60, 186–192. [Google Scholar] [CrossRef]
- Zhou, Q.; Luo, D.; Ma, L.C.; Xie, W.G.; Wang, Y.; Wang, Y.R.; Liu, Z.P. Development and cross-species transferability of EST-SSR markers in Siberian wildrye (Elymus sibiricus L.) using Illumina sequencing. Sci. Rep. 2016, 6, 20549. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.Y.; Zhang, J.C.; Zhao, X.H.; Xie, W.G.; Wang, Y.R. Assessing and broadening genetic diversity of Elymus sibiricus germplasm for the improvement of seed shattering. Molecules 2016, 21, 869. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.H.; Xie, W.G.; Zhang, J.C.; Zhang, Z.Y.; Wang, Y.R. Histological characteristics, cell wall hydrolytic enzymes activity and candidate genes expression associated with seed shattering of Elymus sibiricus accessions. Front. Plant Sci. 2017, 8, 606. [Google Scholar] [CrossRef]
- Wang, M.Y.; Hou, L.Y.; Zhu, Y.Q.; Zhang, Q.; Wang, H.; Xia, F.S.; Chen, L.L.; Mao, P.S.; Hannaway, D.B. Siberian wildrye seed yield limited by assimilate source. Field Crops Res. 2018, 218, 18–23. [Google Scholar] [CrossRef]
- Xie, W.G.; Zhao, X.H.; Zhang, J.Q.; Wang, Y.R.; Liu, W.X. Assessment of genetic diversity of Siberian wild rye (Elymus sibiricus L.) germplasms with variation of seed shattering and implication for future genetic improvement. Biochem. Syst. Ecol. 2015, 58, 211–218. [Google Scholar] [CrossRef]
- Xie, W.G.; Zhang, J.C.; Zhao, X.H.; Zhang, Z.Y.; Wang, Y.R. Transcriptome profiling of Elymus sibiricus, an important forage grass in Qinghai-Tibet plateau, reveals novel insights into candidate genes that potentially connected to seed shattering. BMC Plant Biol. 2017, 17, 78. [Google Scholar] [CrossRef]
- Tong, Z.G.; Gao, Z.H.; Wang, F.; Zhou, J.; Zhang, Z. Selection of reliable reference genes for gene expression studies in peach using real-time PCR. BMC Mol. Biol. 2009, 10, 71. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Gao, Z.; Li, H.; Li, Q.; Zhang, C.X.; Xu, W.P.; Song, S.R.; Ma, C.; Wang, S.P. Selection of reference genes for miRNA qRT-PCR under abiotic stress in grapevine. Sci. Rep. 2018, 8, 4444. [Google Scholar] [CrossRef]
- Narancio, R.; John, U.; Mason, J.; Spangenberg, G. Selection of optimal reference genes for quantitative RT-PCR transcript abundance analysis in white clover (Trifolium repens L.). Funct. Plant Biol. 2018, 45, 737–744. [Google Scholar] [CrossRef]
- Expósito-Rodríguez, M.; Borges, A.A.; Borges-Pérez, A.; Pérez, J.A. Selection of internal control genes for quantitative real-time RT-PCR studies during tomato development process. BMC Plant Biol. 2008, 8, 131. [Google Scholar] [CrossRef] [PubMed]
- Sheshadri, S.; Nishanth, M.; Yamine, V.; Simon, B. Effect of Melatonin on the stability and expression of reference genes in Catharanthus roseus. Sci. Rep. 2018, 8, 2222. [Google Scholar] [CrossRef] [PubMed]
- Guénin, S.; Mauriat, M.; Pelloux, J.; Van Wuytswinkel, O.; Bellini, C.; Gutierrez, L. Normalization of qRT-PCR data: The necessity of adopting a systematic, experimental conditions-specific, validation of references. J. Exp. Bot. 2009, 60, 487–493. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, L.; Mauriat, M.; Guénin, S.; Pelloux, J.; Lefebvre, J.F.; Louvet, R.; Rusterucci, C.; Moritz, T.; Guerineau, F.; Bellini, C. The lack of a systematic validation of reference genes: A serious pitfall undervalued in reverse transcription-polymerase chain reaction (RT-PCR) analysis in plants. Plant Biotechnol. J. 2008, 6, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Ozsolak, F.; Milos, P.M. RNA sequencing: Advances, challenges and opportunities. Nat. Rev. Genet. 2011, 12, 87. [Google Scholar] [CrossRef] [PubMed]
