Dominance of the ON1 Genotype of RSV-A and BA9 Genotype of RSV-B in Respiratory Cases from Jeddah, Saudi Arabia

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Inclusion and Exclusion Criteria
2.2. Sample Collection
2.3. Molecular Detection of Respiratory Viruses
2.4. Virus Isolation
2.5. Whole Genome Sequencing and Typing of RSV
3. Results
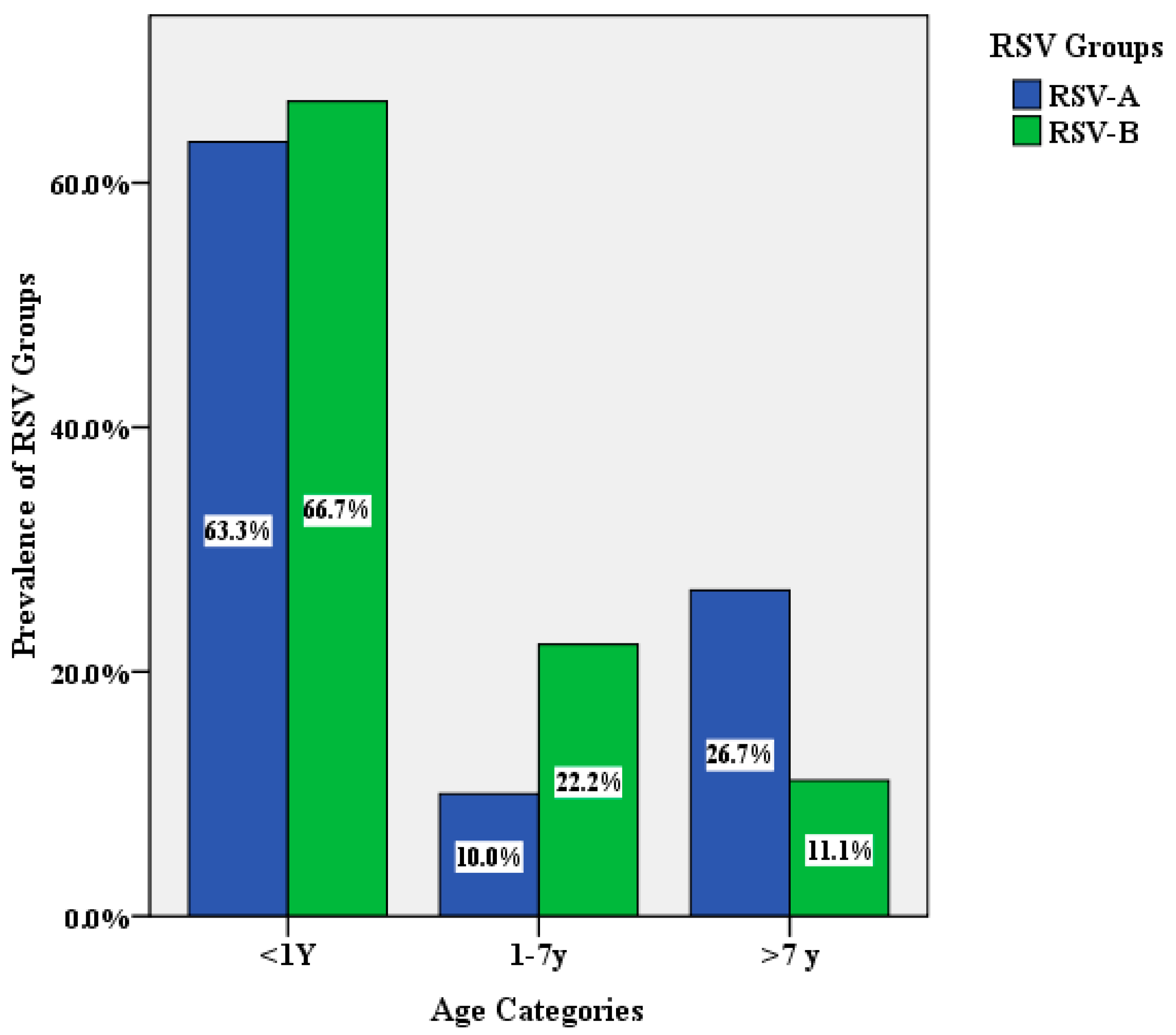
3.1. The Characterization and Prevalence of RSV in Respiratory Samples
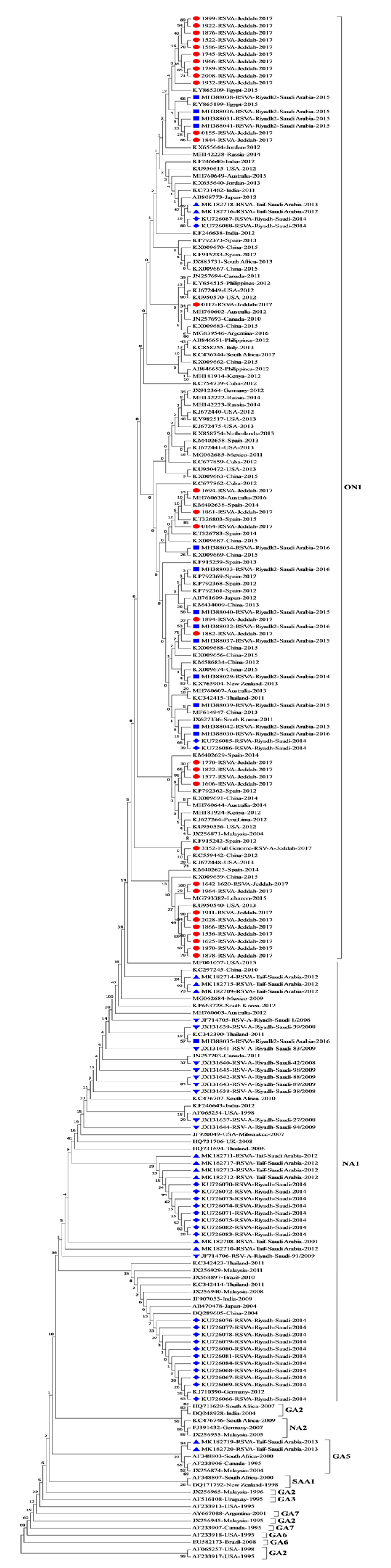
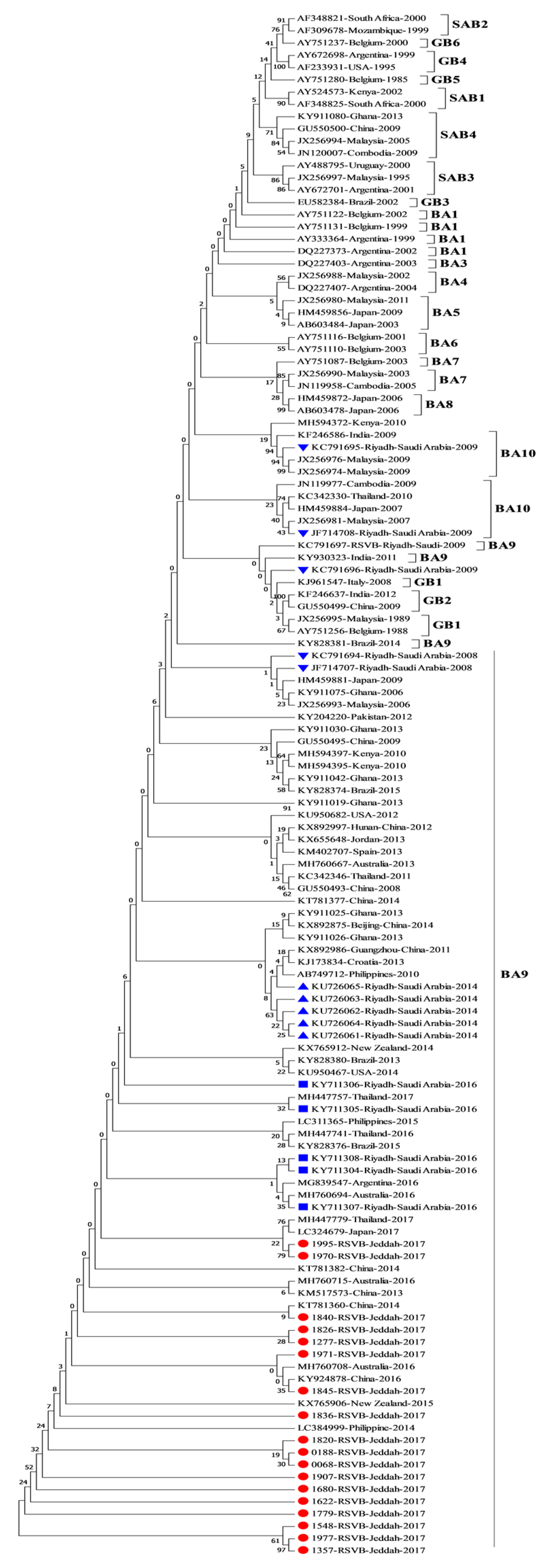
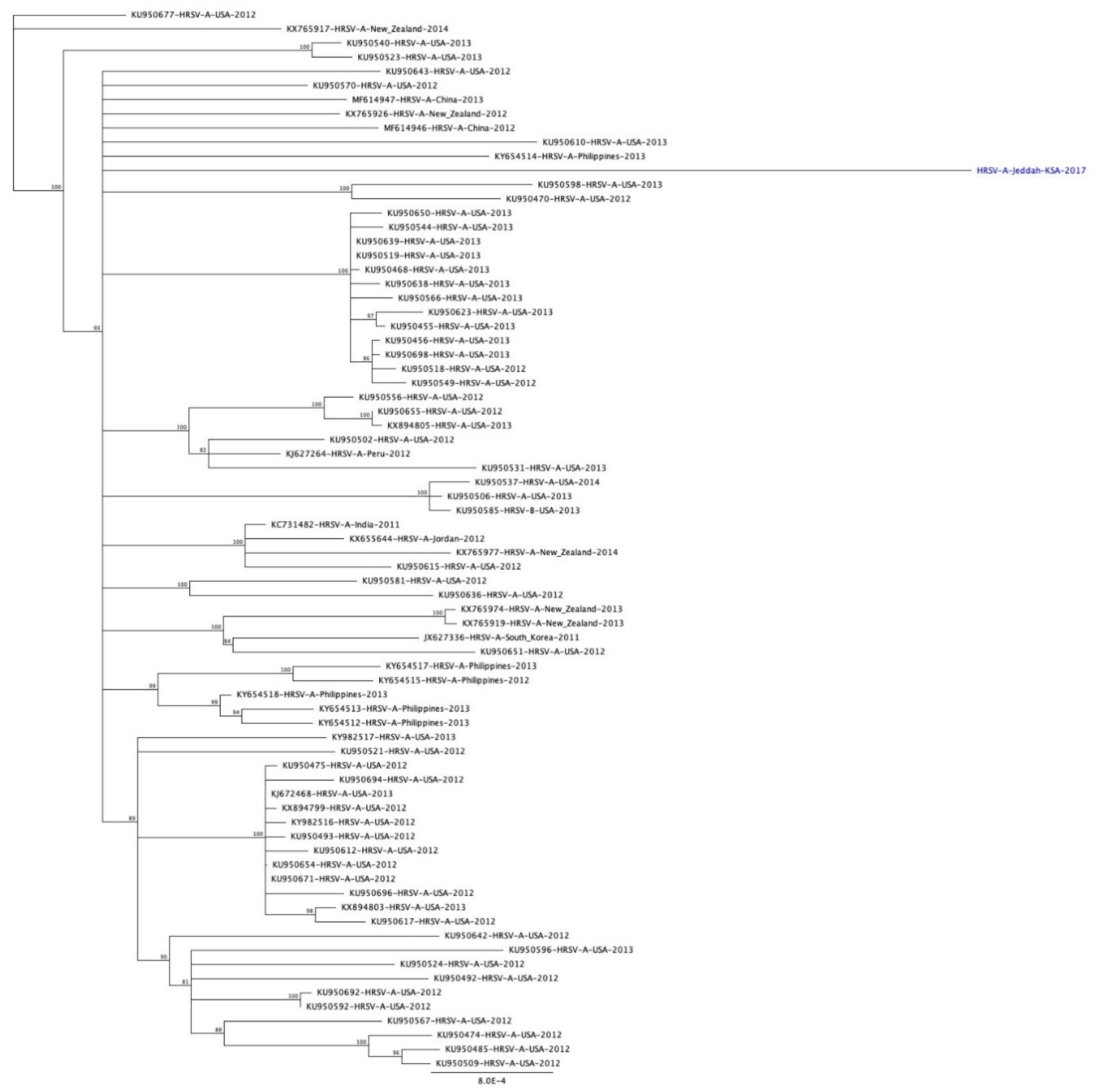
3.2. Phylogenetic Analysis of RSV
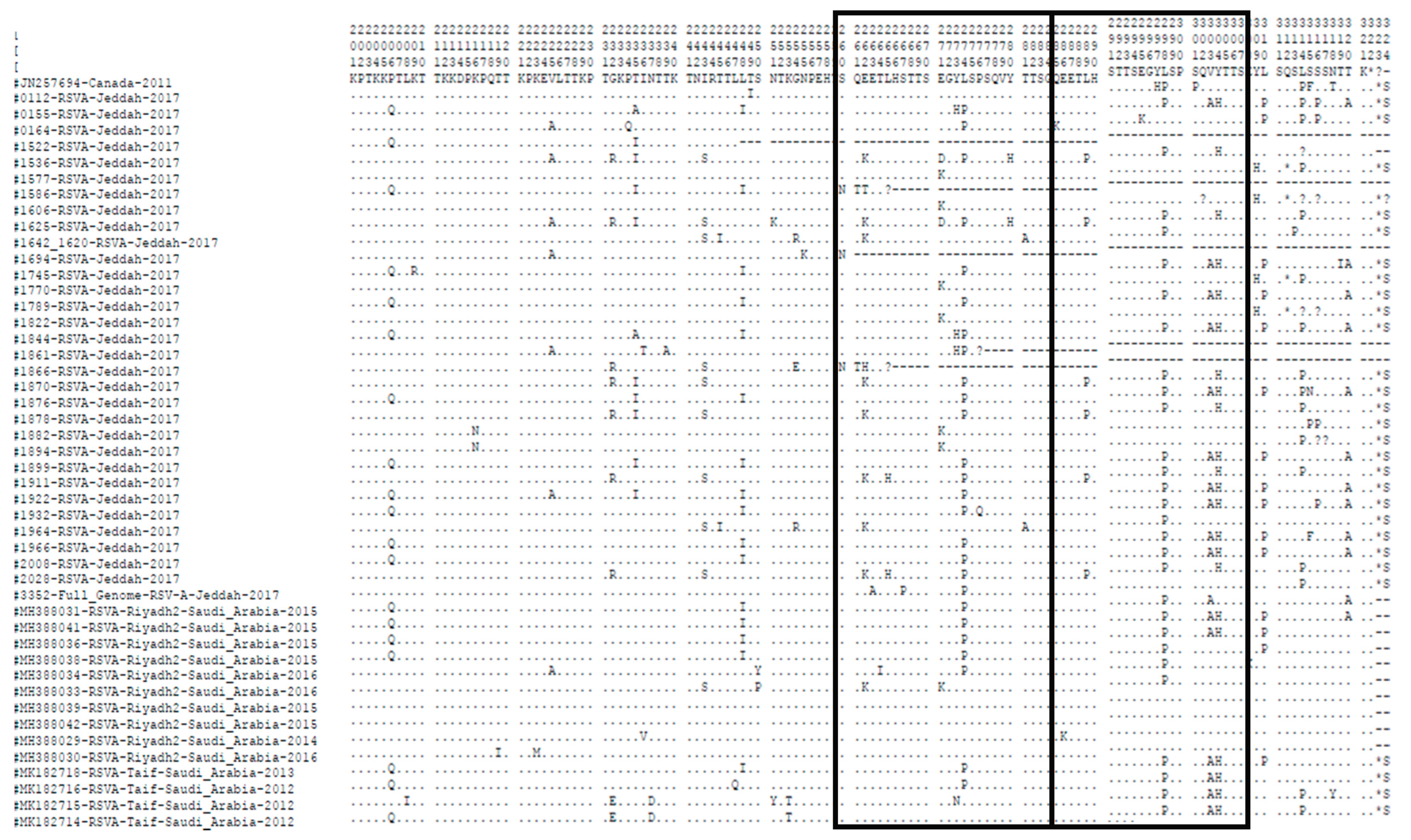
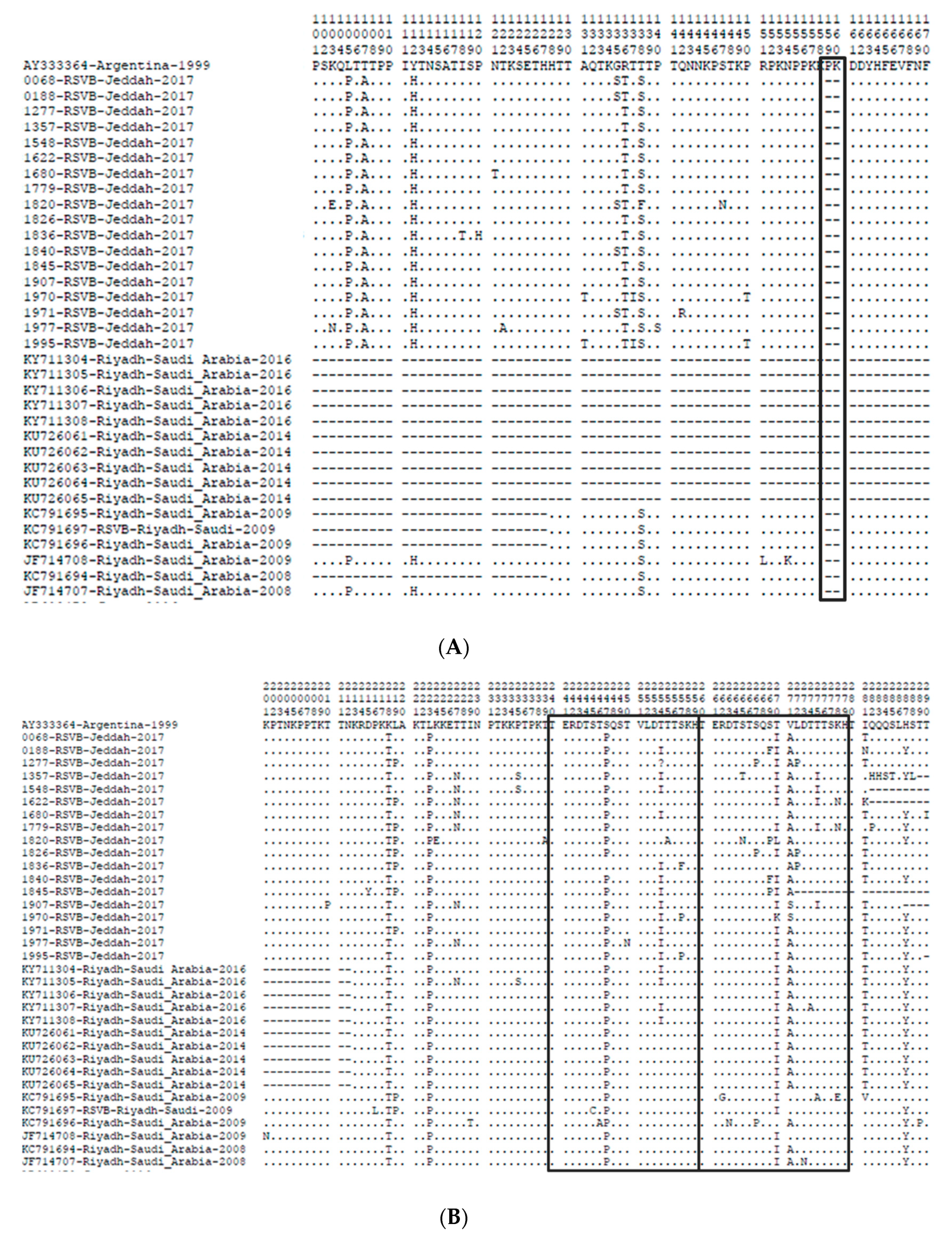
3.3. Amino Acid Sequence Analysis of the G Gene
3.4. N- and O-Glycosylation Site Analysis in Amino Acid Sequence
3.5. Full Genome Sequence
4. Discussions
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yu, X.; Kou, Y.; Xia, D.; Li, J.; Yang, X.; Zhou, Y.; He, X. Human respiratory syncytial virus in children with lower respiratory tract infections or influenza-like illness and its co-infection characteristics with viruses and atypical bacteria in Hangzhou, China. J. Clin. Virol. 