Sex Chromosomes and Internal Telomeric Sequences in Dormitator latifrons (Richardson 1844) (Eleotridae: Eleotrinae): An Insight into Their Origin in the Genus



Abstract
:1. Introduction
2. Materials and Methods
2.1. Specimens Collection and Identification
2.2. Cytogenetic Procedures
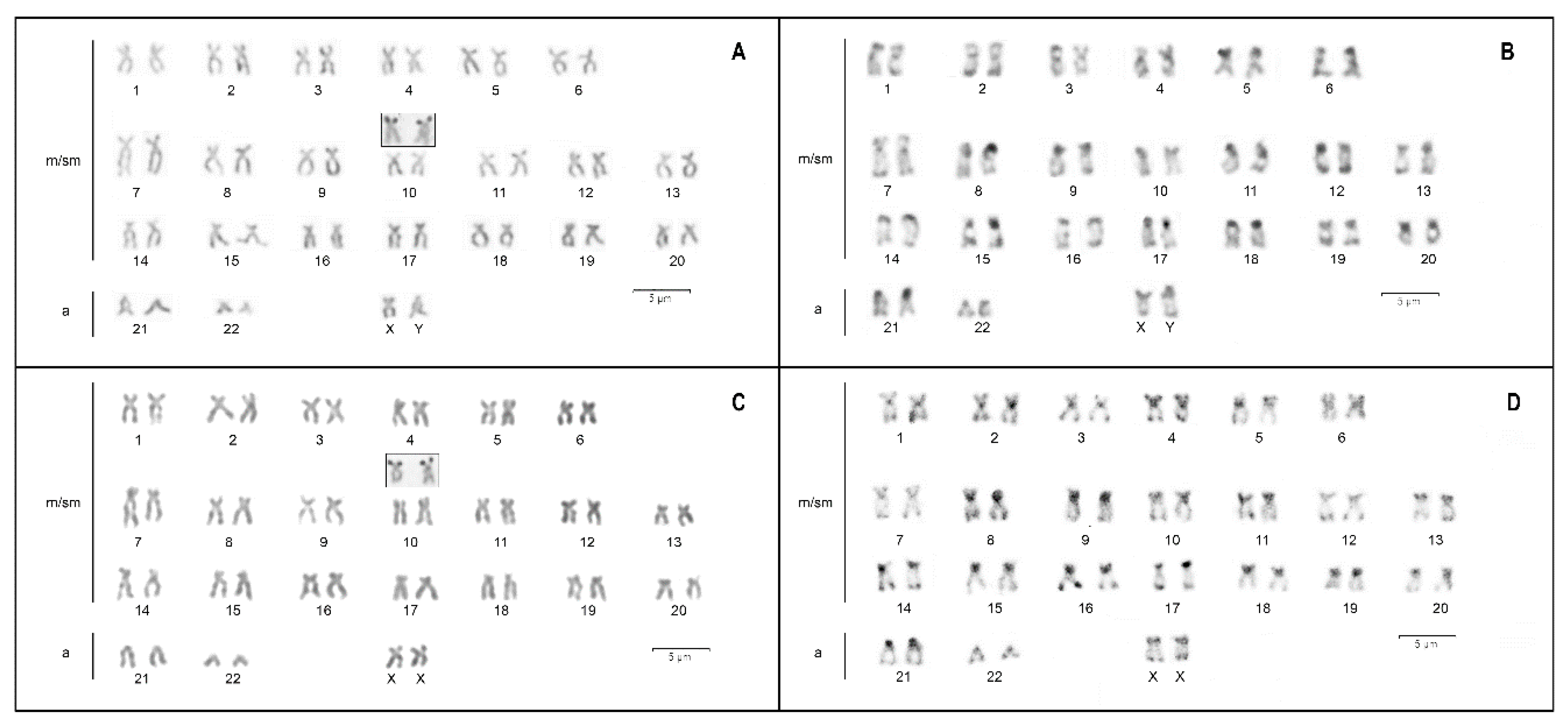
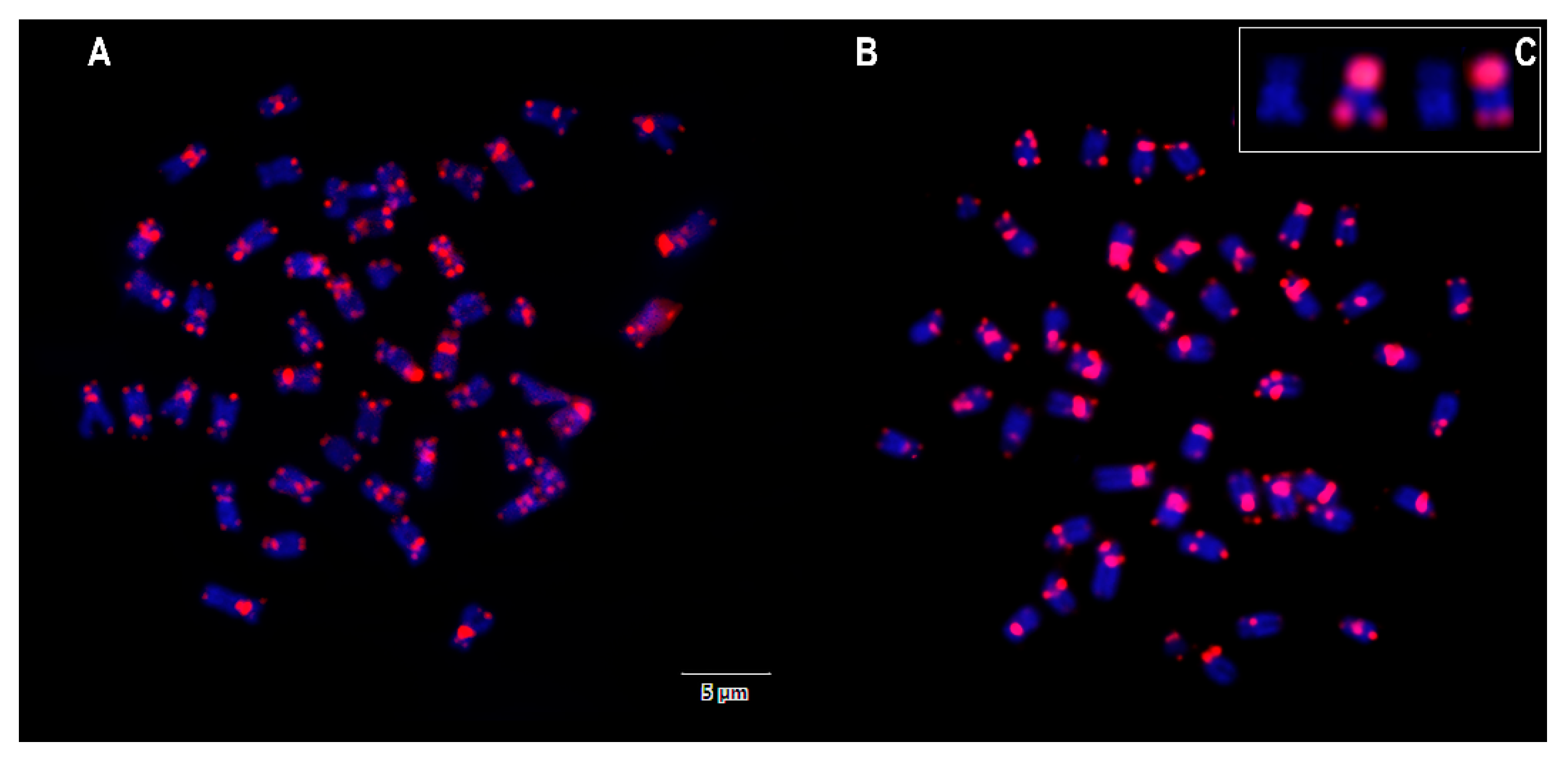
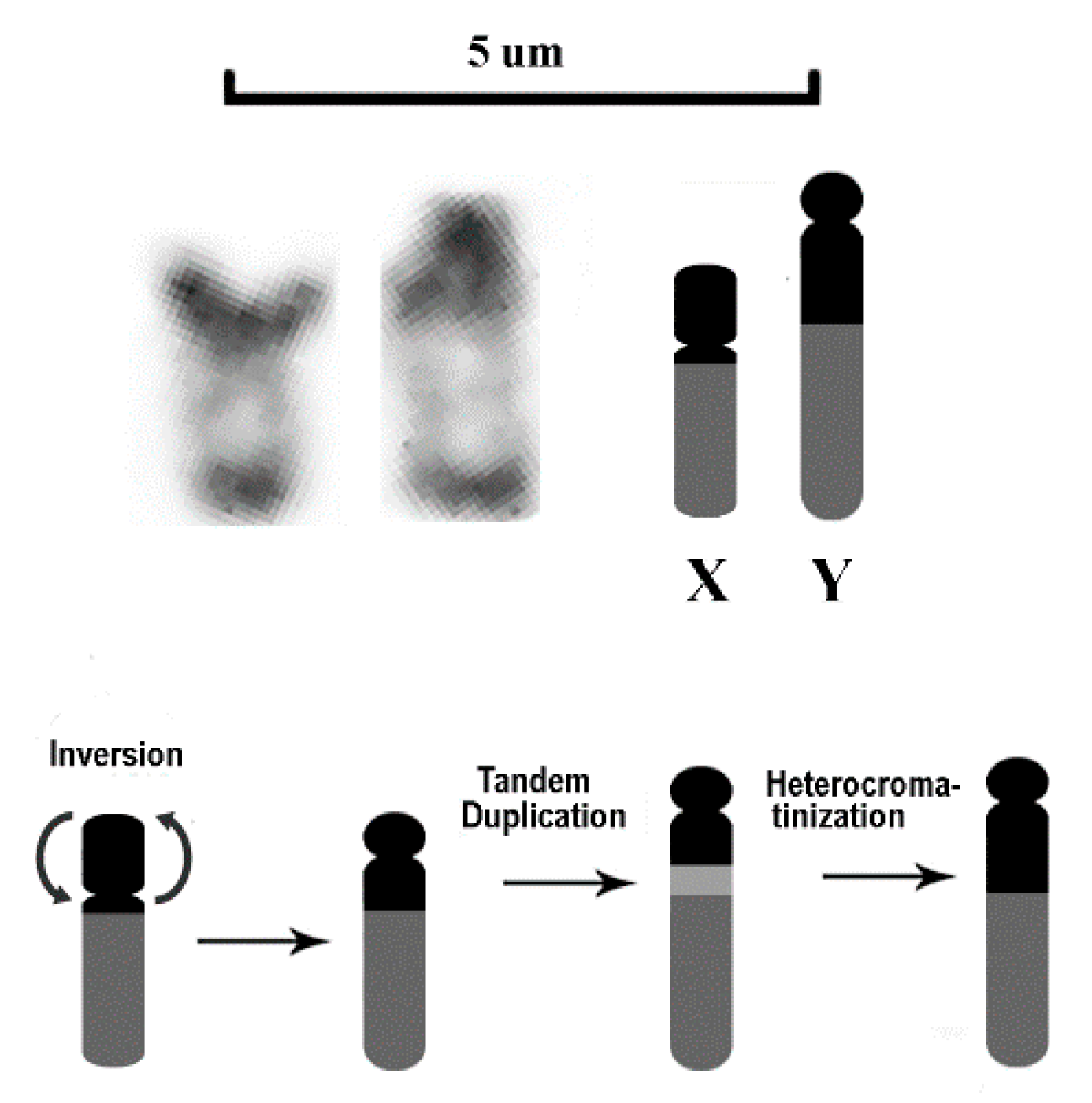
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Gleick, P.H. Water resources. In Encyclopedia of Climate and Weather; Schneider, S.H., Ed.; Oxford University Press: New York, NY, USA, 1996; Volume 2, pp. 817–823. [Google Scholar]
- Nelson, J.S.; Grande, T.C.; Wilson, M.V.H. Fishes of the World, 5th ed.; John Wiley & Sons: Hoboken, NJ, USA, 2016. [Google Scholar]
- Lundberg, J.G.; Kottelat, M.; Smith, G.R.; Stiassny, M.L.J.; Gill, A.C. So many fishes, so little time: An overview of recent ichthyological discovery in continental waters. Ann. Mo. Bot. Gard. 2000, 87, 26–62. [Google Scholar] [CrossRef]
