Differentially Expressed Genes of Natural Killer Cells Can Distinguish Rheumatoid Arthritis Patients from Healthy Controls



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. In Silico Prediction of the Percentage and Status of Immune Cells in Blood of RA Patients
2.2. Using Publicly Available Data to Identify Differentially Expressed Genes (DEGs) in NK Cells
2.3. RA Patients and Healthy Controls Sample Collection
2.4. Isolation of PBMCs and NK Cells from Peripheral Blood
2.5. RNA Extraction, Primer Design and RT-qPCR
2.6. ELISA Quantification of Plasma CXCL16, IFN-γ and IL-1β
2.7. Statistical Analysis
3. Results
3.1. Reduced Activated NK Cells in the Peripheral Blood of RA Patients
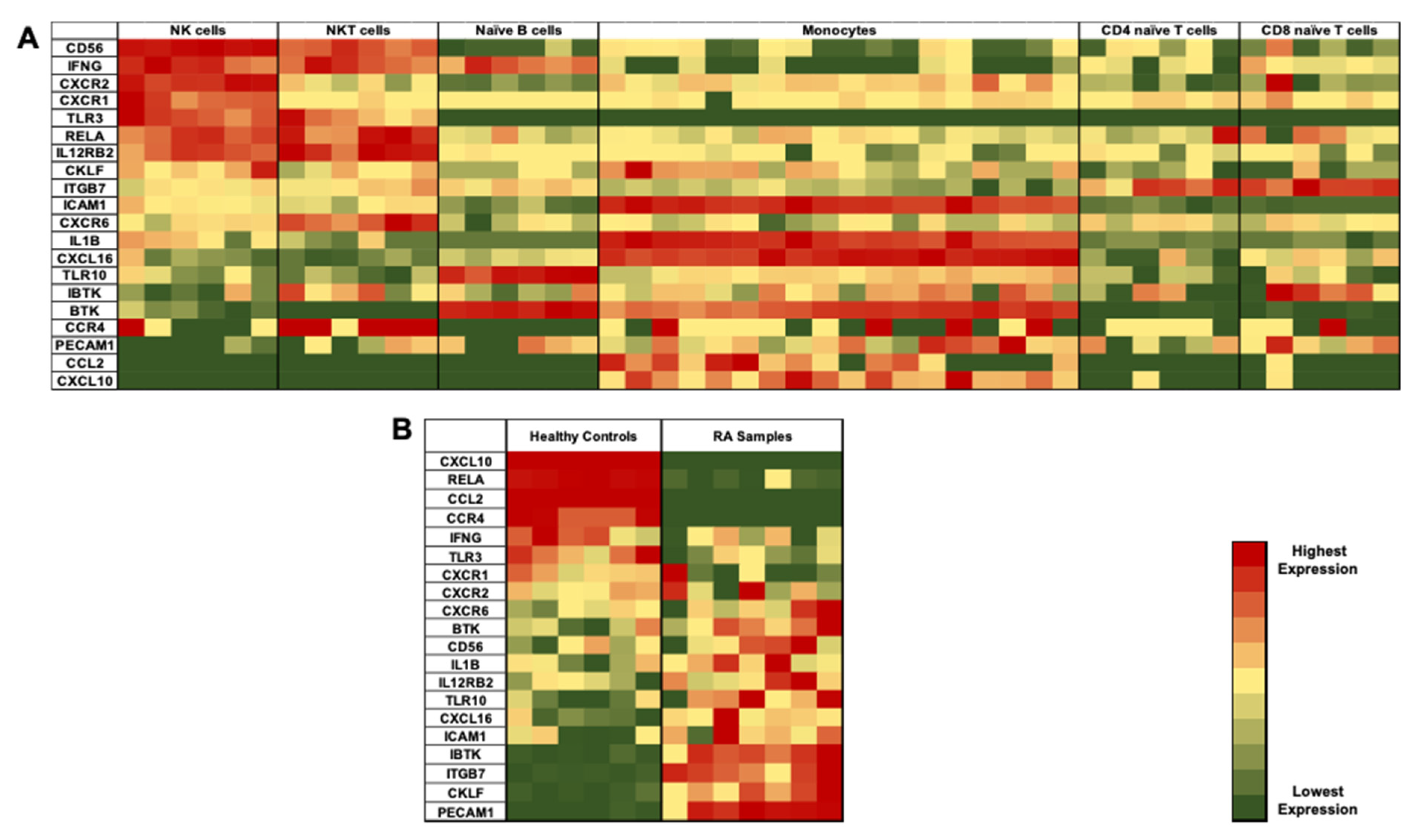
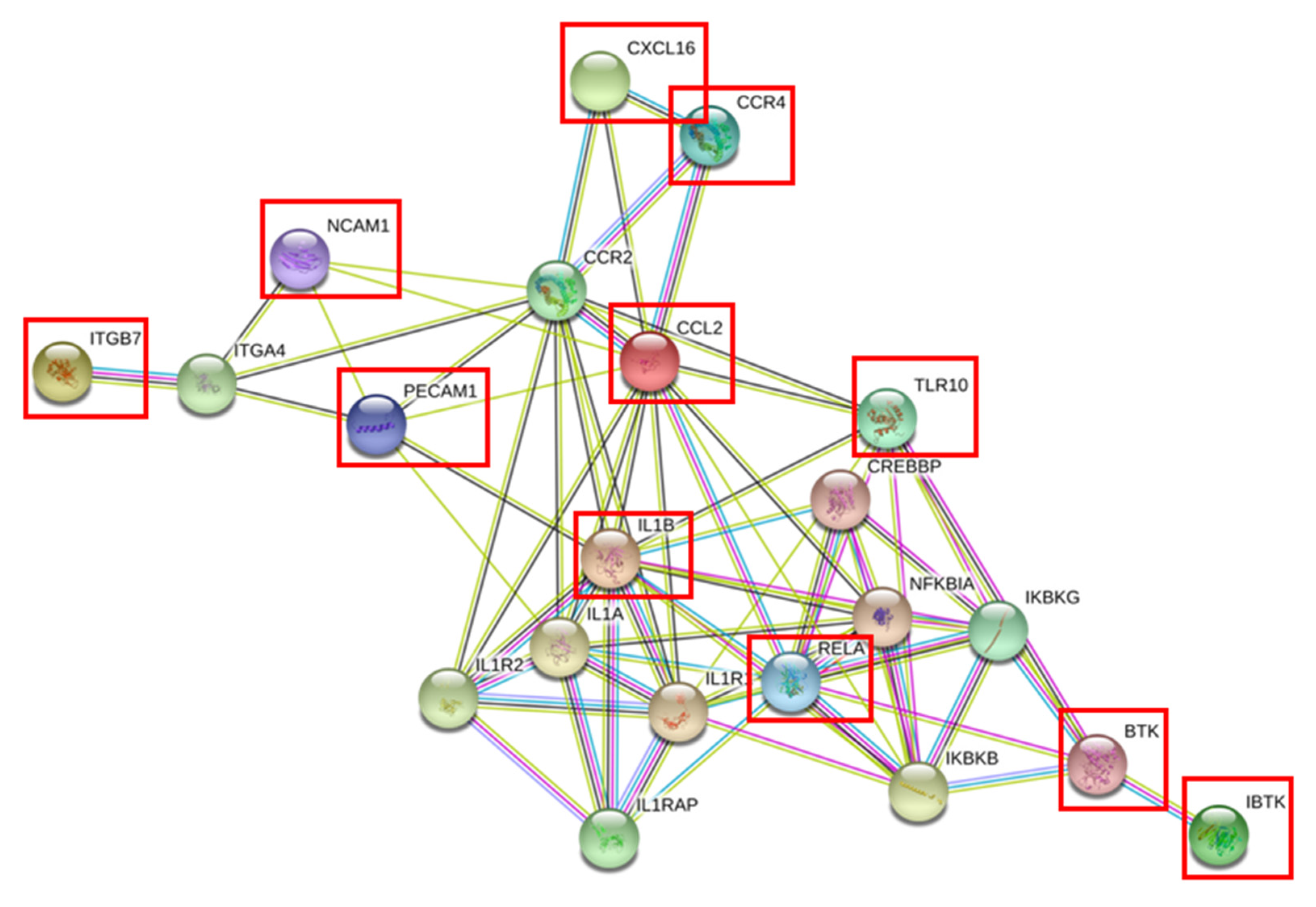
3.2. Identification of Differentially Expressed Genes (DEGs) in NK Cells of RA Patients
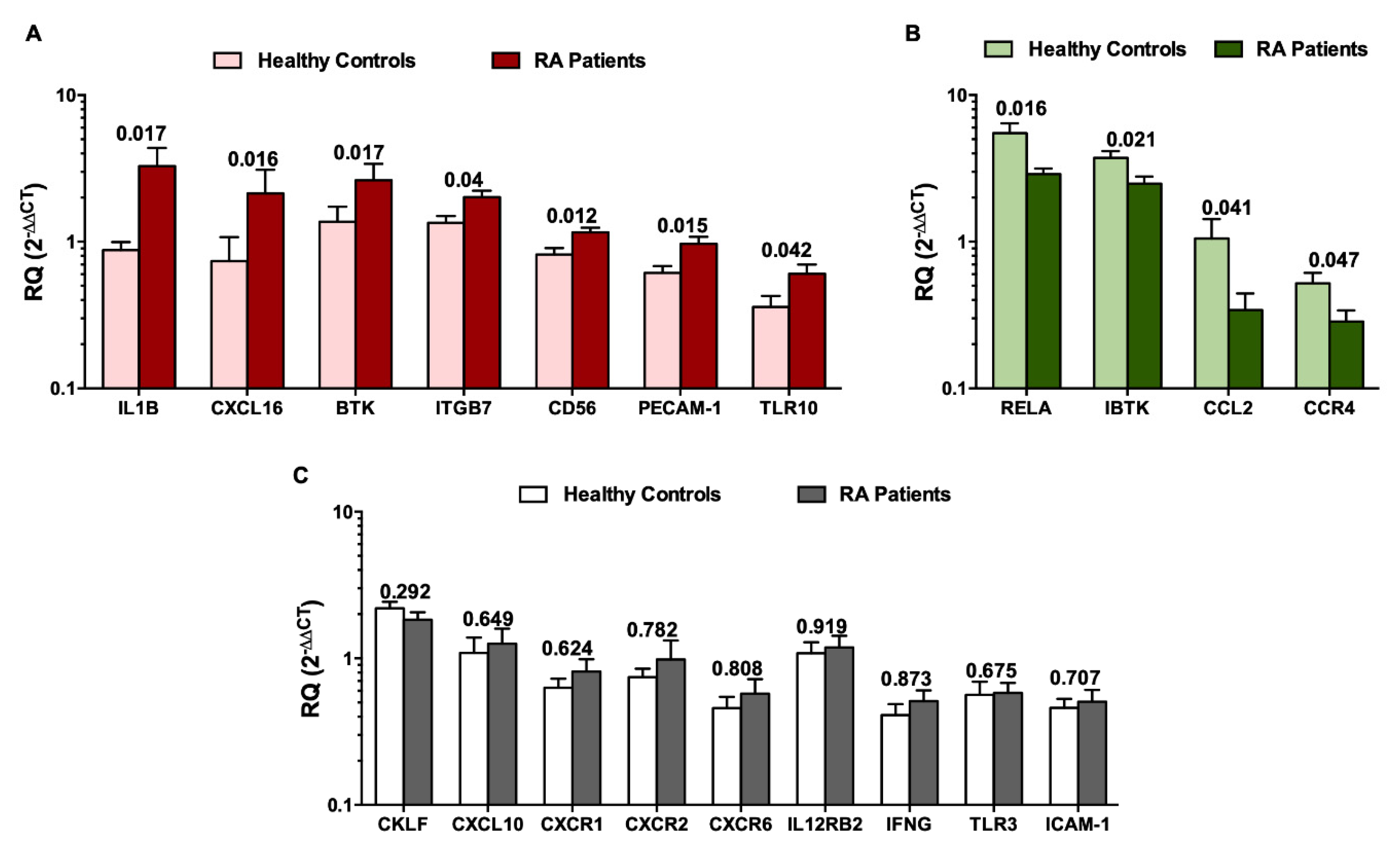
3.3. Validation of DEGs in Recruited Individuals’ PBMCs and NK Cells
3.4. Gene Expression Profiling of NK Cells in RA Patients
3.5. Clinical Data of Recruited Healthy Controls and RA Patients and Correlation with DEGs in NK Cells
3.6. Plasma Cytokines and Chemokines as Potential Biomarkers
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 18S | 18S ribosomal RNA |
| ACPAs | Anti-citrullinated peptide antibodies |
| ACR-EULAR. | American college of rheumatology-european league against rheumatism criteria |
| Anti-CCP | Anti-citrullinated peptide |
| bDMARDs | Biological disease modifying anti-rheumatic drugs |
| BTK | Bruton’s tyrosine kinase |
| CCL | C chemokine ligand |
| CCR | C chemokine receptor |
| CD56 | Cluster of differentiation 56 |
| CIA | Collagen induced arthritis |
| CKLF | Chemokine like factor |
| CRP | C- reactive peptide |
| CXCL | CX chemokine ligand |
| CXCR | CX chemokine receptor |
| DEGs | Differentially expressed genes |
| ELISA | Enzyme linked immunosorbent assay |
| ESR | Erythrocyte sedimentation rate |
| GAPDH | Glyceraldehyde 3-phosphate dehydrogenase |
| GSEA | Gene set enrichment analysis |
| IBTK | Inhibitor of Bruton’s tyrosine kinase |
| ICAM-1 | Intercellular adhesion molecule-1 |
| IFNG | Interferon gamma |
| IL- | Interleukin |
| ILC | Innate lymphoid cells |
| ITGB7 | Integrin beta 7 |
| JAK | Janus kinase |
| MS | Multiple sclerosis |
| NK cells | Natural killer cells |
| OA | Osteoarthritis |
| PBMCs | Peripheral blood mononuclear cells |
| PECAM-1 | Platelet endothelial cell adhesion molecule-1 |
| RA | Rheumatoid arthritis |
| RANK | Receptor activator of nuclear factor kappa-B |
| RELA | p65 subunit of NF-kB |
| RF | Rheumatoid factor |
| RNA | Ribonucleic acid |
| SEM | Standard error of mean |