2015, 69, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Ogunsemowo, O.; Olaleye, D.O.; Odaibo, G.N. Genetic diversity of human respiratory syncytial virus circulating among children in Ibadan, Nigeria. PLoS ONE 2018, 13, e0191494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nair, H.; Nokes, D.J.; Gessner, B.D.; Dherani, M.; Madhi, S.A.; Singleton, R.J.; O’Brien, K.L.; Roca, A.; Wright, P.F.; Bruce, N. Global burden of acute lower respiratory infections due to respiratory syncytial virus in young children: A systematic reviewand meta-analysis. Lancet 2010, 375, 1545–1555. [Google Scholar] [CrossRef] [Green Version]
- Fan, R.; Fan, C.; Zhang, J.; Wen, B.; Lei, Y.; Liu, C.; Chen, L.; Liu, W.; Wang, C.; Qu, X. Respiratory syncytial virus subtype ON1/NA1/BA9 predominates in hospitalized children with lower respiratory tract infections. J. Med. Virol. 2017, 89, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Eshaghi, A.; Duvvuri, V.R.; Lai, R.; Nadarajah, J.T.; Li, A.; Patel, S.N.; Low, D.E.; Gubbay, J.B. Genetic variability of human respiratory syncytial virus A strainscirculating in Ontario: A novel genotype with a 72 nucleotide G gene duplication. PLoS ONE 2012, 7, e32807. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, A.; Haider, S.H.; Parveen, S.; Arshad, M.; Alsenaidy, H.A.; Baaboud, A.O.; Mobaireek, K.F.; AlSaadi, M.M.; Alsenaidy, A.M.; Sullender, W. Co-Circulation of 72bp Duplication Group A and 60bp Duplication Group B Respiratory Syncytial Virus (RSV) Strains in Riyadh, Saudi Arabia during 2014. PLoS ONE 2016, 11, e0166145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, G.; Zhu, R.; Deng, J.; Zhao, L.; Sun, Y.; Wang, F.; Qian, Y. Rapid replacement of prevailing genotype of human respiratory syncytial virus by genotype ON1 in Beijing, 2012–2014. Infect. Genet. Evol. 2015, 33, 163–168. [Google Scholar] [CrossRef]
- Tabatabai, J.; Prifert, C.; Pfeil, J.; Grulich-Henn, J.; Schnitzler, P. Novel respiratory syncytial virus (RSV) genotype ON1 predominates in Germany during winter season 2012–13. PLoS ONE 2014, 9, e109191. [Google Scholar] [CrossRef] [Green Version]
- Pierangeli, A.; Trotta, D.; Scagnolari, C.; Ferreri, M.; Nicolai, A.; Midulla, F.; Marinelli, K.; Antonelli, G.; Bagnarelli, P. Rapid spread of the novel respiratory syncytial virus A ON1 genotype, central Italy, 2011 to 2013. Eurosurveillance 2014, 19, 20843. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.-J.; Kim, D.-W.; Lee, W.-J.; Yun, M.-R.; Lee, H.Y.; Lee, H.S.; Jung, H.-D.; Kim, K. Rapid replacement of human respiratory syncytial virus A with the ON1 genotype having 72 nucleotide duplication in G gene. Infect. Genet. Evol. 2014, 26, 103–112. [Google Scholar] [CrossRef]