- Toussaint, A.; Charpin, N.; Brosse, S.; Villéger, S. Global functional diversity of freshwater fish is concentrated in the Neotropics while functional vulnerability is widespread. Sci. Rep. 2016, 6, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reis, R.E.; Albert, J.S.; Di Dario, F.; Mincarone, M.M.; Petry, P.; Rocha, L.A. Fish biodiversity and conservation in South America. J. Fish Biol. 2016, 89, 12–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hilsdorf, A.W.S.; Hallerman, E.M. Genetic Resources of Neotropical Fishes; Springer: Cham, Germany, 2017. [Google Scholar]
- Nirchio, M.; Gaviria, J.I.; Siccha-Ramirez, Z.R.; Oliveira, C.; Foresti, F.; Milana, V.; Rossi, A.R. Chromosomal polymorphism and molecular variability in the pearly razorfish Xyrichtys novacula (Labriformes, Labridae): Taxonomic and biogeographic implications. Genetica 2019, 147, 47–56. [Google Scholar] [CrossRef] [PubMed]
- de Cioffi, B.M.; Wagner, F.M.; Ferreira, A.R.; Bertollo, L.A.C. Chromosomes as tools for discovering biodiversity–the case of erythrinidae fish family. Recent Trends Cytogenet. Stud. Methodol. Appl. 2012. [Google Scholar] [CrossRef] [Green Version]
- Sember, A.; de Oliveira, E.A.; Ráb, P.; Bertollo, L.A.C.; de Freitas, N.L.; Viana, P.F.; Yano, C.F.; Hatanaka, T.; Marinho, M.M.F.; de Moraes, R.L.R.; et al. Centric Fusions behind the karyotype evolution of neotropical Nannostomus Pencilfishes (Characiforme, Lebiasinidae): First insights from a molecular cytogenetic perspective. Genes 2020, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fricke, R.; Eschmeyer, W.N.; Fong, J.D. Species by Family/Subfamily. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/SpeciesByFamily.asp (accessed on 15 May 2020).
