Indication of Premelanosome Protein (PMEL) Expression Outside of Pigmented Bovine Skin Suggests Functions Beyond Eumelanogenesis

 , , and
, , and 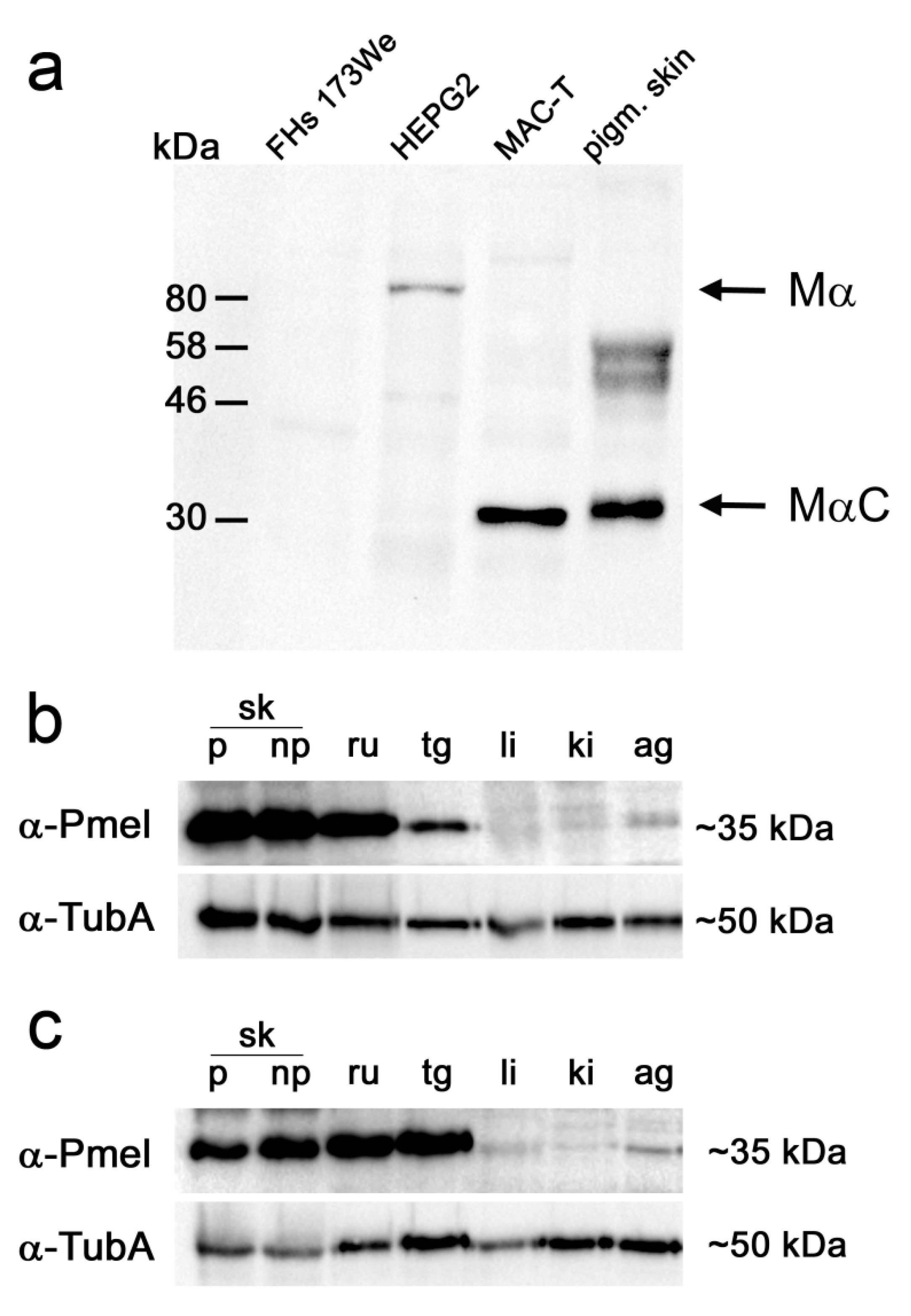{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animals and Sampling
2.3. Protein Isolation
2.4. Western Blot
2.5. Immunohistochemistry (IHC)
2.6. Immunocytochemistry
3. Results
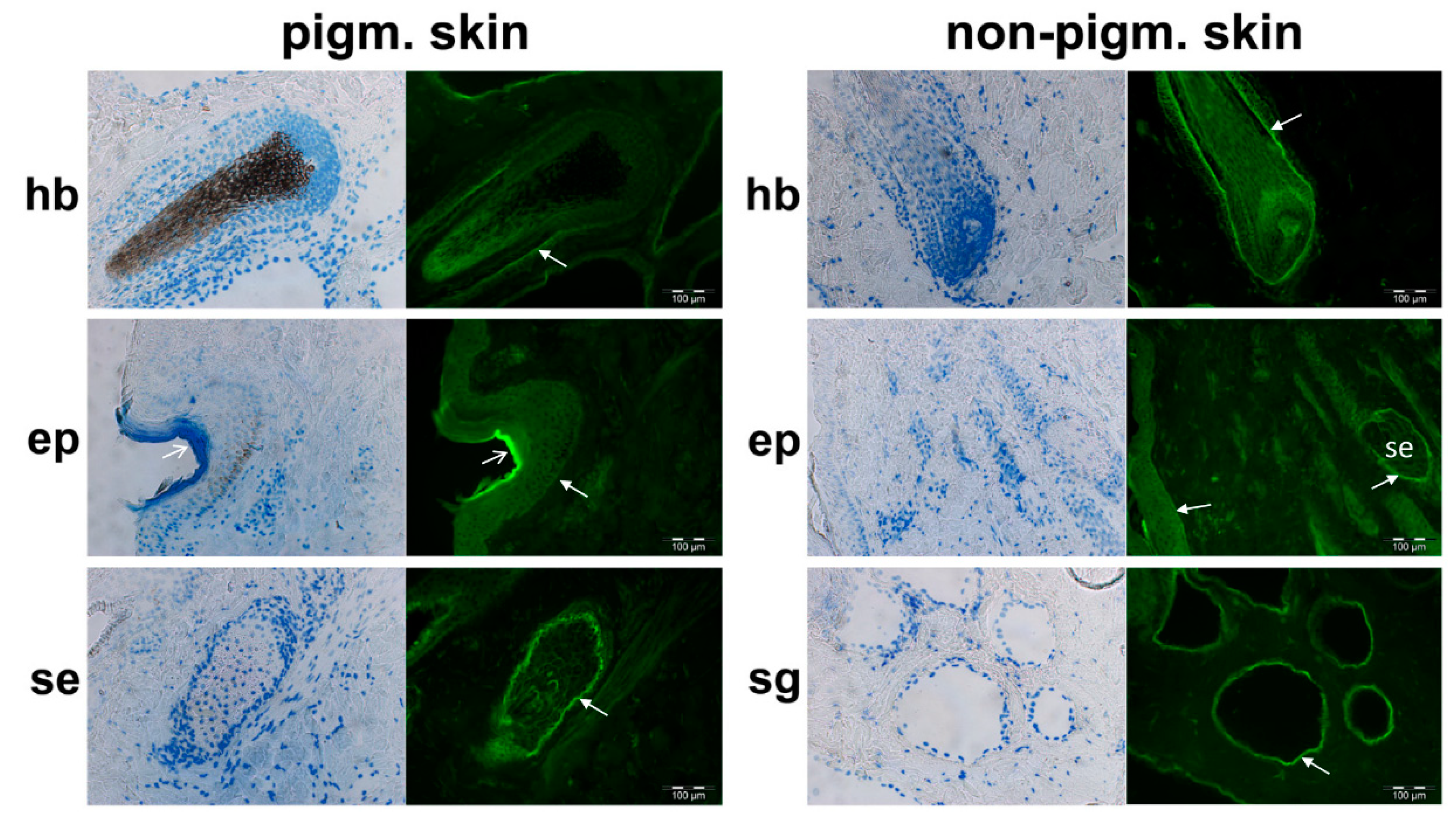
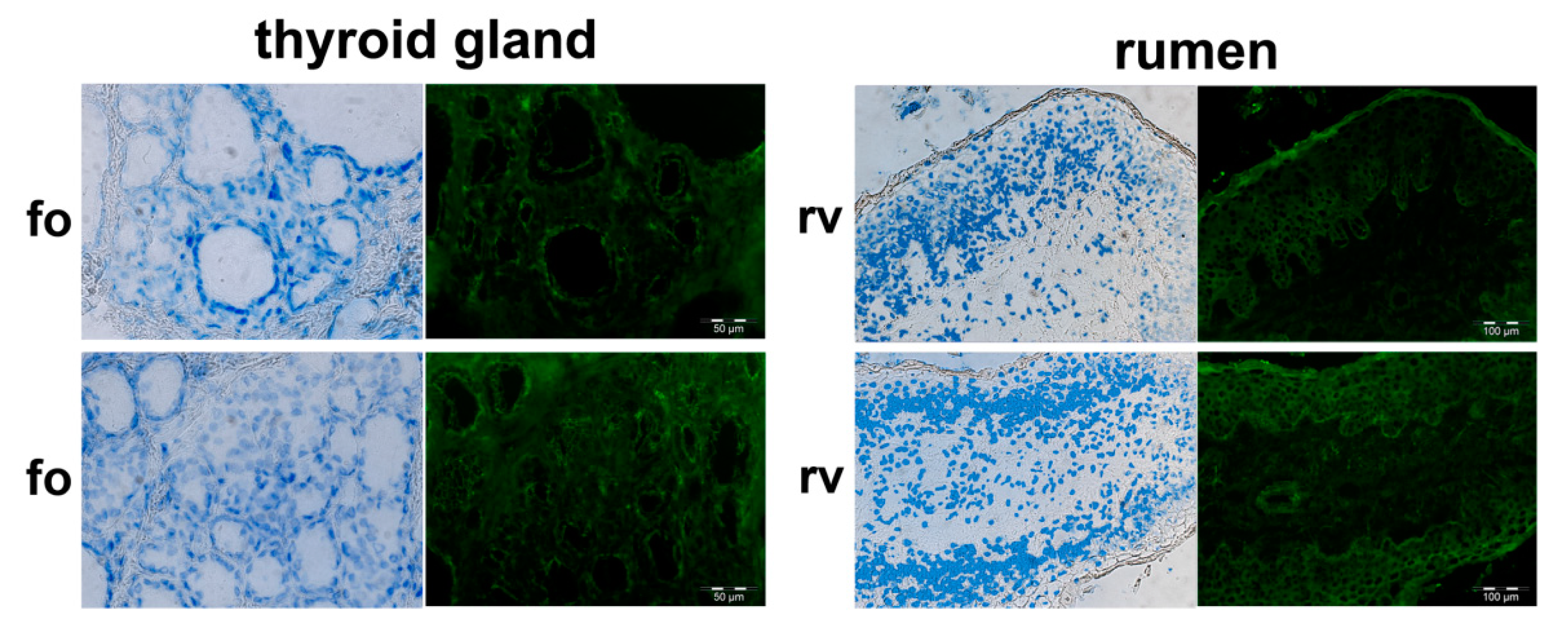
3.1. PMEL Signals Are Detected in Pigmented and Non-Pigmented Tissues
3.2. Bovine PMEL Protein Seems to Be Present not Only in Cells of the Melanocyte Lineage
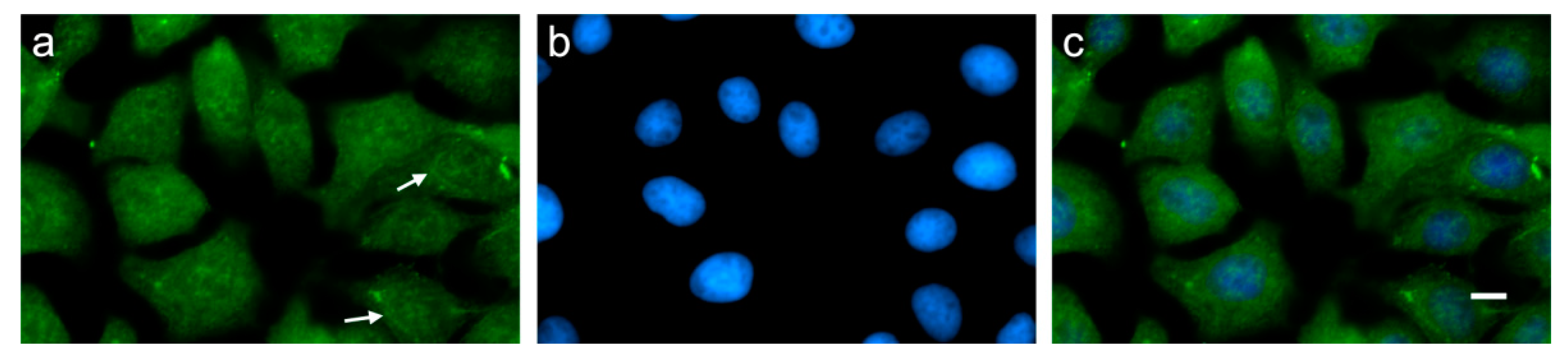
3.3. Indication of Putative Bovine PMEL Protein Expression in non-Melanocyte Cells
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Theos, A.C.; Truschel, S.T.; Raposo, G.; Marks, M.S. The Silver locus product Pmel17/gp100/Silv/ME20: Controversial in name and in function. Pigment Cell Res. 2005, 18, 322–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellstrom, A.R.; Watt, B.; Fard, S.S.; Tenza, D.; Mannstrom, P.; Narfstrom, K.; Ekesten, B.; Ito, S.; Wakamatsu, K.; Larsson, J.; et al. Inactivation of Pmel alters melanosome shape but has only a subtle effect on visible pigmentation. PLoS Genet. 2011, 7, e1002285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, T.; Vieira, W.D.; Potterf, B.; Sakai, C.; Imokawa, G.; Hearing, V.J. Modulation of melanogenic protein expression during the switch from eumelanogenesis to pheomelanogenesis. J. Cell Sci. 1995, 108, 2301–2309. [Google Scholar] [PubMed]
- Watt, B.; van Niel, G.; Raposo, G.; Marks, M.S. PMEL: A pigment cell-specific model for functional amyloid formation. Pigment Cell Melanoma Res. 2013, 26, 300–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Esparza, M.M.; Jimenez-Cervantes, C.; Bennett, D.C.; Lozano, J.A.; Solano, F.; Garcia-Borron, J.C. The mouse silver locus encodes a single transcript truncated by the silver mutation. Mamm. Genome 1999, 10, 1168–1171. [Google Scholar] [CrossRef]
