The Role of Metabolites in the Link between DNA Replication and Central Carbon Metabolism in Escherichia coli

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Metabolites
2.3. Effects of Metabolites on the Growth of Bacterial Strains
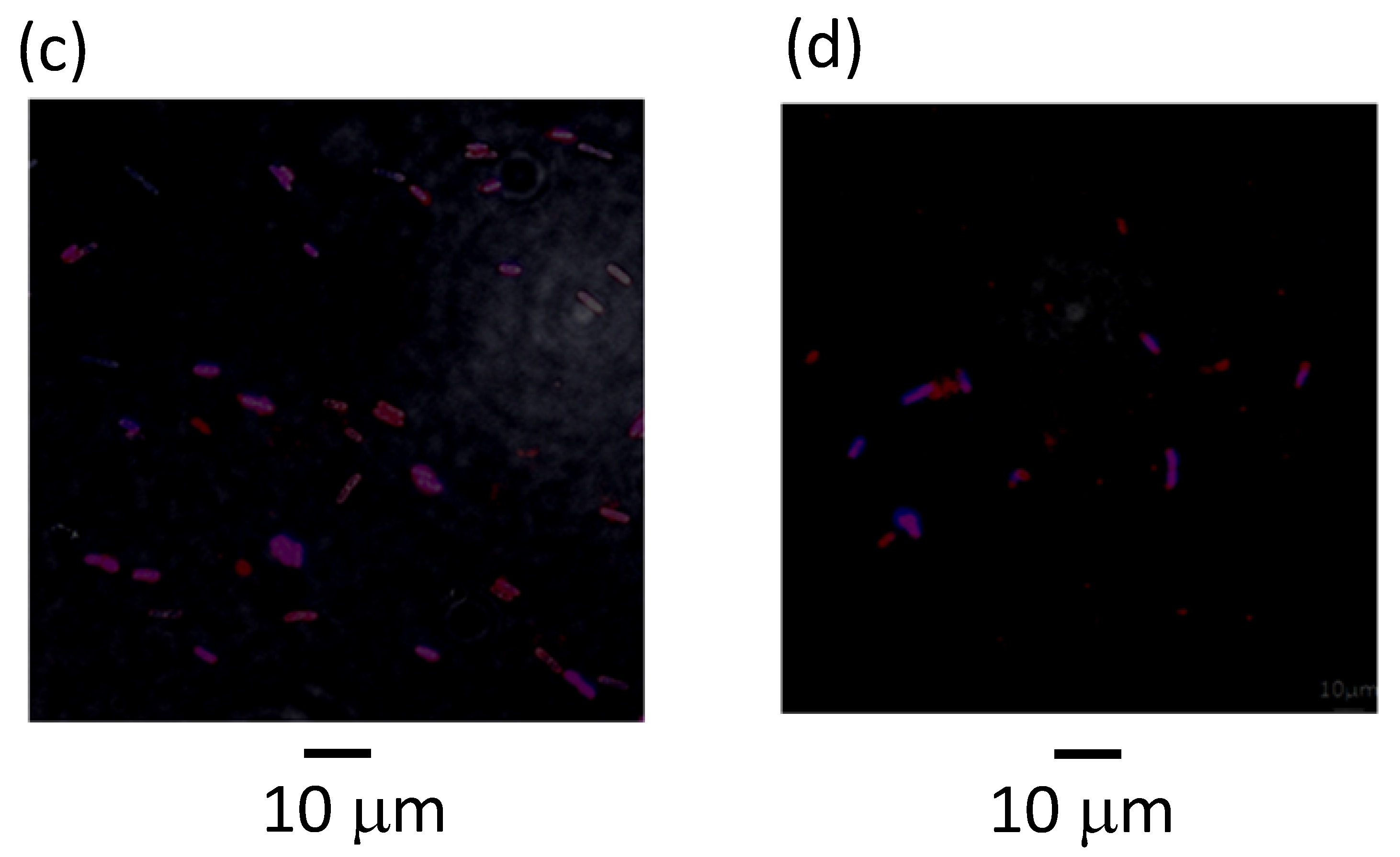
2.4. Microscopic Analysis
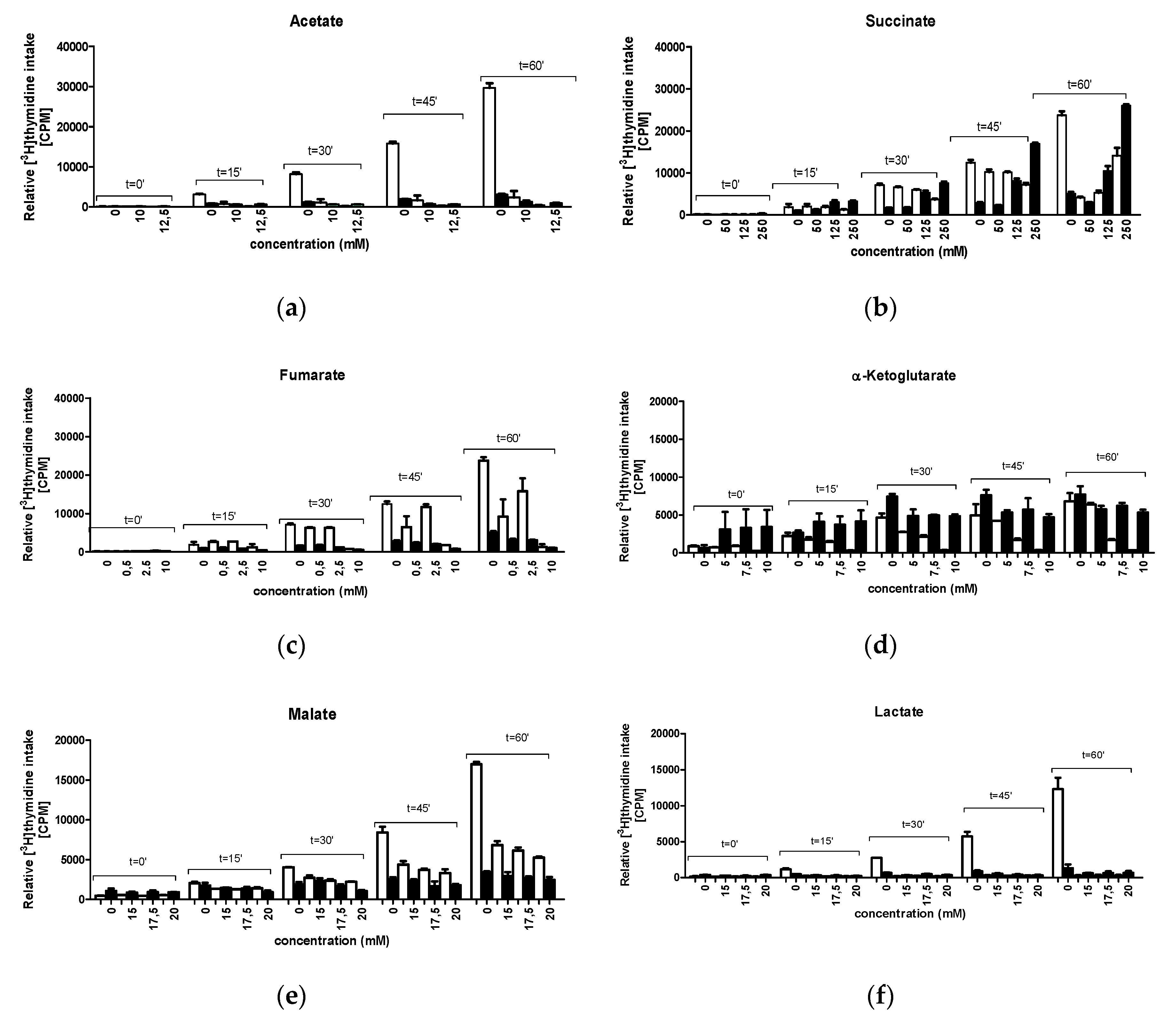
2.5. Kinetics of DNA Replication In Vivo
3. Results
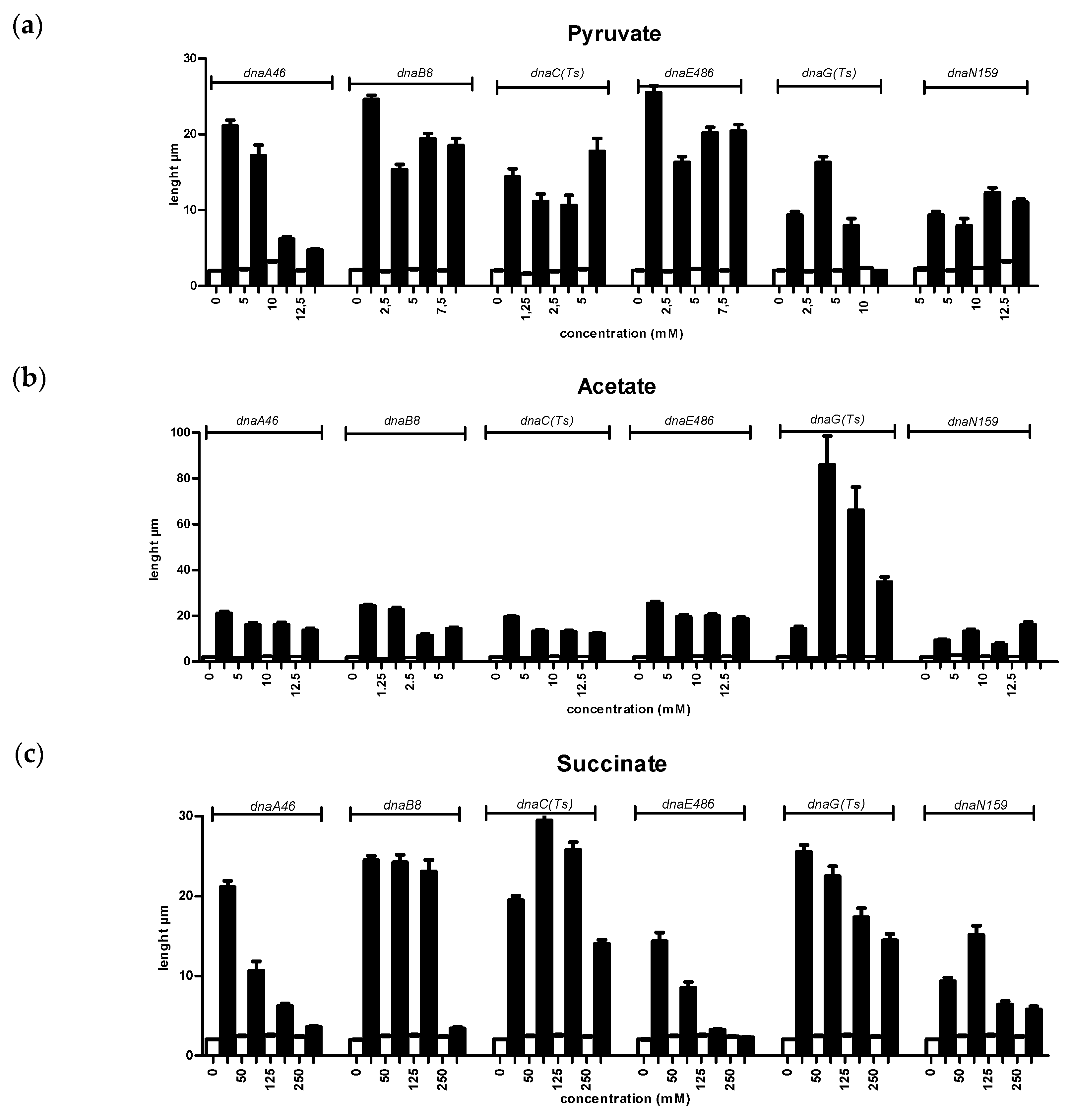
3.1. Growth Defect Suppression of Replication Mutant Strains by Certain CCM Metabolites
3.2. Effects of Metabolites on Filamentation of the Replication Mutants at Elevated Temperature
3.3. Kinetics of DNA Replication in the Presence of CCM Metabolites
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kornberg, A.; Baker, T.A. DNA Replication, 2nd ed.; Lilley, D., Ed.; University Science Books: Mill Valley, CA, USA, 1992; Volume 17, p. 271. [Google Scholar] [CrossRef]
- Gottschalk, G. Bacterial Metabolism, 2nd ed.; Springer: Berlin/Heidelberg, Germay, 1986. [Google Scholar]
- Jannière, L.; Canceill, D.; Suski, C.; Kanga, S.; Dalmais, B.; Lestini, R.; Monnier, A.F.; Chapuis, J.; Bolotin, A.; Titok, M.; et al. Genetic evidence for a link between glycolysis and DNA replication. PLoS ONE 2007, 2, e447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nouri, H.; Monnier, A.F.; Fossum-Raunehaug, S.; Maciag-Dorszynska, M.; Cabin-Flaman, A.; Képès, F.; Wegrzyn, G.; Szalewska-Palasz, A.; Norris, V.; Skarstad, K.; et al. Multiple links connect central carbon metabolism to DNA replication initiation and elongation in Bacillus subtilis. DNA Res. 2018, 25, 