The POU Transcription Factor POU-M2 Regulates Vitellogenin Receptor Gene Expression in the Silkworm, Bombyx mori


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Strains
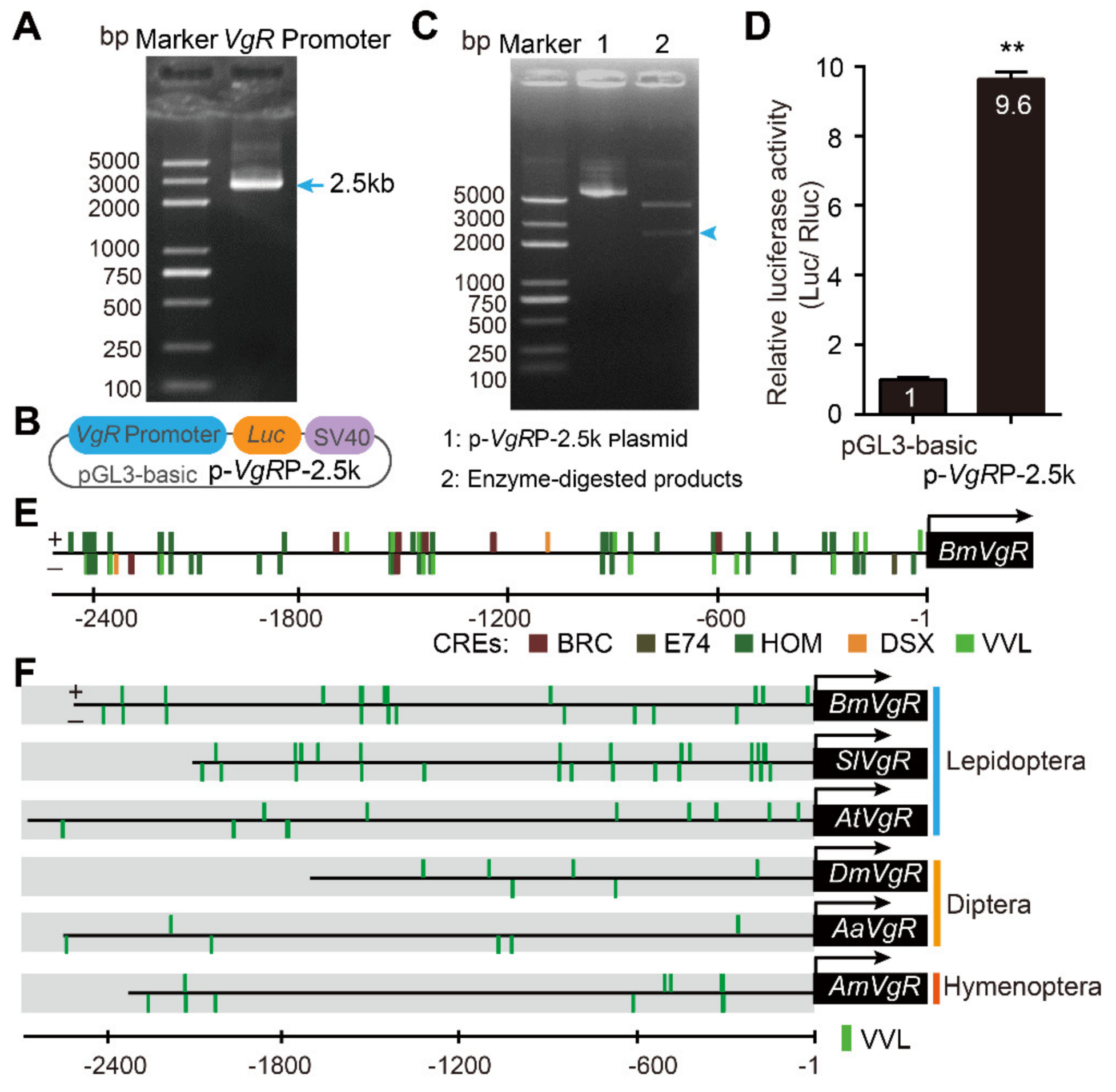
2.2. BmVgR Promoter Isolation and Bioinformatics Analysis
2.3. Vector Construction
2.4. Cell Culture and Dual Luciferase Reporter (DLR) Assay
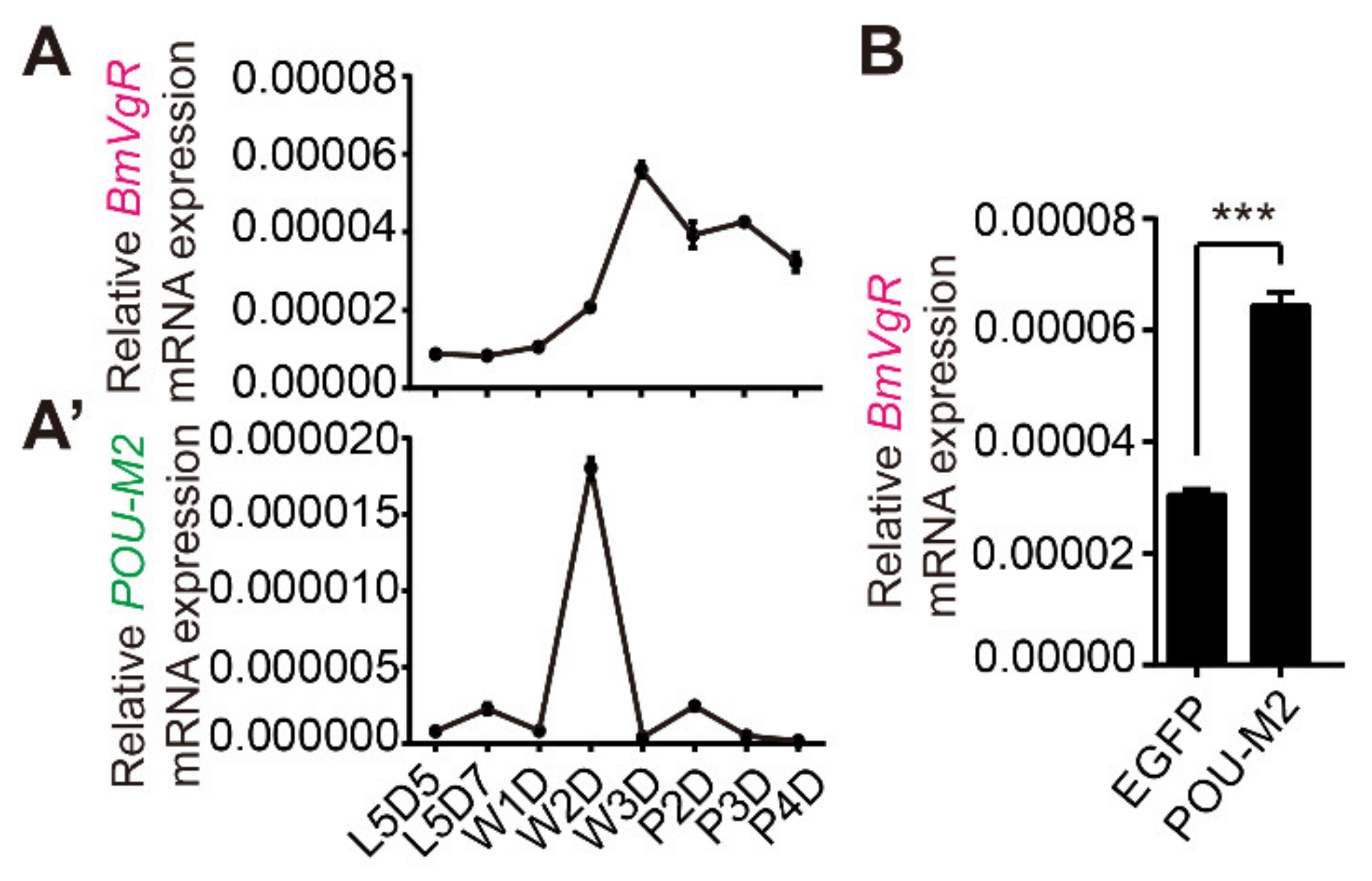
2.5. Quantitative Reverse Transcription PCR (RT-PCR) and RT-PCR
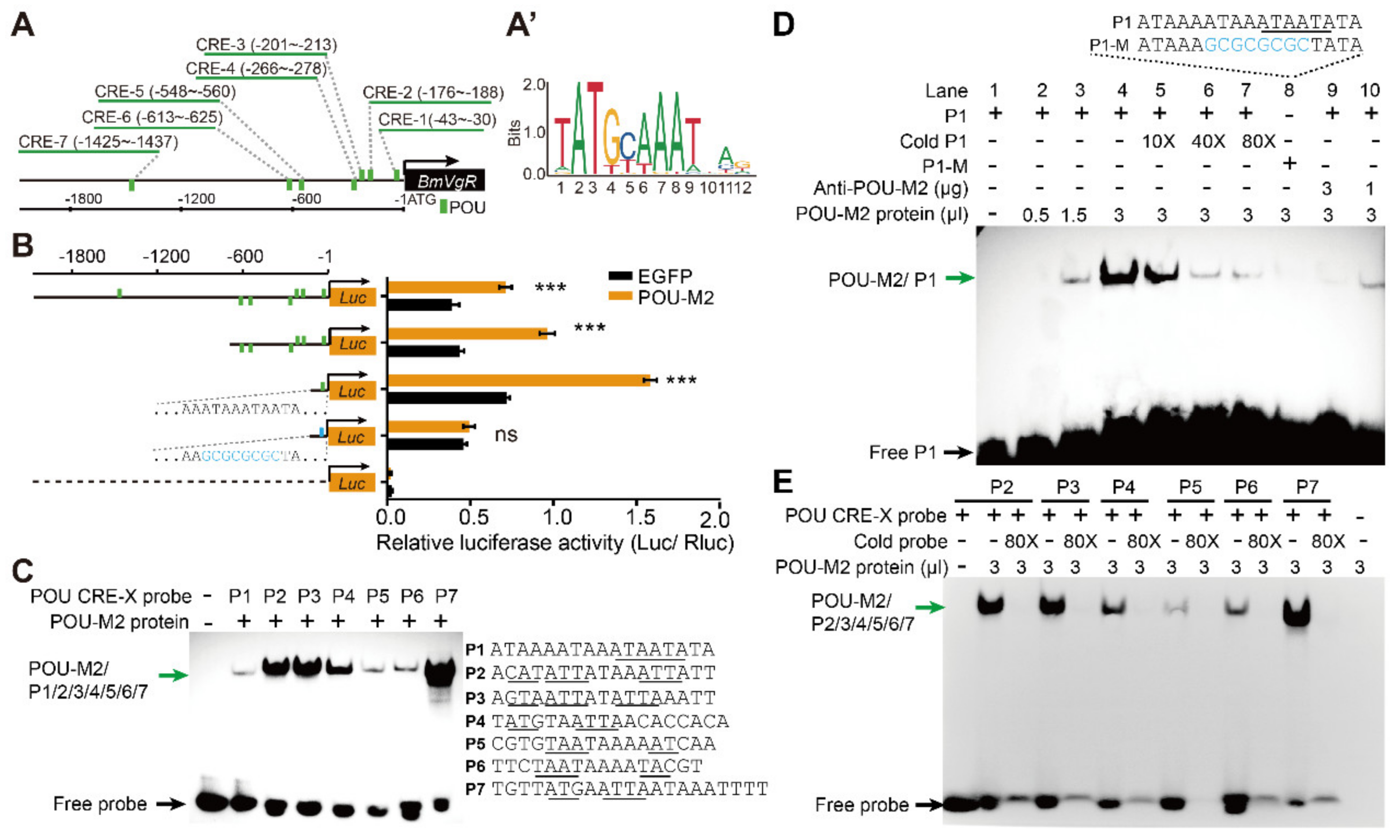
2.6. Electrophoretic Mobility Shift Assay (EMSA)
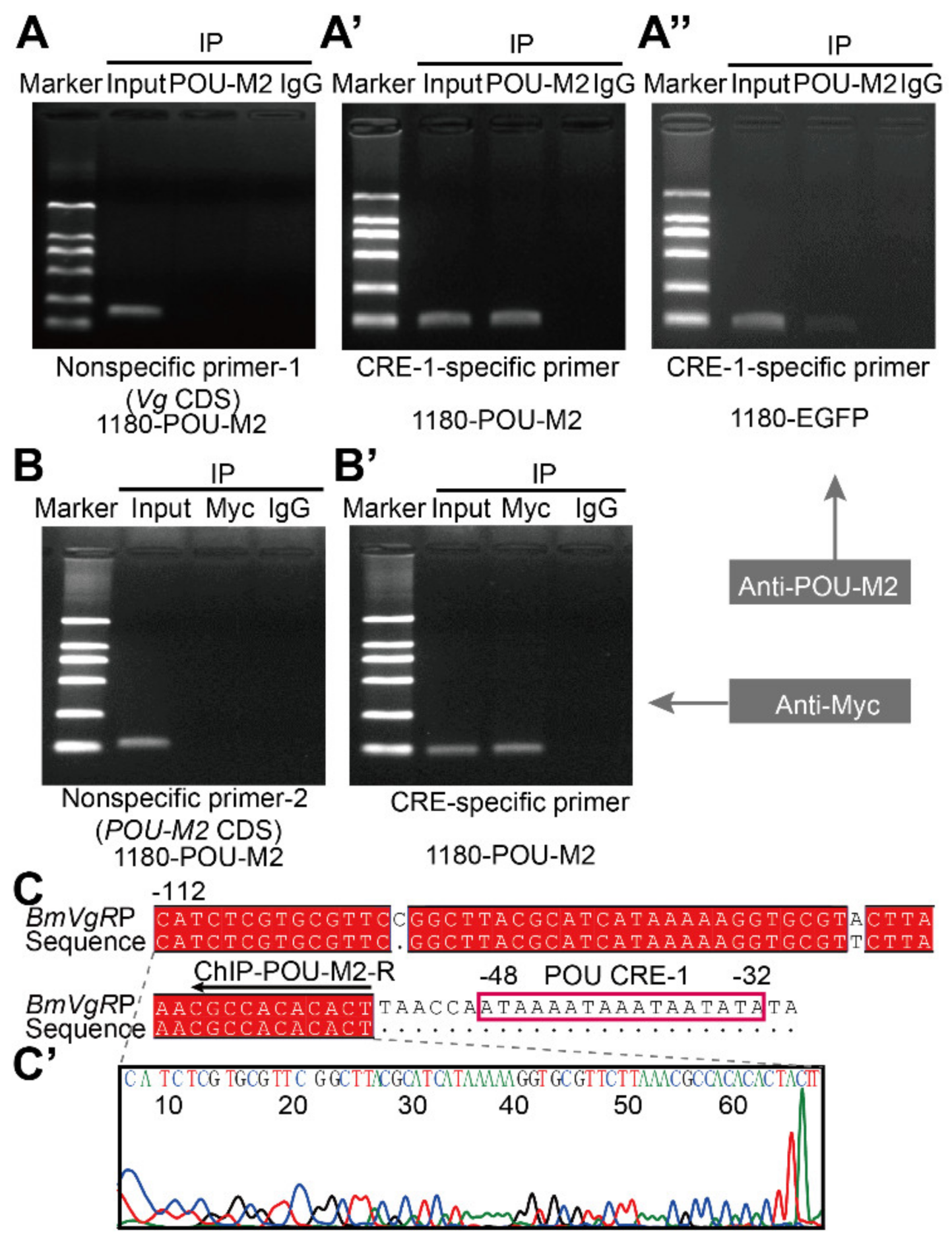
2.7. Chromatin Immunoprecipitation (ChIP)
2.8. RNA Interference (RNAi)
2.9. Statistical Analysis
3. Results
3.1. Cloning and Analysis of the BmVgR Gene Promoter
3.2. Analysis of the Expression Pattern of POU-M2 and BmVgR in Ovary
3.3. POU-M2 Regulated BmVgR Expression by Binding Directly to the POU CREs of the BmVgR Promoter
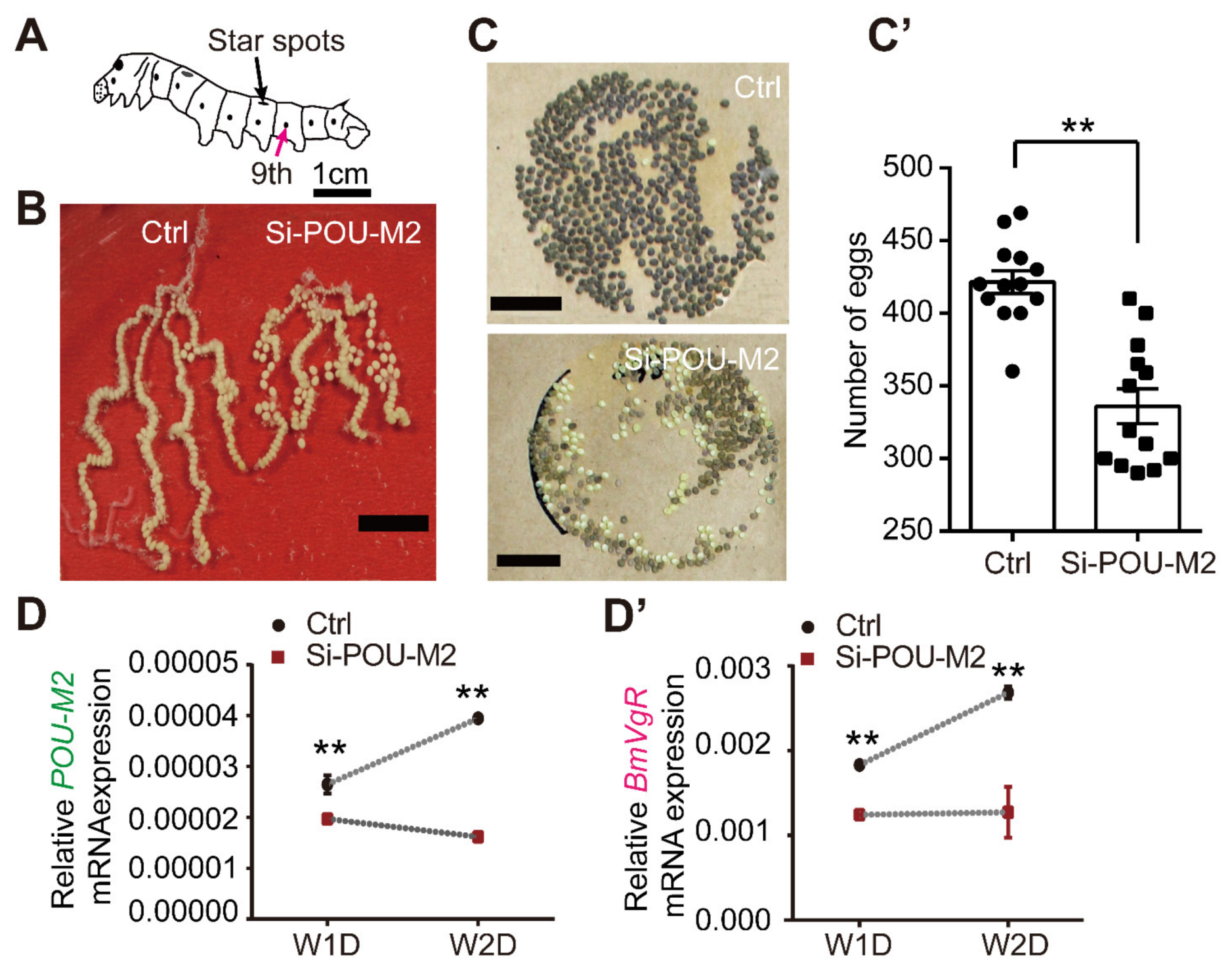
3.4. Suppression of POU-M2 by RNA Interference Reduces the Expression of BmVgR and the Egg-laying Rate
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Thomas, W.S.; Alexander, S.R. Molecular characteristics of insect vitellogenins and vitellogenin receptors. Insect Biochem. Mol. Biol. 1998, 28, 277–300. [Google Scholar]
- Tufail, M.; Takeda, M. Insect vitellogenin/lipophorin receptors: Molecular structures, role in oogenesis, and regulatory mechanisms. J. Insect Physiol. 2009, 55, 87–103. [Google Scholar] [CrossRef] [PubMed]
- Schneider, W.J. Vitellogenin receptors: Oocyte-specific members of the Low-Density Lipoprotein Receptor supergene family. Int. Rev. Cytol. 1996, 166, 103–137. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.E.; Lewis, D.K.; Keeley, L.L.; Pietrantonio, P.V. cDNA cloning and transcriptional regulation of the vitellogenin receptor from the imported fire ant, Solenopsis invicta Buren (Hymenoptera: Formicidae) Insect Mol. Biol. 2004, 13, 195–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, K.H.; Cheon, H.M.; Kokoza, V.; Raikhel, A.S. Regulatory region of the vitellogenin receptor gene sufficient for high-level, germ line cell-specific ovarian expression in transgenic Aedes aegypti mosquitoes. Insect Biochem. Mol. Biol. 2006, 36, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, G.A.; Quattro, J.M.; Denslow, N.D.; Kroll, K.J.; Prucha, M.S.; Porak, W.F.; Grier, H.J.; Sabo-Attwood, T.L. Identification and transcriptional modulation of the largemouth bass, Micropterus salmoides, vitellogenin receptor during oocyte development by insulin and sex steroids. Biol. Reprod. 2012, 87, 67. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, G.A.; Bisesi, J.H., Jr.; Kroll, K.J.; Denslow, N.D.; Sabo-Attwood, T. Control of transcriptional repression of the vitellogenin receptor gene in largemouth bass (Micropterus salmoides) by select estrogen receptors isotypes. Toxicol. Sci. 2014, 141, 423–431. [Google Scholar] [CrossRef] [Green Version]
- Lu, K.; Shu, Y.; Zhou, J.; Zhang, X.; Zhang, X.; Chen, M.; Yao, Q.; Zhou, Q.; Zhang, W. Molecular characterization and RNA interference analysis of vitellogenin receptor from Nilaparvata lugens (Stal). J. Insect Physiol. 2015, 73, 20–29. [Google Scholar] [CrossRef]
- Liu, W.; Guo, S.; Sun, D.; Zhu, L.; Zhu, F.; Lei, C.L.; Sheng, L.; Phelps, B.; Wang, X.P. Molecular characterization and juvenile hormone-regulated transcription of the vitellogenin receptor in the cabbage beetle Colaphellus bowringi. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2019, 229, 69–75. [Google Scholar] [CrossRef]
- Goldsmith, M.R.; Shimada, T.; Abe, H. The genetics and genomics of the silkworm, Bombyx mori. Annu. Rev. Entomol. 2005, 50, 71–100. [Google Scholar] [CrossRef]
