Phased Haplotype Resolution of the SLC6A4 Promoter Using Long-Read Single Molecule Real-Time (SMRT) Sequencing

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples and Publicly Available Data
2.2. PCR Amplification of the SLC6A4 Promoter
2.3. SLC6A4 Promoter Sample Barcoding and Pooling
2.4. Single Molecule Real-Time (SMRT) Sequencing
2.5. Sanger Sequencing Confirmation
3. Results
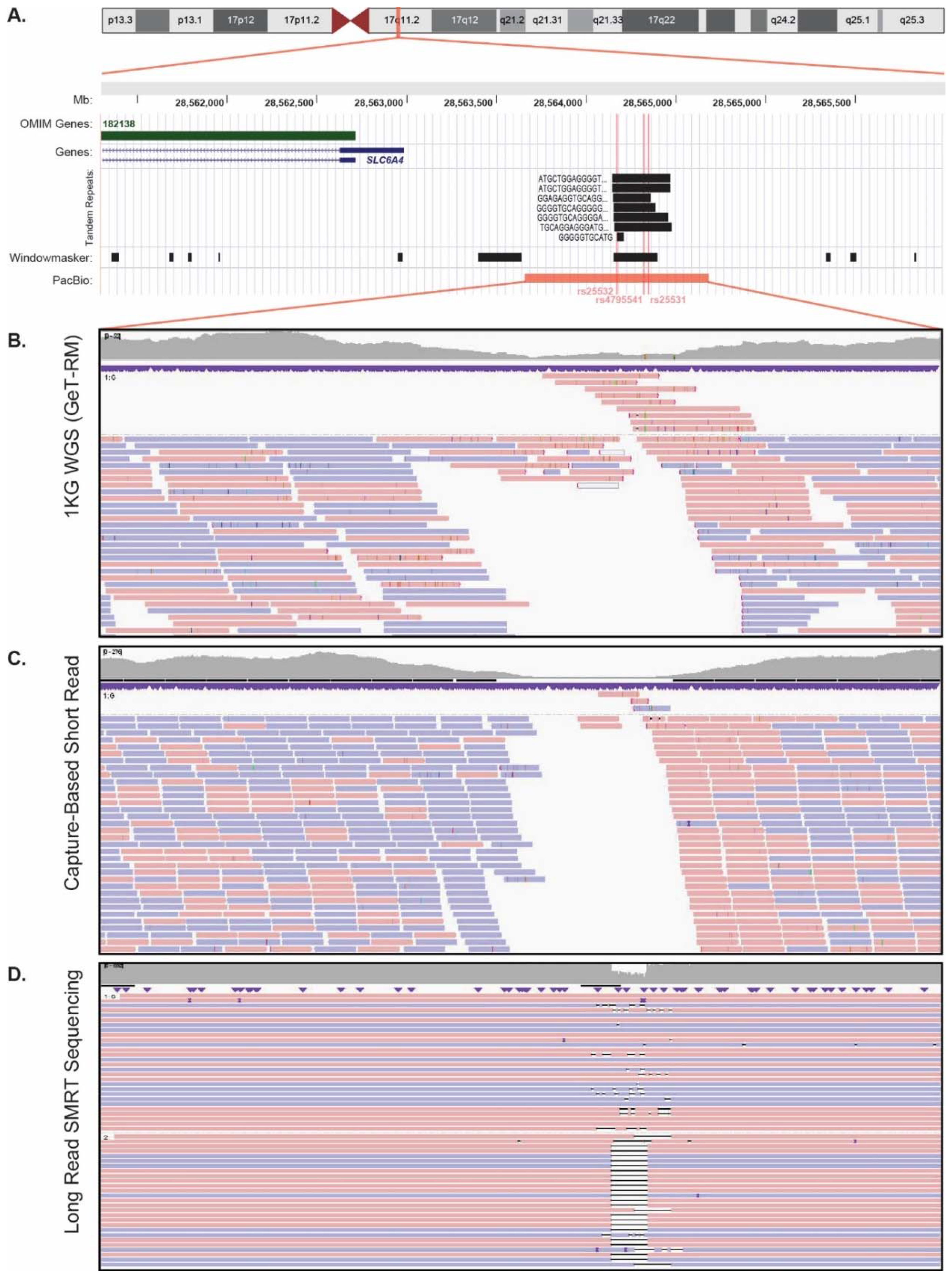
3.1. SLC6A4 Promoter Short-Read Sequencing