- Jain, M. Next-generation sequencing technologies for gene expression profiling in plants. Brief. Funct. Genom. 2011, 11, 63–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinha, P.; Saxena, R.K.; Singh, V.K.; Krishnamurthy, L.; Varshney, R.K. Selection and validation of housekeeping genes as reference for gene expression studies in pigeonpea (Cajanus cajan) under heat and salt stress conditions. Front. Plant Sci. 2015, 6, 1071. [Google Scholar] [CrossRef] [PubMed]
- Bao, W.; Qu, Y.; Shan, X.; Wan, Y. Screening and validation of housekeeping genes of the root and cotyledon of Cunninghamia lanceolata under abiotic stresses by using quantitative real-time PCR. Int. J. Mol. Sci. 2016, 17, 1198. [Google Scholar] [CrossRef]
- Jain, M.; Nijhawan, A.; Tyagi, A.K.; Khurana, J.P. Validation of housekeeping genes as internal control for studying gene expression in rice by quantitative real-time PCR. Biochem. Biophys. Res. Commun. 2006, 345, 646–651. [Google Scholar] [CrossRef]
- Glare, E.; Divjak, M.; Bailey, M.; Walters, E. β-Actin and GAPDH housekeeping gene expression in asthmatic airways is variable and not suitable for normalising mRNA levels. Thorax 2002, 57, 765–770. [Google Scholar] [CrossRef]
- Gao, M.M.; Liu, Y.P.; Ma, X.; Shuai, Q.; Gai, J.Y.; Li, Y. Evaluation of reference genes for normalization of gene expression using quantitative RT-PCR under aluminum, cadmium, and heat stresses in soybean. PLoS ONE 2017, 12, e0168965. [Google Scholar] [CrossRef]
- Thellin, O.; Zorzi, W.; Lakaye, B.; De Borman, B.; Coumans, B.; Hennen, G.; Grisar, T.; Igout, A.; Heinen, E. Housekeeping genes as internal standards: Use and limits. J. Biotechnol. 1999, 75, 291–295. [Google Scholar] [CrossRef]
- Zhou, Z.; Cong, P.H.; Tian, Y.; Zhu, Y.M. Using RNA-seq data to select reference genes for normalizing gene expression in apple roots. PLoS ONE 2017, 12, e0185288. [Google Scholar] [CrossRef]
- Dheda, K.; Huggett, J.; Chang, J.; Kim, L.; Bustin, S.; Johnson, M.; Rook, G.; Zumla, A. The implications of using an inappropriate reference gene for real-time reverse transcription PCR data normalization. Anal. Biochem. 2005, 344, 141–143. [Google Scholar] [CrossRef] [PubMed]
- Sang, J.; Han, X.J.; Liu, M.Y.; Qiao, G.R.; Jiang, J.; Zhuo, R.Y. Selection and validation of reference genes for real-time quantitative PCR in hyperaccumulating ecotype of Sedum alfredii under different heavy metals stresses. PLoS ONE 2013, 8, e82927. [Google Scholar] [CrossRef] [PubMed]
- Karuppaiya, P.; Yan, X.X.; Liao, W.; Wu, J.; Chen, F.; Tang, L. Identification and validation of superior reference gene for gene expression normalization via RT-qPCR in staminate and pistillate flowers of Jatropha curcas—A biodiesel plant. PLoS ONE 2017, 12, e0172460. [Google Scholar]
- Xiao, Z.; Sun, X.B.; Liu, X.Q.; Li, C.; He, L.S.; Chen, S.P.; Su, J.L. Selection of reliable reference genes for gene expression studies on Rhododendron molle G. Don. Front. Plant Sci. 2016, 7, 1547. [Google Scholar] [CrossRef]
- Wu, Z.J.; Tian, C.; Jiang, Q.; Li, X.H.; Zhuang, J. Selection of suitable reference genes for qRT-PCR normalization during leaf development and hormonal stimuli in tea plant (Camellia sinensis). Sci. Rep. 2016, 6, 19748. [Google Scholar] [CrossRef]