| SLE | Systemic lupus erythematosus |
| TLR | Toll like receptor |
| TNF | Tumor necrosis factor |
| WBC | White blood cells |
References
- Cope, A.P. 52-Rheumatoid Arthritis. In Clinical Immunology, 5th ed.; Rich, R.R., Fleisher, T.A., Shearer, W.T., Schroeder, H.W., Frew, A.J., Weyand, C.M., Eds.; Elsevier: London, UK, 2019; pp. 705–721.e701. [Google Scholar]
- Harris, E.D. Rheumatoid arthritis. Pathophysiology and implications for therapy. N. Engl. J. Med. 1990, 322, 1277–1289. [Google Scholar] [PubMed]
- Arts, E.E.; Fransen, J.; Den Broeder, A.A.; van Riel, P.L.C.M.; Popa, C.D. Low disease activity (DAS28 ≤ 3.2) reduces the risk of first cardiovascular event in rheumatoid arthritis: A time-dependent Cox regression analysis in a large cohort study. Ann. Rheum. Dis. 2017, 76, 1693–1699. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Wang, Y.; Xu, D.; Nossent, J.; Pavlos, N.J.; Xu, J. Rheumatoid arthritis: Pathological mechanisms and modern pharmacologic therapies. Bone Res. 2018, 6, 15. [Google Scholar] [CrossRef] [PubMed]
- Gibofsky, A. Overview of epidemiology, pathophysiology, and diagnosis of rheumatoid arthritis. Am. J. Manag. Care 2012, 18, S295–S302. [Google Scholar]
- Doran Michele, F.; Pond Gregory, R.; Crowson Cynthia, S.; O′Fallon, W.M.; Gabriel Sherine, E. Trends in incidence and mortality in rheumatoid arthritis in Rochester, Minnesota, over a forty-year period. Arthritis Rheum. 2002, 46, 625–631. [Google Scholar] [CrossRef]
- Aletaha, D.; Neogi, T.; Silman, A.J.; Funovits, J.; Felson, D.T.; Bingham, C.O.; Birnbaum, N.S.; Burmester, G.R.; Bykerk, V.P.; Cohen, M.D.; et al. 2010 Rheumatoid arthritis classification criteria: An American College of Rheumatology/European League Against Rheumatism collaborative initiative. Ann. Rheum. Dis. 2010, 69, 1580. [Google Scholar] [CrossRef] [PubMed]
- Van Baarsen, L.G.M.; Wijbrandts, C.A.; Timmer, T.C.G.; van der Pouw Kraan, T.C.T.M.; Tak, P.P.; Verweij, C.L. Synovial tissue heterogeneity in rheumatoid arthritis in relation to disease activity and biomarkers in peripheral blood. Arthritis Rheum. 2010, 62, 1602–1607. [Google Scholar] [CrossRef]
- Maghazachi, A.A. Compartmentalization of human natural killer cells. Mol. Immunol. 2005, 42, 523–529. [Google Scholar] [CrossRef]
- Cooper, M.A.; Fehniger, T.A.; Caligiuri, M.A. The biology of human natural killer-cell subsets. Trends Immunol. 2001, 22, 633–640. [Google Scholar] [CrossRef]
- Shegarfi, H.; Naddafi, F.; Mirshafiey, A. Natural killer cells and their role in rheumatoid arthritis: Friend or foe? Sci. World J. 2012, 2012, 491974. [Google Scholar] [CrossRef]
