Evolutionary Understanding of Metacaspase Genes in Cultivated and Wild Oryza Species and Its Role in Disease Resistance Mechanism in Rice

 ,
,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification and Nomenclature of Rice Metacaspase Genes Sequences
2.2. Gene Structure, Motif Analysis, and Phylogenetic Tree Construction
2.3. Chromosomal Localization, Gene Synteny, and Cis-Regulatory Element Analysis
2.4. Localization and Tertiary (3D) Protein Structure Analyses
2.5. Haplotypic Evaluation
2.6. Transcriptomic Evaluation and Co-Expression Analysis of Metacaspase Genes
2.7. Plant Materials and Fungal Infection
2.8. RNA Extraction and Expression Analysis
3. Results
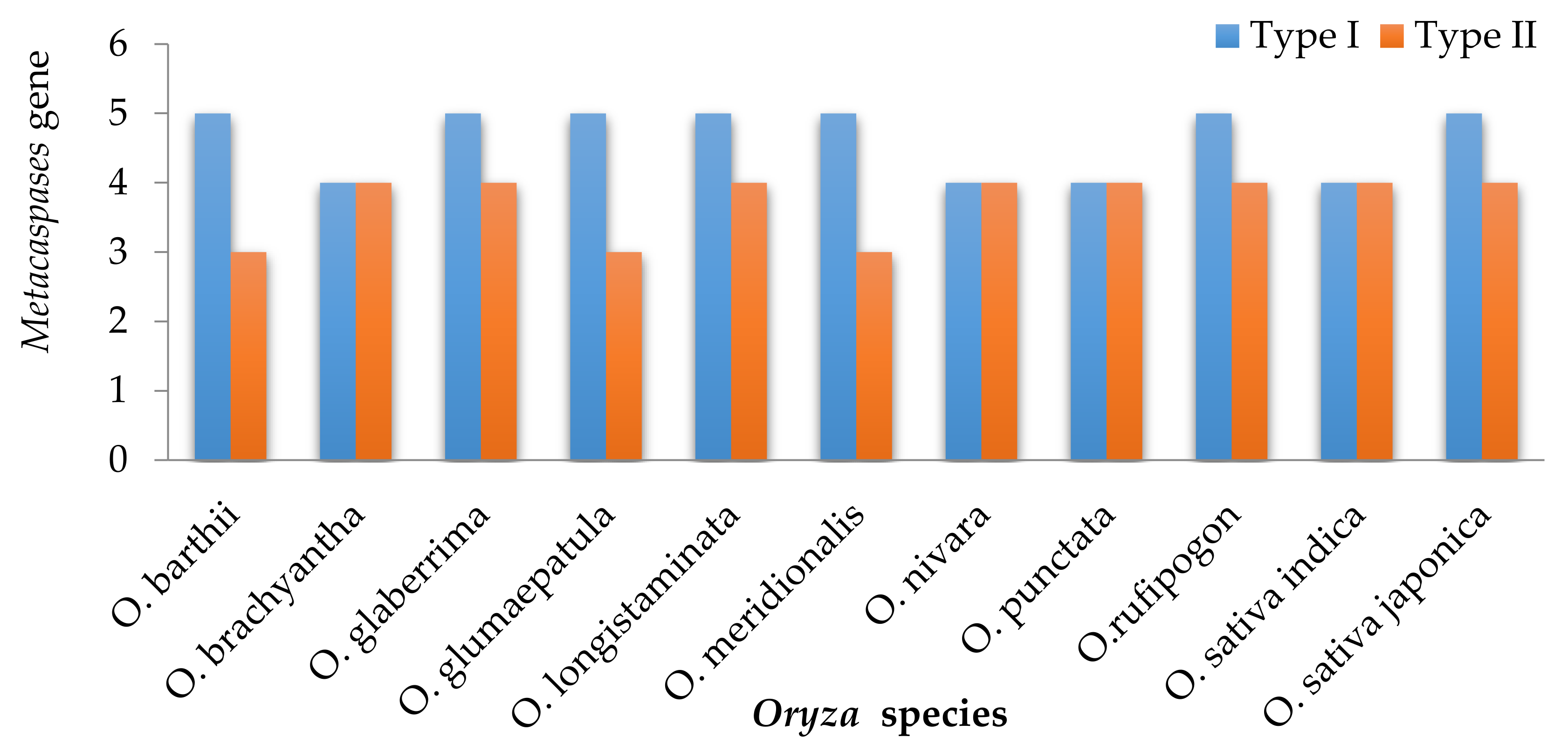
3.1. Metacaspases in Wild and Cultivated Rice Genomes
3.2. Predicted Structural and Functional Attributes of Metacaspase Genes
3.3. Gene Architecture, Conserved Motifs, and Phylogenetic Distribution of Metacaspase Genes
3.4. Chromosomal Distribution, Gene Duplication, and Syntenic Relationship of Metacaspase Genes in Wild and Cultivated Oryza Species
3.5. Cis-Acting Promoter Analysis
3.6. Haplotyping for Genes in Rice
3.7. Expression Analysis of Metacaspase Genes during Different Developmental and Stress Conditions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pennell, R.I.; Lamb, C. Programmed cell death in plants. Plant Cell 1997, 9, 1157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danon, A.; Delorme, V.; Mailhac, N.; Gallois, P. Plant programmed cell death: A common way to die. Plant Physiol. Biochem. 2000, 38, 647–655. [Google Scholar] [CrossRef]
- Thornberry, N.A.; Lazebnik, Y. Caspases: Enemies within. Science 1998, 281, 1312–1316. [Google Scholar] [CrossRef] [PubMed]
- Sanmartín, M.; Jaroszewski, L.; Raikhel, N.V.; Rojo, E. Caspases. Regulating death since the origin of life. Plant Physiol. 2005, 137, 841–847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonneau, L.; Ge, Y.; Drury, G.E.; Gallois, P. What happened to plant caspases? J. Exp. Bot. 2008, 59, 491–499. [Google Scholar] [CrossRef] [Green Version]
- Uren, A.G.; O’Rourke, K.; Aravind, L.A.; Pisabarro, M.T.; Seshagiri, S.; Koonin, E.V.; Dixit, V.M. Identification of paracaspases and metacaspases: Two ancient families of caspase-like proteins, one of which plays a key role in MALT lymphoma. Mol. Cell 2000, 6, 961–967. [Google Scholar] [CrossRef]
