Evaluation of the CRISPR/Cas9 Genetic Constructs in Efficient Disruption of Porcine Genes for Xenotransplantation Purposes Along with an Assessment of the Off-Target Mutation Formation

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Selection of Short Oligonucleotides
2.2. Preparation of Genetic Constructions in the CRISPR/Cas9 System
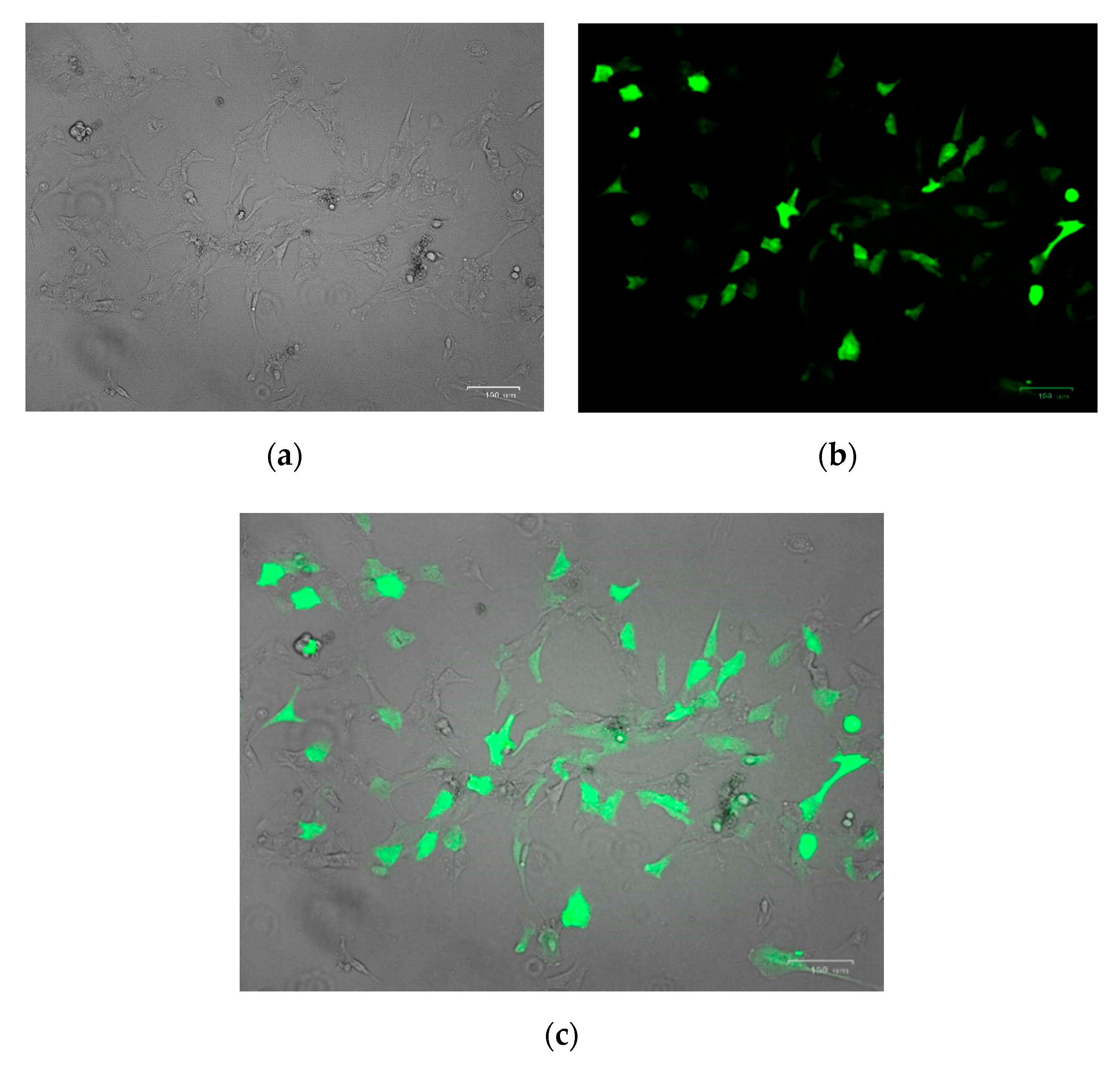
2.3. Isolation and Culture of Porcine Primary Kidney Cells In Vitro
2.4. Nucleofection
2.5. Antibiotic Selection
2.6. Analysis of Introduced Genetic Modifications
2.7. Sequencing Analysis Using the TIDE Tool
3. Results
3.1. Selection of the Potential Modifications Location in Porcine Genome
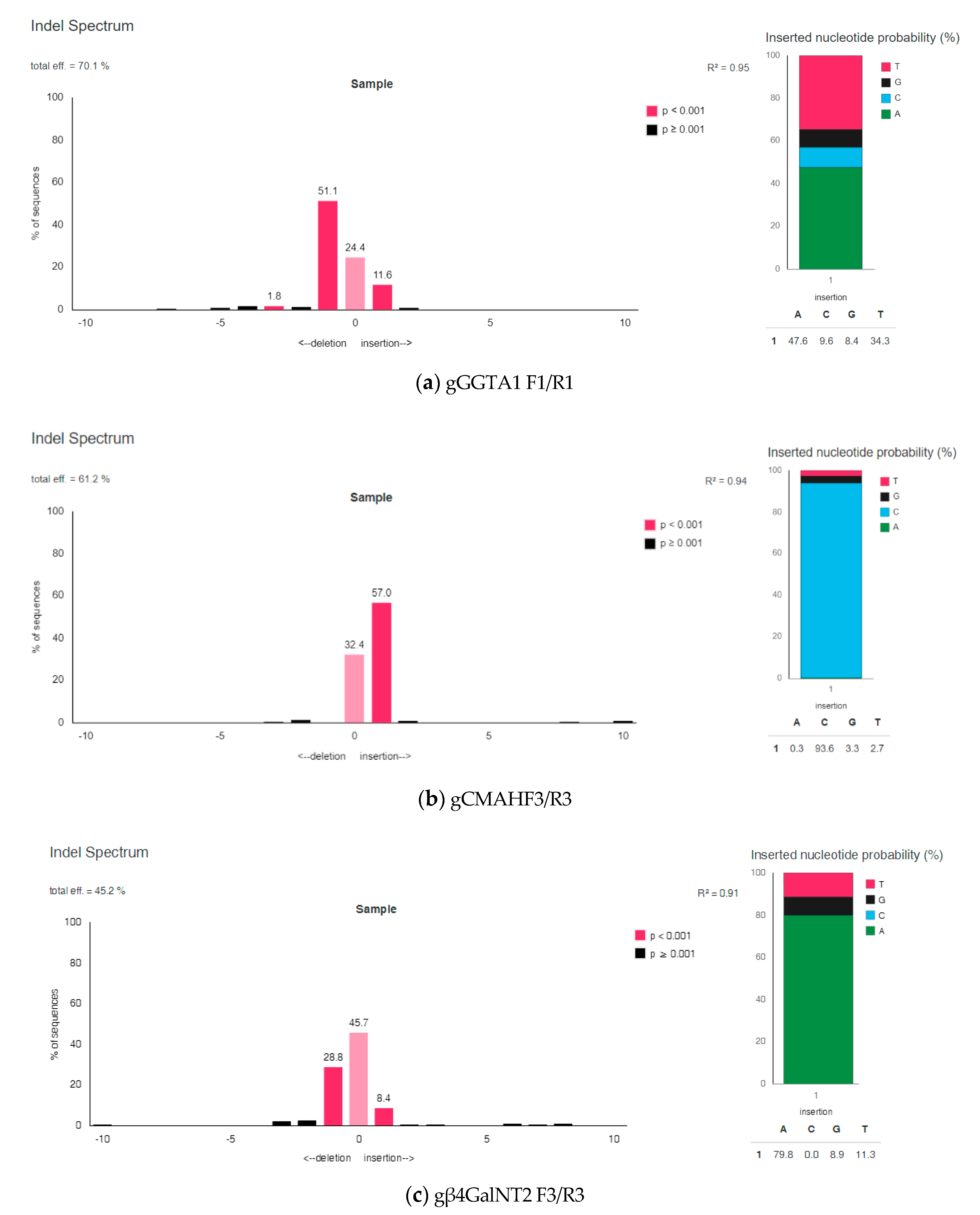
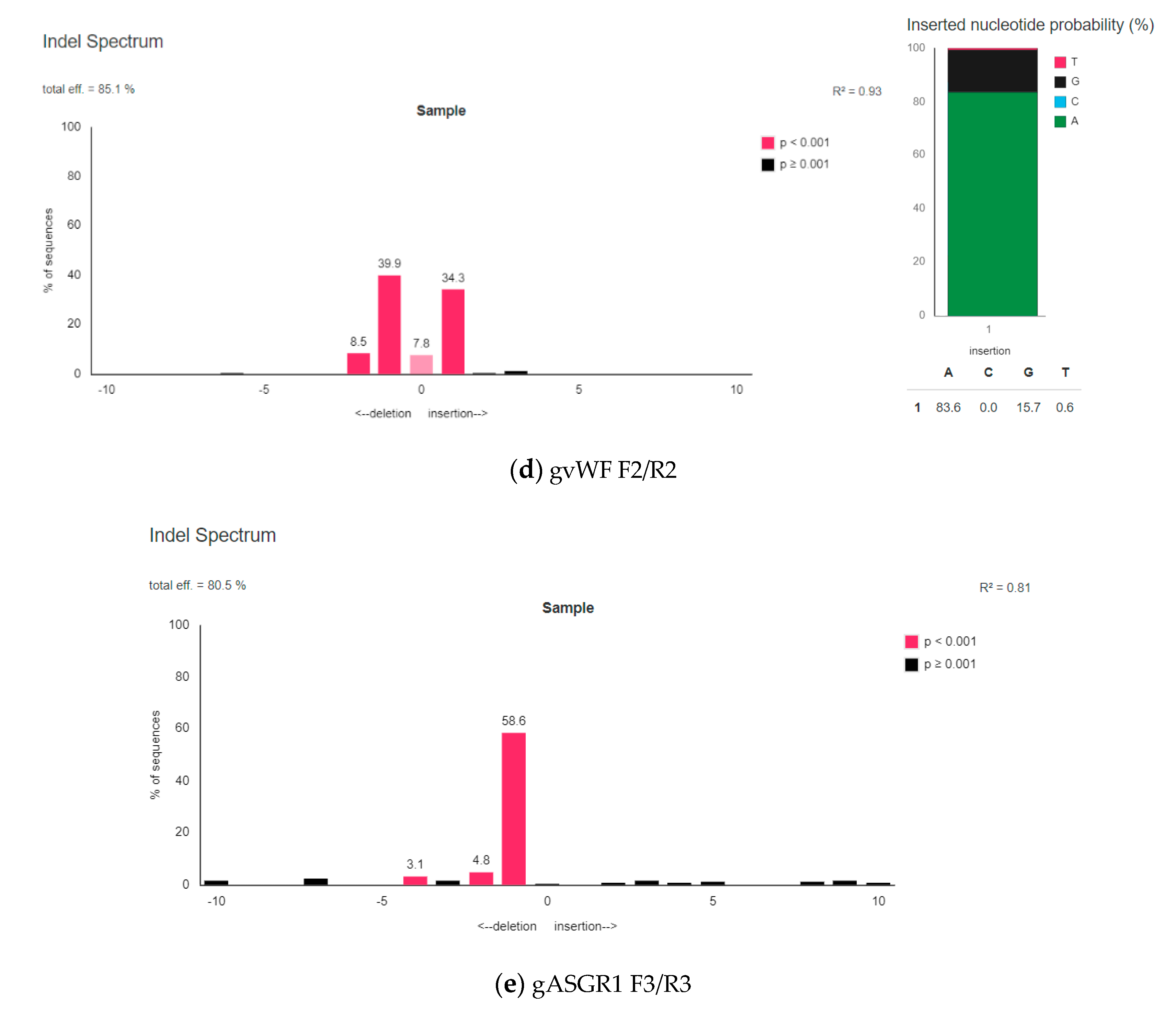
3.2. Analysis of On-Target Modification Sites
3.2.1. Nucleofection Efficiency
3.2.2. The Efficiency of Introducing Modifications within the Examined Porcine Genes Using Genetic Constructions Selected as the Best
3.2.3. Comparison of Bioinformatically Predicted DNA Diruption Efficiency Results with Those Obtained in the In Vitro Cultured Cells
3.3. Analysis of the Potential Off-Target Sites in Porcine Genome
3.3.1. Selection of the Potential Off-Target Sites Location
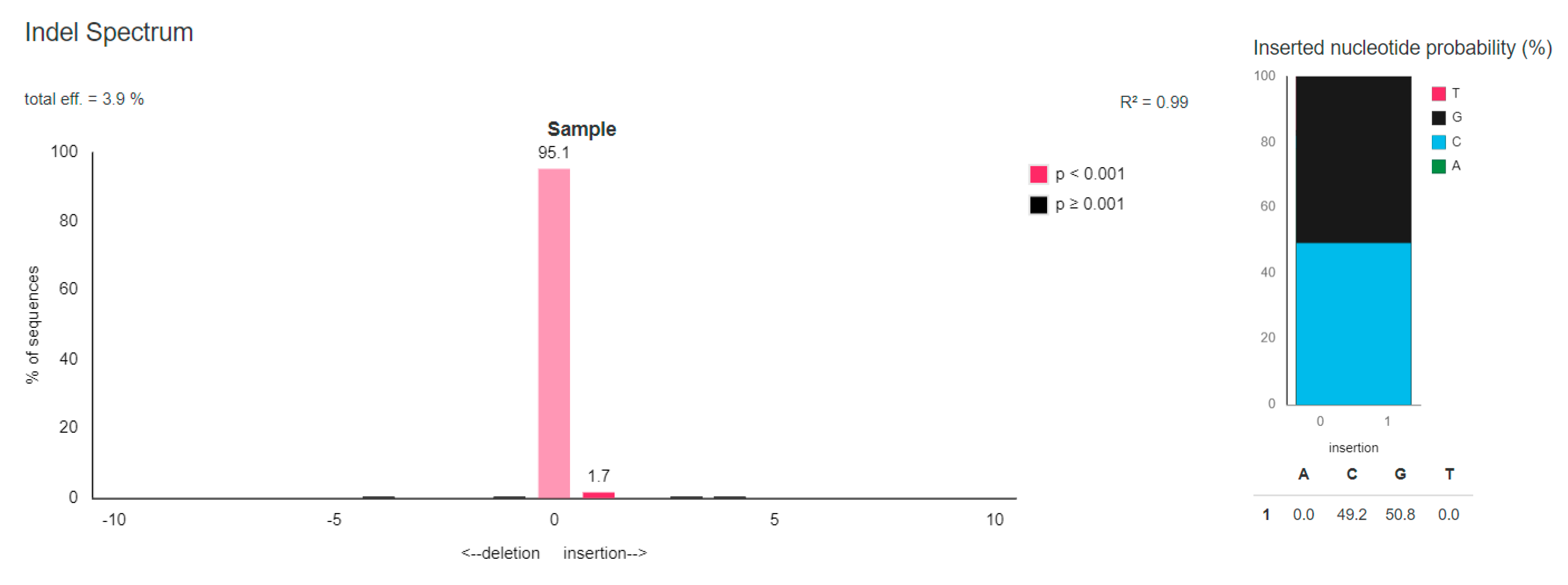
3.3.2. TIDE Analysis of the Chosen Potential Off-Target Sites
3.3.3. Comparison of Bioinformatically Predicted DNA Diruption Efficiency Results in the Off-Target Sites with Those Obtained in the In Vitro Cultured Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ekser, B.; Li, P.; Cooper, D.K.C. Xenotransplantation: Past, present, and future. Curr. Opin. Organ. Transplant. 2017, 22, 513–521. [Google Scholar] [CrossRef] [PubMed]
- Cooper, D.K.C.; Ekser, B.; Tector, J. A brief history of clinical xenotransplantation. Int. J. Surg. 2015, 23, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Aristizabal, A.M.; Caicedo, L.A.; Martínez, J.M.; Moreno, M.; Echeverri, G.J. Clinical xenotransplantation, a closer reality: Literature review. Cirugía Española (English Ed.) 2017, 95, 62–72. [Google Scholar] [CrossRef]
- Cooper, D.K.C.; Iwase, H.; Wang, L.; Yamamoto, T.; Li, Q.; Li, J.; Zhou, H.; Hara, H. Bringing home the bacon: Update on the state of kidney xenotransplantation. Blood Purif. 2018, 45, 254–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vadori, M.; Cozzi, E. The immunological barriers to xenotransplantation. Tissue Antigens 2015, 86, 239–253. [Google Scholar] [CrossRef] [PubMed]
