The Role of Salmonella Genomic Island 4 in Metal Tolerance of Salmonella enterica Serovar I 4,[5],12:i:- Pork Outbreak Isolate USDA15WA-1

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Mutant Construction, and Primers
2.2. Conjugation Studies
2.3. DNA Sequencing and Assembly
2.4. Phenotypic Microarrays
2.5. Transcriptional Analysis of USDA15WA-1 in Response to Copper Exposure
2.6. Swine Study
2.7. Microbiota Analyses
3. Results and Discussion
3.1. Salmonella Genomic Island 4 Transferred by Conjugation from Strain USDA15WA-1 to Serovar Typhimurium
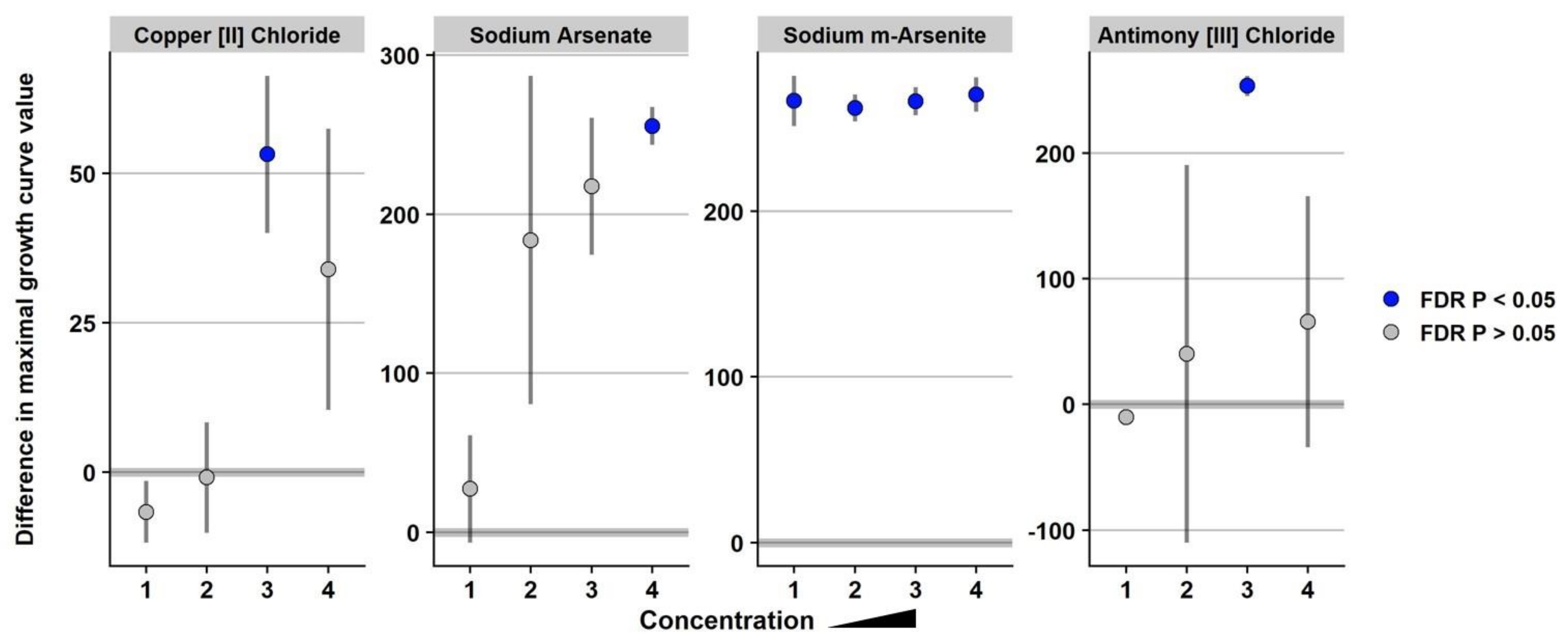