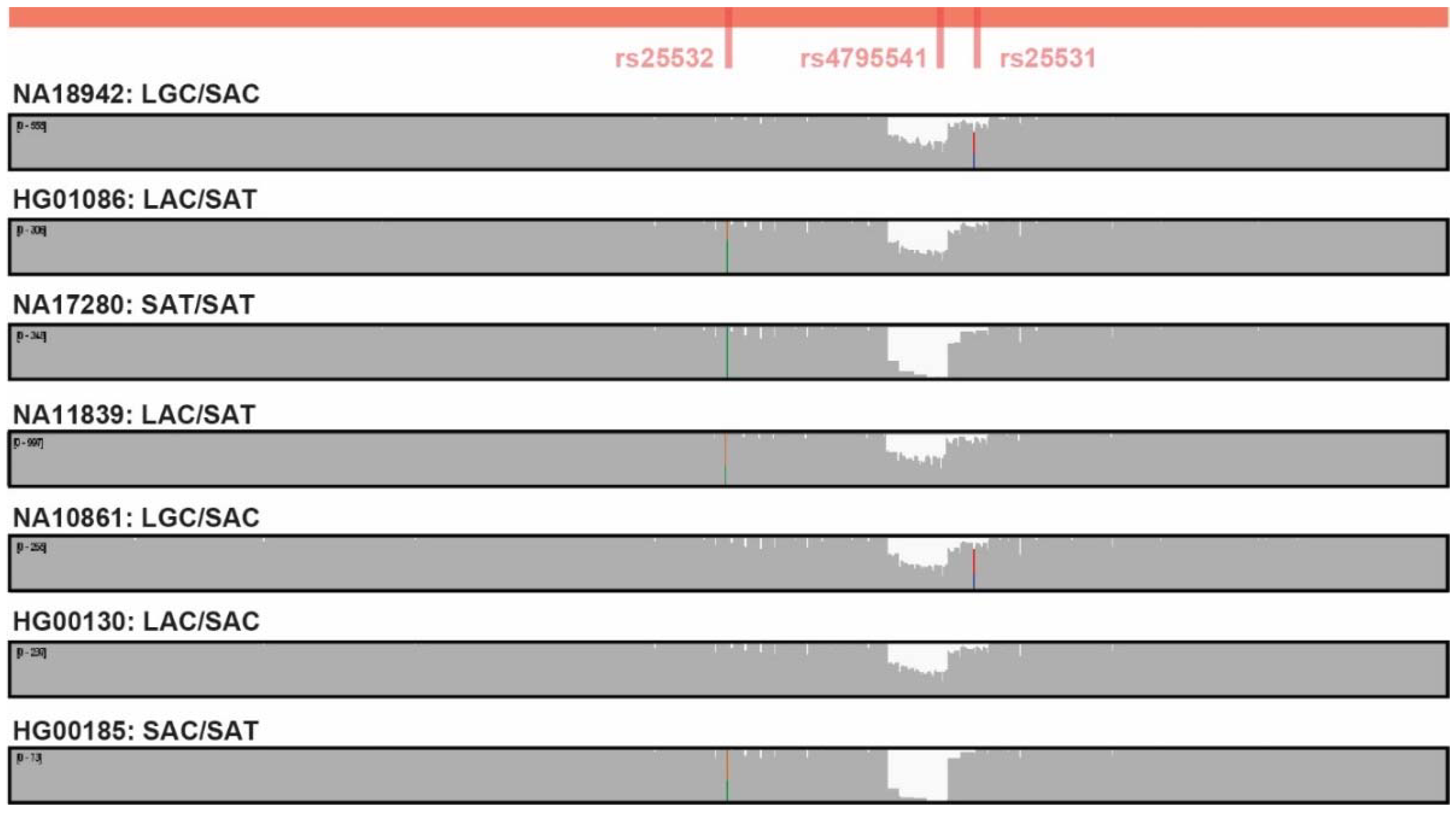
3.2. SLC6A4 Promoter Long-Read SMRT Sequencing
3.3. SLC6A4 Promoter Long-Read SMRT Sequencing Sanger Validation
3.4. SLC6A4 Promoter Long-Read SMRT Sequencing Precision
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- National Center for Health Statistics. Health, United States, 2016: With Chartbook on Long-Term Trends in Health; National Center for Health Statistics: Hyattsville, MD, USA, 2017.
