Changes in Serum Iron and Leukocyte mRNA Levels of Genes Involved in Iron Metabolism in Amateur Marathon Runners—Effect of the Running Pace

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Characteristics of the Subjects and Baseline Laboratory Parameters
2.2. Experimental Procedure
2.3. RNA Extraction and Reverse Transcription
2.4. Quantitative Polymerase Chain Reaction Assay
2.5. Statistical Analysis
2.5.1. Serum Parameters
2.5.2. mRNA Levels
3. Results
3.1. Blood Morphology
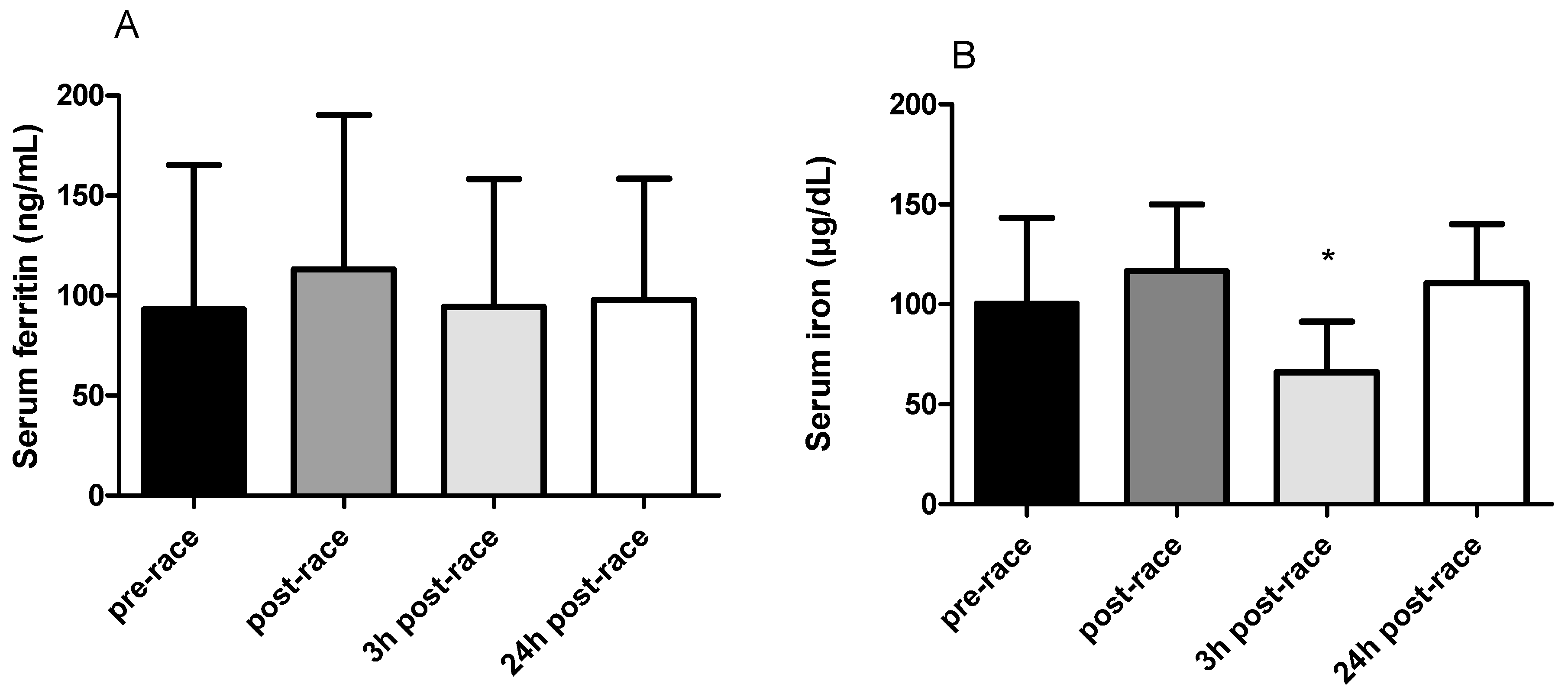
3.2. Serum Ferritin and Iron Concentrations
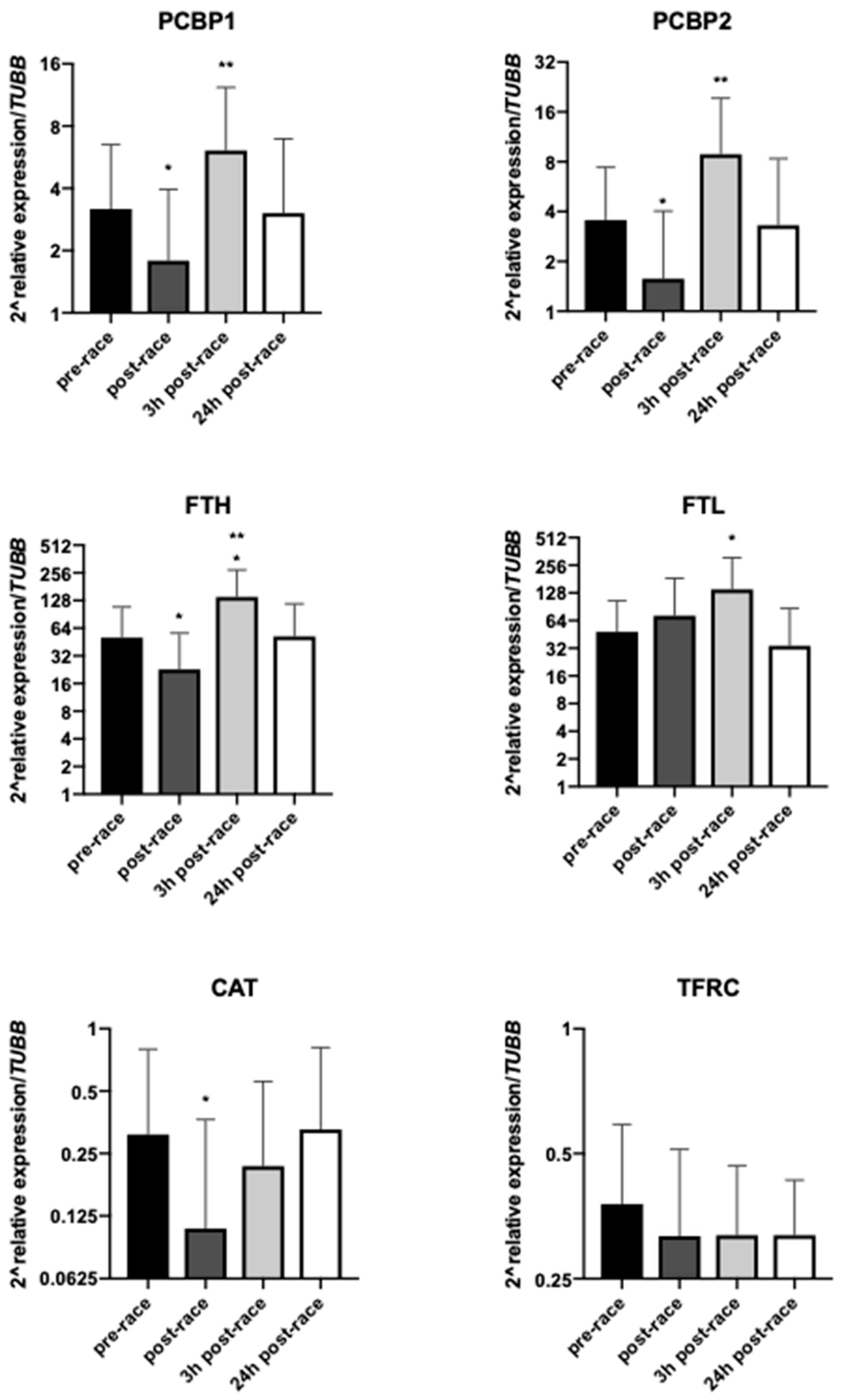
3.3. Effect of Exercise on mRNA Levels of Selected Genes
3.4. Relationship Between Baseline Levels of Serum Iron and Ferritin, and Changes in mRNA Levels with Exercise
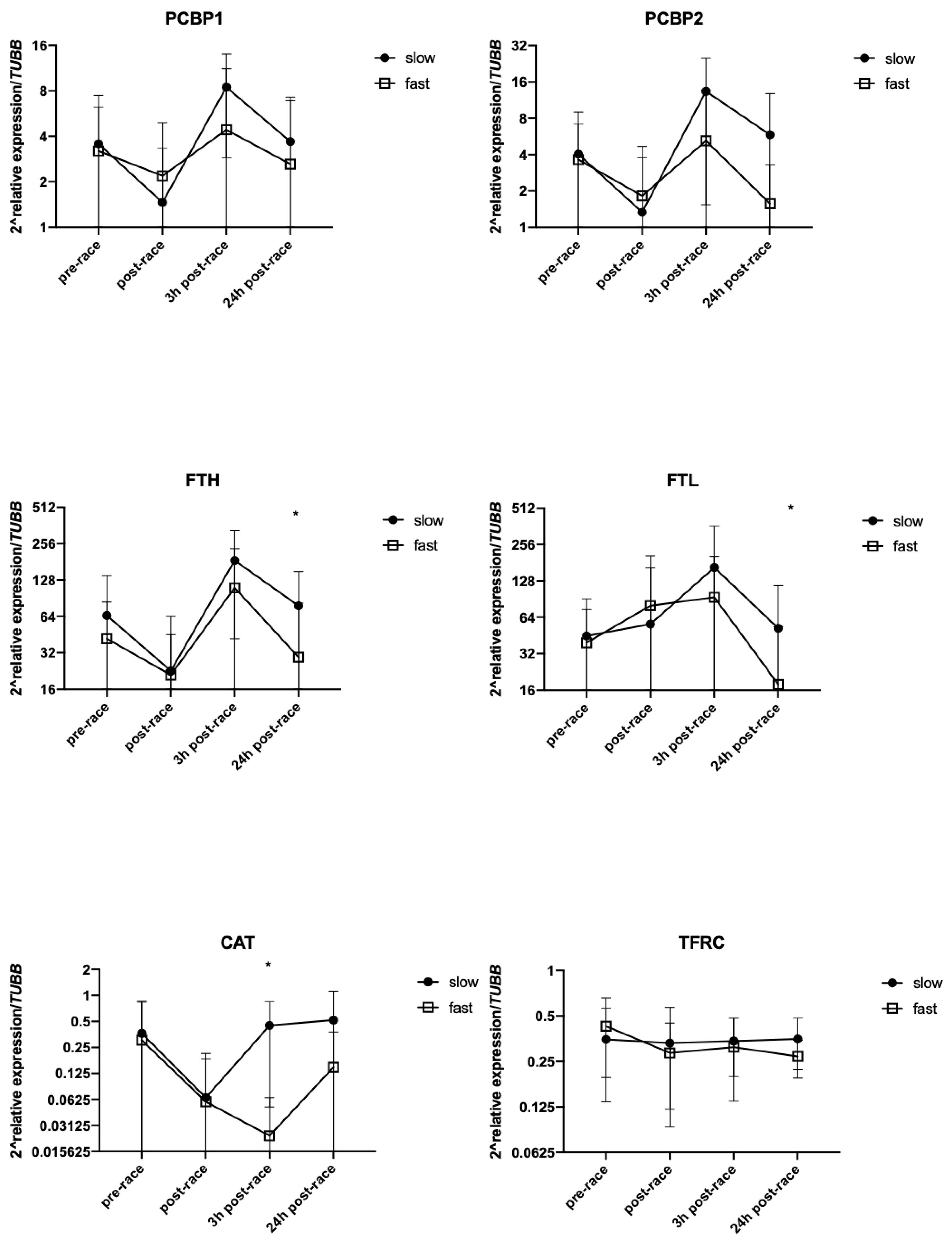
3.5. Effect of Running Pace on mRNA Levels of Selected Genes
4. Discussion
4.1. Changes in Serum Iron and Ferritin Concentrations
4.2. Changes in the mRNA of Genes Involved in Iron Metabolism
4.3. Relationship Between the mRNA of Genes Involved in Iron Metabolism and Running Speed
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Alaunyte, I.; Stojceska, V.; Plunkett, A. Iron and the Female Athlete: A Review of Dietary Treatment Methods for Improving Iron Status and Exercise Performance. J. Int. Soc. Sports Nutr. 2015, 12, 38. [Google Scholar] [CrossRef] [PubMed]
- Clénin, G.E.; Cordes, M.; Huber, A.; Schumacher, Y.; Noack, P.; Scales, J.; Kriemler, S. Iron Deficiency in Sports—Definition, Influence on Performance and Therapy. Schweizerische Zeitschrift für Sportmedizin und Sporttraumatologie 2016, 64, 6–18. [Google Scholar] [CrossRef] [PubMed]
- Sandström, G.; Börjesson, M.; Rödjer, S. Iron Deficiency in Adolescent Female Athletes-Is Iron Status Affected by Regular Sporting Activity? Clin. J. Sport Med. 2012, 22, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Mettler, S.; Zimmermann, M.B. Iron Excess in Recreational Marathon Runners. Eur. J. Clin. Nutr. 2010, 64, 490–494. [Google Scholar] [CrossRef] [PubMed]