3.2. The Presence of SGI-4 in Strain USDA15WA-1 Enhanced Tolerance to Multiple Metals
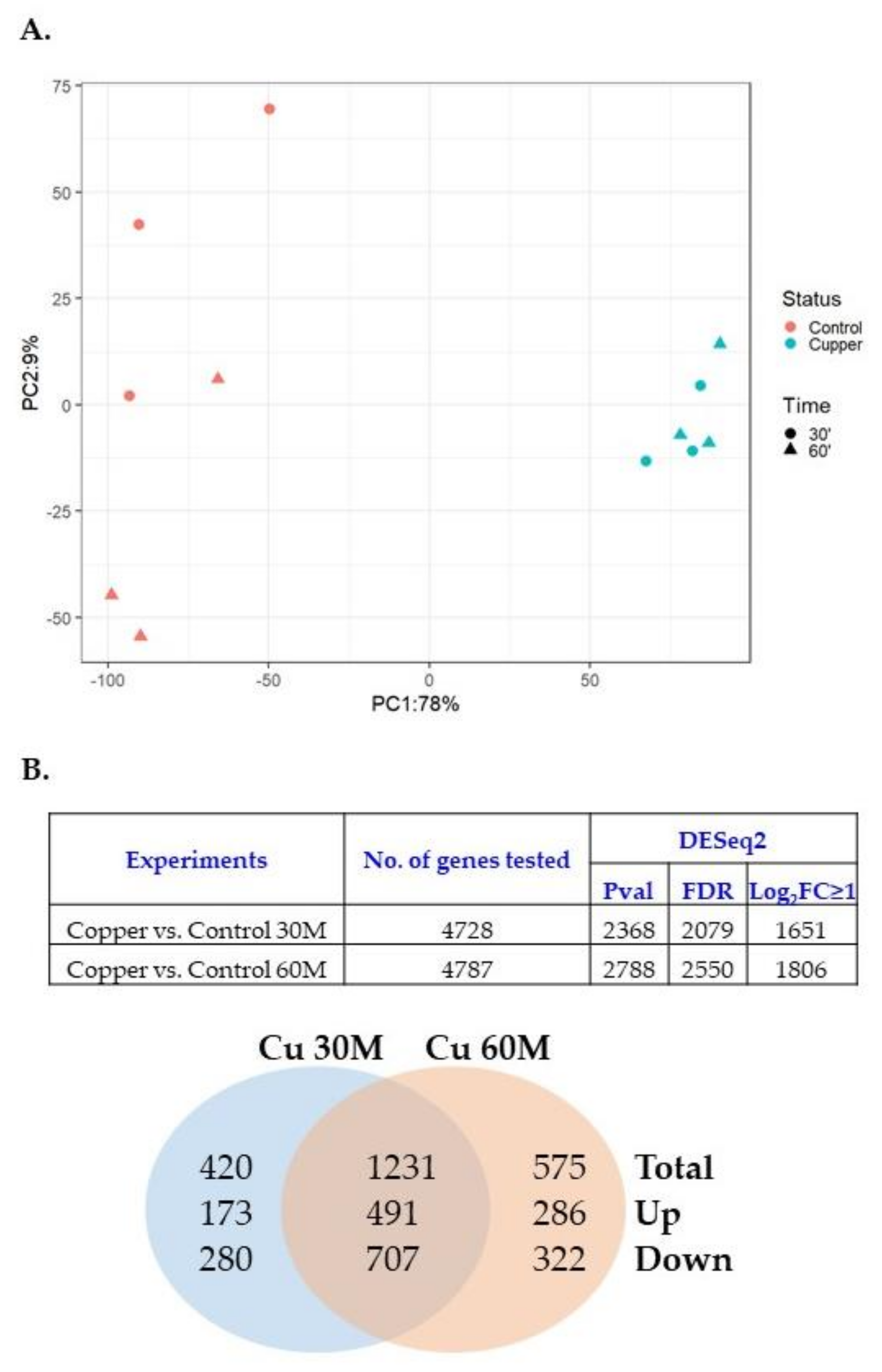
3.3. Copper Induced Gene Expression of Metal Tolerance Genes on Genomic Islands
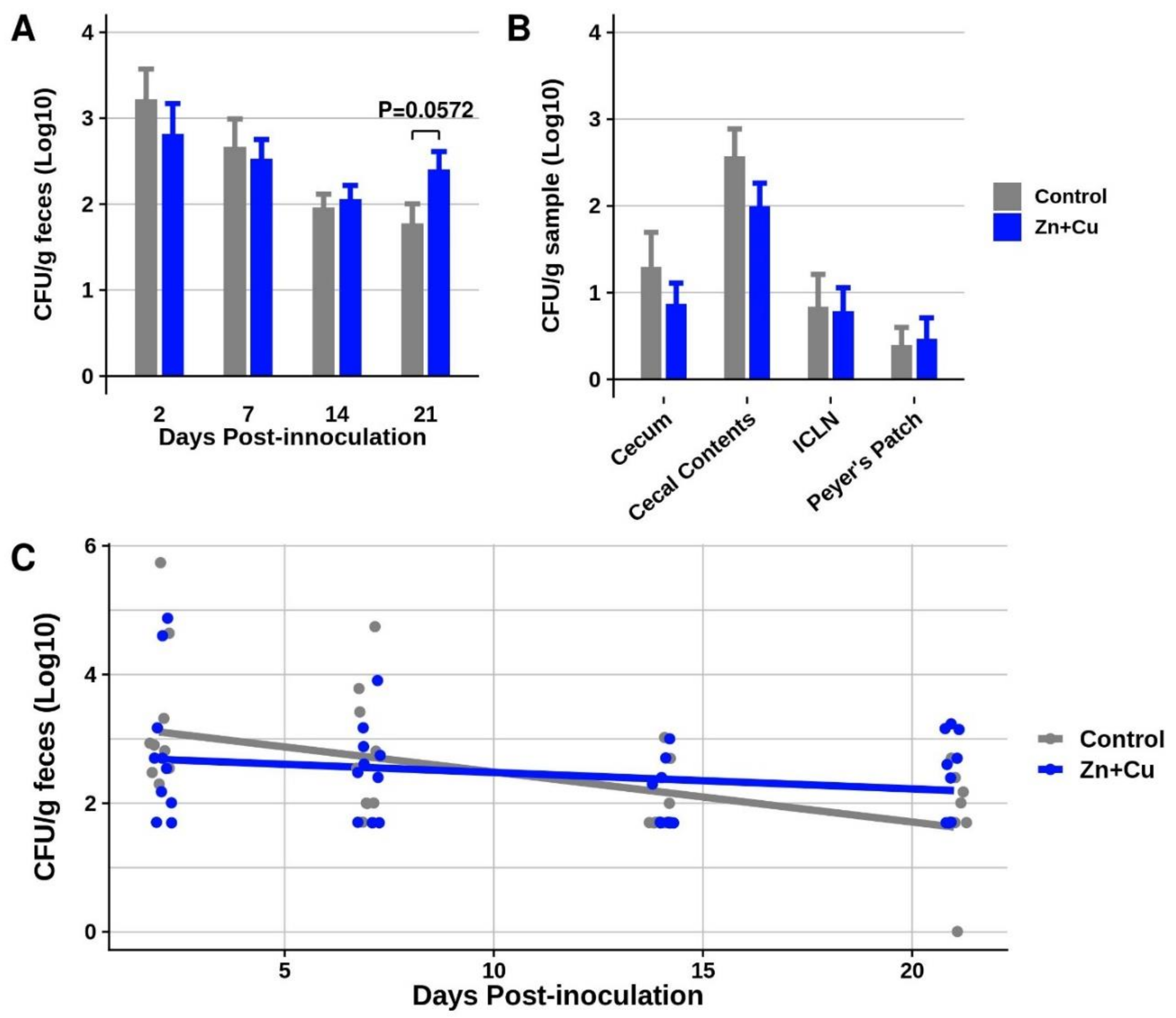
3.4. In-Feed Supplementation with Elevated Concentrations of Zn and Cu as an Antimicrobial Slowed the Decline of Salmonella I 4,[5],12:i:- Fecal Shedding in Pigs
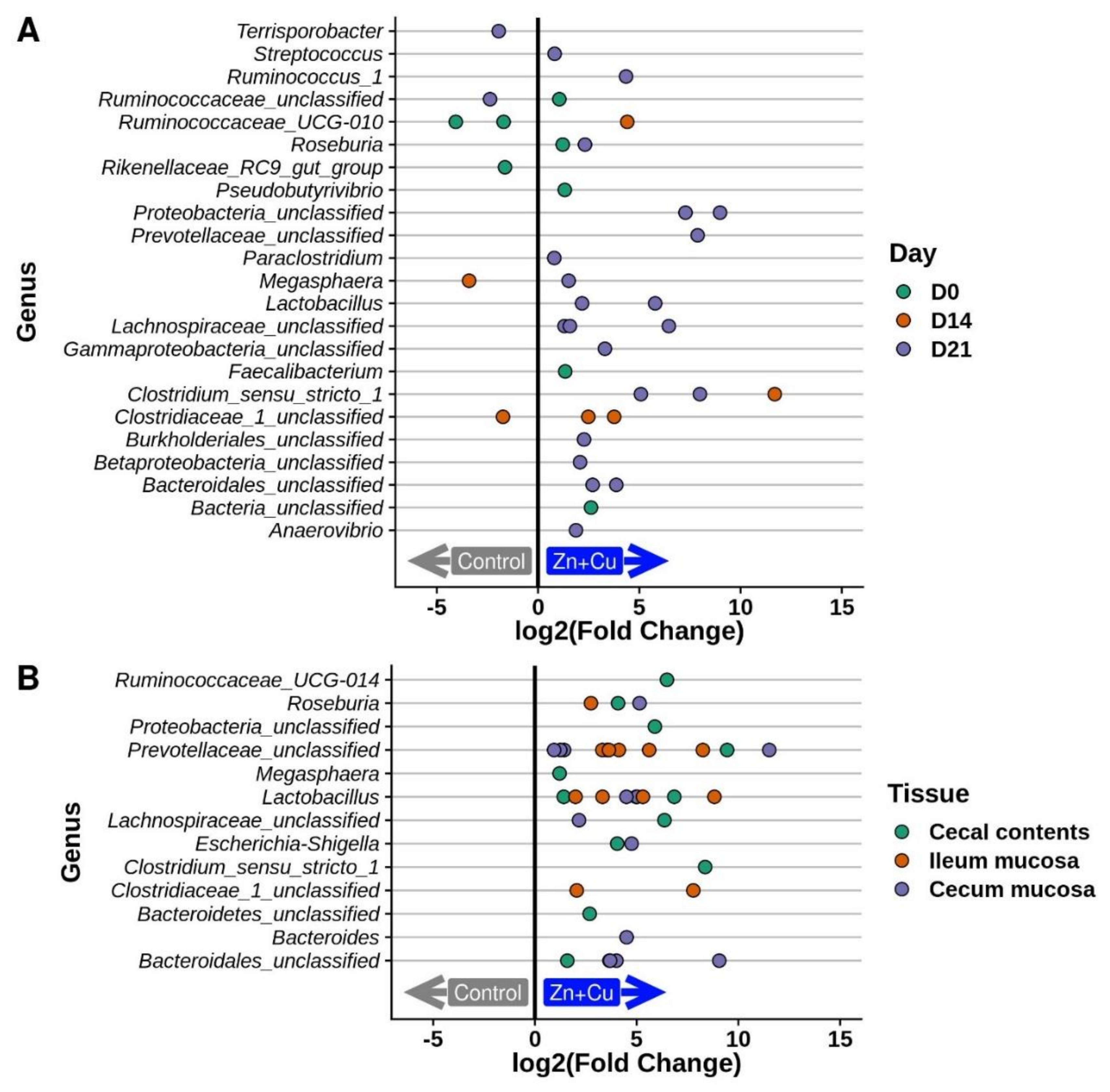
3.5. Alterations in the Porcine Microbiota in Response to In-Feed Zn/Cu Treatment
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
Appendix B
References
- CDC. Salmonella. Available online: https://www.cdc.gov/salmonella/index.html (accessed on 12 May 2020).
- Boyen, F.; Haesebrouck, F.; Maes, D.; Van Immerseel, F.; Ducatelle, R.; Pasmans, F. Non-typhoidal Salmonella infections in pigs: A closer look at epidemiology, pathogenesis and control. Vet. Microbiol. 2008, 130, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Heredia, N.; Garcia, S. Animals as sources of food-borne pathogens: A review. Anim. Nutr. 2018, 4, 250–255. [Google Scholar] [CrossRef] [PubMed]
- CDC. National Outbreak Reporting System Dashboard. Available online: https://wwwn.cdc.gov/norsdashboard/ (accessed on 12 May 2020).