- Froese, R.; Pauly, D. Eleotrinae Bonaparte. 1835. Available online: http://www.marinespecies.org/aphia.php?p=taxdetails&id=267092 (accessed on 15 May 2020).
- Patzner, R.; Van Tassell, J.L.; Kovacic, M.; Kapoor, B.G. The Biology of Gobies; CRC Press: Boca Raton, FL, USA, 2011. [Google Scholar]
- Galván-Quesada, S.; Doadrio, I.; Alda, F.; Perdices, A.; Reina, R.G.; García Varela, M.; Hernández, N.; Campos Mendoza, A.; Bermingham, E.; Domínguez-Domínguez, O. Molecular phylogeny and biogeography of the amphidromous fish genus Dormitator Gill 1861 (Teleostei: Eleotridae). PLoS ONE 2016, 11, e0153538. [Google Scholar] [CrossRef] [Green Version]
- Van Tassell, J. Dormitator Latifrons. Available online: https://0-dx-doi-org.brum.beds.ac.uk/10.2305/IUCN.UK.2010-3.RLTS.T183257A8081686.en (accessed on 10 April 2020).
- Massay, S.; Mosquera, R. Presence of Chame Dormitator latifrons (Richardson, 1844) (Pisces: Eleotridae) in the Galapagos Islands, Ecuador. J. Fish Biol. 1992, 40, 815–816. [Google Scholar] [CrossRef]
- Larumbe, E. Algunos aspectos biológicos de los popoyotes (Dormitator latifrons) en cautiverio. Panor. Acuícola 2002, 24–25. Available online: https://fis.com/panoramacuicola/noticias/noticia%203.htm (accessed on 15 May 2020).
- Freire-Lascano, C.A. Experiencias en el manejo del Chame (Dormitator latifrons) en la Cuenca del Río Guayas, Ecuador. Rev. Electrónica Ing. Prod. Acuícola 2013, 7, 1–13. [Google Scholar]
- de Rodríguez-Montes Oca, G.A.; Medina-Hernández, E.A.; Velázquez-Sandoval, J.; López-López, V.V.; Román-Reyes, J.C.; Dabrowski, K.; Haws, M.C. Producción de larvas de Chame (Dormitator latifrons, Pisces: Eleotridae) usando GnRHa and LHRHa. Rev. Colomb. Cienc. Pecu. 2012, 25, 422–429. [Google Scholar]
- Torell, E.; Tobey, J. Enterprise Strategies for Coastal and Marine Conservation: A Review of Best Practices and Lessons Learned; Coastal Resources Center, University of Rhode Island: Narragansett, RI, USA, 2012. [Google Scholar]
- López-Huerta, J.M.; Vega-Villasante, F.; Viana, M.T.; Carrillo-Farnés, O.; Badillo-Zapata, D. First report of nutritional quality of the native fish Dormitator latifrons (Richardson, 1844) (Perciformes: Eleotridae). Lat. Am. J. Aquat. Res. 2018, 46, 849–854. [Google Scholar] [CrossRef]
- Jácome, J.; Quezada Abad, C.; Sánchez Romero, O.; Pérez, J.E.; Nirchio, M. Tilapia en Ecuador: Paradoja entre la producción acuícola y la protección de la biodiversidad ecuatoriana. Rev. Peru. Biol. 2019, 26, 543–550. [Google Scholar] [CrossRef] [Green Version]
- Fujiwara, A. Recent advancement in fish cytogenetics. J. Anim. Genet. 2005, 32, 113–122. [Google Scholar] [CrossRef]
- Gorshkova, G. Practical use of cytogenetics in fish biology and aquaculture. Isr. J. Aquac. Bamidgeh 2006, 58, 280–285. [Google Scholar]
- Ocalewicz, K.; Jankun, M.; Luczynski, M. Cytogenetic characteristics of interspecific hybrids and chromosome set manipulated finfish. In Fish Cytogenetics; Pisano, E., Ozouf-Costaz, C., Foresti, F., Kapoor, B.G., Eds.; Science Publishers: Enfield, NH, USA, 2007; pp. 289–332. [Google Scholar]
- Uribe-Alcocer, M.; Espinoza, J.A.; Padilla, A.T.; Pérez, A.C. Los cromosomas de Dormitator latifrons (Pisces: Gobiidae). An. Inst. Cienc. Mar Limnol. Univ. Nac. Auton. Mex. 1982, 10, 23–30. [Google Scholar]
- Uribe-Alcocer, M.; Ramirez-Escamilla, A. Comparación citogenética entre las especies del genero Dormitator (Pisces, Gobiidae). An. Inst. Cienc. Mar Limnol. Univ. Nac. Auton. Mex. 1989, 16, 75–80. [Google Scholar]