- Wan, Q.; Chen, S.L.; Shan, Z.H.; Yang, Z.L.; Chen, L.M.; Zhang, C.J.; Yuan, S.L.; Hao, Q.N.; Zhang, X.J.; Qiu, D.Z. Stability evaluation of reference genes for gene expression analysis by RT-qPCR in soybean under different conditions. PLoS ONE 2017, 12, e0189405. [Google Scholar] [CrossRef]
- Wang, H.B.; Wang, J.J.; Jiang, J.F.; Chen, S.M.; Guan, Z.Y.; Liao, Y.; Chen, F.D. Reference genes for normalizing transcription in diploid and tetraploid Arabidopsis. Sci. Rep. 2014, 4, 6781. [Google Scholar] [CrossRef] [PubMed]
- Pabuayon, I.M.; Yamamoto, N.; Trinidad, J.L.; Longkumer, T.; Raorane, M.L.; Kohli, A. Reference genes for accurate gene expression analyses across different tissues, developmental stages and genotypes in rice for drought tolerance. Rice 2016, 9, 32. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Li, P.F.; Luo, X.; Chang, T.L.; Li, J.X.; Zhao, Y.W.; Xu, Y. Selection of appropriate reference genes for the detection of rhythmic gene expression via quantitative real-time PCR in Tibetan hulless barley. PLoS ONE 2018, 13, e0190559. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.K.; Yan, H.D.; Jiang, X.M.; Zhang, Y.; Zhang, X.Q.; Ji, Y.; Zeng, B.; Xu, B.; Yin, G.H.; Lee, S. Reference gene selection for quantitative real-time reverse-transcriptase PCR in orchardgrass subjected to various abiotic stresses. Gene 2014, 553, 158–165. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, J.; Xu, L.; Lai, H.; Chen, Y.; Yang, Z.M.; Huang, B.R. Identification and validation of reference genes for seashore paspalum response to abiotic stresses. Int. J. Mol. Sci. 2017, 18, 1322. [Google Scholar] [CrossRef] [PubMed]
- Niu, K.J.; Shi, Y.; Ma, H.L. Selection of candidate reference genes for gene expression analysis in Kentucky Bluegrass (Poa pratensis L.) under abiotic stress. Front. Plant Sci. 2017, 8, 193. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.Q.; Eamens, A.L.; Grof, C.P. Reference gene identification for reliable normalisation of quantitative RT-PCR data in Setaria viridis. Plant Methods 2018, 14, 24. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.H.; Jiang, X.; Zhao, K.; Zhao, X.H.; Yin, J.; Xie, W.G. Screening of germplasm with low seed shattering rate and evaluation on agronomic traits in Elymus sibiricus L. (Chinese with English abstract). J. Plant Genet. Resour. 2015, 16, 691–699. [Google Scholar]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034–1. [Google Scholar] [CrossRef] [PubMed]
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper–Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Silver, N.; Best, S.; Jiang, J.; Thein, S.L. Selection of housekeeping genes for gene expression studies in human reticulocytes using real-time PCR. BMC Mol. Biol. 2006, 7, 33. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.L.; Xiao, P.; Chen, D.L.; Xu, L.; Zhang, B.H. miRDeepFinder: A miRNA analysis tool for deep sequencing of plant small RNAs. Plant Mol. Biol. 2012, 80, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Tan, Z.Q.; Hu, B.Y.; Yang, Z.M.; Xu, B.; Zhuang, L.L.; Huang, B.R. Selection and validation of reference genes for target gene analysis with quantitative RT-PCR in leaves and roots of bermudagrass under four different abiotic stresses. Physiol. Plant. 2015, 155, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Radonić, A.; Thulke, S.; Mackay, I.M.; Landt, O.; Siegert, W.; Nitsche, A. Guideline to reference gene selection for quantitative real-time PCR. Biochem. Biophys. Res. Commun. 2004, 313, 856–862. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Silveira, É.D.; Alves-Ferreira, M.; Guimarães, L.A.; da Silva, F.R.; de Campos Carneiro, V.T. Selection of reference genes for quantitative real-time PCR expression studies in the apomictic and sexual grass Brachiaria brizantha. BMC Plant Biol. 2009, 9, 84. [Google Scholar] [CrossRef]