- Sinyor, M.; Levitt, A.J.; Cheung, A.H.; Schaffer, A.; Kiss, A.; Dowlati, Y.; Lanctôt, K.L. Does inclusion of a placebo arm influence response to active antidepressant treatment in randomized controlled trials? Results from pooled and meta-analyses. J. Clin. Psychiatry 2010, 71, 270–279. [Google Scholar] [CrossRef] [PubMed]
- Pérez, V.; Salavert, A.; Espadaler, J.; Tuson, M.; Saiz-Ruiz, J.; Sáez-Navarro, C.; Bobes, J.; Baca-García, E.; Vieta, E.; Olivares, J.M.; et al. Efficacy of prospective pharmacogenetic testing in the treatment of major depressive disorder: Results of a randomized, double-blind clinical trial. BMC Psychiatry 2017, 17, 250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haddley, K.; Bubb, V.J.; Breen, G.; Parades-Esquivel, U.M.; Quinn, J.P. Behavioural genetics of the serotonin transporter. Curr. Top Behav. Neurosci. 2012, 12, 503–535. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.Z.; Rush, A.J.; Charney, D.; Wilson, A.F.; Sorant, A.J.; Papanicolaou, G.J.; Fava, M.; Trivedi, M.H.; Wisniewski, S.R.; Laje, G.; et al. Association between a functional serotonin transporter promoter polymorphism and citalopram treatment in adult outpatients with major depression. Arch. Gen. Psychiatry 2007, 64, 783–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masoliver, E.; Menoyo, A.; Pérez, V.; Volpini, V.; Rio, E.D.; Pérez, J.; Alvarez, E.; Baiget, M. Serotonin transporter linked promoter (polymorphism) in the serotonin transporter gene may be associated with antidepressant-induced mania in bipolar disorder. Psychiatr. Genet. 2006, 16, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Perlis, R.H.; Mischoulon, D.; Smoller, J.W.; Wan, Y.J.; Lamon-Fava, S.; Lin, K.M.; Rosenbaum, J.F.; Fava, M. Serotonin transporter polymorphisms and adverse effects with fluoxetine treatment. Biol. Psychiatry 2003, 54, 879–883. [Google Scholar] [CrossRef]
- Putzhammer, A.; Schoeler, A.; Rohrmeier, T.; Sand, P.; Hajak, G.; Eichhammer, P. Evidence of a role for the 5-HTTLPR genotype in the modulation of motor response to antidepressant treatment. Psychopharmacology 2005, 178, 303–308. [Google Scholar] [CrossRef]
- Heils, A.; Teufel, A.; Petri, S.; Seemann, M.; Bengel, D.; Balling, U.; Riederer, P.; Lesch, K.P. Functional promoter and polyadenylation site mapping of the human serotonin (5-HT) transporter gene. J. Neural Transm. Gen. Sect. 1995, 102, 247–254. [Google Scholar] [CrossRef]
- Heils, A.; Teufel, A.; Petri, S.; Stöber, G.; Riederer, P.; Bengel, D.; Lesch, K.P. Allelic variation of human serotonin transporter gene expression. J. Neurochem. 1996, 66, 2621–2624. [Google Scholar] [CrossRef]
- Lesch, K.P.; Bengel, D.; Heils, A.; Sabol, S.Z.; Greenberg, B.D.; Petri, S.; Benjamin, J.; Müller, C.R.; Hamer, D.H.; Murphy, D.L. Association of anxiety-related traits with a polymorphism in the serotonin transporter gene regulatory region. Science 1996, 274, 1527–1531. [Google Scholar] [CrossRef]
- Mortensen, O.V.; Thomassen, M.; Larsen, M.B.; Whittemore, S.R.; Wiborg, O. Functional analysis of a novel human serotonin transporter gene promoter in immortalized raphe cells. Brain Res. Mol. Brain Res. 1999, 68, 141–148. [Google Scholar] [CrossRef]