- Yap, H.-Y.; Tee, S.Z.-Y.; Wong, M.M.-T.; Chow, S.-K.; Peh, S.-C.; Teow, S.-Y. Pathogenic Role of Immune Cells in Rheumatoid Arthritis: Implications in Clinical Treatment and Biomarker Development. Cells 2018, 7, 161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, B.; Khodadoust, M.S.; Liu, C.L.; Newman, A.M.; Alizadeh, A.A. Profiling Tumor Infiltrating Immune Cells with CIBERSORT. Methods Mol. Biol. 2018, 1711, 243–259. [Google Scholar] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barturen, G.; Beretta, L.; Cervera, R.; Van Vollenhoven, R.; Alarcón-Riquelme, M.E. Moving towards a molecular taxonomy of autoimmune rheumatic diseases. Nat. Rev. Rheumatol. 2018, 14, 75. [Google Scholar] [CrossRef]
- Van der Pouw Kraan, T.C.T.M.; van Gaalen, F.A.; Kasperkovitz, P.V.; Verbeet, N.L.; Smeets, T.J.M.; Kraan, M.C.; Fero, M.; Tak, P.-P.; Huizinga, T.W.J.; Pieterman, E.; et al. Rheumatoid arthritis is a heterogeneous disease: Evidence for differences in the activation of the STAT-1 pathway between rheumatoid tissues. Arthritis Rheum. 2003, 48, 2132–2145. [Google Scholar] [CrossRef]
- Yamin, R.; Berhani, O.; Peleg, H.; Aamar, S.; Stein, N.; Gamliel, M.; Hindi, I.; Scheiman-Elazary, A.; Gur, C. High percentages and activity of synovial fluid NK cells present in patients with advanced stage active Rheumatoid Arthritis. Sci. Rep. 2019, 9, 1351. [Google Scholar] [CrossRef] [Green Version]
- Tak, P.P.; Kummer, J.A.; Hack, C.E.; Daha, M.R.; Smeets, T.J.M.; Erkelens, G.W.; Meinders, A.E.; Kluin, P.M.; Breedveld, F.C. Granzyme-positive cytotoxic cells are specifically increased in early rheumatoid synovial tissue. Arthritis Rheum. 1994, 37, 1735–1743. [Google Scholar] [CrossRef]
- Dalbeth, N.; Callan, M.F.C. A subset of natural killer cells is greatly expanded within inflamed joints. Arthritis Rheum. 2002, 46, 1763–1772. [Google Scholar] [CrossRef]
- Dalbeth, N.; Gundle, R.; Davies, R.J.O.; Lee, Y.C.G.; McMichael, A.J.; Callan, M.F.C. CD56bright NK Cells Are Enriched at Inflammatory Sites and Can Engage with Monocytes in a Reciprocal Program of Activation. J. Immunol. 2004, 173, 6418. [Google Scholar] [CrossRef] [Green Version]
- Ahern, D.J.; Brennan, F.M. The role of natural killer cells in the pathogenesis of rheumatoid arthritis: Major contributors or essential homeostatic modulators? Immunol. Lett. 2011, 136, 115–121. [Google Scholar] [CrossRef]