- Udvardi, M.K.; Czechowski, T.; Scheible, W.-R. Eleven golden rules of quantitative RT-PCR. Plant Cell 2008, 20, 1736–1737. [Google Scholar] [CrossRef]
- Fei, X.T.; Shi, Q.Q.; Yang, T.X.; Fei, Z.X.; Wei, A.Z. Expression stabilities of ten candidate reference genes for RT-qPCR in Zanthoxylum bungeanum Maxim. Molecules 2018, 23, 802. [Google Scholar] [CrossRef]
- Xiang, Q.J.; Li, J.; Qin, P.; He, M.L.; Yu, X.M.; Zhao, K.; Zhang, X.P.; Ma, M.G.; Chen, Q.; Chen, X.Q. Identification and evaluation of reference genes for qRT-PCR studies in Lentinula edodes. PLoS ONE 2018, 13, e0190226. [Google Scholar] [CrossRef]
- Dai, F.W.; Zhao, X.T.; Tang, C.M.; Wang, Z.J.; Kuang, Z.S.; Li, Z.Y.; Huang, J.; Luo, G.Q. Identification and validation of reference genes for qRT-PCR analysis in mulberry (Morus alba L.). PLoS ONE 2018, 13, e0194129. [Google Scholar] [CrossRef]
- Zheng, T.C.; Chen, Z.L.; Ju, Y.Q.; Zhang, H.; Cai, M.; Pan, H.T.; Zhang, Q.X. Reference gene selection for qRT-PCR analysis of flower development in Lagerstroemia indica and L. speciosa. PLoS ONE 2018, 13, e0195004. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.C.; Huang, Z.F.; Huang, H.J.; Wei, S.H.; Liu, Y.; Jiang, C.L.; Zhang, J.; Zhang, C.X. Selection of relatively exact reference genes for gene expression studies in goosegrass (Eleusine indica) under herbicide stress. Sci. Rep. 2017, 7, 46494. [Google Scholar] [CrossRef] [PubMed]
- Duan, M.M.; Wang, J.L.; Zhang, X.H.; Yang, H.H.; Wang, H.P.; Qiu, Y.; Song, J.P.; Guo, Y.D.; Li, X.X. Identification of optimal reference genes for expression analysis in Radish (Raphanus sativus L.) and its relatives based on expression stability. Front. Plant Sci. 2017, 8, 1605. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ma, X.; Huang, L.K.; Zhang, X.Q. Identification of the valid reference genes for quantitative RT-PCR in annual ryegrass (Lolium multiflorum) under salt stress. Molecules 2015, 20, 4833–4847. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.K.; Yan, H.D.; Jiang, X.M.; Zhang, X.Q.; Zhang, Y.W.; Huang, X.; Zhang, Y.; Miao, J.M.; Xu, B.; Frazier, T. Evaluation of candidate reference genes for normalization of quantitative RT-PCR in switchgrass under various abiotic stress conditions. BioEnergy Res. 2014, 7, 1201–1211. [Google Scholar] [CrossRef]
- Kesten, C.; Menna, A.; Sanchez-Rodriguez, C. Regulation of cellulose synthesis in response to stress. Curr. Opin. Plant Biol. 2017, 40, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.Q.; Guo, K.; Li, Y.; Tu, Y.Y.; Hu, H.Z.; Wang, B.R.; Cui, X.C.; Peng, L.C. Expression profiling and integrative analysis of the CESA/CSL superfamily in rice. BMC Plant Biol. 2010, 10, 282. [Google Scholar] [CrossRef] [PubMed]
- Hazen, S.P.; Scott-Craig, J.S.; Walton, J.D. Cellulose synthase-like genes of rice. Plant Physiol. 2002, 128, 336–340. [Google Scholar] [CrossRef]