641–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maciąg, M.; Nowicki, D.; Janniere, L.; Szalewska-Pałasz, A.; Węgrzyn, G. Genetic response to metabolic fluctuations: Correlation between central carbon metabolism and DNA replication in Escherichia coli. Microb. Cell Factories 2011, 10, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barańska, S.; Glinkowska, M.; Herman-Antosiewicz, A.; Maciąg-Dorszyńska, M.; Nowicki, D.; Szalewska-Pałasz, A.; Węgrzyn, A.; Węgrzyn, G. Replicating DNA by cell factories: Roles of central carbon metabolism and transcription in the control of DNA replication in microbes, and implications for understanding this process in human cells. Microb. Cell Factories 2013, 12, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konieczna, A.; Szczepańska, A.; Sawiuk, K.; Łyżeń, R.; Węgrzyn, G. Enzymes of the central carbon metabolism: Are they linkers between transcription, DNA replication, and carcinogenesis? Med. Hypotheses 2015, 84, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Fornalewicz, K.; Wieczorek, A.; Węgrzyn, G.; Łyżeń, R. Silencing of the pentose phosphate pathway genes influences DNA replication in human fibroblasts. Gene 2017, 635, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Wieczorek, A.; Fornalewicz, K.; Mocarski, Ł.; Łyżeń, R.; Węgrzyn, G. Double silencing of relevant genes suggests the existence of the direct link between DNA replication/repair and central carbon metabolism in human fibroblasts. Gene 2018, 15, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Glinkowska, M.; Boss, L.; Węgrzyn, G. DNA Replication Control in Microbial Cell Factories. In SpringerBriefs in Microbiology; Springer: Heidelberg, Germany, 2015. [Google Scholar] [CrossRef]