- Delbrück, S.J.; Wendel, B.; Grunewald, I.; Sander, T.; Morris-Rosendahl, D.; Crocq, M.A.; Berrettini, W.H.; Hoehe, M.R. A novel allelic variant of the human serotonin transporter gene regulatory polymorphism. Cytogenet. Cell Genet. 1997, 79, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Frisch, A.; Finkel, B.; Michaelovsky, E.; Sigal, M.; Laor, N.; Weizman, R. A rare short allele of the serotonin transporter promoter region (5-HTTLPR) found in an aggressive schizophrenic patient of Jewish Libyan origin. Psychiatr. Genet. 2000, 10, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Gelernter, J.; Kranzler, H.; Cubells, J.F. Serotonin transporter protein (SLC6A4) allele and haplotype frequencies and linkage disequilibria in African- and European-American and Japanese populations and in alcohol-dependent subjects. Hum. Genet. 1997, 101, 243–246. [Google Scholar] [CrossRef] [PubMed]
- Murdoch, J.D.; Speed, W.C.; Pakstis, A.J.; Heffelfinger, C.E.; Kidd, K.K. Worldwide population variation and haplotype analysis at the serotonin transporter gene SLC6A4 and implications for association studies. Biol. Psychiatry 2013, 74, 879–889. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M.; Ueno, S.; Sano, A.; Tanabe, H. The human serotonin transporter gene linked polymorphism (5-HTTLPR) shows ten novel allelic variants. Mol. Psychiatry 2000, 5, 32–38. [Google Scholar] [CrossRef] [Green Version]
- Vijayendran, M.; Cutrona, C.; Beach, S.R.; Brody, G.H.; Russell, D.; Philibert, R.A. The relationship of the serotonin transporter (SLC6A4) extra long variant to gene expression in an African American sample. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2012, 159B, 611–612. [Google Scholar] [CrossRef] [Green Version]
- Ehli, E.A.; Hu, Y.; Lengyel-Nelson, T.; Hudziak, J.J.; Davies, G.E. Identification and functional characterization of three novel alleles for the serotonin transporter-linked polymorphic region. Mol. Psychiatry 2012, 17, 185–192. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.Z.; Lipsky, R.H.; Zhu, G.; Akhtar, L.A.; Taubman, J.; Greenberg, B.D.; Xu, K.; Arnold, P.D.; Richter, M.A.; Kennedy, J.L.; et al. Serotonin transporter promoter gain-of-function genotypes are linked to obsessive-compulsive disorder. Am. J. Hum. Genet. 2006, 78, 815–826. [Google Scholar] [CrossRef] [Green Version]
- Wendland, J.R.; Moya, P.R.; Kruse, M.R.; Ren-Patterson, R.F.; Jensen, C.L.; Timpano, K.R.; Murphy, D.L. A novel, putative gain-of-function haplotype at SLC6A4 associates with obsessive-compulsive disorder. Hum. Mol. Genet. 2008, 17, 717–723. [Google Scholar] [CrossRef] [Green Version]
- Pratt, V.M.; Everts, R.E.; Aggarwal, P.; Beyer, B.N.; Broeckel, U.; Epstein-Baak, R.; Hujsak, P.; Kornreich, R.; Liao, J.; Lorier, R.; et al. Characterization of 137 Genomic DNA Reference Materials for 28 Pharmacogenetic Genes: A GeT-RM Collaborative Project. J. Mol. Diagn. 2016, 18, 109–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genomes Project, C.; Auton, A.; Brooks, L.D.; Durbin, R.M.; Garrison, E.P.; Kang, H.M.; Korbel, J.O.; Marchini, J.L.; McCarthy, S.; McVean, G.A.; et al. A global reference for human genetic variation. Nature 2015, 526, 68–74. [Google Scholar] [CrossRef] [Green Version]
- PyPGx. Available online: https://github.com/sbslee/pypgx (accessed on 11 November 2020).
- Tange, O. GNU Parallel 2018. 2018. Available online: http://0-doi-org.brum.beds.ac.uk/10.5281/zenodo.1146014 (accessed on 11 November 2020).