- Nakano, T.; Kimbara, J.; Fujisawa, M.; Kitagawa, M.; Ihashi, N.; Maeda, H.; Kasumi, T.; Ito, Y. MACROCALYX and JOINTLESS interact in the transcriptional regulation of tomato fruit abscission zone development. Plant Physiol. 2012, 158, 439–450. [Google Scholar] [CrossRef]
- Nunes, A.; Delatorre, C.; Merotto, A., Jr. Gene expression related to seed shattering and the cell wall in cultivated and weedy rice. Plant Biol. 2014, 16, 888–896. [Google Scholar] [CrossRef]
- Li, Y.H.; Cheng, X.J.; Fu, Y.Q.; Wu, Q.Q.; Guo, Y.L.; Peng, J.Y.; Zhang, W.; He, B. A genome-wide analysis of the cellulose synthase-like (Csl) gene family in maize (Zea mays). PeerJ Preprints 2018, 6, e27374v1. [Google Scholar]
- Hamann, T.; Osborne, E.; Youngs, H.L.; Misson, J.; Nussaume, L.; Somerville, C. Global expression analysis of CESA and CSL genes in Arabidopsis. Cellulose 2004, 11, 279–286. [Google Scholar] [CrossRef]
- Wu, J.Y.; Zhang, H.N.; Liu, L.Q.; Li, W.C.; Wei, Y.Z.; Shi, S.Y. Validation of reference genes for RT-qPCR studies of gene expression in preharvest and postharvest longan fruits under different experimental conditions. Front. Plant Sci. 2016, 7, 780. [Google Scholar] [CrossRef] [PubMed]
- Caño-Delgado, A.; Penfield, S.; Smith, C.; Catley, M.; Bevan, M. Reduced cellulose synthesis invokes lignification and defense responses in Arabidopsis thaliana. Plant J. 2003, 34, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.; Cho, L.-H.; Antt, H.W.; Koh, H.-J.; An, G. KNOX protein OSH15 induces grain shattering by repressing lignin biosynthesis genes. Plant Physiol. 2017, 174, 312–325. [Google Scholar] [CrossRef]
- Stolle-Smits, T.; Beekhuizen, J.G.; Kok, M.T.; Pijnenburg, M.; Recourt, K.; Derksen, J.; Voragen, A.G. Changes in cell wall polysaccharides of green bean pods during development. Plant Physiol. 1999, 121, 363–372. [Google Scholar] [CrossRef] [PubMed]
- Doblin, M.S.; Kurek, I.; Jacob-Wilk, D.; Delmer, D.P. Cellulose biosynthesis in plants: From genes to rosettes. Plant Cell Physiol. 2002, 43, 1407–1420. [Google Scholar] [CrossRef]
- Jithesh, M.; Shukla, P.S.; Kant, P.; Joshi, J.; Critchley, A.T.; Prithiviraj, B. Physiological and transcriptomics analyses reveal that Ascophyllum nodosum extracts induce salinity tolerance in Arabidopsis by regulating the expression of stress responsive genes. J. Plant Growth Regul. 2018, 1–16. [Google Scholar] [CrossRef]
- Zhang, J.L.; Shi, H.Z. Physiological and molecular mechanisms of plant salt tolerance. Photosynth. Res. 2013, 115, 1–22. [Google Scholar] [CrossRef]
- Zheng, M.; Wang, Y.H.; Liu, K.; Shu, H.M.; Zhou, Z.G. Protein expression changes during cotton fiber elongation in response to low temperature stress. J. Plant Physiol. 2012, 169, 399–409. [Google Scholar] [CrossRef]
- Dametto, A.; Sperotto, R.A.; Adamski, J.M.; Blasi, É.A.; Cargnelutti, D.; de Oliveira, L.F.; Ricachenevsky, F.K.; Fregonezi, J.N.; Mariath, J.E.; da Cruz, R.P. Cold tolerance in rice germinating seeds revealed by deep RNAseq analysis of contrasting indica genotypes. Plant Sci. 2015, 238, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ko, J.-H.; Prassinos, C.; Keathley, D.; Han, K.-H. Novel aspects of transcriptional regulation in the winter survival and maintenance mechanism of poplar. Tree Physiol. 2011, 31, 208–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Lv, F.J.; Liu, J.R.; Ma, Y.N.; Wang, Y.H.; Chen, B.L.; Meng, Y.L.; Zhou, Z.G.; Oosterhuis, D.M. Effect of late planting and shading on cellulose synthesis during cotton fiber secondary wall development. PLoS ONE 2014, 9, e105088. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Huang, L.Y.; Wang, W.S.; Zhao, X.Q.; Zhu, L.H.; Fu, B.Y.; Li, Z.K. Genome-wide gene expression profiling of introgressed indica rice alleles associated with seedling cold tolerance improvement in a japonica rice background. BMC Genom. 2012, 13, 461. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Sets | Tissue Type | Sampling Dates | Germplasm | Biological Replicates | Total Number of Samples |