- Korsun, N.; Angelova, S.; Trifonova, I.; Voleva, S.; Grigorova, I.; Tzotcheva, I.; Mileva, S.; Alexiev, I.; Perenovska, P. Predominance of ON1 and BA9 Genotypes of Respiratory Syncytial Virus (RSV) in Bulgaria, 2016–2018. J. Med. Virol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Jagusic, M.; Slovic, A.; Ivancic-Jelecki, J.; Ljubin-Sternak, S.; Vilibić-Čavlek, T.; Tabain, I.; Forcic, D. Molecular epidemiology of human respiratory syncytial virus and human metapneumovirus in hospitalized children with acute respiratory infections in Croatia, 2014–2017. Infect. Genet. Evol. 2019, 76, 104039. [Google Scholar] [CrossRef]
- Korsun, N.; Angelova, S.; Tzotcheva, I.; Georgieva, I.; Lazova, S.; Parina, S.; Alexiev, I.; Perenovska, P. Prevalence and genetic characterisation of respiratory syncytial viruses circulating in Bulgaria during the 2014/15 and 2015/16 winter seasons. Pathog. Glob Health 2017, 111, 351–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trento, A.; Casas, I.; Calderón, A.; Garcia-Garcia, M.L.; Calvo, C.; Perez-Breña, P.; Melero, J.A. Ten years of global evolution of the human respiratory syncytial virus BA genotype with a 60-nucleotide duplication in the Gprotein gene. J. Virol. 2010, 84, 7500–7512. [Google Scholar] [CrossRef] [Green Version]
- Pretorius, M.A.; van Niekerk, S.; Tempia, S.; Moyes, J.; Cohen, C.; Madhi, S.A.; Venter, M.; SARI Surveillance Group. Replacement and Positive Evolution of Subtype A and B Respiratory SyncytialVirus G-Protein Genotypes from 1997–2012 in South Africa. J. Infect. Dis. 2013, 208 (Suppl. 3), S227–S237. [Google Scholar] [CrossRef] [Green Version]