- Cooper, D.K.C.; Ekser, B.; Tector, J. Immunobiological barriers to xenotransplantation. Int. J. Surg. 2015, 23, 211–216. [Google Scholar] [CrossRef]
- Fischer, K.; Rieblinger, B.; Hein, R.; Sfriso, R.; Zuber, J.; Fischer, A.; Klinger, B.; Liang, W.; Flisikowski, K.; Kurome, M.; et al. Viable pigs after simultaneous inactivation of porcine MHC class I and three xenoreactive antigen genes GGTA1, CMAH and B4GALNT2. Xenotransplantation 2020, 27, e12560. [Google Scholar] [CrossRef]
- Ganji, M.-R.; Broumand, B. Acute cellular rejection. Iran. J. Kidney Dis. 2007, 1, 54–56. [Google Scholar]
- Lin, C.C.; Cooper, D.K.C.; Dorling, A. Coagulation dysregulation as a barrier to xenotransplantation in the primate. Transpl. Immunol. 2009, 21, 75–80. [Google Scholar] [CrossRef] [Green Version]
- Cooper, D.K.C.; Ezzelarab, M.B.; Hara, H.; Iwase, H.; Lee, W.; Wijkstrom, M.; Bottino, R. The pathobiology of pig-to-primate xenotransplantation: A historical review. Xenotransplantation 2013, 23, 83–105. [Google Scholar] [CrossRef]
- Chuang, C.K.; Chen, C.H.; Huang, C.L.; Su, Y.H.; Peng, S.H.; Lin, T.Y.; Tai, H.C.; Yang, T.S.; Tu, C.F. Generation of GGTA1 mutant pigs by direct pronuclear microinjection of CRISPR/Cas9 plasmid vectors. Anim. Biotechnol. 2017, 28, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.S.; Hanaway, M.J.; Gonzalez-Stawinski, G.V.; Lau, C.L.; Parker, W.; Davis, R.D.; Byrne, G.W.; Diamond, L.E.; Logan, J.S.; Platt, J.L. The role of anti-Galalpha1-3Gal antibodies in acute vascular rejection and accommodation of xenografts. Transplantation 2000, 70, 1667–1674. [Google Scholar] [CrossRef] [PubMed]
- Scobie, L.; Padler-Karavani, V.; Le Bas-Bernardet, S.; Crossan, C.; Blaha, J.; Matouskova, M.; Hector, R.D.; Cozzi, E.; Vanhove, B.; Charreau, B.; et al. Long-term IgG response to porcine Neu5Gc antigens without transmission of PERV in burn patients treated with porcine skin xenografts. J. Immunol. 2013, 191, 2907–2915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varki, A. Loss of N-glycolylneuraminic acid in humans: Mechanisms, consequences, and implications for hominid evolution. Am. J. Phys. Anthropol. 2001, 116, 54–69. [Google Scholar] [CrossRef] [PubMed]