|---|---|---|---|---|---|
| Different genotypes | Leaf | 4–6 leaves stage | ZN, XH, MQ, HZ, LQ, LT | 3 | 18 |
| Different developmental stages | Leaf | Seedling, Tillering, Jointing, Heading, Flowering | ZN | 3 | 15 |
| Different tissues | Root, Stem, Leaf, Tiller bud, Inflorescence | Flowering stage | ZN | 3 | 15 |
| Salt stress (250 mmol/L NaCl) | Leaf, Root | 4–6 leaves stage, 0, 12, 24 HAT 1 | ZN | 3 | 18 |
| Cold stress (3 °C) | Leaf, Root | 4–6 leaves stage, 0, 12, 24 HAT | ZN | 3 | 18 |
| Heat stress (40 °C) | Leaf, Root | 4–6 leaves stage, 0, 12, 24 HAT | ZN | 3 | 18 |
| Osmotic stress (20% PEG6000) | Leaf, Root | 4–6 leaves stage, 0, 12, 24 HAT | ZN | 3 | 18 |
| Control | Leaf, Root | 4–6 leaves stage, 0, 12, 24 HAT | ZN | 3 | 18 |
| Gene | Gene Description | Arabidopsis Homolog Locus | Amino Acid Identity with E. sibiricus (%) | Rice TIRG Identifier | Amino Acid Identity with E. sibiricus (%) |
|---|---|---|---|---|---|
| ACT2 | Actin2 | At5g09810 | 98.41 | LOC_Os11g06390 | 98.94 |
| CYP19 | Cyclophilin 19 | At2g29960 | 75.12 | LOC_Os06g49480 | 82.73 |
| DNAJ | DNAJ heat shock N-terminal domain-containing protein | At1g76700 | 66.67 | LOC_Os08g41110 | 86.04 |
| eIF-3A | Eukaryotic translation initiation factor 3A | At4g11420 | 62.36 | LOC_Os01g03070 | 79.39 |
| eIF-3C | Eukaryotic translation initiation factor 3C | At3g56150 | 62.18 | LOC_Os07g03230 | 82.43 |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase | At3g04120 | 74.93 | LOC_Os08g03290 | 92.12 |
| HIS3 | Histone 3.3 | At4g40030 | 82.93 | LOC_Os06g04030 | 100.00 |
| PP2A | Protein phosphatase 2A | At1g13320 | 74.28 | LOC_Os09g07510 | 78.36 |
| TBP2 | TATA binding protein 2 | At1g55520 | 64.50 | LOC_Os03g45410 | 59.65 |
| TEF2 | Translation elongation factor 2 | At1g56070 | 91.10 | LOC_Os02g32030 | 94.90 |
| TUA2 | Tubulin α-2 | At4g14960 | 96.67 | LOC_Os11g14220 | 97.56 |
| TUB3 | Tubulin β-3 | At1g75780 | 89.04 | LOC_Os06g46000 | 90.13 |
| U2AF | U2 auxiliary factor | At5g42820 | 74.48 | LOC_Os09g31482 | 69.97 |
| CSLE6 | Cellulose synthase-like protein E6 | At1g55850 | 47.92 | LOC_Os09g30130 | 76.64 |
| Candidate Reference Gene | Primer Sequence F/R (5′–3′) | Tm (°C) | Amplicon Length/Bp | Efficiency (%) | R2 |
|---|---|---|---|---|---|
| ACT2 | F: CCACGAGACGACCTACAATTCCATC | 60.0 | 206 | 98.46 | 0.981 |
| R: CTCCGATCCAGACACTGTACTTCCT | |||||
| CYP19 | F: GGTGGTGAATCAATCTACGGCACAA | 60.4 | 141 | 93.73 | 0.996 |
| R: GCTCGTGGTTACAGTGGTGATGAAG | |||||
| DNAJ | F: GCAATGGCGTCAATGGCTTCAC | 59.6 | 209 | 90.35 | 0.992 |
| R: GCATCACTAAGTCTGGACACCTCAG | |||||
| eIF-3A | F: GAATCAGGCACAAGCACTGGAAGA | 59.9 | 284 | 95.4 | 0.984 |
| R: ACAACCTACGGAACTCGGTGGAT | |||||
| eIF-3C | F: GAATCATAAGGCTGCTGCGAAGGT | 60.0 | 183 | 92.84 | 0.990 |
| R: AACGGTGGTGGTCCTCTATTGTCA | |||||
| GAPDH | F: GTTACTGTCTTCGGCGTCAGGAAC | 60.2 | 137 | 97.05 | 0.998 |
| R: ACCTTCTTGGCACCACCCTTCA | |||||