- Gerosa, F.; Baldani-Guerra, B.; Nisii, C.; Marchesini, V.; Carra, G.; Trinchieri, G. Reciprocal activating interaction between natural killer cells and dendritic cells. J. Exp. Med. 2002, 195, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Piccioli, D.; Sbrana, S.; Melandri, E.; Valiante, N.M. Contact-dependent stimulation and inhibition of dendritic cells by natural killer cells. J. Exp. Med. 2002, 195, 335–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, A.; Filer, A.; Parsonage, G.; Kollnberger, S.; Gundle, R.; Buckley, C.D.; Bowness, P. Mediation of the proinflammatory cytokine response in rheumatoid arthritis and spondylarthritis by interactions between fibroblast-like synoviocytes and natural killer cells. Arthritis Rheum. 2008, 58, 707–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Söderström, K.; Stein, E.; Colmenero, P.; Purath, U.; Müller-Ladner, U.; Teixeira de Matos, C.; Tarner, I.H.; Robinson, W.H.; Engleman, E.G. Natural killer cells trigger osteoclastogenesis and bone destruction in arthritis. Proc. Natl. Acad. Sci. USA 2010, 107, 13028. [Google Scholar] [CrossRef] [Green Version]
- Nedvetzki, S.; Sowinski, S.; Eagle, R.A.; Harris, J.; Vély, F.D.R.; Pende, D.; Trowsdale, J.; Vivier, E.; Gordon, S.; Davis, D.M. Reciprocal regulation of human natural killer cells and macrophages associated with distinct immune synapses. Blood 2007, 109, 3776–3785. [Google Scholar] [CrossRef]
- Rabinovich, B.A.; Li, J.; Shannon, J.; Hurren, R.; Chalupny, J.; Cosman, D.; Miller, R.G. Activated, But Not Resting, T Cells Can Be Recognized and Killed by Syngeneic NK Cells. J. Immunol. 2003, 170, 3572. [Google Scholar] [CrossRef] [Green Version]
- Takayanagi, H.; Ogasawara, K.; Hida, S.; Chiba, T.; Murata, S.; Sato, K.; Takaoka, A.; Yokochi, T.; Oda, H.; Tanaka, K.; et al. T-cell-mediated regulation of osteoclastogenesis by signalling cross-talk between RANKL and IFN-γ. Nature 2000, 408, 600–605. [Google Scholar] [CrossRef]
- Chalan, P.; Bijzet, J.; Kroesen, B.-J.; Boots, A.M.H.; Brouwer, E. Altered Natural Killer Cell Subsets in Seropositive Arthralgia and Early Rheumatoid Arthritis Are Associated with Autoantibody Status. J. Rheumatol. 2016, 43, 1008. [Google Scholar] [CrossRef]
- Aggarwal, A.; Sharma, A.; Bhatnagar, A. Role of cytolytic impairment of natural killer and natural killer T-cell populations in rheumatoid arthritis. Clin. Rheumatol. 2014, 33, 1067–1078. [Google Scholar] [CrossRef]
- Conigliaro, P.; Triggianese, P.; Perricone, C.; Chimenti, M.S.; Di Muzio, G.; Ballanti, E.; Guarino, M.D.; Kroegler, B.; Gigliucci, G.; Grelli, S.; et al. Restoration of peripheral blood natural killer and B cell levels in patients affected by rheumatoid and psoriatic arthritis during etanercept treatment. Clin. Exp. Immunol. 2014, 177, 234–243. [Google Scholar] [CrossRef] [Green Version]
- Sokolove, J.; Bromberg, R.; Deane, K.D.; Lahey, L.J.; Derber, L.A.; Chandra, P.E.; Edison, J.D.; Gilliland, W.R.; Tibshirani, R.J.; Norris, J.M.; et al. Autoantibody epitope spreading in the pre-clinical phase predicts progression to rheumatoid arthritis. PLoS ONE 2012, 7, e35296. [Google Scholar] [CrossRef]