- Byrne, G.; Ahmad-Villiers, S.; Du, Z.; McGregor, C. B4GalNT2 and xenotransplantation: A newly appreciated xenogeneic antigen. Xenotransplantation 2018, 25, e12394. [Google Scholar] [CrossRef] [PubMed]
- Byrne, G.W.; Du, Z.; Stalboerger, P.; Kogelberg, H.; McGregor, C.G.A. Cloning and expression of porcine β1,4 N-acetylgalactosaminyl transferase encoding a new xenoreactive antigen. Xenotransplantation 2014, 21, 543–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.Y.; Burlak, C.; Estrada, J.L.; Li, P.; Tector, M.F.; Tector, A.J. Erythrocytes from GGTA1/CMAH knockout pigs: Implications for xenotransfusion and testing in non-human primates. Xenotransplantation 2014, 21, 376–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, D.K.C.; Bottino, R. Recent advances in understanding xenotransplantation: Implications for the clinic. Expert Rev. Clin. Immunol. 2015, 11, 1379–1390. [Google Scholar] [CrossRef] [Green Version]
- Bühler, L.; Basker, M.; Alwayn, I.P.; Goepfert, C.; Kitamura, H.; Kawai, T.; Gojo, S.; Kozlowski, T.; Ierino, F.L.; Awwad, M.; et al. Coagulation and thrombotic disorders associated with pig organ and hematopoietic cell transplantation in nonhuman primates. Transplantation 2000, 70, 1323–1331. [Google Scholar] [CrossRef]
- Lau, C.L.; Cantu, E.; Gonzalez-Stawinski, G.V.; Holzknecht, Z.E.; Nichols, T.C.; Posther, K.E.; Rayborn, C.A.; Platt, J.L.; Parker, W.; Davis, R.D. The role of antibodies and von Willebrand factor in discordant pulmonary xenotransplantation. Am. J. Transplant. 2003, 3, 1065–1075. [Google Scholar] [CrossRef]
- Cantu, E.; Balsara, K.R.; Li, B.; Lau, C.; Gibson, S.; Wyse, A.; Baig, K.; Gaca, J.; Gonzalez-Stawinski, G.V.; Nichols, T.; et al. Prolonged function of macrophage, von Willebrand factor-deficient porcine pulmonary xenografts. Am. J. Transplant. 2007, 7, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Paris, L.L.; Estrada, J.L.; Li, P.; Blankenship, R.L.; Sidner, R.A.; Reyes, L.M.; Montgomery, J.B.; Burlak, C.; Butler, J.R.; Downey, S.M.; et al. Reduced human platelet uptake by pig livers deficient in the asialoglycoprotein receptor 1 protein. Xenotransplantation 2015, 22, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Estrada, J.L.; Burlak, C.; Montgomery, J.; Butler, J.R.; Santos, R.M.; Wang, Z.Y.; Paris, L.L.; Blankenship, R.L.; Downey, S.M.; et al. Efficient generation of genetically distinct pigs in a single pregnancy using multiplexed single-guide RNA and carbohydrate selection. Xenotransplantation 2015, 22, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef]