- Konieczna, A.; Szczepańska, A.; Sawiuk, K.; Wegrzyn, G.; Łyżeń, R. Effects of partial silencing of genes coding for enzymes involved in glycolysis and tricarboxylic acid cycle on the enterance of human fibroblasts to the S phase. BMC Cell Biol. 2015, 16, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, K.F. The Escherichia coli K-12 "wild types" W3110 and MG1655 have an rph frameshift mutation that leads to pyrimidine starvation due to low pyrE expression levels. J. Bacteriol. 1993, 175, 3401–3407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fayet, O.; Louarn, J.M.; Georgopoulos, C. Suppression of the Escherichia coli dnaA46 mutation by amplification of the groES and groEL genes. Mol. Gen. Genet. 1986, 202, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Carl, P.L. Escherichia coli mutants with temperature-sensitive synthesis of DNA. Mol. Gen. Genet. 1970, 109, 107–122. [Google Scholar] [CrossRef] [PubMed]
- Dabbs, E.R. The gene for ribosomal protein S21, rpsU, maps close to dnaG at 66.5 min on the Escherichia coli chromosomal linkage map. J. Bacteriol. 1980, 144, 603–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maciąg, M.; Nowicki, D.; Szalewska-Pałasz, A.; Węgrzyn, G. Central carbon metabolism influences fidelity of DNA replication in Escherichia coli. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2012, 731, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Maciąg-Dorszyńska, M.; Ignatowska, M.; Jannière, L.; Węgrzyn, G.; Szalewska-Pałasz, A. Mutations in central carbon metabolism genes suppress defects in nucleoid position and cell division of replication mutants in Escherichia coli. Gene 2012, 503, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Tymecka-Mulik, J.; Boss, L.; Maciąg-Dorszyńska, M.; Matias Rodrigues, J.F.; Gaffke, L.; Wosinski, A.; Cech, G.M.; Szalewska-Pałasz, A.; Węgrzyn, G.; Glinkowska, M. Suppression of the Escherichia coli dnaA46 mutation by changes in the activities of the pyruvate-acetate node links DNA replication regulation to central carbon metabolism. PLoS ONE 2017, 12, e0176050. [Google Scholar] [CrossRef] [PubMed]
- Murray, H.; Koh, A. Multiple regulatory systems coordinate DNA replication with cell growth in Bacillus subtilis. PLoS Genet. 2014, 10, e1004731. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Protein (Gene Product) | Protein Activity |
|---|---|---|
| dnaA | DnaA; replication initiator | Binding to the replication origin |
| dnaB | DnaB; DNA helicase | DNA unwinding in replication forks |
| dnaC | DnaC | Delivery of DnaB to the origin |
| dnaE | DnaE; a subunit of DNA polymerase III | DNA synthesis during replication |
| dnaG | DnaG; primase | Synthesis of primers during replication |
| dnaN | DnaN; b subunit of DNA polymerase III | Formation of the clamp conferring processivity during replication |
| ackA | Acetate kinase | Formation of acetyl phosphate from acetate and ATP |
| gpmA | 2,3-bisphosphoglycerate-dependent phosphoglycerate mutase | Interconversion of 2-phosphoglycerate and 3-phosphoglycerate |
| pgi | Glucose-6-phosphate isomerase | Isomerization of aldehydo-D-glucose 6-phosphate to keto-D-fructose 6-phosphate |
| pta | Phosphate acetyltransferase | Interconversion of acetyl-CoA and acetyl phosphate |
| tktB | Transketolase 2 | Production of xylulose-5-phosphate and ribose-5-phosphate from sedoheptulose-7-phosphate and glyceraldehyde-3-phosphate |