- Xia, Q.; Li, S.; Feng, Q. Advances in silkworm studies accelerated by the genome sequencing of Bombyx mori. Annu. Rev. Entomol. 2014, 59, 513–536. [Google Scholar] [CrossRef] [PubMed]
- Xia, Q.Y.; Zhou, Z.Y.; Lu, C.; Cheng, D.J.; Dai, F.Y.; Li, B.; Zhao, P.; Zha, X.F.; Cheng, T.C.; Chai, C.L.; et al. A draft sequence for the genome of the domesticated silkworm (Bombyx mori). Science 2004, 306, 1937–1940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, Q.Y.; Guo, Y.R.; Zhang, Z.; Li, D.; Xuan, Z.L.; Li, Z.; Dai, F.Y.; Li, Y.R.; Cheng, D.J.; Li, R.Q.; et al. Complete Resequencing of 40 Genomes Reveals Domestication Events and Genes in Silkworm (Bombyx). Science 2009, 326, 433–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiuchi, T.; Koga, H.; Kawamoto, M.; Shoji, K.; Sakai, H.; Arai, Y.; Ishihara, G.; Kawaoka, S.; Sugano, S.; Shimada, T.; et al. A single female-specific piRNA is the primary determiner of sex in the silkworm. Nature 2014, 509, 633–636. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Sun, W.; Sun, B.; Xiao, Y.; Zhang, Z. The dynamic landscape of gene regulation during Bombyx mori oogenesis. BMC Genom. 2017, 18, 714. [Google Scholar] [CrossRef]
- Yamauchi, H.; Yoshitake, N. Developmental stages of ovarian follicles of the Silkworm, Bombyx mori L. J. Morphol. 1984, 179, 21–31. [Google Scholar] [CrossRef]
- Chen, E.; Chen, Z.; Li, S.; Xing, D.; Guo, H.; Liu, J.; Ji, X.; Lin, Y.; Liu, S.; Xia, Q. MicroRNAs bmo-miR-2739 and novel-miR-167 coordinately regulate the expression of the vitellogenin receptor in Bombyx mori oogenesis. Development 2020. [Google Scholar] [CrossRef]
- Nitta, K.R.; Jolma, A.; Yin, Y.; Morgunova, E.; Kivioja, T.; Akhtar, J.; Hens, K.; Toivonen, J.; Deplancke, B.; Furlong, E.E.; et al. Conservation of transcription factor binding specificities across 600 million years of bilateria evolution. Elife 2015, 4. [Google Scholar] [CrossRef] [Green Version]