- Kerje, S.; Sharma, P.; Gunnarsson, U.; Kim, H.; Bagchi, S.; Fredriksson, R.; Schutz, K.; Jensen, P.; von Heijne, G.; Okimoto, R.; et al. The Dominant white, Dun and Smoky color variants in chicken are associated with insertion/deletion polymorphisms in the PMEL17 gene. Genetics 2004, 168, 1507–1518. [Google Scholar] [CrossRef] [Green Version]
- Clark, L.A.; Wahl, J.M.; Rees, C.A.; Murphy, K.E. Retrotransposon insertion in SILV is responsible for merle patterning of the domestic dog. Proc. Natl. Acad. Sci. USA 2006, 103, 1376–1381. [Google Scholar] [CrossRef] [Green Version]
- Brunberg, E.; Andersson, L.; Cothran, G.; Sandberg, K.; Mikko, S.; Lindgren, G. A missense mutation in PMEL17 is associated with the Silver coat color in the horse. BMC Genet. 2006, 7, 46. [Google Scholar] [CrossRef] [PubMed]
- Kuehn, C.; Weikard, R. An investigation into the genetic background of coat colour dilution in a Charolais x German Holstein F2 resource population. Anim. Genet. 2007, 38, 109–113. [Google Scholar] [CrossRef]
- Schmutz, S.M.; Dreger, D.L. Interaction of MC1R and PMEL alleles on solid coat colors in Highland cattle. Anim. Genet. 2013, 44, 9–13. [Google Scholar] [CrossRef]
- Gutierrez-Gil, B.; Wiener, P.; Williams, J. Genetic effects on coat colour in cattle: Dilution of eumelanin and phaeomelanin pigments in an F2-Backcross Charolais * Holstein population. BMC Genet. 2007, 8, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knaust, J.; Hadlich, F.; Weikard, R.; Kuehn, C. Epistatic interactions of at least three loci determine the ‘rat-tail’ phenotype in cattle. Genet. Sel. Evol. 2016, 48, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ordonez, N.G. Value of melanocytic-associated immunohistochemical markers in the diagnosis of malignant melanoma: A review and update. Hum. Pathol. 2014, 45, 191–205. [Google Scholar] [CrossRef]