- Maldonado-Monroy, M.C.; Uribe-Alcocer, M. Karyotypical studies on Dormitator maculatus bloch and Gobiomorus dormitor Lacépède (Gobiidae: Perciformes). Cytologia 1985, 50, 663–669. [Google Scholar] [CrossRef] [Green Version]
- Molina, W.F. Intraspecific karyotypical diversity in brackish water fishes of the Eleotridae family (Pisces, Perciformes). Cytologia 2005, 70, 39–45. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, C.; de Toledo, L.F.A. Evidence of an XX/XY sex chromosome system in the fish Dormitator maculatus (Teleostei, Eleotrididae). Genet. Mol. Biol. 2006, 29, 653–655. [Google Scholar] [CrossRef] [Green Version]
- Arai, R. Fish Karyotypes: A Check List; Springer: Tokyo, Japan, 2011. [Google Scholar]
- Uribe-Alcocer, M.; Díaz-Jaimes, P. Chromosome complements of Gobionellus microdon and Eleotris picta collected in Mexico. J. Fish Biol. 1996, 48, 796–798. [Google Scholar] [CrossRef]
- Rocon-Stange, E.A. Caracterização cromossômica de Eleotris pisonis (Perciformes, Eleotrididae). In Proceedings of the IV Simpósio de Citogenética Evolutiva e Aplicada de Peixes Neotropicais, Rio de Janeiro, Brazil, 22–24 September 1992; p. 45. [Google Scholar]
- Masagca, J.T.; Sumantadinata, K. Chromosomal characters of the Indonesian sand goby, Oxyeleotris marmorata Blkr. 1874 (Eleotridae). Biotropia Southeast Asian J. Trop. Biol. 1994, 41–46. [Google Scholar] [CrossRef]
- Pinthong, K.; Supiwong, W.; Tanomtong, A.; Hongsachart, P.; Phaengphairee, P.; Chantarangsee, M. A first karyological analysis of the sand goby, Oxyeleotris marmoratus (Teleostei, Eleotridae) in Thailand by Ag-NOR Staining Technique. Cytologia 2014, 79, 127–132. [Google Scholar] [CrossRef] [Green Version]
- Charlesworth, B. Sex determination: Primitive Y chromosomes in fish. Curr. Biol. 2004, 14, R745–R747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, K.; Takehana, Y.; Naruse, K.; Hamaguchi, S.; Sakaizumi, M. Evidence for different origins of sex chromosomes in closely related Oryzias fishes: Substitution of the master sex-determining gene. Genetics 2007, 177, 2075–2081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gornung, E. Twenty years of physical mapping of major ribosomal RNA genes across the Teleosts: A review of research. Cytogenet. Genome Res. 2013, 141, 90–102. [Google Scholar] [CrossRef]
- Symonová, R.; Howell, W.M. Vertebrate genome evolution in the light of fish Cytogenomics and rDNAomics. Genes 2018, 9, 96. [Google Scholar] [CrossRef] [Green Version]
- Cioffi, M.B.; Martins, C.; Bertollo, L.A.C. Chromosome spreading of associated transposable elements and ribosomal DNA in the fish Erythrinus erythrinus. Implications for genome change and karyoevolution in fish. BMC Evol. Biol. 2010, 10, 271. [Google Scholar] [CrossRef] [Green Version]
- Jimenez-Prado, P.; Agruirre, W.; Moncayo, E.; Amaya, R.; Salazar, N.; Iván, F.; Rivera, J. Guía de Peces Para Aguas Continentales en la Vertiente Occidental del Ecuador; Pontificia Universidad Católica del Ecuador Sede Esmeraldas (PUCESE): Quito, Ecuador, 2015. [Google Scholar]
- Lozano, R.; Ruiz Rejon, C.; Ruiz Rejon, M. A method for increasing the number of mitoses available for cytogenetic analysis in rainbow trout. Stain Technol. 1988, 63, 335–338. [Google Scholar] [CrossRef]