- Bárány, P. Inflammation, Serum C-Reactive Protein, and Erythropoietin Resistance. Nephrol. Dial. Transplant. 2001, 16, 224–227. [Google Scholar] [CrossRef] [PubMed]
- Borghini, A.; Giardini, G.; Tonacci, A.; Mastorci, F.; Mercuri, A.; Mrakic-Sposta, S.; Moretti, S.; Andreassi, M.G.; Pratali, L. Chronic and Acute Effects of Endurance Training on Telomere Length. Mutagenesis 2015, 30, 711–716. [Google Scholar] [CrossRef] [PubMed]
- Dalle Carbonare, L.; Manfredi, M.; Caviglia, G.; Conte, E.; Robotti, E.; Marengo, E.; Cheri, S.; Zamboni, F.; Gabbiani, D.; Deiana, M.; et al. Can Half-Marathon Affect Overall Health? The Yin-Yang of Sport. J. Proteomics 2018, 170, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Kell, D.B.; Pretorius, E. Serum Ferritin Is an Important Inflammatory Disease Marker, as It Is Mainly a Leakage Product from Damaged Cells. Metallomics 2014, 6, 748–773. [Google Scholar] [CrossRef]
- Peeling, P.; Dawson, B.; Goodman, C.; Landers, G.; Wiegerinck, E.T.; Swinkels, D.W.; Trinder, D. Cumulative Effects of Consecutive Running Sessions on Hemolysis, Inflammation and Hepcidin Activity. Eur. J. Appl. Physiol. 2009, 106, 51–59. [Google Scholar] [CrossRef]
- Spanidis, Y.; Priftis, A.; Stagos, D.; Stravodimos, G.A.; Leonidas, D.D.; Spandidos, D.A.; Tsatsakis, A.M.; Kouretas, D. Oxidation of Human Serum Albumin Exhibits Inter-Individual Variability after an Ultra-Marathon Mountain Race. Exp. Ther. Med. 2017, 13, 2382–2390. [Google Scholar] [CrossRef]
- Tan, D.; Dawson, B.; Peeling, P. Hemolytic Effects of a Football-Specific Training Session in Elite Female Players. Int. J. Sports Physiol. Perform. 2012, 7, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Telford, R.D.; Sly, G.J.; Hahn, A.G.; Cunningham, R.B.; Bryant, C.; Smith, J.A. Footstrike Is the Major Cause of Hemolysis during Running. J. Appl. Physiol. 2015, 94, 38–42. [Google Scholar] [CrossRef] [PubMed]