- Aramaki, T.; Ida, H.; Izumi, Y.; Fujikawa, K.; Huang, M.; Arima, K.; Tamai, M.; Kamachi, M.; Nakamura, H.; Kawakami, A.; et al. A significantly impaired natural killer cell activity due to a low activity on a per-cell basis in rheumatoid arthritis. Mod. Rheumatol. 2009, 19, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Conigliaro, P.; Scrivo, R.; Valesini, G.; Perricone, R. Emerging role for NK cells in the pathogenesis of inflammatory arthropathies. Autoimmun. Rev. 2011, 10, 577–581. [Google Scholar] [CrossRef] [PubMed]
- Saraste, M.; Irjala, H.; Airas, L. Expansion of CD56Bright natural killer cells in the peripheral blood of multiple sclerosis patients treated with interferon-beta. Neurol. Sci. 2007, 28, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Schepis, D.; Gunnarsson, I.; Eloranta, M.-L.; Lampa, J.; Jacobson, S.H.; Kärre, K.; Berg, L. Increased proportion of CD56bright natural killer cells in active and inactive systemic lupus erythematosus. Immunology 2009, 126, 140–146. [Google Scholar] [CrossRef]
- Bao, Y.; Zheng, J.; Han, C.; Jin, J.; Han, H.; Liu, Y.; Lau, Y.-L.; Tu, W.; Cao, X. Tyrosine kinase Btk is required for NK cell activation. J. Biol. Chem. 2012, 287, 23769–23778. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; Kim, T.; Jeong, D.; Kim, N.; Choi, Y. The tec family tyrosine kinase Btk Regulates RANKL-induced osteoclast maturation. J. Biol. Chem. 2008, 283, 11526–11534. [Google Scholar] [CrossRef] [Green Version]
- Shinohara, M.; Koga, T.; Okamoto, K.; Sakaguchi, S.; Arai, K.; Yasuda, H.; Takai, T.; Kodama, T.; Morio, T.; Geha, R.S.; et al. Tyrosine kinases Btk and Tec regulate osteoclast differentiation by linking RANK and ITAM signals. Cell 2008, 132, 794–806. [Google Scholar] [CrossRef] [Green Version]
- Shinners, N.P.; Carlesso, G.; Castro, I.; Hoek, K.L.; Corn, R.A.; Woodland, R.T.; Scott, M.L.; Wang, D.; Khan, W.N. Bruton’s tyrosine kinase mediates NF-κappa B activation and B cell survival by B cell-activating factor receptor of the TNF-R family. J. Immunol. 2007, 179, 3872–3880. [Google Scholar] [CrossRef] [Green Version]
- De Matos, C.T.; Berg, L.; Michaëlsson, J.; Felländer-Tsai, L.; Kärre, K.; Söderström, K. Activating and inhibitory receptors on synovial fluid natural killer cells of arthritis patients: Role of CD94/NKG2A in control of cytokine secretion. Immunology 2007, 122, 291–301. [Google Scholar] [CrossRef]