- Leary, S.L.; Underwood, W.; Anthony, R.; Gwaltney-Brant, S.; Poison, A.; Meyer, R. AVMA Guidelines for the Euthanasia of Animals, 2013 ed.; American Veterinary Medical Association: Schaumburg, IL, USA, 2013. [Google Scholar]
- Nirchio, M.; Oliveira, C. Citogenética de Peces; Universidad de Oriente: Cumaná, Venezuela, 2006. [Google Scholar]
- Howell, W.M.; Black, D.A. Controlled silver-staining of nucleolus organizer regions with a protective colloidal developer: A 1-step method. Experientia 1980, 36, 1014–1015. [Google Scholar] [CrossRef]
- Sumner, A.T. A simple technique for demonstrating centromeric heterochromatin. Exp. Cell Res. 1972, 75, 304–306. [Google Scholar] [CrossRef]
- Pinkel, D.; Straume, T.; Gray, J.W. Cytogenetic analysis using quantitative, high-sensitivity, fluorescence hybridization. Proc. Natl. Acad. Sci. USA 1986, 83, 2934–2938. [Google Scholar] [CrossRef] [Green Version]
- Levan, A.; Fredga, K.; Sandberg, A.A. Nomenclature for centromeric position on chromosomes. Hereditas 1964, 52, 201–220. [Google Scholar] [CrossRef]
- Mank, J.E.; Avise, J.C. Phylogenetic conservation of chromosome numbers in Actinopterygiian fishes. Genetica 2006, 127, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Kottler, V.A.; Schartl, M. The colorful sex chromosomes of Teleost fish. Genes 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Devlin, R.H.; Nagahama, Y. Sex determination and sex differentiation in fish: An overview of genetic, physiological, and environmental influences. Aquaculture 2002, 208, 191–364. [Google Scholar] [CrossRef]
- Oliveira, C.; Almeida-Toledo, L.F.; Foresti, F. Karyotypic evolution in neotropical fishes. In Fish Cytogenetics; Pisano, E., Ozouf-Costaz, C., Foresti, F., Kapoor, B.G., Eds.; Science Publishers: Enfield, NH, USA, 2007; pp. 111–164. [Google Scholar]
- Cioffi, M.B.; Camacho, J.P.M.; Bertollo, L.A.C. Repetitive DNAs and differentiation of sex chromosomes in neotropical fishes. Cytogenet. Genome Res. 2011, 132, 188–194. [Google Scholar] [CrossRef]
- de Cioffi, M.B.; Yano, C.F.; Sember, A.; Bertollo, L.A.C. Chromosomal evolution in lower Vertebrates: Sex chromosomes in Neotropical fishes. Genes 2017, 8. [Google Scholar] [CrossRef]
- Kitano, J.; Peichel, C.L. Turnover of sex chromosomes and speciation in fishes. Environ. Biol. Fishes 2012, 94, 549–558. [Google Scholar] [CrossRef] [Green Version]
- Abbott, J.K.; Nordén, A.K.; Hansson, B. Sex chromosome evolution: Historical insights and future perspectives. Proc. Biol. Sci. 2017, 284. [Google Scholar] [CrossRef] [Green Version]
- Schultheis, C.; Böhne, A.; Schartl, M.; Volff, J.N.; Galiana-Arnoux, D. Sex determination diversity and sex chromosome evolution in poeciliid fish. Sex Dev. 2009, 3, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Chalopin, D.; Volff, J.-N.; Galiana, D.; Anderson, J.L.; Schartl, M. Transposable elements and early evolution of sex chromosomes in fish. Chromosome Res. 2015, 23, 545–560. [Google Scholar] [CrossRef] [PubMed]
- Ezaz, T.; Deakin, J.E. Repetitive sequence and sex chromosome evolution in Vertebrates. Adv. Evol. Biol. 2014, 2014. [Google Scholar] [CrossRef] [Green Version]
- Wright, A.E.; Dean, R.; Zimmer, F.; Mank, J.E. How to make a sex chromosome. Nat. Commun. 2016, 7, 12087. [Google Scholar] [CrossRef]
- Crown, K.N.; Miller, D.E.; Sekelsky, J.; Hawley, R.S. Local inversion heterozygosity alters recombination throughout the genome. Curr. Biol. 2018, 28, 2984–2990. [Google Scholar] [CrossRef] [Green Version]