- Ingraham, H.A.; Chen, R.; Mangalam, H.J.; Elsholtz, H.P.; Flynn, S.E.; Lin, C.R.; Simmons, D.M.; Swanson, L.; Rosenfeld, M.G. A tissue-specific transcription factor containing a homeodomain specifies a pituitary phenotype. Cell 1988, 55, 519–529. [Google Scholar] [CrossRef]
- Herr, W.; Sturm, R.A.; Clerc, R.G.; Corcoran, L.M.; Baltimore, D.; Sharp, P.A.; Ingraham, H.A.; Rosenfeld, M.G.; Finney, M.; Ruvkun, G.; et al. The POU domain: A large conserved region in the mammalian pit-1, oct-1, oct-2, and Caenorhabditis elegans unc-86 gene products. Genes Dev. 1988, 2, 1513–1516. [Google Scholar] [CrossRef]
- Clerc, R.G.; Corcoran, L.M.; LeBowitz, J.H.; Baltimore, D.; Sharp, P.A. The B-cell-specific Oct-2 protein contains POU box- and homeo box-type domains. Genes Dev. 1988, 2, 1570–1581. [Google Scholar] [CrossRef] [PubMed]
- Finney, M.; Ruvkun, G.; Horvitz, H.R. The C. elegans Cell Lineage and Differentiation Gene uric-86 Encodes a Protein with a Homeodomain and Extended Similarity to Transcription Factors. Cell 1988, 55, 757–769. [Google Scholar] [CrossRef]
- Tang, X.; Engstrom, Y. Regulation of immune and tissue homeostasis by Drosophila POU factors. Insect Biochem. Mol. Biol. 2019, 109, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Ryan, A.K.; Rosenfeld, M.G. POU domain family values: Flexibility, partnerships, and developmental codes. Genes Dev. 1997, 11, 1207–1225. [Google Scholar] [CrossRef] [Green Version]
- Takiya, S.; Kokubo, H.; Suzuki, Y. Transcriptional regulatory elements in the upstream and intron of the fibroin gene bind three specific factors POU-M1, Bm Fkh and FMBP-1. Biochem. J. 1997, 321 Pt 3, 645–653. [Google Scholar] [CrossRef] [Green Version]
- Matsunami, K.; Kokubo, H.; Ohno, K.; Suzuki, Y. Expression pattern analysis of SGF-3/POU-M1 in relation to sericin-1 gene expression in the silk gland. Dev. Growth Differ. 1998, 40, 591–597. [Google Scholar] [CrossRef]