| HIS3 | F: CTACAACTGGAGGAGTGAAGAAGCC | 59.6 | 164 | 98.85 | 0.993 |
| R: GAAGCGGAGGTCAGTCTTGAAGTC | |||||
| PP2A | F: GTGATAATGAGGCTGAAGTGCGGAT | 60.0 | 140 | 101.08 | 0.990 |
| R: GCGAACATGCTGAGACGAATCTGA | |||||
| TBP2 | F: CGGATGAGGCAGCCAAAGATTGT | 59.6 | 103 | 98.65 | 0.993 |
| R: TGTTCTCGAAGGCAGTGTATGTCTC | |||||
| TEF2 | F: AGAAGTCCTGCCGTACCGTTATGA | 60.1 | 107 | 98.97 | 0.995 |
| R: GCCATCATCAATAGCCTCAGCCAAT | |||||
| TUA2 | F: TGGTGATGTTGTGCCGAAGGATG | 59.8 | 195 | 100.6 | 0.996 |
| R: ACGACACTGGTGGAGTTGGAGAT | |||||
| TUB3 | F: GTGCAGAACAAGAACTCGTCCTACT | 59.5 | 233 | 99.97 | 0.998 |
| R: TCGGTGAACTCCATCTCGTCCAT | |||||
| U2AF | F: ATCGCTGCTCTCGCATCCATAAC | 59.7 | 281 | 98.5 | 0.998 |
| R: TGCTGCTGCCTGATCTTCCTCT | |||||
| Target gene | |||||
| CSLE6 | F: AAGGATGGTGGAATGGACAGAGGAT | 60.0 | 149 | 98.37 | 0.994 |
| R: CTTGGACTCGTCTTCGTCGCTTAC | |||||
| Method | Ranking Order (Better—Good—Average) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | |
| Different genotypes | |||||||||||||
| Delta Ct | TBP2 | HIS3 | eIF-3A | CYP19 | PP2A | ACT2 | TUA2 | TEF2 | DNAJ | eIF-3C | TUB3 | GAPDH | U2AF |
| BestKeeper | HIS3 | ACT2 | CYP19 | DNAJ | eIF-3C | TUA2 | TBP2 | TEF2 | TUB3 | eIF-3A | PP2A | GAPDH | U2AF |
| NormFinder | TBP2 | eIF-3A | TUA2 | PP2A | CYP19 | TEF2 | HIS3 | ACT2 | DNAJ | eIF-3C | TUB3 | GAPDH | U2AF |
| GeNorm | DNAJ | eIF-3C | CYP19 | ACT2 | HIS3 | TBP2 | TUA2 | eIF-3A | PP2A | TEF2 | TUB3 | GAPDH | U2AF | |
| RefFinder | TBP2 | HIS3 | CYP19 | DNAJ | ACT2 | eIF-3A | eIF-3C | TUA2 | PP2A | TEF2 | TUB3 | GAPDH | U2AF |
| Different developmental stages | |||||||||||||
| Delta Ct | PP2A | TUA2 | HIS3 | DNAJ | TUB3 | CYP19 | eIF-3C | ACT2 | GAPDH | TEF2 | TBP2 | eIF-3A | U2AF |
| BestKeeper | DNAJ | TUA2 | HIS3 | ACT2 | CYP19 | eIF-3C | PP2A | TUB3 | GAPDH | TBP2 | TEF2 | eIF-3A | U2AF |
| NormFinder | PP2A | TUA2 | TUB3 | HIS3 | DNAJ | CYP19 | eIF-3C | ACT2 | GAPDH | TEF2 | TBP2 | eIF-3A | U2AF |
| GeNorm | ACT2 | HIS3 | DNAJ | TUA2 | PP2A | CYP19 | eIF-3C | TUB3 | GAPDH | TBP2 | TEF2 | eIF-3A | U2AF | |
| RefFinder | TUA2 | PP2A | HIS3 | DNAJ | ACT2 | TUB3 | CYP19 | eIF-3C | GAPDH | TBP2 | TEF2 | eIF-3A | U2AF |
| Different tissues | |||||||||||||
| Delta Ct | PP2A | TBP2 | eIF-3A | CYP19 | TEF2 | DNAJ | HIS3 | GAPDH | eIF-3C | TUA2 | U2AF | TUB3 | ACT2 |
| BestKeeper | TBP2 | DNAJ | PP2A | eIF-3C | eIF-3A | CYP19 | HIS3 | TEF2 | GAPDH | TUA2 | U2AF | TUB3 | ACT2 |
| NormFinder | TBP2 | CYP19 | PP2A | eIF-3A | TEF2 | DNAJ | eIF-3C | HIS3 | GAPDH | TUA2 | U2AF | TUB3 | ACT2 |
| GeNorm | eIF-3A | PP2A | CYP19 | TBP2 | TEF2 | DNAJ | HIS3 | eIF-3C | GAPDH | TUA2 | U2AF | TUB3 | ACT2 | |
| RefFinder | TBP2 | PP2A | eIF-3A | CYP19 | DNAJ | TEF2 | eIF-3C | HIS3 | GAPDH | TUA2 | U2AF | TUB3 | ACT2 |
| Salt stress | |||||||||||||
| Delta Ct | ACT2 | PP2A | TBP2 | TUA2 | HIS3 | TUB3 | U2AF | CYP19 | TEF2 | DNAJ | eIF-3A | eIF-3C | GAPDH |
| BestKeeper | eIF-3C | DNAJ | U2AF | TUB3 | TEF2 | PP2A | TBP2 | ACT2 | TUA2 | HIS3 | CYP19 | eIF-3A | GAPDH |
| NormFinder | ACT2 | PP2A | TBP2 | TUA2 | HIS3 | CYP19 | TUB3 | TEF2 | U2AF | DNAJ | eIF-3A | eIF-3C | GAPDH |
| GeNorm | ACT2 | TBP2 | PP2A | TUB3 | U2AF | TEF2 | DNAJ | TUA2 | HIS3 | CYP19 | eIF-3A | eIF-3C | GAPDH | |
| RefFinder | ACT2 | TBP2 | PP2A | TUB3 | U2AF | TUA2 | DNAJ | eIF-3C | TEF2 | HIS3 | CYP19 | eIF-3A | GAPDH |
| Heat stress | |||||||||||||
| Delta Ct | ACT2 | TUA2 | PP2A | CYP19 | DNAJ | TBP2 | TUB3 | U2AF | HIS3 | TEF2 | eIF-3A | eIF-3C | GAPDH |