- Minina, E.; Coll, N.; Tuominen, H.; Bozhkov, P. Metacaspases versus caspases in development and cell fate regulation. Cell Death Differ. 2017, 24, 1314. [Google Scholar] [CrossRef]
- Vercammen, D.; Declercq, W.; Vandenabeele, P.; Van Breusegem, F. Are metacaspases caspases? J. Cell Biol. 2007, 179, 375–380. [Google Scholar] [CrossRef]
- Bozhkov, P.V.; Smertenko, A.P.; Zhivotovsky, B. Aspasing out metacaspases and caspases: Proteases of many trades. Sci. Signal. 2010, 3, pe48. [Google Scholar] [CrossRef]
- Carmona-Gutierrez, D.; Fröhlich, K.; Kroemer, G.; Madeo, F. Metacaspases Are Caspases. Doubt No More; Nature Publishing Group: Berlin, Germany, 2010. [Google Scholar]
- Enoksson, M.; Salvesen, G. Metacaspases Are Not Caspases—Always Doubt. Cell Death Differ. 2010, 8, 1221. [Google Scholar] [CrossRef]
- Tsiatsiani, L.; Van Breusegem, F.; Gallois, P.; Zavialov, A.; Lam, E.; Bozhkov, P. Metacaspases. Cell Death Differ. 2011, 18, 1279. [Google Scholar] [CrossRef] [PubMed]
- Fagundes, D.; Bohn, B.; Cabreira, C.; Leipelt, F.; Dias, N.; Bodanese-Zanettini, M.H.; Cagliari, A. Caspases in plants: Metacaspase gene family in plant stress responses. Funct. Integr. Genom. 2015, 15, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Zhang, H.; Hong, Y.; Liu, S.; Li, D.; Song, F. Stress-responsive expression, subcellular localization and protein–protein interactions of the rice metacaspase family. Int. J. Mol. Sci. 2015, 16, 16216–16241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, S.I.; Hwang, D.J. Expression analysis of the metacaspase gene family in Arabidopsis. J. Plant Biol. 2013, 56, 391–398. [Google Scholar] [CrossRef]
- Watanabe, N.; Lam, E. Arabidopsis metacaspase 2d is a positive mediator of cell death induced during biotic and abiotic stresses. Plant J. 2011, 66, 969–982. [Google Scholar] [CrossRef]
- Kim, S.M.; Bae, C.; Oh, S.K.; Choi, D. A pepper (Capsicum annuum L.) metacaspase 9 (Camc9) plays a role in pathogen-induced cell death in plants. Mol. Plant Pathol. 2013, 14, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Bostancioglu, S.M.; Tombuloglu, G.; Tombuloglu, H. Genome-wide identification of barley MCs (metacaspases) and their possible roles in boron-induced programmed cell death. Mol. Biol. Rep. 2018, 45, 211–225. [Google Scholar] [CrossRef]