- Terink, R.; ten Haaf, D.; Bongers, C.W.G.; Balvers, M.G.J.; Witkamp, R.F.; Mensink, M.; Eijsvogels, T.M.H.; Gunnewiek, J.M.T.K.; Hopman, M.T.E. Changes in Iron Metabolism during Prolonged Repeated Walking Exercise in Middle-Aged Men and Women. Eur. J. Appl. Physiol. 2018, 118, 2349–2357. [Google Scholar] [CrossRef] [PubMed]
- Jastrzȩbski, Z.; Zychowska, M.; Radzimiński, Ł.; Konieczna, A.; Kortas, J. Damage to Liver and Skeletal Muscles in Marathon Runners during a 100 Km Run with Regard to Age and Running Speed. J. Hum. Kinet. 2015, 45, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Orino, K.; Lehman, L.; Tsuji, Y.; Ayaki, H.; Torti, S.V.; Torti, F.M. Ferritin and the Response to Oxidative Stress. Biochem. J. 2015, 357, 241–247. [Google Scholar] [CrossRef]
- Peeling, P.; Dawson, B.; Goodman, C.; Landers, G.; Trinder, D. Athletic Induced Iron Deficiency: New Insights into the Role of Inflammation, Cytokines and Hormones. Eur. J. Appl. Physiol. 2008, 103, 381–391. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Bencze, K.Z.; Stemmler, T.L.; Philpott, C.C. A Cytosolic Iron Chaperone That Delivers Iron to Ferritin. Science 2008, 320, 1207–1210. [Google Scholar] [CrossRef] [Green Version]
- Völgyi, E.; Tylavsky, F.A.; Lyytikäinen, A.; Suominen, H.; Alén, M.; Cheng, S. Assessing Body Composition with DXA and Bioimpedance: Effects of Obesity, Physical Activity, and Age. Obesity 2008, 16, 700–705. [Google Scholar] [CrossRef]
- Chomczynski, P.; Sacchi, N. Single-Step Method of RNA Isolation by Acid Guanidinium Extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Dill, D.B.; Costill, D.L. Calculation of Percentage Changes in Volumes of Blood, Plasma, and Red Cells in Dehydration. J. Appl. Physiol. 1974, 37, 247–248. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing Real-Time PCR Data by the Comparative CTmethod. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Atamaniuk, J.; Stuhlmeier, K.M.; Vidotto, C.; Tschan, H.; Dossenbach-Glaninger, A.; Mueller, M.M. Effects of Ultra-Marathon on Circulating DNA and MRNA Expression of pro- and Anti-Apoptotic Genes in Mononuclear Cells. Eur. J. Appl. Physiol. 2008, 104, 711–717. [Google Scholar] [CrossRef] [PubMed]
- Roecker, L.; Meier-Buttermilch, R.; Brechtel, L.; Nemeth, E.; Ganz, T. Iron-Regulatory Protein Hepcidin Is Increased in Female Athletes after a Marathon. Eur. J. Appl. Physiol. 2005, 95, 569–571. [Google Scholar] [CrossRef] [PubMed]
- Chiu, Y.H.; Lai, J.I.; Wang, S.H.; How, C.K.; Li, L.H.; Kao, W.F.; Yang, C.C.; Chen, R.J. Early Changes of the Anemia Phenomenon in Male 100-Km Ultramarathoners. J. Chin. Med. Assoc. 2015, 78, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Peeling, P.; Sim, M.; Badenhorst, C.E.; Dawson, B.; Govus, A.D.; Abbiss, C.R.; Swinkels, D.W.; Trinder, D. Iron Status and the Acute Post-Exercise Hepcidin Response in Athletes. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Buchman, A.L.; Dunn, J.K.; Keen, C.; Commisso, J.; Dunn, J.K.; Killip, D.; Dunn, J.K.; Ou, C.N.; Rognerud, C.L.; Dunn, J.K.; et al. The Effect of a Marathon Run on Plasma and Urine Mineral and Metal Concentrations. J. Am. Coll. Nutr. 1998, 17, 124–127. [Google Scholar] [CrossRef] [PubMed]
- Duca, L.; Da Ponte, A.; Cozzi, M.; Carbone, A.; Pomati, M.; Nava, I.; Cappellini, M.D.; Fiorelli, G. Changes in Erythropoiesis, Iron Metabolism and Oxidative Stress after Half-Marathon. Intern. Emerg. Med. 2006, 1, 30–34. [Google Scholar] [CrossRef]
- Weight, L.M.; Byrne, M.J.; Jacobs, P. Haemolytic Effects of Exercise. Clin. J. Sport Med. 2006, 2, 74. [Google Scholar] [CrossRef]