- Du, J.; Yin, N.; Xie, T.; Zheng, Y.; Xia, N.; Shang, J.; Chen, F.; Zhang, H.; Yu, J.; Liu, F. Quantitative assessment of HR and NHEJ activities via CRISPR/Cas9-induced oligodeoxynucleotide-mediated DSB repair. DNA Repair 2018, 70, 67–71. [Google Scholar] [CrossRef]
- Miyaoka, Y.; Mayerl, S.J.; Chan, A.H.; Conklin, B.R. Detection and quantification of HDR and NHEJ induced by genome editing at endogenous gene loci using droplet digital PCR. Methods Mol. Biol. 2018, 1768, 349–362. [Google Scholar] [CrossRef]
- Fu, Y.; Foden, J.A.; Khayter, C.; Maeder, M.L.; Reyon, D.; Joung, J.K.; Sander, J.D. High-frequency off-target mutagenesis induced by CRISPR-Cas nucleases in human cells. Nat. Biotechnol. 2013, 31, 822–826. [Google Scholar] [CrossRef] [Green Version]
- Lentsch, E.; Li, L.; Pfeffer, S.; Ekici, A.B.; Taher, L.; Pilarsky, C.; Grützmann, R. CRISPR/Cas9-mediated knock-out of krasG12D mutated pancreatic cancer cell lines. Int. J. Mol. Sci. 2019, 20, 5706. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Wang, J.; Zhao, M.; Zhu, J.; Zhang, M.; Wang, Z.; Gao, Y.; Zhu, W.; Lu, H. Feasible development of stable HEK293 clones by CRISPR/Cas9-mediated site-specific integration for biopharmaceuticals production. Biotechnol. Lett. 2019, 41, 941–950. [Google Scholar] [CrossRef]
- Steyer, B.; Bu, Q.; Cory, E.; Jiang, K.; Duong, S.; Sinha, D.; Steltzer, S.; Gamm, D.; Chang, Q.; Saha, K. Scarless Genome Editing of Human Pluripotent Stem Cells via Transient Puromycin Selection. Stem Cell Rep. 2018, 10, 642–654. [Google Scholar] [CrossRef] [Green Version]
- Doench, J.G.; Fusi, N.; Sullender, M.; Hegde, M.; Vaimberg, E.W.; Donovan, K.F.; Smith, I.; Tothova, Z.; Wilen, C.; Orchard, R.; et al. Optimized sgRNA design to maximize activity and minimize off-target effects of CRISPR-Cas9. Nat. Biotechnol. 2016, 34, 184–191. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.; Xu, J.; Cheng, M.; Liao, X.; Peng, S. Review of CRISPR/Cas9 sgRNA Design Tools. Interdiscip. Sci. 2018, 10, 455–465. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Steentoft, C.; Hauge, C.; Hansen, L.; Thomsen, A.L.; Niola, F.; Vester-Christensen, M.B.; Frödin, M.; Clausen, H.; Wandall, H.H.; et al. Fast and sensitive detection of indels induced by precise gene targeting. Nucleic Acids Res. 2015, 43, e59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sentmanat, M.F.; Peters, S.T.; Florian, C.P.; Connelly, J.P.; Pruett-Miller, S.M. A survey of validation strategies for CRISPR-Cas9 editing. Sci. Rep. 2018, 8, 888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosicki, M.; Rajan, S.S.; Lorenzetti, F.C.; Wandall, H.H.; Narimatsu, Y.; Metzakopian, E.; Bennett, E.P. Dynamics of indel profiles induced by various CRISPR/Cas9 delivery methods. Prog. Mol. Biol. Transl. Sci. 2017, 152, 49–67. [Google Scholar] [CrossRef]
- Peng, H.; Zheng, Y.; Zhao, Z.; Liu, T.; Li, J. Recognition of CRISPR/Cas9 off-target sites through ensemble learning of uneven mismatch distributions. Bioinformatics 2018, 34, i757–i765. [Google Scholar] [CrossRef] [Green Version]
- Kuscu, C.; Arslan, S.; Singh, R.; Thorpe, J.; Adli, M. Genome-wide analysis reveals characteristics of off-target sites bound by the Cas9 endonuclease. Nat. Biotechnol. 2014, 32, 677–683. [Google Scholar] [CrossRef]
- Zheng, T.; Hou, Y.; Zhang, P.; Zhang, Z.; Xu, Y.; Zhang, L.; Niu, L.; Yang, Y.; Liang, D.; Yi, F.; et al. Profiling single-guide RNA specificity reveals a mismatch sensitive core sequence. Sci. Rep. 2017, 7, 40638. [Google Scholar] [CrossRef] [Green Version]
- Wienert, B.; Wyman, S.K.; Richardson, C.D.; Yeh, C.D.; Akcakaya, P.; Porritt, M.J.; Morlock, M.; Vu, J.T.; Kazane, K.R.; Watry, H.L.; et al. Unbiased detection of CRISPR off-targets in vivo using DISCOVER-Seq. Science 2019, 364, 286–289. [Google Scholar] [CrossRef]
- Tsai, S.Q.; Nguyen, N.T.; Malagon-Lopez, J.; Topkar, V.V.; Aryee, M.J.; Joung, J.K. CIRCLE-seq: A highly sensitive in vitro screen for genome-wide CRISPR-Cas9 nuclease off-targets. Nat. Methods 2017, 14, 607–614. [Google Scholar] [CrossRef] [Green Version]
- Pliatsika, V.; Rigoutsos, I. Off-Spotter’: Very fast and exhaustive enumeration of genomic lookalikes for designing CRISPR/Cas guide RNAs. Biol. Direct 2015, 10, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, A.; Cheng, Z.; Kong, L.; Zhu, Z.; Lin, S.; Gao, G.; Zhang, B. CasOT: A genome-wide Cas9/gRNA off-target searching tool. Bioinformatics 2014, 30, 1180–1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, Y.; Xie, X.; Wang, Y.; Ma, W.; Liang, P.; Songyang, Z.; Dai, Z. pgRNAFinder: A web-based tool to design distance independent paired-gRNA. Bioinformatics 2017, 33, 3642–3644. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.J.; Holmes, B.R.; Aronin, N.; Brodsky, M.H. CRISPRseek: A Bioconductor package to identify target-specific guide RNAs for CRISPR-Cas9 genome-editing systems. PLoS ONE 2014, 9, e108424. [Google Scholar] [CrossRef] [Green Version]
- Cong, L.; Ran, F.A.; Cox, D.; Lin, S.; Barretto, R.; Habib, N.; Hsu, P.D.; Wu, X.; Jiang, W.; Marraffini, L.A.; et al. Multiplex Genome Engineering Using CRISPR/Cas Systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef] [Green Version]
- Fu, B.; Smith, J.D.; Fuchs, R.T.; Mabuchi, M.; Curcuru, J.; Robb, G.B.; Fire, A.Z. Target-dependent nickase activities of the CRISPR–Cas nucleases Cpf1 and Cas9. Nat. Microbiol. 2019, 4, 888–897. [Google Scholar] [CrossRef]
- Lyu, P.; Javidi-Parsijani, P.; Atala, A.; Lu, B. Delivering Cas9/sgRNA ribonucleoprotein (RNP) by lentiviral capsid-based bionanoparticles for efficient ‘hit-and-run’ genome editing. Nucleic Acids Res. 2019, 47, e99. [Google Scholar] [CrossRef] [Green Version]
- Martin, R.M.; Ikeda, K.; Cromer, M.K.; Uchida, N.; Nishimura, T.; Romano, R.; Tong, A.J.; Lemgart, V.T.; Camarena, J.; Pavel-Dinu, M.; et al. Highly Efficient and Marker-free Genome Editing of Human Pluripotent Stem Cells by CRISPR-Cas9 RNP and AAV6 Donor-Mediated Homologous Recombination. Cell Stem Cell 2019, 24, 821–828.e5. [Google Scholar] [CrossRef]
- Oh, S.A.; Seki, A.; Rutz, S. Ribonucleoprotein Transfection for CRISPR/Cas9-Mediated Gene Knockout in Primary T Cells. Curr. Protoc. Immunol. 2019, 124, e69. [Google Scholar] [CrossRef] [Green Version]
- Seki, A.; Rutz, S. Optimized RNP transfection for highly efficient CRI SPR/Cas9-mediated gene knockout in primary T cells. J. Exp. Med. 2018, 215, 985–997. [Google Scholar] [CrossRef]
- Vakulskas, C.A.; Behlke, M.A. Evaluation and reduction of crispr off-target cleavage events. Nucleic Acid Ther. 2019, 29, 167–174. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Porcine Genome Locus | gRNA | Exon | Chromosome Localization 1 |
|---|---|---|---|
| GGTA1 | gGGTA1 F1/R1 | Exon 8 | Chromosome 1, c261513705-261513686 (NC_010443.5) |
| gGGTA1 F2/R2 | Exon 8 | Chromosome 1, c261513541-261513522 (NC_010443.5) | |