| Strain | Genotype | Reference |
|---|---|---|
| MG1655 | F- l- ilvG rfb-50 rph-1 | [12] |
| dnaA46 | F- l- ilvG rfb-50 rph-1 dnaA46 tna::Tn10 | [13] |
| dnaC(ts) | leu thy rpsL dnaC(ts) | [14] |
| dnaB8 | F- l- ilvG rfb-50 rph-1 dnaB8 cmR | [5,14] |
| dnaE486 | F- l- ilvG rfb-50 rph-1 dnaE486 zae502::Tn10 | [5] |
| dnaG(ts) | leu thy rpsL dnaG(ts) | [15] |
| dnaN159 | F- l- ilvG rfb-50 rph-1 dnaN159 zid501::Tn10 | [16] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krause, K.; Maciąg-Dorszyńska, M.; Wosinski, A.; Gaffke, L.; Morcinek-Orłowska, J.; Rintz, E.; Bielańska, P.; Szalewska-Pałasz, A.; Muskhelishvili, G.; Węgrzyn, G. The Role of Metabolites in the Link between DNA Replication and Central Carbon Metabolism in Escherichia coli. Genes 2020, 11, 447. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11040447
Krause K, Maciąg-Dorszyńska M, Wosinski A, Gaffke L, Morcinek-Orłowska J, Rintz E, Bielańska P, Szalewska-Pałasz A, Muskhelishvili G, Węgrzyn G. The Role of Metabolites in the Link between DNA Replication and Central Carbon Metabolism in Escherichia coli. Genes. 2020; 11(4):447. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11040447
Chicago/Turabian StyleKrause, Klaudyna, Monika Maciąg-Dorszyńska, Anna Wosinski, Lidia Gaffke, Joanna Morcinek-Orłowska, Estera Rintz, Patrycja Bielańska, Agnieszka Szalewska-Pałasz, Georgi Muskhelishvili, and Grzegorz Węgrzyn. 2020. "The Role of Metabolites in the Link between DNA Replication and Central Carbon Metabolism in Escherichia coli" Genes 11, no. 4: 447. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11040447