- Kawakami, V.M.; Bottichio, L.; Angelo, K.; Linton, N.; Kissler, B.; Basler, C.; Lloyd, J.; Inouye, W.; Gonzales, E.; Rietberg, K.; et al. Notes from the Field: Outbreak of Multidrug-Resistant Salmonella Infections Linked to Pork--Washington, 2015. MMWR Morb. Mortal. Wkly. Rep. 2016, 65, 379–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bearson, B.L.; Trachsel, J.M.; Holman, D.B.; Brunelle, B.W.; Sivasankaran, S.K.; Simmons, M.; Wasilenko, J.; Tillman, G.; Johnston, J.J.; Bearson, S.M.D. Complete Genome Sequence of Multidrug-Resistant Salmonella enterica Serovar I 4,[5],12:i:- 2015 U.S. Pork Outbreak Isolate USDA15WA-1. Microbiol. Resour. Announc. 2019, 8, e00791-19. [Google Scholar] [CrossRef] [Green Version]
- Mastrorilli, E.; Pietrucci, D.; Barco, L.; Ammendola, S.; Petrin, S.; Longo, A.; Mantovani, C.; Battistoni, A.; Ricci, A.; Desideri, A.; et al. A Comparative Genomic Analysis Provides Novel Insights Into the Ecological Success of the Monophasic Salmonella Serovar 4,[5],12:i. Front. Microbiol. 2018, 9, 715. [Google Scholar] [CrossRef]
- Switt, A.I.; Soyer, Y.; Warnick, L.D.; Wiedmann, M. Emergence, distribution, and molecular and phenotypic characteristics of Salmonella enterica serotype 4,5,12:i. Foodborne Pathog. Dis. 2009, 6, 407–415. [Google Scholar] [CrossRef] [Green Version]
- CDC. National Antimicrobial Resistance Monitoring System for Enteric Bacteria (NARMS): Human Isolates Surveillance Report for 2015 (Final Report); U.S. Department of Health and Human Services, CDC: Atlanta, GA, USA, 2018.
- Garcia, P.; Malorny, B.; Rodicio, M.R.; Stephan, R.; Hachler, H.; Guerra, B.; Lucarelli, C. Horizontal Acquisition of a Multidrug-Resistance Module (R-type ASSuT) Is Responsible for the Monophasic Phenotype in a Widespread Clone of Salmonella Serovar 4,[5],12:i. Front. Microbiol. 2016, 7, 680. [Google Scholar] [CrossRef] [Green Version]
- Petrovska, L.; Mather, A.E.; AbuOun, M.; Branchu, P.; Harris, S.R.; Connor, T.; Hopkins, K.L.; Underwood, A.; Lettini, A.A.; Page, A.; et al. Microevolution of Monophasic Salmonella Typhimurium during Epidemic, United Kingdom, 2005–2010. Emerg. Infect. Dis. 2016, 22, 617–624. [Google Scholar] [CrossRef] [Green Version]
- Arai, N.; Sekizuka, T.; Tamamura, Y.; Tanaka, K.; Barco, L.; Izumiya, H.; Kusumoto, M.; Hinenoya, A.; Yamasaki, S.; Iwata, T.; et al. Phylogenetic Characterization of Salmonella enterica Serovar Typhimurium and Its Monophasic Variant Isolated from Food Animals in Japan Revealed Replacement of Major Epidemic Clones in the Last 4 Decades. J. Clin. Microbiol. 2018, 56, e01758-17. [Google Scholar] [CrossRef] [Green Version]
- Mourao, J.; Novais, C.; Machado, J.; Peixe, L.; Antunes, P. Metal tolerance in emerging clinically relevant multidrug-resistant Salmonella enterica serotype 4,[5],12:i:- clones circulating in Europe. Int. J. Antimicrob. Agents 2015, 45, 610–616. [Google Scholar] [CrossRef]
- Medardus, J.J.; Molla, B.Z.; Nicol, M.; Morrow, W.M.; Rajala-Schultz, P.J.; Kazwala, R.; Gebreyes, W.A. In-feed use of heavy metal micronutrients in U.S. swine production systems and its role in persistence of multidrug-resistant Salmonellae. Appl. Environ. Microbiol. 2014, 80, 2317–2325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bearson, B.L.; Bearson, S.M.; Uthe, J.J.; Dowd, S.E.; Houghton, J.O.; Lee, I.; Toscano, M.J.; Lay, D.C., Jr. Iron regulated genes of Salmonella enterica serovar Typhimurium in response to norepinephrine and the requirement of fepDGC for norepinephrine-enhanced growth. Microbes Infect. 2008, 10, 807–816. [Google Scholar] [CrossRef]
- Doublet, B.; Douard, G.; Targant, H.; Meunier, D.; Madec, J.Y.; Cloeckaert, A. Antibiotic marker modifications of lambda Red and FLP helper plasmids, pKD46 and pCP20, for inactivation of chromosomal genes using PCR products in multidrug-resistant strains. J. Microbiol. Methods 2008, 75, 359–361. [Google Scholar] [CrossRef] [PubMed]
- Shippy, D.C.; Bearson, B.L.; Holman, D.B.; Brunelle, B.W.; Allen, H.K.; Bearson, S.M.D. Porcine Response to a Multidrug-Resistant Salmonella enterica serovar I 4,[5],12:i:- Outbreak Isolate. Foodborne Pathog. Dis. 2018, 15, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Bearson, S.M.; Bearson, B.L.; Rasmussen, M.A. Identification of Salmonella enterica serovar Typhimurium genes important for survival in the swine gastric environment. Appl. Environ. Microbiol. 2006, 72, 2829–2836. [Google Scholar] [CrossRef] [Green Version]
- Bushnell, B. BBMap. Available online: https://sourceforge.net/projects/bbmap/ (accessed on 27 October 2020).
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [Green Version]
- Team, R.C. R: A Language and Environment for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 7 August 2020).