- Wang, X.; Wang, X.; Feng, H.; Tang, C.; Bai, P.; Wei, G.; Huang, L.; Kang, Z. TaMCA4, a novel wheat metacaspase gene functions in programmed cell death induced by the fungal pathogen Puccinia striiformis f. sp. tritici. Mol. Plant Microbe Interact. 2012, 25, 755–764. [Google Scholar] [CrossRef] [Green Version]
- Hoeberichts, F.A.; Ten Have, A.; Woltering, E.J. A tomato metacaspase gene is upregulated during programmed cell death in Botrytis cinerea-infected leaves. Planta 2003, 217, 517–522. [Google Scholar] [CrossRef]
- Atwell, B.J.; Wang, H.; Scafaro, A.P. Could abiotic stress tolerance in wild relatives of rice be used to improve Oryza sativa? Plant Sci. 2014, 215, 48–58. [Google Scholar] [CrossRef]
- Garg, R.; Verma, M.; Agrawal, S.; Shankar, R.; Majee, M.; Jain, M. Deep transcriptome sequencing of wild halophyte rice, Porteresia coarctata, provides novel insights into the salinity and submergence tolerance factors. DNA Res. 2014, 21, 69–84. [Google Scholar] [CrossRef] [PubMed]
- Giuliani, R.; Koteyeva, N.; Voznesenskaya, E.; Evans, M.A.; Cousins, A.B.; Edwards, G.E. Coordination of leaf photosynthesis, transpiration, and structural traits in rice and wild relatives (genus Oryza). Plant Physiol. 2013, 162, 1632–1651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, W.-Y.; Wang, G.-L.; Chen, L.-L.; Kim, H.-S.; Pi, L.-Y.; Holsten, T.; Gardner, J.; Wang, B.; Zhai, W.-X.; Zhu, L.-H. A receptor kinase-like protein encoded by the rice disease resistance gene, Xa21. Science 1995, 270, 1804–1806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, T.-T. The origin, evolution, cultivation, dissemination, and diversification of Asian and African rices. Euphytica 1976, 25, 425–441. [Google Scholar] [CrossRef]
- Jena, K.K. The species of the genus Oryza and transfer of useful genes from wild species into cultivated rice, O. sativa. Breed. Sci. 2010, 60, 518–523. [Google Scholar] [CrossRef] [Green Version]
- Ensembl Plants. Oryza sativa Japonica Group (IRGSP-1.0). Available online: https://plants.ensembl.org/Oryza_sativa/Info/Index (accessed on 1 January 2020).