- Galanello, R.; Giagu, N.; Barella, S.; Maccioni, L.; Origa, R. Lack of Correlation between Serum Ferritin and Liver Iron Concentration in Beta-Zero Thalassemia Intermedia. Blood 2004, 104, 3620. [Google Scholar]
- Huang, B.W.; Miyazawa, M.; Tsuji, Y. Distinct Regulatory Mechanisms of the Human Ferritin Gene by Hypoxia and Hypoxia Mimetic Cobalt Chloride at the Transcriptional and Post-Transcriptional Levels. Cell. Signal. 2014, 26, 2702–2709. [Google Scholar] [CrossRef]
- Mieszkowski, J.; Kochanowicz, M.; Ma, B.; Kochanowicz, A.; Grzybkowska, A.; Anczykowska, K.; Sawicki, P.; Borkowska, A.; Niespodzinski, B.B.; Antosiewicz, J. Ferritin Genes Overexpression in PBMC and a Rise in Exercise Performance as an Adaptive Response to Ischaemic Preconditioning in Young Men. BioMed Res. Int. 2019, 2019, 9576876. [Google Scholar] [CrossRef] [PubMed]
- Maqueda, M.; Roca, E.; Brotons, D.; Soria, J.M.; Perera, A. Affected Pathways and Transcriptional Regulators in Gene Expression Response to an Ultra-Marathon Trail: Global and Independent Activity Approaches. PLoS ONE 2017, 12, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Leidgens, S.; Bullough, K.Z.; Shi, H.; Li, F.; Shakoury-Elizeh, M.; Yabe, T.; Subramanian, P.; Hsu, E.; Natarajan, N.; Nandal, A.; et al. Each Member of the Poly-r(C)-Binding Protein 1 (PCBP) Family Exhibits Iron Chaperone Activity toward Ferritin. J. Biol. Chem. 2013, 288, 17791–17802. [Google Scholar] [CrossRef] [PubMed]
- Sureda, A.; del Mar Bibiloni, M.; Julibert, A.; Bouzas, C.; Argelich, E.; Llompart, I.; Pons, A.; Tur, J.A. Adherence to the Mediterranean Diet and Inflammatory Markers. Nutrients 2018, 10, 62. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Participant’s Characteristic | |
|---|---|
| Baseline (n = 24) | |
| Age (years) | 48.8 ± 6.5 |
| Body mass (kg) | 80.1 ± 8.5 |
| Height (cm) | 178.7 ± 5.3 |
| BMI (kg/m2) | 25.1 ± 2.3 |
| PBF (%) | 15.5 ± 5.0 |
| Pace during the run (km/h) | 10.9 ± 1.4 |