- Almajhdi, F.N.; Farrag, M.A.; Amer, H.M. Group B strains of human respiratory syncytial virus in Saudi Arabia: Molecularand phylogenetic analysis. Virus Genes 2014, 48, 252–259. [Google Scholar] [CrossRef]
- Xia, Q.; Zhou, L.; Peng, C.; Hao, R.; Ni, K.; Zang, N.; Ren, L.; Deng, Y.; Xie, X.; He, L. Detection of respiratory syncytial virus fusion protein variants between 2009 and 2012 in China. Arch. Virol. 2014, 159, 1089–1098. [Google Scholar] [CrossRef]
- Runan, Z.; Xuehui, G.; Zhiliang, W. Identification of subgroups of respiratory syncytial virus by reverse transcription polymerase chain reaction. Chin. J. Pediatr. 1998, 9, 433–437. [Google Scholar]
- Rebuffo-Scheer, C.; Bose, M.; He, J.; Khaja, S.; Ulatowski, M.; Beck, E.T.; Fan, J.; Kumar, S.; Nelson, M.I.; Henrickson, K.J. Whole genome sequencing and evolutionary analysis of human respiratory syncytial virus A and B from Milwaukee, WI 1998–2010. PLoS ONE 2011, 6, e25468. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Gupta, R.; Jung, E.; Brunak, S. Prediction of N-Glycosylation Sites in Human Proteins. 2004. Available online: http://www.cbs.dtu.dk/services/NetNGlyc (accessed on 26 October 2020).