- Vaas, L.A.I.; Sikorski, J.; Hofner, B.; Fiebig, A.; Buddruhs, N.; Klenk, H.-P.; Goeker, M. opm: An R package for analysing OmniLog((R)) phenotype microarray data. Bioinformatics 2013, 29, 1823–1824. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, S.; Otto, C.; Kurtz, S.; Sharma, C.M.; Khaitovich, P.; Vogel, J.; Stadler, P.F.; Hackermuller, J. Fast mapping of short sequences with mismatches, insertions and deletions using index structures. PLoS Comput. Biol. 2009, 5, e1000502. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gene Ontology Consortium. The Gene Ontology Resource: 20 years and still GOing strong. Nucleic Acids Res. 2019, 47, D330–D338. [Google Scholar] [CrossRef] [Green Version]
- Jones, P.; Binns, D.; Chang, H.Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. InterProScan 5: Genome-scale protein function classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexa, A.; Rahnenfuhrer, J. topGO: Enrichment Analysis for Gene Ontology; R Package Version 2.40.0. 2020. Available online: https://rdrr.io/bioc/topGO/ (accessed on 27 October 2020).
- Bearson, B.L.; Bearson, S.M.; Lee, I.S.; Brunelle, B.W. The Salmonella enterica serovar Typhimurium QseB response regulator negatively regulates bacterial motility and swine colonization in the absence of the QseC sensor kinase. Microb. Pathog. 2010, 48, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Bearson, S.M.; Bearson, B.L.; Loving, C.L.; Allen, H.K.; Lee, I.; Madson, D.; Kehrli, M.E., Jr. Prophylactic Administration of Vector-Encoded Porcine Granulocyte-Colony Stimulating Factor Reduces Salmonella Shedding, Tonsil Colonization, and Microbiota Alterations of the Gastrointestinal Tract in Salmonella-Challenged Swine. Front. Vet. Sci. 2016, 3, 66. [Google Scholar] [CrossRef] [Green Version]
- Bates, D.; Machler, M.; Bolker, B.M.; Walker, S.C. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest Package: Tests in Linear Mixed Effects Models. J. Stat. Softw. 2017, 82, 26. [Google Scholar] [CrossRef] [Green Version]
- Trachsel, J.; Briggs, C.; Gabler, N.K.; Allen, H.K.; Loving, C.L. Dietary Resistant Potato Starch Alters Intestinal Microbial Communities and Their Metabolites, and Markers of Immune Regulation and Barrier Function in Swine. Front. Immunol. 2019, 10, 1381. [Google Scholar] [CrossRef] [Green Version]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. vegan: Community Ecology Package, 2.5.6. 2019. Available online: https://rdrr.io/cran/vegan/ (accessed on 27 October 2020).
- McMurdie, P.J.; Holmes, S. phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Branchu, P.; Charity, O.J.; Bawn, M.; Thilliez, G.; Dallman, T.J.; Petrovska, L.; Kingsley, R.A. SGI-4 in Monophasic Salmonella Typhimurium ST34 Is a Novel ICE That Enhances Resistance to Copper. Front. Microbiol. 2019, 10, 1118. [Google Scholar] [CrossRef] [Green Version]
- Arai, N.; Sekizuka, T.; Tamamura, Y.; Kusumoto, M.; Hinenoya, A.; Yamasaki, S.; Iwata, T.; Watanabe-Yanai, A.; Kuroda, M.; Akiba, M. Salmonella Genomic Island 3 Is an Integrative and Conjugative Element and Contributes to Copper and Arsenic Tolerance of Salmonella enterica. Antimicrob. Agents Chemother. 2019, 63, e00429-19. [Google Scholar] [CrossRef] [Green Version]
- Chalmers, G.; Rozas, K.M.; Amachawadi, R.G.; Scott, H.M.; Norman, K.N.; Nagaraja, T.G.; Tokach, M.D.; Boerlin, P. Distribution of the pco Gene Cluster and Associated Genetic Determinants among Swine Escherichia coli from a Controlled Feeding Trial. Genes 2018, 9, 504. [Google Scholar] [CrossRef] [Green Version]
- Osman, D.; Cavet, J.S. Metal sensing in Salmonella: Implications for pathogenesis. Adv. Microb. Physiol. 2011, 58, 175–232. [Google Scholar] [CrossRef]
- Macomber, L.; Imlay, J.A. The iron-sulfur clusters of dehydratases are primary intracellular targets of copper toxicity. Proc. Natl. Acad. Sci. USA 2009, 106, 8344–8349. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, M.; Udagedara, S.R.; Sze, C.M.; Ryan, T.M.; Howlett, G.J.; Xiao, Z.; Wedd, A.G. PcoE—A metal sponge expressed to the periplasm of copper resistance Escherichia coli. Implication of its function role in copper resistance. J. Inorg. Biochem. 2012, 115, 186–197. [Google Scholar] [CrossRef]
- Pontel, L.B.; Scampoli, N.L.; Porwollik, S.; Checa, S.K.; McClelland, M.; Soncini, F.C. Identification of a Salmonella ancillary copper detoxification mechanism by a comparative analysis of the genome-wide transcriptional response to copper and zinc excess. Microbiology 2014, 160, 1659–1669. [Google Scholar] [CrossRef] [Green Version]