- Zhang, T.Y.; Kang, L.; Zhang, Z.F.; Xu, W.H. Identification of a POU factor involved in regulating the neuron-specific expression of the gene encoding diapause hormone and pheromone biosynthesis-activating neuropeptide in Bombyx mori. Biochem. J. 2004, 380, 255–263. [Google Scholar] [CrossRef]
- Deng, H.; Zheng, S.; Yang, X.; Liu, L.; Feng, Q. Transcription factors BmPOUM2 and BmbetaFTZ-F1 are involved in regulation of the expression of the wing cuticle protein gene BmWCP4 in the silkworm, Bombyx mori. Insect Mol. Biol. 2011, 20, 45–60. [Google Scholar] [CrossRef]
- Deng, H.; Zhang, J.; Li, Y.; Zheng, S.; Liu, L.; Huang, L.; Xu, W.H.; Palli, S.R.; Feng, Q. Homeodomain POU and Abd-A proteins regulate the transcription of pupal genes during metamorphosis of the silkworm, Bombyx mori. Proc. Natl. Acad. Sci. USA 2012, 109, 12598–12603. [Google Scholar] [CrossRef] [Green Version]
- Meng, M.; Cheng, D.J.; Peng, J.; Qian, W.L.; Li, J.R.; Dai, D.D.; Zhang, T.L.; Xia, Q.Y. The homeodomain transcription factors antennapedia and POU-M2 regulate the transcription of the steroidogenic enzyme gene Phantom in the silkworm. J. Biol. Chem. 2015, 290, 24438–24452. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Liu, H.; Yang, C.; Gu, J.; Shen, G.; Zhang, H.; Chen, E.; Han, C.; Zhang, Y.; Xu, Y.; et al. The POU homeodomain transcription factor POUM2 and broad complex isoform 2 transcription factor induced by 20-hydroxyecdysone collaboratively regulate vitellogenin gene expression and egg formation in the silkworm Bombyx mori. Insect Mol. Biol. 2017, 26, 496–506. [Google Scholar] [CrossRef] [PubMed]
- Silkworm genome database. Available online: http://silkworm.swu.edu.cn/silkdb (accessed on 1 December 2018).
- MatInspector program. Available online: http://www.genomatix.de/ (accessed on 10 January 2019).