- Pamuk, O.N.; Tozkir, H.; Uyanik, M.S.; Gurkan, H.; Saritas, F.; Duymaz, J.; Donmez, S.; Yazar, M.; Pamuk, G.E. PECAM-1 gene polymorphisms and soluble PECAM-1 level in rheumatoid arthritis and systemic lupus erythematosus patients: Any link with clinical atherosclerotic events? Clin. Rheumatol. 2014, 33, 1737–1743. [Google Scholar] [CrossRef] [PubMed]
- Berman, M.E.; Xie, Y.; Muller, W.A. Roles of platelet/endothelial cell adhesion molecule-1 (PECAM-1, CD31) in natural killer cell transendothelial migration and beta 2 integrin activation. J. Immunol. 1996, 156, 1515. [Google Scholar] [PubMed]
- Szekanecz, Z.; Vegvari, A.; Szabo, Z.; Koch, A.E. Chemokines and chemokine receptors in arthritis. Front. Biosci. 2010, 2, 153–167. [Google Scholar] [CrossRef] [Green Version]
- Coelho, F.M.; Pinho, V.; Amaral, F.A.; Sachs, D.; Costa, V.V.; Rodrigues, D.H.; Vieira, A.T.; Silva, T.A.; Souza, D.G.; Bertini, R.; et al. The chemokine receptors CXCR1/CXCR2 modulate antigen-induced arthritis by regulating adhesion of neutrophils to the synovial microvasculature. Arthritis Rheum. 2008, 58, 2329–2337. [Google Scholar] [CrossRef] [PubMed]
- Sucur, A.; Jajic, Z.; Artukovic, M.; Matijasevic, M.I.; Anic, B.; Flegar, D.; Markotic, A.; Kelava, T.; Ivcevic, S.; Kovacic, N.; et al. Chemokine signals are crucial for enhanced homing and differentiation of circulating osteoclast progenitor cells. Arthritis Res. Ther. 2017, 19, 142. [Google Scholar] [CrossRef] [Green Version]
- Bahlas, S.; Damiati, L.; Dandachi, N.; Sait, H.; Alsefri, M.; Pushparaj, P.N. Rapid immunoprofiling of cytokines, chemokines and growth factors in patients with active rheumatoid arthritis using Luminex Multiple Analyte Profiling technology for precision medicine. Clin. Exp. Rheumatol. 2019, 37, 112–119. [Google Scholar] [PubMed]
- Katrib, A.; Tak, P.P.; Bertouch, J.V.; Cuello, C.; McNeil, H.P.; Smeets, T.J.M.; Kraan, M.C.; Youssef, P.P. Expression of chemokines and matrix metalloproteinases in early rheumatoid arthritis. Rheumatology 2001, 40, 988–994. [Google Scholar] [CrossRef] [Green Version]
- Koch, A.E.; Kunkel, S.L.; Harlow, L.A.; Johnson, B.; Evanoff, H.L.; Haines, G.K.; Burdick, M.D.; Pope, R.M.; Strieter, R.M. Enhanced production of monocyte chemoattractant protein-1 in rheumatoid arthritis. J. Clin. Investig. 1992, 90, 772–779. [Google Scholar] [CrossRef] [Green Version]
- Van Hooij, A.; Boeters, D.M.; Tjon Kon Fat, E.M.; van den Eeden, S.J.F.; Corstjens, P.L.A.M.; van der Helm-van Mil, A.H.M.; Geluk, A. Longitudinal IP-10 Serum Levels Are Associated with the Course of Disease Activity and Remission in Patients with Rheumatoid Arthritis. Clin. Vaccine Immunol. 2017, 24, e00060-17. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhang, Y.; Yang, X.; Han, W.; Liu, Y.; Xu, Q.; Zhao, R.; Di, C.; Song, Q.; Ma, D. Chemokine-like factor 1 is a functional ligand for CC chemokine receptor 4 (CCR4). Life Sci. 2006, 78, 614–621. [Google Scholar] [CrossRef]