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Vercammen, D.; Van De Cotte, B.; De Jaeger, G.; Eeckhout, D.; Casteels, P.; Vandepoele, K.; Vandenberghe, I.; Van Beeumen, J.; Inzé, D.; Van Breusegem, F. Type II metacaspases Atmc4 and Atmc9 of Arabidopsis thaliana cleave substrates after arginine and lysine. J. Biol. Chem. 2004, 279, 45329–45336. [Google Scholar] [CrossRef] [Green Version]
- Marchler-Bauer, A.; Bo, Y.; Han, L.; He, J.; Lanczycki, C.J.; Lu, S.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R. CDD/SPARCLE: Functional classification of proteins via subfamily domain architectures. Nucleic Acids Res. 2017, 45, D200–D203. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.; Guo, A.-Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [Green Version]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Voorrips, R. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higo, K.; Ugawa, Y.; Iwamoto, M.; Korenaga, T. Plant cis-acting regulatory DNA elements (PLACE) database: 1999. Nucleic Acids Res. 1999, 27, 297–300. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.S.; Chen, Y.C.; Lu, C.H.; Hwang, J.K. Prediction of protein subcellular localization. Proteins Struct. Funct. Bioinform. 2006, 64, 643–651. [Google Scholar] [CrossRef]
- Krogh, A.; Larsson, B.; Von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [Green Version]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein identification and analysis tools on the ExPASy server. In The Proteomics Protocols Handbook; Springer: Berlin/Heidelberg, Germany, 2005; pp. 571–607. [Google Scholar]
- Yang, J.; Yan, R.; Roy, A.; Xu, D.; Poisson, J.; Zhang, Y. The I-TASSER Suite: Protein structure and function prediction. Nat. Methods 2015, 12, 7. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Yao, W.; Ouyang, Y.; Yang, W.; Wang, G.; Lian, X.; Xing, Y.; Chen, L.; Xie, W. RiceVarMap: A comprehensive database of rice genomic variations. Nucleic Acids Res. 2015, 43, D1018–D1022. [Google Scholar] [CrossRef]
- Valent, B.; Farrall, L.; Chumley, F.G. Magnaporthe grisea genes for pathogenicity and virulence identified through a series of backcrosses. Genetics 1991, 127, 87–101. [Google Scholar]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Abe, H.; Urao, T.; Ito, T.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Arabidopsis AtMYC2 (bHLH) and AtMYB2 (MYB) function as transcriptional activators in abscisic acid signaling. Plant Cell 2003, 15, 63–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eulgem, T.; Rushton, P.J.; Robatzek, S.; Somssich, I.E. The WRKY superfamily of plant transcription factors. Trends Plant Sci. 2000, 5, 199–206. [Google Scholar] [CrossRef]