- Lopez, C.; Checa, S.K.; Soncini, F.C. CpxR/CpxA Controls scsABCD Transcription to Counteract Copper and Oxidative Stress in Salmonella enterica Serovar Typhimurium. J. Bacteriol. 2018, 200, e00126-18. [Google Scholar] [CrossRef] [Green Version]
- Pezza, A.; Pontel, L.B.; Lopez, C.; Soncini, F.C. Compartment and signal-specific codependence in the transcriptional control of Salmonella periplasmic copper homeostasis. Proc. Natl. Acad. Sci. USA 2016, 113, 11573–11578. [Google Scholar] [CrossRef] [Green Version]
- Hahn, J.D.; Baker, D.H. Growth and plasma zinc responses of young pigs fed pharmacologic levels of zinc. J. Anim. Sci. 1993, 71, 3020–3024. [Google Scholar] [CrossRef]
- Hu, C.; Song, J.; You, Z.; Luan, Z.; Li, W. Zinc oxide-montmorillonite hybrid influences diarrhea, intestinal mucosal integrity, and digestive enzyme activity in weaned pigs. Biol. Trace Elem. Res. 2012, 149, 190–196. [Google Scholar] [CrossRef]
- Case, C.L.; Carlson, M.S. Effect of feeding organic and inorganic sources of additional zinc on growth performance and zinc balance in nursery pigs. J. Anim. Sci. 2002, 80, 1917–1924. [Google Scholar] [CrossRef]
- Kloubert, V.; Blaabjerg, K.; Dalgaard, T.S.; Poulsen, H.D.; Rink, L.; Wessels, I. Influence of zinc supplementation on immune parameters in weaned pigs. J. Trace Elem. Med. Biol. 2018, 49, 231–240. [Google Scholar] [CrossRef]
- Pieper, R.; Vahjen, W.; Neumann, K.; Van Kessel, A.G.; Zentek, J. Dose-dependent effects of dietary zinc oxide on bacterial communities and metabolic profiles in the ileum of weaned pigs. J. Anim. Physiol. Anim. Nutr. (Berl.) 2012, 96, 825–833. [Google Scholar] [CrossRef]
- Castillo, M.; Martin-Orue, S.M.; Taylor-Pickard, J.A.; Perez, J.F.; Gasa, J. Use of mannanoligosaccharides and zinc chelate as growth promoters and diarrhea preventative in weaning pigs: Effects on microbiota and gut function. J. Anim. Sci. 2008, 86, 94–101. [Google Scholar] [CrossRef]
- Liu, P.; Pieper, R.; Rieger, J.; Vahjen, W.; Davin, R.; Plendl, J.; Meyer, W.; Zentek, J. Effect of dietary zinc oxide on morphological characteristics, mucin composition and gene expression in the colon of weaned piglets. PLoS ONE 2014, 9, e91091. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Guo, Y. Supplemental zinc reduced intestinal permeability by enhancing occludin and zonula occludens protein-1 (ZO-1) expression in weaning piglets. Br. J. Nutr. 2009, 102, 687–693. [Google Scholar] [CrossRef] [Green Version]
- Hojberg, O.; Canibe, N.; Poulsen, H.D.; Hedemann, M.S.; Jensen, B.B. Influence of dietary zinc oxide and copper sulfate on the gastrointestinal ecosystem in newly weaned piglets. Appl. Environ. Microbiol. 2005, 71, 2267–2277. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Harper, A.F.; Estienne, M.J.; Webb, K.E., Jr.; McElroy, A.P.; Denbow, D.M. Growth performance and intestinal morphology responses in early weaned pigs to supplementation of antibiotic-free diets with an organic copper complex and spray-dried plasma protein in sanitary and nonsanitary environments. J. Anim. Sci. 2007, 85, 1302–1310. [Google Scholar] [CrossRef] [Green Version]
- Stahly, T.S.; Cromwell, G.L.; Monegue, H.J. Effects of the dietary inclusion of copper and(or) antibiotics on the performance of weanling pigs. J. Anim. Sci. 1980, 51, 1347–1351. [Google Scholar] [CrossRef]
- Perez, V.G.; Waguespack, A.M.; Bidner, T.D.; Southern, L.L.; Fakler, T.M.; Ward, T.L.; Steidinger, M.; Pettigrew, J.E. Additivity of effects from dietary copper and zinc on growth performance and fecal microbiota of pigs after weaning. J. Anim. Sci. 2011, 89, 414–425. [Google Scholar] [CrossRef] [PubMed]
- Namkung, H.; Gong, J.; Yu, H.; de Lange, C.F.M. Effect of pharmacological intakes of zinc and copper on growth performance, circulating cytokines and gut microbiota of newly weaned piglets challenged with coliform lipopolysaccharides. Can. J. Anim. Sci. 2006, 86, 511–522. [Google Scholar] [CrossRef]
- Mourao, J.; Marcal, S.; Ramos, P.; Campos, J.; Machado, J.; Peixe, L.; Novais, C.; Antunes, P. Tolerance to multiple metal stressors in emerging non-typhoidal MDR Salmonella serotypes: A relevant role for copper in anaerobic conditions. J. Antimicrob. Chemother. 2016, 71, 2147–2157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, X.; Luthje, F.L.; Qin, Y.; McDevitt, S.F.; Lutay, N.; Hobman, J.L.; Asiani, K.; Soncini, F.C.; German, N.; Zhang, S.; et al. Survival in amoeba—A major selection pressure on the presence of bacterial copper and zinc resistance determinants? Identification of a “copper pathogenicity island”. Appl. Microbiol. Biotechnol. 2015, 99, 5817–5824. [Google Scholar] [CrossRef]