| Training units per week | Between 1 and 7 |
| Training regimen (km/week) | Between 20 and 115 |
| Training experience (years) | Between 4 and 24 |
| Number of finished marathons | Between 1 and 62 |
| Pre-Race | Post-Race | 3 h Post-Race | 24 h Post-Race | |
|---|---|---|---|---|
| %Delta PV | - | 1.05% | 11.58% * | 12.43% * |
| - | p > 0.9999 | p < 0.0001 | p < 0.0001 | |
| Hemoglobin (g/dL) | 14.94 ± 0.85 | 14.78 ± 1.36 | 12.63 ± 1.06 * | 12.41 ± 0.96 * |
| - | p = 0.9545 | p < 0.0001 | p < 0.0001 | |
| RBC (×1012/L) | 4.97 ± 0.33 | 4.91 ± 0.44 | 4.19 ± 0.35 * | 4.13 ± 0.32 * |
| - | p = 0.9445 | p < 0.0001 | p < 0.0001 | |
| Hematocrite (%) | 43.33 ± 2.17 | 42.61 ± 3.96 | 35.91 ± 3.09 * | 35.91 ± 2.58 * |
| - | p > 0.9999 | p < 0.0001 | p < 0.0001 | |
| Reticulocytes (×109/L) | 59.44 ± 11.6 | 61.78 ± 13.77 | 49.38 ± 11.67 * | 46.63 ± 12.52 * |
| - | p = 0.9088 | p = 0.0261 | p = 0.0027 | |
| White blood cells (×109/L) | 5.27 ± 1.22 | 14.74 ± 3.45 * | 12.02 ± 2.07 * | 6.35 ± 1.54 |
| - | p < 0.0001 | p < 0.0001 | p = 0.9970 | |
| Neutrophils (×109/L) | 2.84 ± 0.9 | 12.2 ± 3.11 * | 9.82 ± 1.97 * | 3.58 ± 1.63 |
| - | p < 0.0001 | p < 0.0001 | p > 0.9999 | |
| Lymphocytes (×109/L) | 1.7 ± 0.31 | 1.48 ± 0.47 * | 1.26 ± 0.28 | 1.94 ± 0.45 |
| - | p = 0.1485 | p = 0.0007 | p = 0.9166 | |
| Monocytes (×109/L) | 0.47 ± 0.14 | 0.95 ± 0.26 * | 0.89 ± 0.21 * | 0.56 ± 0.13 |
| - | p < 0.0001 | p < 0.0001 | p > 0.9999 | |
| Eosinophils (×109/L) | 0.22 ± 0.14 | 0.04 ± 0.04 * | 0.01 ± 0.01 * | 0.19 ± 0.12 |
| - | p < 0.0001 | p < 0.0001 | p > 0.9999 | |
| Basophils (×109/L) | 0.04 ± 0.02 | 0.06 ± 0.02 * | 0.03 ± 0.01 | 0.04 ± 0.01 |
| - | p = 0.0032 | p > 0.9999 | p > 0.9999 | |
| CRP (mg/L) | 1.4 ± 3.7 | 1.31 ± 3.26 | 1.87 ± 3.12 | 9.79 ± 7.28 * |
| - | p > 0.9999 | p = 0.0807 | p < 0.0001 | |
| Uric acid (mg/dL) | 5.26 ± 1.08 | 5.9 ± 1.07 * | 5.71 ± 1.03 * | 4.86 ± 0.96 * |
| - | p < 0.0001 | p = 0.0007 | p = 0.0113 | |