- JASPAR 2020. Available online: http://jaspar.genereg.net/ (accessed on 1 January 2020).
- Yang, C.; Lin, Y.; Liu, H.; Shen, G.; Luo, J.; Zhang, H.; Peng, Z.; Chen, E.; Xing, R.; Han, C.; et al. The Broad Complex isoform 2 (BrC-Z2) transcriptional factor plays a critical role in vitellogenin transcription in the silkworm Bombyx mori. Biochim. Biophys. Acta 2014, 1840, 2674–2684. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.-H.; Xia, Q.-Y.; Cheng, D.-J.; Duan, J.; Zhao, P.; Chen, J.; Zhu, L. Reference genes identified in the silkwormBombyx moriduring metamorphism based on oligonucleotide microarray and confirmed by qRT-PCR. Insect Sci. 2008, 15, 405–413. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2—ΔΔC(T) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Liu, L.; Li, Y.; Wang, Y.; Zhao, P.; Wei, S.; Li, Z.; Chang, H.; He, H. Biochemical characterization and functional analysis of the POU transcription factor POU-M2 of Bombyx mori. Int. J. Biol. Macromol. 2016, 86, 701–708. [Google Scholar] [CrossRef]
- Suzuki, M.G.; Ohbayashi, F.; Mita, K.; Shimada, T. The mechanism of sex-specific splicing at the doublesex gene is different between Drosophila melanogaster and Bombyx mori. Insect Biochem. Mol. Biol. 2001, 31, 1201–1211. [Google Scholar] [CrossRef]
- Vonkalm, L.; Crossgrove, K.; Vonseggern, D.; Guild, G.M.; Beckendorf, S.K. The Broad-Complex Directly Controls a Tissue-Specific Response to the Steroid-Hormone Ecdysone at the Onset of Drosophila Metamorphosis. Embo J. 1994, 13, 3505–3516. [Google Scholar] [CrossRef]
- Urness, L.D.; Thummel, C.S. Molecular-Interactions within the Ecdysone Regulatory Hierarchy—DNA-Binding Properties of the Drosophila Ecdysone-Inducible E74a Protein. Cell 1990, 63, 47–61. [Google Scholar] [CrossRef]