- Qiao, W.; Yang, Y.; Sebra, R.; Mendiratta, G.; Gaedigk, A.; Desnick, R.J.; Scott, S.A. Long-Read Single Molecule Real-Time Full Gene Sequencing of Cytochrome P450-2D6. Hum. Mutat. 2016, 37, 315–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borras, D.M.; Vossen, R.; Liem, M.; Buermans, H.P.J.; Dauwerse, H.; van Heusden, D.; Gansevoort, R.T.; den Dunnen, J.T.; Janssen, B.; Peters, D.J.M.; et al. Detecting PKD1 variants in polycystic kidney disease patients by single-molecule long-read sequencing. Hum. Mutat. 2017, 38, 870–879. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Li, S.; Wang, Q.; Pan, L.; Jiang, F.; Yang, X.; Zhang, N.; Han, M.; Jia, C. Association of 5-HTTLPR polymorphism with smoking behaviors: A meta-analysis. Physiol. Behav. 2015, 152, 32–40. [Google Scholar] [CrossRef]
- Kranzler, H.R.; Scott, D.; Tennen, H.; Feinn, R.; Williams, C.; Armeli, S.; Taylor, R.E.; Briggs-Gowan, M.J.; Covault, J. The 5-HTTLPR polymorphism moderates the effect of stressful life events on drinking behavior in college students of African descent. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2012, 159B, 484–490. [Google Scholar] [CrossRef] [Green Version]
- Arieff, Z.; Kaur, M.; Gameeldien, H.; van der Merwe, L.; Bajic, V.B. 5-HTTLPR polymorphism: Analysis in South African autistic individuals. Hum. Biol. 2010, 82, 291–300. [Google Scholar] [CrossRef] [Green Version]
- Karczewski, K.J.; Francioli, L.C.; Tiao, G.; Cummings, B.B.; Alfoldi, J.; Wang, Q.; Collins, R.L.; Laricchia, K.M.; Ganna, A.; Birnbaum, D.P.; et al. The mutational constraint spectrum quantified from variation in 141,456 humans. Nature 2020, 581, 434–443. [Google Scholar] [CrossRef]
- Iurescia, S.; Seripa, D.; Rinaldi, M. Role of the 5-HTTLPR and SNP Promoter Polymorphisms on Serotonin Transporter Gene Expression: A Closer Look at Genetic Architecture and In Vitro Functional Studies of Common and Uncommon Allelic Variants. Mol. Neurobiol. 2016, 53, 5510–5526. [Google Scholar] [CrossRef]
- Mandelker, D.; Schmidt, R.J.; Ankala, A.; McDonald Gibson, K.; Bowser, M.; Sharma, H.; Duffy, E.; Hegde, M.; Santani, A.; Lebo, M.; et al. Navigating highly homologous genes in a molecular diagnostic setting: A resource for clinical next-generation sequencing. Genet. Med. 2016, 18, 1282–1289. [Google Scholar] [CrossRef] [Green Version]
- Ardui, S.; Ameur, A.; Vermeesch, J.R.; Hestand, M.S. Single molecule real-time (SMRT) sequencing comes of age: Applications and utilities for medical diagnostics. Nucleic Acids Res. 2018, 46, 2159–2168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaedigk, A.; Sangkuhl, K.; Whirl-Carrillo, M.; Twist, G.P.; Klein, T.E.; Miller, N.A.; PharmVar Steering, C. The Evolution of PharmVar. Clin. Pharmacol. Ther. 2019, 105, 29–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Primer ID | Primer Sequence * | Amplicon Size (bp) |
|---|---|---|
| Forward primer | ||
| SLC6A4F | 5’-ATGGGTTCCAGAGTCAATCCTTTGCGTTTTCTGTTGCCCT-3’ | - |
| Reverse primers | ||
| SLC6A4R1 | 5’-GAAAGGTCTGGAGTCTTGATGAGGGACTGAGCTGGACAACCAC-3’ | 699 |
| SLC6A4R2 | 5’-GAAAGGTCTGGAGTCTTGATCCCAGCAGGAGCCTATTGTT-3’ | 1026 |
| SLC6A4R3 | 5’-GAAAGGTCTGGAGTCTTGATTCTCTTGACCTCGGACACCT-3’ | 1536 |
| SLC6A4R4 | 5’-GAAAGGTCTGGAGTCTTGATGAAAGAAACGTGGGTTCGAGG-3’ | 2096 |
| LAC | SAC | LGC | SAT | XLAC | SGC | |
|---|---|---|---|---|---|---|
| n (freq) * | 101 (0.421) | 89 (0.371) | 26 (0.108) | 21 (0.088) | 2 (0.008) | 1 (0.004) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Botton, M.R.; Yang, Y.; Scott, E.R.; Desnick, R.J.; Scott, S.A. Phased Haplotype Resolution of the SLC6A4 Promoter Using Long-Read Single Molecule Real-Time (SMRT) Sequencing. Genes 2020, 11, 1333. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11111333
Botton MR, Yang Y, Scott ER, Desnick RJ, Scott SA. Phased Haplotype Resolution of the SLC6A4 Promoter Using Long-Read Single Molecule Real-Time (SMRT) Sequencing. Genes. 2020; 11(11):1333. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11111333
Chicago/Turabian StyleBotton, Mariana R., Yao Yang, Erick R. Scott, Robert J. Desnick, and Stuart A. Scott. 2020. "Phased Haplotype Resolution of the SLC6A4 Promoter Using Long-Read Single Molecule Real-Time (SMRT) Sequencing" Genes 11, no. 11: 1333. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11111333