- Baumann, K.; De Paolis, A.; Costantino, P.; Gualberti, G. The DNA binding site of the Dof protein NtBBF1 is essential for tissue-specific and auxin-regulated expression of the rolB oncogene in plants. Plant Cell 1999, 11, 323–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubey, N.; Trivedi, M.; Varsani, S.; Vyas, V.; Farsodia, M.; Singh, S.K. Genome-wide characterization, molecular evolution and expression profiling of the metacaspases in potato (Solanum tuberosum L.). Heliyon 2019, 5, e01162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, S.; Liu, A.; Zhang, Z.; Zou, X.; Jiang, X.; Huang, J.; Fan, L.; Zhang, Z.; Deng, X.; Ge, Q. Genome-Wide Identification and Expression Analysis of the Metacaspase Gene Family in Gossypium Species. Genes 2019, 10, 527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coll, N.S.; Vercammen, D.; Smidler, A.; Clover, C.; Van Breusegem, F.; Dangl, J.L.; Epple, P. Arabidopsis type I metacaspases control cell death. Science 2010, 330, 1393–1397. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, R.K.; Sonah, H.; Singh, N.K. Intron gain, a dominant evolutionary process supporting high levels of gene expression in rice. J. Plant Biochem. Biotechnol. 2016, 25, 142–146. [Google Scholar] [CrossRef]
- Zhang, C.; Gong, P.; Wei, R.; Li, S.; Zhang, X.; Yu, Y.; Wang, Y. The metacaspase gene family of Vitis vinifera L.: Characterization and differential expression during ovule abortion in stenospermocarpic seedless grapes. Gene 2013, 528, 267–276. [Google Scholar] [CrossRef]
- Deshmukh, R.K.; Vivancos, J.; Ramakrishnan, G.; Guérin, V.; Carpentier, G.; Sonah, H.; Labbé, C.; Isenring, P.; Belzile, F.J.; Bélanger, R.R. A precise spacing between the NPA domains of aquaporins is essential for silicon permeability in plants. Plant J. 2015, 83, 489–500. [Google Scholar] [CrossRef]
- Cao, Y.; Meng, D.; Chen, T.; Chen, Y.; Zeng, W.; Zhang, L.; Wang, Q.; Hen, W.; Abdullah, M.; Jin, Q. Metacaspase gene family in Rosaceae genomes: Comparative genomic analysis and their expression during pear pollen tube and fruit development. PLoS ONE 2019, 14, e0211635. [Google Scholar] [CrossRef] [Green Version]
- Zhu, P.; Yu, X.-H.; Wang, C.; Zhang, Q.; Liu, W.; McSweeney, S.; Shanklin, J.; Lam, E.; Liu, Q. Structural basis for Ca2+-dependent activation of a plant metacaspase. Nat. Commun. 2020, 11, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Deng, Z.; Chen, J.; Wang, S.; Hao, L.; Li, D. Genome-wide identification and expression analysis of the metacaspase gene family in Hevea brasiliensis. Plant Physiol. Biochem. 2016, 105, 90–101. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sr. No. | Selected Genes | MC Type | Gene | Length (aa) | MW (Kd) | pI | Protein Localization |
|---|---|---|---|---|---|---|---|
| 1 | OBART01G33990.1 | II | OBaMC7 | 417 | 45.8 | 5.52 | Nuclear |
| 2 | OBART03G19910.1 | I | OBaMC1 | 368 | 39.6 | 7.85 | Nuclear and Extracellular |
| 3 | OBART03G19930.1 | I | OBaMC2 | 372 | 40.0 | 5.73 | Chloroplast |
| 4 | OBART03G19940.1 | I | OBaMC5 | 280 | 30.9 | 6.12 | Nuclear |