- Lin, Y.; Meng, Y.; Wang, Y.X.; Luo, J.; Katsuma, S.; Yang, C.W.; Banno, Y.; Kusakabe, T.; Shimada, T.; Xia, Q.Y. Vitellogenin receptor mutation leads to the oogenesis mutant phenotype “scanty vitellin” of the silkworm, Bombyx mori. J. Biol. Chem. 2013, 288, 13345–13355. [Google Scholar] [CrossRef] [Green Version]
- Andersen, B.; Rosenfeld, M.G. POU domain factors in the neuroendocrine system: Lessons from developmental biology provide insights into human disease. Endocr. Rev. 2001, 22, 2–35. [Google Scholar] [CrossRef]
- Singh, H.; Sen, R.; Baltimore, D.; Sharp, P.A. A nuclear factor that binds to a conserved sequence motif in transcriptional control elements of immunoglobulin genes. Nature 1986, 319, 154–158. [Google Scholar] [CrossRef] [PubMed]
- Certel, K.; Anderson, M.G.; Shrigley, R.J.; Johnson, W.A. Distinct variant DNA-binding sites determine cell-specific autoregulated expression of the Drosophila POU domain transcription factor drifter in midline glia or trachea. Mol. Cell Biol. 1996, 16, 1813–1823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Junell, A.; Uvell, H.; Davis, M.M.; Edlundh-Rose, E.; Antonsson, A.; Pick, L.; Engstrom, Y. The POU transcription factor Drifter/Ventral veinless regulates expression of Drosophila immune defense genes. Mol. Cell Biol. 2010, 30, 3672–3684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, P.X.; Fukuta, M.; Takiya, S.; Matsuno, K.; Xu, X.; Suzuki, Y. Promoter of the Pou-M1/Sgf-3 Gene Involved in the Expression of Bombyx Silk Genes. J. Biol. Chem. 1994, 269, 2733–2742. [Google Scholar] [PubMed]
- Fukuta, M.; Matsuno, K.; Hui, C.C.; Nagata, T.; Takiya, S.; Xu, P.X.; Ueno, K.; Suzuki, Y. Molecular cloning of a POU domain-containing factor involved in the regulation of the Bombyx sericin-1 gene. J. Biol. Chem. 1993, 268, 19471–19475. [Google Scholar]
- Venkatarama, T.; Lai, F.F.; Luo, X.T.; Zhou, Y.; Newman, K.; King, M.L. Repression of zygotic gene expression in the Xenopus germline. Development 2010, 137, 651–660. [Google Scholar] [CrossRef] [Green Version]