- Li, T.; Zhong, J.; Chen, Y.; Qiu, X.; Zhang, T.; Ma, D.; Han, W. Expression of chemokine-like factor 1 is upregulated during T lymphocyte activation. Life Sci. 2006, 79, 519–524. [Google Scholar] [CrossRef] [PubMed]
- Flytlie, H.A.; Hvid, M.; Lindgreen, E.; Kofod-Olsen, E.; Petersen, E.L.; Jørgensen, A.; Deleuran, M.; Vestergaard, C.; Deleuran, B. Expression of MDC/CCL22 and its receptor CCR4 in rheumatoid arthritis, psoriatic arthritis and osteoarthritis. Cytokine 2010, 49, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Thompson, S.D.; Luyrink, L.K.; Graham, T.B.; Tsoras, M.; Ryan, M.; Passo, M.H.; Glass, D.N. Chemokine receptor CCR4 on CD4+ T cells in juvenile rheumatoid arthritis synovial fluid defines a subset of cells with increased IL-4:IFN-γamma mRNA ratios. J. Immunol. 2001, 166, 6899–6906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nanki, T.; Shimaoka, T.; Hayashida, K.; Taniguchi, K.; Yonehara, S.; Miyasaka, N. Pathogenic role of the CXCL16-CXCR6 pathway in rheumatoid arthritis. Arthritis Rheum. 2005, 52, 3004–3014. [Google Scholar] [CrossRef]
- Kim, C.H.; Kunkel, E.J.; Boisvert, J.; Johnston, B.; Campbell, J.J.; Genovese, M.C.; Greenberg, H.B.; Butcher, E.C. Bonzo/CXCR6 expression defines type 1-polarized T-cell subsets with extralymphoid tissue homing potential. J. Clin. Investig. 2001, 107, 595–601. [Google Scholar] [CrossRef] [Green Version]
- Ruth, J.H.; Haas, C.S.; Park, C.C.; Amin, M.A.; Martinez, R.J.; Haines, G.K., 3rd; Shahrara, S.; Campbell, P.L.; Koch, A.E. CXCL16-mediated cell recruitment to rheumatoid arthritis synovial tissue and murine lymph nodes is dependent upon the MAPK pathway. Arthritis Rheum. 2006, 54, 765–778. [Google Scholar] [CrossRef]
- Van der Voort, R.; van Lieshout, A.W.T.; Toonen, L.W.J.; Slöetjes, A.W.; van den Berg, W.B.; Figdor, C.G.; Radstake, T.R.D.J.; Adema, G.J. Elevated CXCL16 expression by synovial macrophages recruits memory T cells into rheumatoid joints. Arthritis Rheum. 2005, 52, 1381–1391. [Google Scholar] [CrossRef]
- Adib-Conquy, M.; Scott-Algara, D.; Cavaillon, J.-M.; Souza-Fonseca-Guimaraes, F. TLR-mediated activation of NK cells and their role in bacterial/viral immune responses in mammals. Immunol. Cell Biol. 2014, 92, 256–262. [Google Scholar] [CrossRef]
- Huang, Q.-Q.; Pope, R.M. The role of toll-like receptors in rheumatoid arthritis. Curr. Rheumatol. Rep. 2009, 11, 357–364. [Google Scholar] [CrossRef] [Green Version]