| 5 | OBART03G19960.1 | I | OBaMC3 | 255 | 27.5 | 4.95 | Extracellular and Plasma membrane |
| 6 | OBART05G21540.1 | II | OBaMC6 | 409 | 44.5 | 6.53 | Nuclear |
| 7 | OBART05G21550.1 | II | OBaMC8 | 420 | 46.0 | 6.04 | Nuclear |
| 8 | OBART10G18850.1 | I | OBaMC4 | 334 | 36.1 | 6.16 | Extracellular |
| 9 | OB01G42780.1 | II | OBrMC5 | 417 | 45.8 | 5.46 | Nuclear |
| 10 | OB03G29120.1 | I | OBrMC1 | 247 | 27.0 | 7.6 | Nuclear and Mitochondrial |
| 11 | OB03G29130.1 | I | OBrMC2 | 344 | 36.7 | 5.62 | Chloroplast and Nuclear |
| 12 | OB03G29140.1 | I | OBrMC3 | 294 | 32.0 | 5.44 | Extracellular and Plasma membrane |
| 13 | OB05G29040.1 | II | OBrMC7 | 440 | 47.9 | 8.86 | Nuclear |
| 14 | OB05G29050.1 | II | OBrMC6 | 419 | 45.8 | 6.48 | Nuclear |
| 15 | OB10G25980.1 | I | OBrMC4 | 262 | 28.4 | 5.15 | Extracellular |
| 16 | OB12G12140.1 | II | OBrMC8 | 409 | 42.2 | 4.79 | Cytoplasmic |
| 17 | ORGLA01G0279200.1 | II | OGbMC7 | 417 | 45.8 | 5.52 | Nuclear |
| 18 | ORGLA03G0186200.1 | I | OGbMC1 | 368 | 39.6 | 7.85 | Extracellular, Nuclear |
| 19 | ORGLA03G0186300.1 | I | OGbMC2 | 397 | 42.7 | 11.24 | Nuclear |
| 20 | ORGLA03G0186400.1 | I | OGbMC5 | 302 | 33.5 | 6.46 | Nuclear |
| 21 | ORGLA03G0186500.1 | I | OGbMC3 | 369 | 39.1 | 7.91 | Chloroplast and Plasma Membrane |
| 22 | ORGLA05G0187100.1 | II | OGbMC6 | 409 | 44.5 | 6.53 | Nuclear |
| 23 | ORGLA05G0187200.1 | II | OGbMC8 | 420 | 46.0 | 6.04 | Nuclear |
| 24 | ORGLA10G0139900.1 | I | OGbMC4 | 355 | 38.5 | 5.8 | Extracellular |
| 25 | ORGLA11G0018800.1 | II | OGbMC9 | 341 | 35.6 | 5.09 | Cytoplasmic |
| 26 | OGLUM01G38250.1 | II | OGlMC7 | 417 | 45.8 | 5.37 | Nuclear |
| 27 | OGLUM03G20520.1 | I | OGlMC1 | 368 | 39.7 | 7.85 | Nuclear |
| 28 | OGLUM03G20560.1 | I | OGlMC2 | 392 | 41.4 | 7.91 | Chloroplast |
| 29 | OGLUM03G20570.1 | I | OGlMC5 | 176 | 19.7 | 4.98 | Cytoplasmic |
| 30 | OGLUM03G20600.1 | I | OGlMC3 | 443 | 48.0 | 8.67 | Chloroplast |
| 31 | OGLUM05G22740.1 | II | OGlMC6 | 409 | 44.5 | 6.35 | Nuclear |
| 32 | OGLUM05G22750.1 | II | OGlMC8 | 416 | 45.5 | 6.03 | Nuclear |
| 33 | OGLUM10G18910.1 | I | OGlMC4 | 582 | 63.1 | 7.91 | Extracellular |
| 34 | AMDW01032852.1_FGP001 | I | OLoMC1 | 120 | 12.6 | 4.35 | Cytoplasmic, Nuclear |
| 35 | KN538687.1_FGP194 | II | OLoMC8 | 823 | 87.3 | 5.32 | Chloroplast |
| 36 | KN538687.1_FGP207 | II | OLoMC6 | 420 | 46.0 | 6.04 | Nuclear |
| 37 | KN538892.1_FGP005 | I | OLoMC4 | 235 | 26.0 | 5.46 | Nuclear |
| 38 | KN538892.1_FGP007 | I | OLoMC2 | 272 | 29.8 | 5.16 | Nuclear |
| 39 | KN539149.1_FGP003 | I | OLoMC3 | 496 | 54.4 | 6.81 | Extracellular and Nuclear |
| 40 | KN539612.1_FGP001 | II | OLoMC9 | 452 | 49.2 | 6.63 | Plasma Membrane |
| 41 | KN540137.1_FGP001 | I | OLoMC5 | 157 | 16.7 | 4.77 | Plasma Membrane, Chloroplast |
| 42 | KN540619.1_FGP006 | II | OLoMC7 | 417 | 45.7 | 5.44 | Nuclear |