| BestKeeper | eIF-3C | TEF2 | U2AF | TUB3 | TBP2 | PP2A | DNAJ | ACT2 | TUA2 | CYP19 | HIS3 | eIF-3A | GAPDH |
| NormFinder | ACT2 | TUA2 | PP2A | CYP19 | DNAJ | TBP2 | TUB3 | HIS3 | U2AF | TEF2 | eIF-3A | eIF-3C | GAPDH |
| GeNorm | CYP19 | TUA2 | ACT2 | DNAJ | PP2A | TBP2 | TUB3 | U2AF | TEF2 | HIS3 | eIF-3C | eIF-3A | GAPDH | |
| RefFinder | ACT2 | TUA2 | CYP19 | PP2A | DNAJ | TBP2 | TUB3 | eIF-3C | U2AF | TEF2 | HIS3 | eIF-3A | GAPDH |
| Cold stress | |||||||||||||
| Delta Ct | PP2A | ACT2 | DNAJ | TBP2 | TEF2 | TUA2 | CYP19 | HIS3 | U2AF | TUB3 | eIF-3C | GAPDH | eIF-3A |
| BestKeeper | eIF-3C | TUB3 | TBP2 | DNAJ | ACT2 | TUA2 | TEF2 | U2AF | PP2A | CYP19 | HIS3 | GAPDH | eIF-3A |
| NormFinder | PP2A | ACT2 | DNAJ | HIS3 | TEF2 | TBP2 | CYP19 | U2AF | TUA2 | TUB3 | eIF-3C | GAPDH | eIF-3A |
| GeNorm | TEF2 | U2AF | PP2A | DNAJ | TBP2 | ACT2 | TUA2 | CYP19 | HIS3 | TUB3 | eIF-3C | GAPDH | eIF-3A | |
| RefFinder | PP2A | ACT2 | DNAJ | TEF2 | TBP2 | U2AF | eIF-3C | TUB3 | TUA2 | HIS3 | CYP19 | GAPDH | eIF-3A |
| Osmotic stress | |||||||||||||
| Delta Ct | DNAJ | PP2A | ACT2 | TUB3 | CYP19 | U2AF | TBP2 | HIS3 | TEF2 | TUA2 | eIF-3C | eIF-3A | GAPDH |
| BestKeeper | eIF-3C | TEF2 | U2AF | DNAJ | TUB3 | TBP2 | PP2A | ACT2 | HIS3 | CYP19 | TUA2 | eIF-3A | GAPDH |
| NormFinder | ACT2 | CYP19 | HIS3 | PP2A | TUB3 | DNAJ | TBP2 | TUA2 | U2AF | TEF2 | eIF-3C | eIF-3A | GAPDH |
| GeNorm | DNAJ | U2AF | TEF2 | TUB3 | TBP2 | PP2A | ACT2 | CYP19 | HIS3 | TUA2 | eIF-3C | eIF-3A | GAPDH | |
| RefFinder | DNAJ | U2AF | ACT2 | PP2A | TUB3 | TEF2 | CYP19 | eIF-3C | TBP2 | HIS3 | TUA2 | eIF-3A | GAPDH |
| All samples | |||||||||||||
| Delta Ct | PP2A | TBP2 | CYP19 | TEF2 | DNAJ | HIS3 | TUA2 | TUB3 | ACT2 | U2AF | eIF-3C | eIF-3A | GAPDH |
| BestKeeper | eIF-3C | DNAJ | U2AF | TBP2 | TUB3 | TEF2 | PP2A | ACT2 | CYP19 | TUA2 | HIS3 | eIF-3A | GAPDH |
| NormFinder | PP2A | CYP19 | TBP2 | HIS3 | TEF2 | TUA2 | DNAJ | TUB3 | ACT2 | U2AF | eIF-3C | eIF-3A | GAPDH |
| GeNorm | PP2A | TBP2 | DNAJ | TEF2 | TUB3 | CYP19 | TUA2 | HIS3 | ACT2 | U2AF | eIF-3C | eIF-3A | GAPDH | |
| RefFinder | PP2A | TBP2 | DNAJ | CYP19 | TEF2 | eIF-3C | TUB3 | HIS3 | TUA2 | U2AF | ACT2 | eIF-3A | GAPDH |
| Gene name | ACT2 | CYP19 | DNAJ | eIF-3A | eIF-3C | GAPDH | HIS3 | PP2A | TBP2 | TEF2 | TUA2 | TUB3 | U2AF |
| Number of times the best gene appears | 9 | 1 | 5 | 1 | 6 | 0 | 2 | 11 | 8 | 1 | 2 | 0 | 2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Xie, W.; Yu, X.; Zhang, Z.; Zhao, Y.; Wang, N.; Wang, Y. Selection of Suitable Reference Genes for RT-qPCR Gene Expression Analysis in Siberian Wild Rye (Elymus sibiricus) under Different Experimental Conditions. Genes 2019, 10, 451. https://0-doi-org.brum.beds.ac.uk/10.3390/genes10060451
Zhang J, Xie W, Yu X, Zhang Z, Zhao Y, Wang N, Wang Y. Selection of Suitable Reference Genes for RT-qPCR Gene Expression Analysis in Siberian Wild Rye (Elymus sibiricus) under Different Experimental Conditions. Genes. 2019; 10(6):451. https://0-doi-org.brum.beds.ac.uk/10.3390/genes10060451
Chicago/Turabian StyleZhang, Junchao, Wengang Xie, Xinxuan Yu, Zongyu Zhang, Yongqiang Zhao, Na Wang, and Yanrong Wang. 2019. "Selection of Suitable Reference Genes for RT-qPCR Gene Expression Analysis in Siberian Wild Rye (Elymus sibiricus) under Different Experimental Conditions" Genes 10, no. 6: 451. https://0-doi-org.brum.beds.ac.uk/10.3390/genes10060451