- Brouwenstijn, N.; Slager, E.H.; Bakker, A.B.; Schreurs, M.W.; Van der Spek, C.W.; Adema, G.J.; Schrier, P.I.; Figdor, C.G. Transcription of the gene encoding melanoma-associated antigen gp100 in tissues and cell lines other than those of the melanocytic lineage. Br. J. Cancer 1997, 76, 1562–1566. [Google Scholar] [CrossRef] [Green Version]
- Kuehn, C.; Weikard, R. Multiple splice variants within the bovine silver homologue (SILV) gene affecting coat color in cattle indicate a function additional to fibril formation in melanophores. BMC Genom. 2007, 8, 335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klungland, H.; Vage, D.I.; Gomezraya, L.; Adalsteinsson, S.; Lien, S. The Role of Melanocyte-Stimulating Hormone (Msh) Receptor in Bovine Coat Color Determination. Mamm. Genome 1995, 6, 636–639. [Google Scholar] [CrossRef]
- Haase, B.; Brooks, S.A.; Schlumbaum, A.; Azor, P.J.; Bailey, E.; Alaeddine, F.; Mevissen, M.; Burger, D.; Poncet, P.A.; Rieder, S.; et al. Allelic heterogeneity at the equine KIT locus in dominant white (W) horses. PLoS Genet. 2007, 3, 2101–2108. [Google Scholar] [CrossRef] [Green Version]
- Leonhardt, R.M.; Vigneron, N.; Hee, J.S.; Graham, M.; Cresswella, P. Critical residues in the PMEL/Pmel17 N-terminus direct the hierarchical assembly of melanosomal fibrils. Mol. Biol. Cell 2013, 24, 964–981. [Google Scholar] [CrossRef]
- Hayakawa, K.; Nagamine, T. Fucoidan-dependent Increased Membrane Components in HepG2 Cells: Effect of Fucoidan Is Not Due to Gene Expression. Cancer Genom. Proteom. 2014, 11, 93–113. [Google Scholar]
- Huynh, H.T.; Robitaille, G.; Turner, J.D. Establishment of Bovine Mammary Epithelial-Cells (Mac-T)—An Invitro Model for Bovine Lactation. Exp. Cell Res. 1991, 197, 191–199. [Google Scholar] [CrossRef]