- Julenius, K.; Mølgaard, A.; Gupta, R.; Brunak, S. Prediction, conservation analysis, and structural characterization of mammalian mucin-type O-glycosylation sites. Glycobiology 2005, 15, 153–164. [Google Scholar] [CrossRef] [Green Version]
- Albogami, S.S.; Alotaibi, M.R.; Alsahli, S.A.; Masuadi, E.; Alshaalan, M. Seasonal variations of respiratory viruses detected from children with respiratory tract infections in Riyadh, Saudi Arabia. J. Infect. Public Health 2018, 11, 183–186. [Google Scholar] [CrossRef]
- Etemadi, M.R.; Sekawi, Z.; Othman, N.; Lye, M.-S.; Moghaddam, F.Y. Circulation of human respiratory syncytial virus strains among hospitalized children with acute lower respiratory infection in malaysia. Evol. Bioinform. 2013, 9, S10999. [Google Scholar] [CrossRef] [PubMed]
- Comas-García, A.; Noyola, D.E.; Cadena-Mota, S.; Rico-Hernández, M.; Bernal-Silva, S. Respiratory syncytial virus-A ON1 genotype emergence in central Mexico in 2009 and evidence of multiple duplication events. J. Infect. Dis. 2018, 217, 1089–1098. [Google Scholar] [CrossRef] [Green Version]
- Mejias, A.; Ramilo, O. New options in the treatment of respiratory syncytial virus disease. J. Infect. 2015, 71, S80–S87. [Google Scholar] [CrossRef] [PubMed]
- Taleb, S.A.; Al Thani, A.A.; Al Ansari, K.; Yassine, H.M. Human respiratory syncytial virus: Pathogenesis, immune responses, and current vaccine approaches. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 1817–1827. [Google Scholar] [CrossRef]
- Gimferrer, L.; Vila, J.; Piñana, M.; Andrés, C.; Rodrigo-Pendás, J.A.; Peremiquel-Trillas, P.; Codina, M.G.; CMartín, M.D.; Esperalba, J.; Fuentes, F.; et al. Virological surveillance of human respiratory syncytial virus A and B at a tertiary hospital in Catalonia (Spain) during five consecutive seasons (2013–2018). Future Microbiol. 2019, 14, 373–381. [Google Scholar] [CrossRef]
- Thongpan, I.; Mauleekoonphairoj, J.; Vichiwattana, P.; Korkong, S.; Wasitthankasem, R.; Vongpunsawad, S.; Poovorawan, Y. Respiratory syncytial virus genotypes NA1, ON1, and BA9 are prevalent in Thailand, 2012–2015. PeerJ 2017, 5, e3970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valley-Omar, Z.; Muloiwa, R.; Hu, N.-C.; Eley, B.; Hsiao, N.-Y. Novel respiratory syncytial virus subtype ON1 among children, Cape Town, South Africa, 2012. Emerg. Infect. Dis. 2013, 19, 668. [Google Scholar] [CrossRef]
- Calderón, A.; Pozo, F.; Calvo, C.; García-García, M.; González-Esguevillas, M.; Molinero, M.; Casas, I. Genetic variability of respiratory syncytial virus A in hospitalized children in the last five consecutive winter seasons in Central Spain. J. Med. Virol. 2017, 89, 767–774. [Google Scholar] [CrossRef] [PubMed]
- Schobel, S.A.; Stucker, K.M.; Moore, M.L.; Anderson, L.J.; Larkin, E.K.; Shankar, J.; Bera, J.; Puri, V.; Shilts, M.H.; Rosas-Salazar, C. Respiratory Syncytial Virus whole-genome sequencing identifies convergent evolution of sequence duplication in the C-terminus of the G gene. Sci. Rep. 2016, 6, 26311. [Google Scholar] [CrossRef] [PubMed]
- Gaymard, A.; Bouscambert-Duchamp, M.; Pichon, M.; Frobert, E.; Vallee, J.; Lina, B.; Casalegno, J.-S.; Morfin, F. Genetic characterization of respiratory syncytial virus highlights a new BA genotype and emergence of the ON1 genotype in Lyon, France, between 2010 and 2014. J. Clin. Virol. 2018, 102, 12–18. [Google Scholar] [CrossRef]
- Panayiotou, C.; Richter, J.; Koliou, M.; Kalogirou, N.; Georgiou, E.; Christodoulou, C. Epidemiology of respiratory syncytial virusin children in Cyprus during three consecutive winter seasons (2010–2013): Age distribution, seasonality and association between prevalent genotypes and disease severity. Epidemiol. Infect. 2014, 142, 2406–2411. [Google Scholar] [CrossRef]
- Madi, N.; Chehadeh, W.; Asadzadeh, M.; Al-Turab, M.; Al-Adwani, A. Analysis of genetic variability of respiratory syncytial virus groups A and B in Kuwait. Arch. Virol. 2018, 163, 2405–2413. [Google Scholar] [CrossRef]
- Abdel-Moneim, A.S.; Soliman, M.S.; Kamel, M.M.; El-Kholy, A.A. Sequence analysis of the G gene of hRSVA ON1 genotype from Egyptian children with acute respiratory tract infections. J. Med. Microbiol. 2018, 67, 387–391. [Google Scholar] [CrossRef]
- Abou-El-Hassan, H.; Massaad, E.; Soudani, N.; Assaf-Casals, A.; Shaker, R.; Khoury, M.L.; Ghanem, S.; Karam, M.; Andary, R.; Saito, R. Detection of ON1 and novel genotypes of human respiratory syncytial virus and emergence of palivizumab resistance in Lebanon. PLoS ONE 2019, 14, e0212687. [Google Scholar] [CrossRef] [Green Version]
- Malekshahi, S.S.; Razaghipour, S.; Samieipoor, Y.; Hashemi, F.B.; Manesh, A.A.R.; Izadi, A.; Faghihloo, E.; Ghavami, N.; Mokhtari-Azad, T.; Salimi, V. Molecular characterization of the glycoprotein and fusion protein in human respiratory syncytial virus subgroup A: Emergence of ON-1 genotype in Iran. Infect. Genet. Evol. 2019, 71, 166–178. [Google Scholar] [CrossRef]