- Moran, P.; Martinez, J.L.; Garcia-Vazquez, E.; Pendas, A.M. Sex chromosome linkage of 5S rDNA in rainbow trout (Oncorhynchus mykiss). Cytogenet. Cell Genet. 1996, 75, 145–150. [Google Scholar] [CrossRef]
- Scacchetti, P.C.; Utsunomia, R.; Pansonato-Alves, J.C.; Vicari, M.R.; Artoni, R.F.; Oliveira, C.; Foresti, F. Chromosomal mapping of repetitive DNAs in Characidium (Teleostei, Characiformes): Genomic organization and diversification of ZW sex chromosomes. Cytogenet. Genome Res. 2015, 146, 136–143. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, F.A.; Feldberg, E.; Carvalho, N.D.M.; Hernandez Rangel, S.M.; Schneider, C.H.; Carvalho-Zilse, G.A.; Da Silva, V.F.; Gross, M.C. Effects of environmental pollution on the rDNAomics of Amazonian fish. Environ. Pollut. 2019, 252, 180–187. [Google Scholar] [CrossRef]
- Iida, T.; Kobayashi, T. How do cells count multi-copy genes? “Musical Chair” model for preserving the number of rDNA copies. Curr. Genet. 2019, 65, 883–885. [Google Scholar] [CrossRef]
- Bolzán, A.D. Interstitial telomeric sequences in vertebrate chromosomes: Origin, function, instability and evolution. Mutat. Res. Mutat. Res. 2017, 773, 51–65. [Google Scholar] [CrossRef]
- Ocalewicz, K.; Furgala-Selezniow, G.; Szmyt, M.; Lisboa, R.; Kucinski, M.; Lejk, A.M.; Jankun, M. Pericentromeric location of the telomeric DNA sequences on the European grayling chromosomes. Genetica 2013, 141, 409–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandrioli, M.; Bizzaro, D.; Manicardi, G.C.; Gionghi, D.; Bassoli, L.; Bianchi, U. Cytogenetic and molecular characterization of a highly repeated DNA sequence in the peach potato aphid Myzus persicae. Chromosoma 1999, 108, 436–442. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Herrera, A.; Nergadze, S.G.; Santagostino, M.; Giulotto, E. Telomeric repeats far from the ends: Mechanisms of origin and role in evolution. Cytogenet. Genome Res. 2008, 122, 219–228. [Google Scholar] [CrossRef]
- Salvadori, S.; Deiana, A.; Elisabetta, C.; Floridia, G.; Rossi, E.; Zuffardi, O. Colocalization of (TTAGGG)n telomeric sequences and ribosomal genes in Atlantic eels. Chromosome Res. 1995, 3, 54–58. [Google Scholar] [CrossRef]
- Gornung, E.; Mannarelli, M.E.; Rossi, A.R.; Sola, L. Chromosomal evolution in Mugilidae (Pisces, Mugiliformes): FISH mapping of the (TTAGGG) telomeric repeat in the six Mediterranean mullets. Hereditas 2004, 140, 158–159. [Google Scholar] [CrossRef]
- Rossi, A.R.; Gornung, E.; Sola, L.; Nirchio, M. Comparative molecular cytogenetic analysis of two congeneric species, Mugil curema and M. liza (Pisces, Mugiliformes), characterized by significant karyotype diversity. Genetica 2005, 125, 27–32. [Google Scholar] [CrossRef]
- Libertini, A.; Vitturi, R.; Lannino, A.; Maone, M.C.; Franzoi, P.; Riccato, F.; Colomba, S. FISH mapping of 18S rDNA and (TTAGGG)n sequences in two pipefish species (Gasteroisteiformes: Syngnathidae). J. Genet. 2006, 85, 153–156. [Google Scholar] [CrossRef] [PubMed]
- Aksenova, A.Y.; Mirkin, S.M. At the beginning of the end and in the middle of the beginning: Structure and maintenance of telomeric DNA repeats and interstitial telomeric sequences. Genes 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ocalewicz, K. Telomeres in fishes. Cytogenet. Genome Res. 2013, 141, 114–125. [Google Scholar] [CrossRef]