| gGGTA1 F3/R3 | Exon 8 | Chromosome 1, c261513764-261513745 (NC_010443.5) | |
| CMAH | gCMAH F1/R1 | Exon 6 | Chromosome 7, c19902027-19902008 (NC_010449.5) |
| gCMAH F2/R2 | Exon 3 | Chromosome 7, c19917616-19917597 (NC_010449.5) | |
| gCMAH F3/R3 | Exon 5 | Chromosome 7, c19903792-19903773 (NC_010449.5) | |
| β4GalNT2 | gβ4GalNT2 F1/R1 | Exon 2 | Chromosome 12, c25388178-25388159 (NC_010454.4) |
| gβ4GalNT2 F2/R2 | Exon 3 | Chromosome 12, c25386323-25386304 (NC_010454.4) | |
| gβ4GalNT2 F3/R3 | Exon 6 | Chromosome 12, c25381330-25381311 (NC_010454.4) | |
| vWR | gvWR F1/R1 | Exon 2 | Chromosome 5, 64553818-64553837 (NC_010447.5) |
| gvWR F2/R2 | Exon 3 | Chromosome 5, 64556041-64556060 (NC_010447.5) | |
| gvWR F3/R3 | Exon 4 | Chromosome 5, 64557621-64557640 (NC_010447.5) | |
| ASGR1 | gASGR1 F1/R1 | Exon 3 | Chromosome 12, c52538530-52538511 (NC_010454.4) |
| gASGR1 F2/R2 | Exon 7 | Chromosome 12, c52537633-52537614 (NC_010454.4) | |
| gASGR1 F3/R3 | Exon 9 | Chromosome 12, c52537146-52537127 (NC_010454.4) |
| gRNA | In Silico Analysis Predicted Total Efficiency | In Vitro Analysis Total Efficiency |
|---|---|---|
| gGGTA1 F1/R1 | 78.9% | 70.1% * |
| gGGTA1 F2/R2 | 72.2% | 27.1% |
| gGGTA1 F3/R3 | 70.6% | 18.2% |
| gCMAH F1/R1 | 61.5% | 7.8% |
| gCMAH F2/R2 | 73.5% | 12.9% |
| gCMAH F3/R3 | 70.1% | 61.2% * |
| gβ4GalNT2 F1/R1 | 53.3% | 8.1% |
| gβ4GalNT2 F2/R2 | 66% | 17.6% |
| gβ4GalNT2 F3/R3 | 75.8% | 45.2% * |
| gvWF F1/R1 | 48.9% | 5.2% |
| gvWF F2/R2 | 73.9% | 85.1% * |
| gvWF F3/R3 | 76% | 39.6% |
| gASGR1 F1/R1 | 72.7% | 13.9% |
| gASGR1 F2/R2 | 67.3% | 33.5% |
| gASGR1 F3/R3 | 68.7% | 80.5% * |
| Chosen gRNA | * | Sequence | Porcine Genome Localization |
|---|---|---|---|
| gGGTA1 F1/R1 | 1. | GCTGCACTTGAAGACCATCG | chr7: +33592796 |
| 2. | GATAGTCATGGAGACCATCG | chr7: +1925303 (RIPK1 gene: ENSSSCG00000001009) | |
| 3. | CCTGCGCGTGAAGACCAACG | chr2: -44588168 (OTOG gene: ENSSSCG00000013376) | |
| 4. | GAGGTGCATGAAGAACATCT | chr2: +13174290 | |
| gCMAH F3/R3 | 5. | ATTCGATCCTCCTAACCCCT | chr15: +40937188 |
| 6. | TCTTAACCCTCATAACCCGT | chr4: -97128319 | |
| 7. | AATAAATCACCCTAACCAGT | chr4: +116709576 (HIPK1 gene: ENSSSCG00000006760) | |
| gβ4Gal NT2 F3/R3 | 8. | AAACTACCAGCTCCACAGAG | chr16: -5677136 |
| 9. | ATTGTACCACCTCCACAGAC | chr10: -13115375 | |
| 10. | TCAGTATCACCTCCACAGAG | chr7: -109695811 | |
| gvWF F2/R2 | 11. | CCTTCTGCTTCATGCCCGCG | chr6: +157052374 |