| 43 | OMERI03G17880.1 | I | OMeMC1 | 370 | 39.7 | 8.16 | Nuclear |
| 44 | OMERI03G17890.1 | I | OMeMC2 | 391 | 41.3 | 7.91 | Chloroplast |
| 45 | OMERI03G17900.1 | I | OMeMC5 | 303 | 33.4 | 5.83 | Nuclear |
| 46 | OMERI03G17920.1 | I | OMeMC3 | 395 | 42.0 | 8.87 | Plasma Membrane and Chloroplast |
| 47 | OMERI05G19430.1 | II | OMeMC6 | 420 | 45.9 | 5.94 | Nuclear |
| 48 | OMERI08G00110.2 | I | OMeMC4 | 450 | 49.0 | 8.59 | Extracellular and Nuclear |
| 49 | OMERI09G06400.1 | II | OMeMC7 | 518 | 56.8 | 6.81 | Nuclear |
| 50 | OMERI11G02500.2 | II | OMeMC8 | 833 | 90.9 | 8.27 | Plasma Membrane |
| 51 | ONIVA01G38790.1 | II | ONiMC6 | 417 | 45.8 | 5.44 | Nuclear |
| 52 | ONIVA03G21600.1 | I | ONiMC1 | 368 | 39.6 | 7.85 | Nuclear and Extracellular |
| 53 | ONIVA03G21620.1 | I | ONiMC2 | 679 | 73.0 | 6.77 | Chloroplast and Nuclear |
| 54 | ONIVA03G21660.1 | I | ONiMC3 | 360 | 38.3 | 7.5 | Plasma Membrane |
| 55 | ONIVA05G22210.1 | II | ONiMC5 | 409 | 44.5 | 6.57 | Nuclear |
| 56 | ONIVA05G22220.1 | II | ONiMC7 | 420 | 46.0 | 6.04 | Nuclear |
| 57 | ONIVA10G21340.1 | I | ONiMC4 | 814 | 87.8 | 9.05 | Extracellular and Nuclear |
| 58 | ONIVA11G02360.1 | II | ONiMC8 | 1086 | 117.6 | 6.95 | Plasma Membrane |
| 59 | OPUNC01G33030.1 | II | OPuMC5 | 417 | 45.9 | 5.37 | Nuclear |
| 60 | OPUNC03G18760.1 | I | OPuMC1 | 660 | 71.9 | 6.78 | Plasma Membrane and Nuclear |
| 61 | OPUNC03G18780.1 | I | OPuMC2 | 555 | 60.6 | 6.1 | Cytoplasmic and Nuclear |
| 62 | OPUNC03G18790.1 | I | OPuMC3 | 294 | 32.0 | 5.61 | Plasma membrane and Chloroplast |
| 63 | OPUNC05G19180.1 | II | OPuMC6 | 416 | 44.3 | 5.46 | Nuclear |
| 64 | OPUNC05G19190.1 | II | OPuMC7 | 419 | 46.0 | 6.14 | Nuclear |
| 65 | OPUNC10G17260.1 | I | OPuMC4 | 968 | 105.6 | 9.13 | Extracellular, Plasma membrane, and Nuclear |
| 66 | OPUNC12G01940.1 | II | OPuMC8 | 1019 | 109.9 | 6.54 | Plasma Membrane |
| 67 | ORUFI01G37300.1 | II | ORuMC7 | 417 | 45.8 | 5.54 | Nuclear |
| 68 | ORUFI03G20550.1 | I | ORuMC1 | 368 | 39.6 | 7.85 | Extracellular |
| 69 | ORUFI03G20570.1 | I | ORuMC2 | 400 | 42.1 | 7.01 | Chloroplast |
| 70 | ORUFI03G20590.1 | I | ORuMC5 | 268 | 29.5 | 5.75 | Nuclear |
| 71 | ORUFI03G20620.1 | I | ORuMC3 | 323 | 33.7 | 6.24 | Plasma Membrane |
| 72 | ORUFI05G22800.1 | II | ORuMC6 | 409 | 44.5 | 6.35 | Nuclear |
| 73 | ORUFI05G22810.1 | II | ORuMC8 | 420 | 46.0 | 6.04 | Nuclear |
| 74 | ORUFI10G20040.1 | I | ORuMC4 | 363 | 38.8 | 6.21 | Extracellular |
| 75 | ORUFI11G02080.1 | II | ORuMC9 | 341 | 35.7 | 5.32 | Cytoplasmic |
| 76 | BGIOSGA004618-PA | II | OInMC6 | 417 | 45.7 | 5.36 | Nuclear |
| 77 | BGIOSGA010607-PA | I | OInMC1 | 369 | 39.0 | 7.5 | Plasma Membrane |
| 78 | BGIOSGA010608-PA | I | OInMC4 | 301 | 33.6 | 6.46 | Nuclear |
| 79 | BGIOSGA010610-PA | I | OInMC2 | 400 | 42.1 | 7.91 | Chloroplast |
| 80 | BGIOSGA010611-PA | I | OInMC3 | 368 | 39.6 | 7.85 | Nuclear & Extracellular |