- Phillips, R.; Ráb, P. Chromosome evolution in the Salmonidae (Pisces): An update. Biol. Rev. Camb. Philos. Soc. 2001, 76, 1–25. [Google Scholar] [CrossRef]
- Slijepcevic, P. Telomeres and mechanisms of Robertsonian fusion. Chromosoma 1998, 107, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.G. Manipulation of chromosomes in fish: Review of various techniques and their implications in aquaculture. Bangladesh J. Fish. Res. 1998, 2, 99–108. [Google Scholar]
- Arai, K.; Fujimoto, T. Chromosome manipulation techniques and applications to aquaculture. In Sex Control in Aquaculture; Wang, H.-P., Piferrer, F., Chen, S.-H., Shen, Z.-G., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2018; pp. 137–162. [Google Scholar] [CrossRef]
- Zheng, S.; Wang, X.; Zhang, S.; Long, J.; Tao, W.; Li, M.; Wang, D. Screening and characterization of sex-linked DNA markers and marker-assisted selection in the Southern catfish (Silurus meridionalis). Aquaculture 2020, 517, 734783. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | 2n | Karyotype Composition | FN | Sampling Area | References |
|---|---|---|---|---|---|
| Eleotrinae | |||||
| D. latifrons | 46 | 44m/sm + 2st/a | 90 | Mexico | [25] |
| D. latifrons | 46 | 44m/sm + 2st/a | 90 | Mexico | [26] |
| D. latifrons | 46 | 42m/sm + 4a (♀) 41m/sm + 5a (♂) | 88/87 | Ecuador | This paper |
| D. maculatus | 46 | 44m/sm + 2st/a | 90 | Mexico | [26] |
| D. maculatus | 46 | 34m/sm + 12st/a | 80 | Brazil | [27] |
| D. maculatus | 46 | 40m/sm + 6st/a | 86 | Brazil | [28] |
| D. maculatus | 46 | 14m + 28sm + 2st + 2a (♀); 13m + 28sm + 3st + 2a (♂) | 90 | Brazil | [29] |
| Eleotris acanthopoma | 46 | 46st/a | 46 | Japan | [30] |
| Eleotris oxycephala | 46 | 46a | 46 | China | [30] |
| E. oxycephala | 46 | 46a | 46 | China | [30] |
| Eleotris picta | 52 | 52a | 52 | Mexico | [31] |
| E. pisonis | 46 | 2m/sm + 42st/a | 46 | Mexico | [31] |
| E. pisonis | 46 | 46a | 46 | Unknown | [32] |
| E. pisonis | 46 | 46a | 46 | Brazil | [28] |
| Gobiomorus dormitor | 48 | 6m/sm + 42a | 54 | Mexico | [27] |
| Hypseleotris cyprinoides | 48 | 48a | 48 | Japan | [30] |
| Mogurnda mogurnda | 46 | 6sm + 40st/a | 52 | Australia | [30] |
| Butinae | |||||
| Ophiocara porocephala | 48 | 48a | 48 | Thailand | [30] |
| Oxyeleotris marmorata | 46 | 2m + 2sm + 42a | 50 | Thailand | [30] |
| O. marmorata | 46 | 4sm + 42a | 50 | Indonesia | [33] |
| O. marmorata | 46 | 2m + 2sm+ 42a | 50 | Thailand | [34] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paim, F.G.; Nirchio, M.; Oliveira, C.; Rossi, A.R. Sex Chromosomes and Internal Telomeric Sequences in Dormitator latifrons (Richardson 1844) (Eleotridae: Eleotrinae): An Insight into Their Origin in the Genus. Genes 2020, 11, 659. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11060659
Paim FG, Nirchio M, Oliveira C, Rossi AR. Sex Chromosomes and Internal Telomeric Sequences in Dormitator latifrons (Richardson 1844) (Eleotridae: Eleotrinae): An Insight into Their Origin in the Genus. Genes. 2020; 11(6):659. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11060659
Chicago/Turabian StylePaim, Fabilene Gomes, Mauro Nirchio, Claudio Oliveira, and Anna Rita Rossi. 2020. "Sex Chromosomes and Internal Telomeric Sequences in Dormitator latifrons (Richardson 1844) (Eleotridae: Eleotrinae): An Insight into Their Origin in the Genus" Genes 11, no. 6: 659. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11060659