- Mourao, J.; Rebelo, A.; Ribeiro, S.; Peixe, L.; Novais, C.; Antunes, P. Tolerance to arsenic contaminant among multidrug-resistant and copper-tolerant Salmonella successful clones is associated with diverse ars operons and genetic contexts. Environ. Microbiol. 2020, 22, 2829–2842. [Google Scholar] [CrossRef]
- Katouli, M.; Melin, L.; Jensen-Waern, M.; Wallgren, P.; Mollby, R. The effect of zinc oxide supplementation on the stability of the intestinal flora with special reference to composition of coliforms in weaned pigs. J. Appl. Microbiol. 1999, 87, 564–573. [Google Scholar] [CrossRef] [Green Version]
- Holm, A. Zinc oxide in treating E. coli diarrhea in pigs after weaning. Comp. Cont. Educ. Pract. 1996, 18, S26–S48. [Google Scholar]
- Mach, N.; Berri, M.; Estelle, J.; Levenez, F.; Lemonnier, G.; Denis, C.; Leplat, J.J.; Chevaleyre, C.; Billon, Y.; Dore, J.; et al. Early-life establishment of the swine gut microbiome and impact on host phenotypes. Environ. Microbiol. Rep. 2015, 7, 554–569. [Google Scholar] [CrossRef] [PubMed]
- Hiippala, K.; Jouhten, H.; Ronkainen, A.; Hartikainen, A.; Kainulainen, V.; Jalanka, J.; Satokari, R. The Potential of Gut Commensals in Reinforcing Intestinal Barrier Function and Alleviating Inflammation. Nutrients 2018, 10, 988. [Google Scholar] [CrossRef] [Green Version]
- Tamanai-Shacoori, Z.; Smida, I.; Bousarghin, L.; Loreal, O.; Meuric, V.; Fong, S.B.; Bonnaure-Mallet, M.; Jolivet-Gougeon, A. Roseburia spp.: A marker of health? Future Microbiol. 2017, 12, 157–170. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Kim, J.; Lee, J.S.; Rhee, S.K.; Kim, H. Characterization of the fecal microbiome in different swine groups by high-throughput sequencing. Anaerobe 2014, 28, 157–162. [Google Scholar] [CrossRef]
- Tinge, S.A.; Curtiss, R., 3rd. Isolation of the replication and partitioning regions of the Salmonella typhimurium virulence plasmid and stabilization of heterologous replicons. J. Bacteriol. 1990, 172, 5266–5277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carattoli, A.; Zankari, E.; Garcia-Fernandez, A.; Voldby Larsen, M.; Lund, O.; Villa, L.; Moller Aarestrup, F.; Hasman, H. In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain No. | Strain Background | Genotype | Phenotype * | Source |
|---|---|---|---|---|
| BSX 137 | USDA15WA-1, FSIS1503788, SX 238 | serovar I 4,[5],12:i:- | ASSuT | Glenn Tillman FSIS [6] |
| SX 240 | USDA15WA-1, FSIS1503788 | serovar I 4,[5],12:i:- | ASSuT | Swine passaged [17] |
| BSX 120 | UK-1, SB 26 | serovar Typhimurium | Nal | [18] |
| BBS 1268 | USDA15WA-1 | pKD46-Gm | ASSuTGm, 30 °C | BSX 137/pKD46-GM; plasmid source [16] |
| BBS 1270 | USDA15WA-1 | ΔSGI-4: neo | ASSuTKn | BBS 1268/oBBI 528/529-neo |
| BBS 1356 | USDA15WA-1 | SGI-4Ωneo | ASSuTKn | BBS 1268/oBBI 542/543-neo |
| BBS 1358 | UK-1 | SGI-4Ωneo | NalKn | BSX 120 x BBS 1356 |
| BBS 1359 | UK-1 | SGI-4Ωneo | NalKn | BSX 120 x BBS 1356 |
| BBS 1406 | UK-1 | SGI-4Ωneo | NalKn | BBS 1358small |
| BBS 1407 | UK-1 | SGI-4Ωneo | NalKn | BBS 1358large |
| Gene ID | Gene Name | 30 Min | 60 Min | Description | Location | ||
|---|---|---|---|---|---|---|---|
| Log2FC | FDR | Log2FC | FDR | ||||
| AOL22_00855 | cueO | 4.3 | 5.02 × 10−34 | 4.2 | 7.55 × 10−31 | multicopper oxidase CueO | Core |
| AOL22_02225 | mdsC | 3 | 7.80 × 10−9 | 1.7 | 1.85 × 10−9 | multidrug efflux transporter outer membrane subunit MdsC (GesC) | Core |
| AOL22_02230 | mdsB | 3.5 | 5.39 × 10−8 | 2.8 | 2.35× 10−11 | multidrug efflux RND transporter permease subunit MdsB (GesB) | Core |
| AOL22_02235 | mdsA | 5.3 | 6.59 × 10−12 | 4.8 | 6.80 × 10−16 | multidrug efflux RND transporter periplasmic adaptor subunit MdsA (GesA) | Core |
| AOL22_02240 | golT | 7 | 2.09 × 10−28 | 6.3 | 1.33 × 10−18 | gold/copper-translocating P-type ATPase GolT | Core |
| AOL22_02245 | golS | 5.9 | 3.38 × 10−29 | 5 | 2.55 × 10−27 | Au(I) sensor transcriptional regulator GolS | Core |
| AOL22_02250 | golB | 6.9 | 1.35 × 10−97 | 6.5 | 2.12 × 10−102 | gold resistance metallochaperone GolB | Core |
| AOL22_02995 | copA | 5.7 | 2.64 × 10−25 | 5.2 | 4.89 × 10−48 | copper-exporting P-type ATPase CopA | Core |
| AOL22_05910 | scsA | 2.4 | 9.70 × 10−6 | 4.1 | 1.13 × 10−22 | copper resistance protein | Core |
| AOL22_05915 | scsB | 6.2 | 7.68 × 10−37 | 7.8 | 2.48 × 10−88 | protein disulfide reductase | Core |
| AOL22_05920 | scsC | 5.4 | 4.80 × 10−29 | 6.7 | 1.42 × 10−75 | disulfide bond formation protein DsbA | Core |