- Ho, T.; Watt, B.; Spruce, L.A.; Seeholzer, S.H.; Marks, M.S. The Kringle-like Domain Facilitates Post-endoplasmic Reticulum Changes to Premelanosome Protein (PMEL) Oligomerization and Disulfide Bond Configuration and Promotes Amyloid Formation. J. Biol. Chem. 2016, 291, 3595–3612. [Google Scholar] [CrossRef] [Green Version]
- Hee, J.S.; Mitchell, S.M.; Liu, X.; Leonhardt, R.M. Melanosomal formation of PMEL core amyloid is driven by aromatic residues. Sci. Rep. 2017, 7, 44064. [Google Scholar] [CrossRef] [PubMed]
- Harper, D.C.; Theos, A.C.; Herman, K.E.; Tenza, D.; Raposo, G.; Marks, M.S. Premelanosome amyloid-like fibrils are composed of only Golgi-processed forms of Pmel17 that have been proteolytically processed in Endosomes. J. Biol. Chem. 2008, 283, 2307–2322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yasumoto, K.; Watabe, H.; Valencia, J.C.; Kushimoto, T.; Kobayashi, T.; Appella, E.; Hearing, V.J. Epitope mapping of the melanosomal matrix protein gp100 (PMEL17): Rapid processing in the endoplasmic reticulum and glycosylation in the early Golgi compartment. J. Biol. Chem. 2004, 279, 28330–28338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giordano, F.; Bonetti, C.; Surace, E.M.; Marigo, V.; Raposo, G. The ocular albinism type 1 (OA1) G-protein-coupled receptor functions with MART-1 at early stages of melanogenesis to control melanosome identity and composition. Hum. Mol. Genet. 2009, 18, 4530–4545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yajima, I.; Larue, L. The location of heart melanocytes is specified and the level of pigmentation in the heart may correlate with coat color. Pigment Cell Melanoma Res. 2008, 21, 471–476. [Google Scholar] [CrossRef]
- Peinado, H.; Aleckovic, M.; Lavotshkin, S.; Matei, I.; Costa-Silva, B.; Moreno-Bueno, G.; Hergueta-Redondo, M.; Williams, C.; Garcia-Santos, G.; Ghajar, C.M.; et al. Melanoma exosomes educate bone marrow progenitor cells toward a pro-metastatic phenotype through MET. Nat. Med. 2012, 18, 883–891. [Google Scholar] [CrossRef] [Green Version]