| Creatine kinase (U/L) | 171.56 ± 68.52 | 569.55 ± 490.71 * | 871.04 ± 900.02 * | 1410.66 ± 1444.06 * |
| - | p < 0.0001 | p < 0.0001 | p < 0.0001 |
| Slow and Fast Groups Characteristics | ||
|---|---|---|
| Slow (n = 12) | Fast (n = 12) | |
| Pace during the run (km/h) | 10.04 ± 0.52 | 12.18 ± 0.71 * |
| Age (years) | 53.58 ± 5.45 | 44.25 ± 3.49 * |
| BMI (kg/m2) | 26.28 ± 1.88 | 23.83 ± 1.95 * |
| Training units per week | 3.00 ± 1.04 | 4.58 ± 1.26 |
| Training regimen (km/week) | 43.58 ± 16.53 | 71.27 ± 25.22 |
| Training experience (years) | 10.83 ± 6.90 | 8.08 ± 5.02 |
| Number of finished marathons | 16.83 ± 20.85 | 13.58 ± 9.18 |
| Baseline iron level (µg/dL) | 112.75 ± 40.54 | 92.92 ± 42.44 |
| Baseline ferritin level (ng/mL) | 93.41 ± 36.58 | 88.18 ± 98.29 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grzybkowska, A.; Anczykowska, K.; Ratkowski, W.; Aschenbrenner, P.; Antosiewicz, J.; Bonisławska, I.; Żychowska, M. Changes in Serum Iron and Leukocyte mRNA Levels of Genes Involved in Iron Metabolism in Amateur Marathon Runners—Effect of the Running Pace. Genes 2019, 10, 460. https://0-doi-org.brum.beds.ac.uk/10.3390/genes10060460
Grzybkowska A, Anczykowska K, Ratkowski W, Aschenbrenner P, Antosiewicz J, Bonisławska I, Żychowska M. Changes in Serum Iron and Leukocyte mRNA Levels of Genes Involved in Iron Metabolism in Amateur Marathon Runners—Effect of the Running Pace. Genes. 2019; 10(6):460. https://0-doi-org.brum.beds.ac.uk/10.3390/genes10060460
Chicago/Turabian StyleGrzybkowska, Agata, Katarzyna Anczykowska, Wojciech Ratkowski, Piotr Aschenbrenner, Jędrzej Antosiewicz, Iwona Bonisławska, and Małgorzata Żychowska. 2019. "Changes in Serum Iron and Leukocyte mRNA Levels of Genes Involved in Iron Metabolism in Amateur Marathon Runners—Effect of the Running Pace" Genes 10, no. 6: 460. https://0-doi-org.brum.beds.ac.uk/10.3390/genes10060460