| 12. | GCACGTACTCCTTGCCCGCG | chr4: -347951 (ARHGAP39 gene: ENSSSCG00000005894) | |
| 13. | CCGTGTCGTCCAGGCCCGCG | chr6: -9684887 (WWOX gene: ENSSSCG00000027415) | |
| 14. | CCCTGTCCTGCAGGCCTGCG | chr14: -55036169 (COMT gene: ENSSSCG00000010132) | |
| gASGR1 F3/R3 | 15. | GCATATGTCTGGTACGGGCA | chr6: +4310372 |
| 16. | GCATATGTCTGGTACGGGCA | chr6: -4126764 | |
| 17. | CACAATGACAGGTACGGGCA | chr5: +67768944 (KCNA1 gene: ENSSSCG00000000716) | |
| 18. | CCAGACGACTGGCACGGGCA | chr12: -54751930 | |
| 19. | CCCGCTGTCTGGGACGGGCA | chr6: +155398812 (C1orf210 gene: ENSSSCG00000003951) |
| gRNA | * | DNA Hydrolysis Efficiency at a Potential Off-Target Site | |
|---|---|---|---|
| Bioinformatic Analysis | In Vitro Cultured Cells | ||
| gGGTA1 F1/R1 | 1. | 1.6% | 3.9% |
| 2. | 0.5% | 0% | |
| 3. | 0.4% | 0% | |
| 4. | 0.2% | 0.8% | |
| gCMAH F3/R3 | 5. | 0.6% | 1.1% |
| 6. | 0.6% | 0% | |
| 7. | 0.2% | 0% | |
| gB4GalNT2 F3/R3 | 8. | 2.6% | 0% |
| 9. | 1.8% | 0% | |
| 10. | 1.8% | 2.2% | |
| gvWF F2/R2 | 11. | 0.9% | 1.8% |
| 12. | 0.8% | 0% | |
| 13. | 0.4% | 0% | |
| 14. | 0.2% | 3.4% | |
| gASGR1 F3/R3 | 15. | 1.5% | 0% |
| 16. | 1.5% | 0% | |
| 17. | 1.4% | 0% | |
| 18. | 0.7% | 1.7% | |
| 19. | 0.6% | 1.6% | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ryczek, N.; Hryhorowicz, M.; Lipiński, D.; Zeyland, J.; Słomski, R. Evaluation of the CRISPR/Cas9 Genetic Constructs in Efficient Disruption of Porcine Genes for Xenotransplantation Purposes Along with an Assessment of the Off-Target Mutation Formation. Genes 2020, 11, 713. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11060713
Ryczek N, Hryhorowicz M, Lipiński D, Zeyland J, Słomski R. Evaluation of the CRISPR/Cas9 Genetic Constructs in Efficient Disruption of Porcine Genes for Xenotransplantation Purposes Along with an Assessment of the Off-Target Mutation Formation. Genes. 2020; 11(6):713. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11060713
Chicago/Turabian StyleRyczek, Natalia, Magdalena Hryhorowicz, Daniel Lipiński, Joanna Zeyland, and Ryszard Słomski. 2020. "Evaluation of the CRISPR/Cas9 Genetic Constructs in Efficient Disruption of Porcine Genes for Xenotransplantation Purposes Along with an Assessment of the Off-Target Mutation Formation" Genes 11, no. 6: 713. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11060713