- Almajhdi, F.N.; Farrag, M.A.; Amer, H.M. Genetic diversity in the G protein gene of group A human respiratory syncytial viruses circulating in Riyadh, Saudi Arabia. Arch. Virol. 2014, 159, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Al Aboud, D.; Al Aboud, N.M.; Al-Malky, M.I.; Abdel-Moneim, A.S. Genotyping of Type A Human Respiratory Syncytial Virus Based on Direct F Gene Sequencing. Medicina 2019, 55, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auksornkitti, V.; Kamprasert, N.; Thongkomplew, S.; Suwannakarn, K.; Theamboonlers, A.; Samransamruajkij, R.; Poovorawan, Y. Molecular characterization of human respiratory syncytial virus, 2010-2011: Identification of genotype ON1 and a new subgroup B genotype in Thailand. Arch. Virol. 2014, 159, 499–507. [Google Scholar] [CrossRef]
- Goya, S.; Valinotto, L.E.; Tittarelli, E.; Rojo, G.L.; Jodar, M.S.N.; Greninger, A.L.; Zaiat, J.J.; Marti, M.A.; Mistchenko, A.S.; Viegas, M. An optimized methodology for whole genome sequencing of RNArespiratory viruses from nasopharyngeal aspirates. PLoS ONE 2018, 13, e0199714. [Google Scholar] [CrossRef] [Green Version]
- Zou, L.; Yi, L.; Wu, J.; Song, Y.; Huang, G.; Zhang, X.; Liang, L.; Ni, H.; Pybus, O.G.; Ke, C. Evolution and transmission of respiratory syncytial group a (RSV-A) viruses in Guangdong, China 2008–2015. Front. Microbiol. 2016, 7, 1263. [Google Scholar] [CrossRef]
- Prifert, C.; Streng, A.; Krempl, C.D.; Liese, J.; Weissbrich, B. Novel respiratory syncytial virus a genotype, Germany, 2011–2012. Emerg. Infect. Dis. 2013, 19, 1029. [Google Scholar] [CrossRef] [PubMed]
- Otieno, J.R.; Kamau, E.M.; Oketch, J.W.; Ngoi, J.M.; Gichuki, A.M.; Binter, Š.; Otieno, G.P.; Ngama, M.; Agoti, C.N.; Cane, P.A. Whole genome analysis of local Kenyan and global sequences unravels the epidemiological and molecularevolutionary dynamics of RSV genotype ON1 strains. Virus Evol. 2018, 4, vey027. [Google Scholar] [CrossRef] [Green Version]
- Khor, C.-S.; Sam, I.-C.; Hooi, P.-S.; Chan, Y.-F. Displacement of predominant respiratory syncytial virus genotypes in Malaysia between 1989 and 2011. Infect. Genet. Evol. 2013, 14, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, M.; Anand, S.; Wadhwa, B.; Chadha, M. Genetic variability of human respiratory syncytial virus in Pune, Western India. Infect. Genet. Evol. 2013, 20, 369–377. [Google Scholar] [CrossRef]
- Ezzeddine, A.; Soudani, N.; Lee, C.K.; Dbaibo, G.; Elbahesh, H.; Zaraket, H. Complete Genome Sequence of an ON1 Human Respiratory Syncytial Virus Strain Isolated in Lebanon in 2015. Genome Announc. 2018, 6, e00316-18. [Google Scholar] [CrossRef] [Green Version]
- Malasao, R.; Okamoto, M.; Chaimongkol, N.; Imamura, T.; Tohma, K.; Dapat, I.; Dapat, C.; Suzuki, A.; Saito, M.; Saito, M. Molecular characterization of human respiratory syncytial virus in the Philippines, 2012–2013. PLoS ONE 2015, 10, e0142192. [Google Scholar] [CrossRef] [Green Version]
- Cane, P. Molecular epidemiology and evolution of RSV. Perspect. Med. Virol. 2006, 14, 89–114. [Google Scholar]
- Trento, A.; Galiano, M.; Videla, C.; Carballal, G.; García-Barreno, B.; Melero, J.A.; Palomo, C. Major changes in the G protein of human respiratory syncytial virus isolates introducedby a duplication of 60 nucleotides. J. Gen. Virol. 2003, 84, 3115–3120. [Google Scholar] [CrossRef]
- Zlateva, K.T.; Lemey, P.; Moës, E.; Vandamme, A.-M.; Van Ranst, M. Genetic variability and molecular evolution of the human respiratory syncytial virus subgroup B attachment G protein. J. Virol. 2005, 79, 9157–9167. [Google Scholar] [CrossRef] [Green Version]
- Okamoto, M.; Dapat, C.P.; Sandagon, A.M.D.; Batangan-Nacion, L.P.; Lirio, I.C.; Tamaki, R.; Saito, M.; Saito-Obata, M.; Lupisan, S.P.; Oshitani, H. Molecular characterization of respiratory syncytial virus in children with repeated infections with subgroup B in the Philippines. J. Infect. Dis. 2018, 218, 1045–1053. [Google Scholar] [CrossRef] [PubMed]
- Di Giallonardo, F.; Kok, J.; Fernandez, M.; Carter, I.; Geoghegan, J.L.; Dwyer, D.E.; Holmes, E.C.; Eden, J.-S. Evolution of human respiratory syncytial virus (RSV) over multiple seasons in New South Wales, Australia. Viruses 2018, 10, 476. [Google Scholar] [CrossRef] [Green Version]
- Fedechkin, S.O.; George, N.L.; Wolff, J.T.; Kauvar, L.M.; DuBois, R.M. Structures of respiratory syncytial virus G antigen bound to broadly neutralizing antibodies. Sci. Immunol. 2018, 3, ar3534. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Frequency N (%) | ||
|---|---|---|
| Age | <1 year | 60 (71.4) |
| 1–7 years | 9 (10.7) | |
| >7 years | 15 (17.9) | |
| Gender | F | 36 (42.9) |
| M | 48 (57.1) | |
| Month | January 17 | 7 (8.3) |
| March 17 | 2 (2.4) | |
| September 17 | 2 (2.4) | |
| October 17 | 2 (2.4) | |
| November 17 | 28 (33.3) | |
| December 17 | 43 (51.2) | |
| Ward | ER | 57 (67.9) |