- Uhlen, M.; Fagerberg, L.; Hallstrom, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, A.; Kampf, C.; Sjostedt, E.; Asplund, A.; et al. Proteomics. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Nolte, W.; Weikard, R.; Brunner, R.M.; Albrecht, E.; Hammon, H.M.; Reverter, A.; Kühn, C. Biological Network Approach for the Identification of Regulatory Long Non-Coding RNAs Associated With Metabolic Efficiency in Cattle. Front. Genet. 2019, 10, 1130. [Google Scholar] [CrossRef] [Green Version]
- Donatien, P.D.; Orlow, S.J. Interaction of melanosomal proteins with melanin. Eur. J. Biochem. 1995, 232, 159–164. [Google Scholar] [CrossRef]
- Raposo, G.; Marks, M.S. Melanosomes-dark organelles enlighten endosomal membrane transport. Nat. Rev. Mol. Cell Biol. 2007, 8, 786–797. [Google Scholar] [CrossRef] [Green Version]
- Okamoto, N.; Aoto, T.; Uhara, H.; Yamazaki, S.; Akutsu, H.; Umezawa, A.; Nakauchi, H.; Miyachi, Y.; Saida, T.; Nishimura, E.K. A melanocyte-melanoma precursor niche in sweat glands of volar skin. Pigment Cell Melanoma Res. 2014, 27, 1039–1050. [Google Scholar] [CrossRef] [Green Version]
- Barden, H.; Levine, S. Histochemical-Observations on rodent brain melanin. Brain Res. Bull. 1983, 10, 847–851. [Google Scholar] [CrossRef]
- Markert, C.L.; Silvers, W.K. The effects of genotype and cell environment on melanoblast differentiation in the house mouse. Genetics 1956, 41, 429–450. [Google Scholar] [PubMed]
- Nichols, S.E.; Reams, W.M. The occurrence and morphogenesis of melanocytes in the connective tissues of the pet/mcv mouse strain. J. Embryol. Exp. Morphol. 1960, 8, 24–32. [Google Scholar] [PubMed]