- Ospelt, C.; Brentano, F.; Rengel, Y.; Stanczyk, J.; Kolling, C.; Tak, P.P.; Gay, R.E.; Gay, S.; Kyburz, D. Overexpression of toll-like receptors 3 and 4 in synovial tissue from patients with early rheumatoid arthritis: Toll-like receptor expression in early and longstanding arthritis. Arthritis Rheum. 2008, 58, 3684–3692. [Google Scholar] [CrossRef] [Green Version]
- Thwaites, R.; Chamberlain, G.; Sacre, S. Emerging Role of Endosomal Toll-Like Receptors in Rheumatoid Arthritis. Front. Immunol. 2014, 5, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Cao, R.; Ying, H.; Du, J.; Chen, S.; Wang, N.; Shen, B. Increased Expression of TLR10 in B Cell Subsets Correlates with Disease Activity in Rheumatoid Arthritis. Mediat. Inflamm. 2018, 2018, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eastgate, J.A.; Symons, J.A.; Wood, N.C.; Grinlinton, F.M.; di Giovine, F.S.; Duff, G.W. Correlation of plasma interleukin 1 levels with disease activity in rheumatoid arthritis. Lancet 1988, 2, 706–709. [Google Scholar] [CrossRef]
- Kageyama, Y.; Torikai, E.; Nagano, A. Anti-tumor necrosis factor-alpha antibody treatment reduces serum CXCL16 levels in patients with rheumatoid arthritis. Rheumatol. Int. 2007, 27, 467–472. [Google Scholar] [CrossRef] [PubMed]
- Li, C.-H.; Xu, L.-L.; Zhao, J.-X.; Sun, L.; Yao, Z.-Q.; Deng, X.-L.; Liu, R.; Yang, L.; Xing, R.; Liu, X.-Y. CXCL16 upregulates RANKL expression in rheumatoid arthritis synovial fibroblasts through the JAK2/STAT3 and p38/MAPK signaling pathway. Inflamm. Res. 2016, 65, 193–202. [Google Scholar] [CrossRef]
- Elemam, N.M.; Hachim, M.Y.; Hannawi, S.; Maghazachi, A.A. Natural Killer Cells Gene Expression Can Differentiate Rheumatoid Arthritis Patients from Healthy Controls [abstract]. In Proceedings of the ACR/ARP Annual Meeting, Supplement of Arthritis & Rheumatology, Atlanta, GA, USA, 8–13 November 2019. [Google Scholar]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elemam, N.M.; Hachim, M.Y.; Hannawi, S.; Maghazachi, A.A. Differentially Expressed Genes of Natural Killer Cells Can Distinguish Rheumatoid Arthritis Patients from Healthy Controls. Genes 2020, 11, 492. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11050492
Elemam NM, Hachim MY, Hannawi S, Maghazachi AA. Differentially Expressed Genes of Natural Killer Cells Can Distinguish Rheumatoid Arthritis Patients from Healthy Controls. Genes. 2020; 11(5):492. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11050492
Chicago/Turabian StyleElemam, Noha Mousaad, Mahmood Yaseen Hachim, Suad Hannawi, and Azzam A. Maghazachi. 2020. "Differentially Expressed Genes of Natural Killer Cells Can Distinguish Rheumatoid Arthritis Patients from Healthy Controls" Genes 11, no. 5: 492. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11050492