| 81 | BGIOSGA017791-PA | II | OInMC7 | 420 | 46.0 | 6.04 | Nuclear |
| 82 | BGIOSGA017792-PA | II | OInMC5 | 409 | 44.5 | 6.35 | Nuclear |
| 83 | BGIOSGA034558-PA | II | OInMC8 | 341 | 35.6 | 5.24 | Cytoplasmic |
| 84 | Os01t0799900-01 | II | OJaMC7 | 417 | 45.8 | 5.51 | Nuclear |
| 85 | Os03t0388900-01 | I | OJaMC1 | 368 | 39.6 | 7.85 | Extracellular and Nuclear |
| 86 | Os03t0389000-00 | I | OJaMC4 | 424 | 45.2 | 11.59 | Nuclear |
| 87 | Os03t0389100-01 | I | OJaMC5 | 302 | 33.5 | 6.46 | Nuclear |
| 88 | Os03t0389501-00 | I | OJaMC2 | 208 | 21.9 | 6.94 | Chloroplast |
| 89 | Os05t0496400-00 | II | OJaMC6 | 305 | 33.2 | 8.46 | Nuclear |
| 90 | Os05t0496500-01 | II | OJaMC8 | 420 | 46.0 | 6.04 | Nuclear |
| 91 | Os10t0565100-00 | I | OJaMC3 | 457 | 49.8 | 8.47 | Nuclear |
| 92 | Os11t0134700-01 | II | OJaMC9 | 341 | 35.6 | 5.24 | Cytoplasmic |
| Sr. No. | Gene ID | Gene Nomenclature | Chr. | Chr. Position | Haplotypes | SNPs/InDel | Non-Syn. SNP | Deleterious Mut. | Gene Diversity | |
|---|---|---|---|---|---|---|---|---|---|---|
| Pi/BP | Theta/BP | |||||||||
| 1 | Os03t388900 | OJaMC1 | 3 | 15530630–15532454 | 8 | 15 | 7 | 1 | 0.15 | 0.10 |
| 2 | Os03t389501 | OJaMC2 | 3 | 15586221–15587152 | 8 | 11 | 5 | 0 | 0.31 | 0.11 |
| 3 | Os03t0389000 | OJaMC4 | 3 | 15563374–15564927 | 6 | 16 | 2 | 0 | 0.14 | 0.11 |
| 4 | Os05t0496400 | OJaMC6 | 5 | 24383180–24384097 | 3 | 8 | 4 | 2 | 0.05 | 0.12 |
| 5 | Os05t0496500 | OJaMC8 | 5 | 24384902–24387337 | 4 | 8 | 3 | 1 | 0.01 | 0.12 |
| 6 | Os01t799900 | OJaMC7 | 1 | 33869142–33871835 | 10 | 31 | 3 | 0 | 0.06 | 0.11 |
| 7 | Os11t0134700 | OJaMC9 | 11 | 1616990–1618300 | 9 | 22 | 14 | 0 | 0.14 | 0.11 |
| 8 | Os03t0389100 | OJaMC5 | 3 | 15567899–15572135 | 128 | 372 | 49 | 5 | 0.16 | 0.11 |
| 9 | Os10t0565100 | OJaMC3 | 10 | 22320808–22323911 | 5 | 25 | 0 | NA | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bansal, R.; Rana, N.; Singh, A.; Dhiman, P.; Mandlik, R.; Sonah, H.; Deshmukh, R.; Sharma, T.R. Evolutionary Understanding of Metacaspase Genes in Cultivated and Wild Oryza Species and Its Role in Disease Resistance Mechanism in Rice. Genes 2020, 11, 1412. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11121412
Bansal R, Rana N, Singh A, Dhiman P, Mandlik R, Sonah H, Deshmukh R, Sharma TR. Evolutionary Understanding of Metacaspase Genes in Cultivated and Wild Oryza Species and Its Role in Disease Resistance Mechanism in Rice. Genes. 2020; 11(12):1412. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11121412
Chicago/Turabian StyleBansal, Ruchi, Nitika Rana, Akshay Singh, Pallavi Dhiman, Rushil Mandlik, Humira Sonah, Rupesh Deshmukh, and Tilak Raj Sharma. 2020. "Evolutionary Understanding of Metacaspase Genes in Cultivated and Wild Oryza Species and Its Role in Disease Resistance Mechanism in Rice" Genes 11, no. 12: 1412. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11121412