| AOL22_05925 | scsD | 4.8 | 4.60 × 10−18 | 6.4 | 2.60 × 10−58 | protein disulfide oxidoreductase | Core |
| AOL22_14670 | merE | 1.8 | 2.10 × 10−3 | 1.3 | 6.18 × 10−3 | broad-spectrum mercury transporter MerE | MDR |
| AOL22_14675 | merD | 1.4 | 7.46 × 10−3 | 1.5 | 1.72 × 10−3 | mercury resistance co-regulator MerD | MDR |
| AOL22_14680 | merA | 1.3 | 1.40 × 10−2 | 1.1 | 1.09 × 10−2 | mercury(II) reductase | MDR |
| AOL22_14685 | merC | 1.4 | 4.45 × 10−3 | 1.2 | 4.41 × 10−3 | organomercurial transporter MerC | MDR |
| AOL22_14690 | merP | 1.4 | 3.21 × 10−3 | 1.1 | 5.62 × 10−3 | mercury resistance system periplasmic binding protein MerP | MDR |
| AOL22_14695 | merT | 1.6 | 1.49 × 10−3 | 1.5 | 1.00 × 10−3 | mercuric ion transporter MerT | MDR |
| AOL22_23325 | arsR | 2.6 | 1.35 × 10−3 | 2 | 1.65 × 10−3 | transcriptional regulator | SGI-4 |
| AOL22_23330 | arsD | 4.8 | 1.34 × 10−9 | 4 | 4.09 × 10−7 | arsenite efflux transporter metallochaperone ArsD | SGI-4 |
| AOL22_23335 | arsA | 4.6 | 4.25 × 10−9 | 3.7 | 1.39 × 10−6 | arsenite efflux transporter ATPase subunit ArsA | SGI-4 |
| AOL22_23340 | arsB | 4.5 | 9.60 × 10−8 | 3.9 | 1.56 × 10−6 | arsenic transporter | SGI-4 |
| AOL22_23345 | arsC | 3.7 | 2.37 × 10−10 | 3 | 1.23 × 10−6 | arsenate reductase (glutaredoxin) | SGI-4 |
| AOL22_23510 | silE | 9 | 2.29 × 10−53 | 7.5 | 1.55 × 10−32 | silver-binding protein SilE | SGI-4 |
| AOL22_23515 | silS | 2.7 | 4.93 × 10−10 | 2 | 1.42 × 10−6 | copper/silver sensor histidine kinase SilS | SGI-4 |
| AOL22_23520 | silR | 3.2 | 6.17 × 10−8 | 2.4 | 2.57 × 10−6 | copper/silver response regulator transcription factor SilR | SGI-4 |
| AOL22_23525 | silC | 7 | 1.04 × 10−19 | 6.5 | 6.81 × 10−19 | Cu(+)/Ag(+) efflux RND transporter outer membrane channel SilC | SGI-4 |
| AOL22_23530 | silF | 7.3 | 4.31 × 10−32 | 6.9 | 4.25 × 10−23 | copper ABC transporter substrate-binding protein | SGI-4 |
| AOL22_23535 | silB | 7.5 | 3.63 × 10−25 | 7 | 2.39 × 10−21 | Cu(+)/Ag(+) efflux RND transporter periplasmic adaptor subunit SilB | SGI-4 |
| AOL22_23540 | silA | 6.7 | 8.15 × 10−26 | 5.8 | 1.80 × 10−19 | Cu(+)/Ag(+) efflux RND transporter permease subunit SilA | SGI-4 |
| AOL22_23545 | orf105 | 6.9 | 8.38 × 10−204 | 4.8 | 1.30 × 10−64 | hypothetical protein | SGI-4 |
| AOL22_23550 | silP | 5.7 | 1.94 × 10−47 | 3.5 | 1.66 × 10−15 | Ag(+)-translocating P-type ATPase SilP | SGI-4 |
| AOL22_23565 | pcoG | 2.3 | 6.85 × 10−8 | 0.71 | 5.20 × 10−2 | copper resistance protein | SGI-4 |
| AOL22_23570 | pcoA | 2.4 | 9.92 × 10−19 | 2.3 | 1.10 × 10−25 | multicopper oxidase PcoA | SGI-4 |
| AOL22_23575 | pcoB | 2.4 | 5.26 × 10−16 | 2.3 | 2.71 × 10−17 | copper resistance outer membrane transporter PcoB | SGI-4 |
| AOL22_23580 | pcoC | 2.5 | 4.30 × 10−15 | 2.1 | 2.41 × 10−20 | copper resistance system metallochaperone PcoC | SGI-4 |
| AOL22_23585 | pcoD | 1.7 | 4.43 × 10−7 | 1.4 | 4.73 × 10−6 | copper resistance inner membrane protein PcoD | SGI-4 |
| AOL22_23590 | pcoR | 0.4 | 2.43 × 10−1 | 0.7 | 2.27 × 10−2 | copper response regulator transcription factor PcoR | SGI-4 |
| AOL22_23595 | pcoS | 0.3 | 2.57 × 10−1 | 0.8 | 1.73 × 10−4 | copper resistance membrane spanning protein PcoS | SGI-4 |
| AOL22_23600 | pcoE | 7.7 | 6.64 × 10−25 | 6.7 | 1.68 × 10−21 | copper-binding protein | SGI-4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bearson, B.L.; Trachsel, J.M.; Shippy, D.C.; Sivasankaran, S.K.; Kerr, B.J.; Loving, C.L.; Brunelle, B.W.; Curry, S.M.; Gabler, N.K.; Bearson, S.M.D. The Role of Salmonella Genomic Island 4 in Metal Tolerance of Salmonella enterica Serovar I 4,[5],12:i:- Pork Outbreak Isolate USDA15WA-1. Genes 2020, 11, 1291. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11111291
Bearson BL, Trachsel JM, Shippy DC, Sivasankaran SK, Kerr BJ, Loving CL, Brunelle BW, Curry SM, Gabler NK, Bearson SMD. The Role of Salmonella Genomic Island 4 in Metal Tolerance of Salmonella enterica Serovar I 4,[5],12:i:- Pork Outbreak Isolate USDA15WA-1. Genes. 2020; 11(11):1291. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11111291
Chicago/Turabian StyleBearson, Bradley L., Julian M. Trachsel, Daniel C. Shippy, Sathesh K. Sivasankaran, Brian J. Kerr, Crystal L. Loving, Brian W. Brunelle, Shelby M. Curry, Nicholas K. Gabler, and Shawn M. D. Bearson. 2020. "The Role of Salmonella Genomic Island 4 in Metal Tolerance of Salmonella enterica Serovar I 4,[5],12:i:- Pork Outbreak Isolate USDA15WA-1" Genes 11, no. 11: 1291. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11111291