- Theriault, L.L.; Hurley, L.S. Ultrastructure of developing melanosomes in c57 black mice and pallid mice. Dev. Biol. 1970, 23, 261–275. [Google Scholar] [CrossRef]
- Sommer, L. Generation of melanocytes from neural crest cells. Pigment Cell Melanoma Res. 2011, 24, 411–421. [Google Scholar] [CrossRef]
- Weiner, L.; Fu, W.Y.; Chirico, W.J.; Brissette, J.L. Skin as a living coloring book: How epithelial cells create patterns of pigmentation. Pigment Cell Melanoma Res. 2014, 27, 1014–1031. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.F.; Hammer, J.A. Melanosome transfer: It is best to give and receive. Curr. Opin. Cell Biol. 2014, 29, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.Y.; Fisher, D.E. Melanocyte biology and skin pigmentation. Nature 2007, 445, 843–850. [Google Scholar] [CrossRef]
- Schalles, R.R.; Cundiff, L.V. Inheritance of the ‘rat-tail’ syndrome and its effect on calf performance. J. Anim. Sci. 1999, 77, 1144–1147. [Google Scholar] [CrossRef] [PubMed]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Knaust, J.; Weikard, R.; Albrecht, E.; Brunner, R.M.; Günther, J.; Kühn, C. Indication of Premelanosome Protein (PMEL) Expression Outside of Pigmented Bovine Skin Suggests Functions Beyond Eumelanogenesis. Genes 2020, 11, 788. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11070788
Knaust J, Weikard R, Albrecht E, Brunner RM, Günther J, Kühn C. Indication of Premelanosome Protein (PMEL) Expression Outside of Pigmented Bovine Skin Suggests Functions Beyond Eumelanogenesis. Genes. 2020; 11(7):788. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11070788
Chicago/Turabian StyleKnaust, Jacqueline, Rosemarie Weikard, Elke Albrecht, Ronald M. Brunner, Juliane Günther, and Christa Kühn. 2020. "Indication of Premelanosome Protein (PMEL) Expression Outside of Pigmented Bovine Skin Suggests Functions Beyond Eumelanogenesis" Genes 11, no. 7: 788. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11070788