- Morichika, K.; Sugimoto, M.; Yasuda, K.; Kinoshita, T. Possible regulation of Oct60 transcription by a positive feedback loop in Xenopus oocytes. Zygote 2014, 22, 266–274. [Google Scholar] [CrossRef]
- Greenstein, D.; Furuta, T. The POU-homeobox gene ceh-18 is a crucial determinant of gonadal sheath cell function during oogenesis of Caenorhabditis elegans. Dev. Biol. 1996, 175, C13. [Google Scholar]
- Gao, J.N.; Wang, X.B.; Zhang, Q.Q. Evolutionary Conservation of pou5f3 Genomic Organization and Its Dynamic Distribution during Embryogenesis and in Adult Gonads in Japanese Flounder Paralichthys olivaceus. Int. J. Mol. Sci. 2017, 18, 231. [Google Scholar] [CrossRef] [Green Version]
- Roy, S.; Saha, T.T.; Zou, Z.; Raikhel, A.S. Regulatory Pathways Controlling Female Insect Reproduction. Annu. Rev. Entomol. 2018, 63, 489–511. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, G.; Chen, E.; Ji, X.; Liu, L.; Liu, J.; Hua, X.; Li, D.; Xiao, Y.; Xia, Q. The POU Transcription Factor POU-M2 Regulates Vitellogenin Receptor Gene Expression in the Silkworm, Bombyx mori. Genes 2020, 11, 394. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11040394
Shen G, Chen E, Ji X, Liu L, Liu J, Hua X, Li D, Xiao Y, Xia Q. The POU Transcription Factor POU-M2 Regulates Vitellogenin Receptor Gene Expression in the Silkworm, Bombyx mori. Genes. 2020; 11(4):394. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11040394
Chicago/Turabian StyleShen, Guanwang, Enxiang Chen, Xiaocun Ji, Lina Liu, Jianqiu Liu, Xiaoting Hua, Dan Li, Yingdan Xiao, and Qingyou Xia. 2020. "The POU Transcription Factor POU-M2 Regulates Vitellogenin Receptor Gene Expression in the Silkworm, Bombyx mori" Genes 11, no. 4: 394. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11040394