| ISO | 3 (3.6) | |
| MIC | 1 (1.2) | |
| PICU | 23 (27.4) |
| Sequence Name | Position Site | ON1 Nucleotide | Variant Nucleotide | Amino Acid Change | Codon Change | Protein Effect |
|---|---|---|---|---|---|---|
| JN257694-Canada-2011 | 910 | U | C | Y to H | UAU to CAU | Substitution |
| 653 | M | A | X to Q | CMA to CAA | Substitution | |
| 908 | U | C | V to A | GUC to GCC | Substitution | |
| 704 | C | T | T to I | ACC to ATC | Substitution | |
| 958 | A | G | T to A | ACA to GCA | Substitution | |
| 617 | C | A | P to Q | CCA to CAA | Substitution | |
| 821 | U | C | L to P | CUA to CCA | Substitution | |
| 893 | U | C | L to P | CUA to CCA | Substitution | |
| 929 | U | C | L to P | CUA to CCA | Substitution | |
| 941 | U | C | L to P | CUA to CCA | Substitution | |
| 742 | C | A | L to I | CUC to AUC | Substitution | |
| 401 | A | T | K to I | AAA to ATA | Substitution | |
| 728 | U | G | I to S | AUC to AGC | Substitution | |
| 784 | G | A | E to K | GAG to AAG | Substitution | |
| 69 | G | A | CUG to CUA | None | ||
| 369 | G | A | GAG to GAA | None | ||
| 717 | U | C | ACU to ACC | None | ||
| 840 | U | C | UAU to UAC | None | ||
| 945 | U | C | UCU to UCC | None |
| Sequence Name | Position Site | BA Nucleotide | Variant Nucleotide | Amino Acid Change | Codon Change | Protein Effect |
|---|---|---|---|---|---|---|
| AY333364-Argentina-1999 | 30 | G | A | AAG to AAA | None | |
| 177 | C | T | CUC to CUT | None | ||
| 276 | C | A | AUC to AUA | None | ||
| 283 | U | C | Y to H | UAC to CAC | Substitution | |
| 286 | C | T | L to F | CUU to TUU | Substitution | |
| 329 | U | C | L to P | CUC to CCC | Substitution | |
| 334 | A | G | T to A | ACC to GCC | Substitution | |
| 339 | A | G | ACA to ACG | None | ||
| 349 | U | C | Y to H | UAC to CAC | Substitution | |
| 375 | U | C | CCU to CCC | None | ||
| 422 | G | C | R to T | AGA to ACA | Substitution | |
| 427 | AC | TCT | T to S | ACC to TCT | Substitution | |
| 490 | CCAAAA | PK to no Amino Acid | CCA, AAA to no codon | Deletion | ||
| 525 | C | T | UUC to UUT | None | ||
| 591 | U | C | AAU to AAC | None | ||
| 612 | C | T | ACC to ACT | None | ||
| 614 | U | C | I to T | AUA to ACA | Substitution | |
| 636 | C | T | CCC to CCT | None | ||
| 663 | A | C | CCA to CCC | None | ||
| 668 | A | C | K to T | AAA to ACA | Substitution | |
| 671 | U | C | L to P | CUA to CCA | Substitution | |
| 683 | U | C | L to P | CUG to CCG | Substitution | |
| 695 | C | A | T to N | ACC to AAC | Substitution | |
| 702 | C | T | AUC to AUT | None | ||
| 732 | C | T | ACC to ACT | None | ||
| 754 | U | C | S to P | UCA to CCA | Substitution | |
| 776 | C | T | T to I | ACA to ATA | Substitution | |
| 824 | C | T | T to I | ACU to ATU | Substitution | |
| 827 | U | C | V to A | GUG to GCG | Substitution | |
| 831 | C | T | CUC to CUT | None | ||
| 857 | U | C | I to T | AUC to ACC | Substitution | |
| 870 | C | T | UCC to UCT | None | ||
| 874 | C | T | H to Y | CAC to TAC | Substitution | |
| 927 | C | T | UCC to UCT | None | ||
| 948 | C | T | UCC to UCT | None | ||
| 952 | C | T | Truncation | |||
| 955 | AAA | CTC | K to L | AAA to CTC | Substitution | |
| 956 | AA | CT | K to T | AAA to ACT | Substitution | |
| 959 | UC | AG | L to Q | CUC to CAG | Substitution | |
| 959 | UC | CA | L to P | CUC to CCA | Substitution | |
| 961 | UAG | GTC | Extension | |||
| 962 | AG | CA | Extension | |||
| 964 | U | A | ||||
| 965 | C | A | ||||
| 965 | C | T | ||||
| 966 | AU | TG | ||||
| 968 | A | C | ||||
| 968 | A | G | ||||
| 969 | U | C | ||||
| 970 | G | T | ||||
| 971 | C | A | ||||
| 971 | C | T | ||||
| 972 | U | A | ||||
| 972 | U | G | ||||
| 973 | U | G | ||||
| 974 | A | T | ||||
| 975 | G | A | ||||
| 975 | G | T | ||||
| 976 | U | A | ||||
| 978 | A | T | ||||
| 979 | UUC | AAA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
A. Al-Sharif, H.; El-Kafrawy, S.A.; Yousef, J.M.; Kumosani, T.A.; Kamal, M.A.; Khathlan, N.A.; Kaki, R.M.; Alnajjar, A.A.; Azhar, E.I. Dominance of the ON1 Genotype of RSV-A and BA9 Genotype of RSV-B in Respiratory Cases from Jeddah, Saudi Arabia. Genes 2020, 11, 1323. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11111323
A. Al-Sharif H, El-Kafrawy SA, Yousef JM, Kumosani TA, Kamal MA, Khathlan NA, Kaki RM, Alnajjar AA, Azhar EI. Dominance of the ON1 Genotype of RSV-A and BA9 Genotype of RSV-B in Respiratory Cases from Jeddah, Saudi Arabia. Genes. 2020; 11(11):1323. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11111323
Chicago/Turabian StyleA. Al-Sharif, Hessa, Sherif A. El-Kafrawy, Jehad M. Yousef, Taha A. Kumosani, Mohammad A. Kamal, Norah A. Khathlan, Reham M. Kaki, Abeer A. Alnajjar, and Esam I. Azhar. 2020. "Dominance of the ON1 Genotype of RSV-A and BA9 Genotype of RSV-B in Respiratory Cases from Jeddah, Saudi Arabia" Genes 11, no. 11: 1323. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11111323