TLR4 Receptor D299G/T399I Haplotype Polymorphism Is Associated with Insulin Resistance in Obese Female Subjects

 ,
,
Abstract
:1. Introduction
2. Methods
2.1. Study Protocol
2.2. Study Design
2.3. Anthropometrics Measurements
2.4. Biochemical Assays
2.5. Genotyping of the Polymorphisms of the Variants rs4986790 and rs4986791
2.6. Flowcytometry and Expression of TLR4 Receptors on Monocytes and Macrophages
2.7. Definitions and Cut-Off Values in the Study
2.8. Statistical Analysis
2.9. Power Calculations
3. Results
3.1. Phenotype Characteristics: Anthropometric and Biochemical Data of the Study Subjects
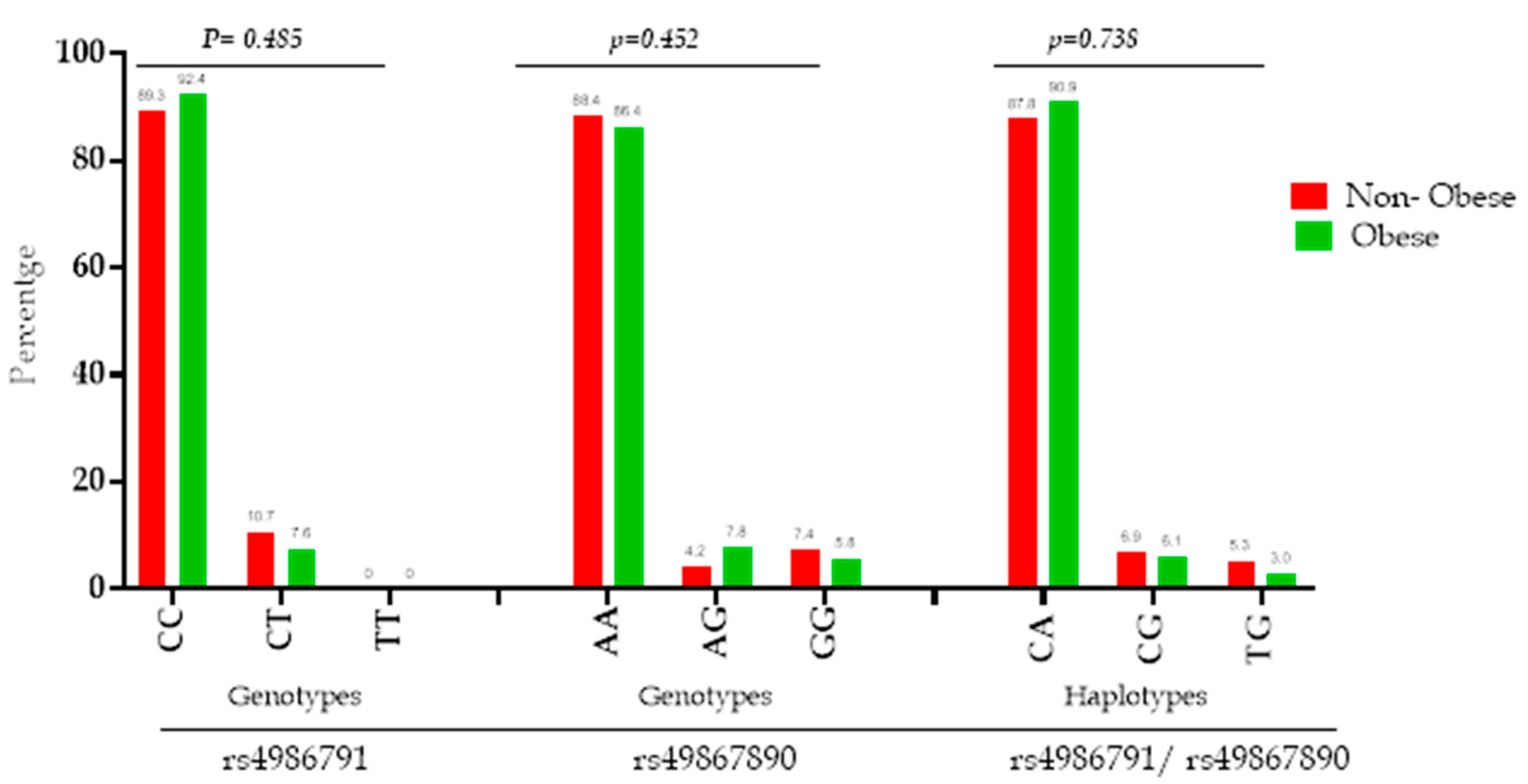
3.2. TLR4 Genetic and Haplotype Distribution and Its Association with Obesity Phenotypes
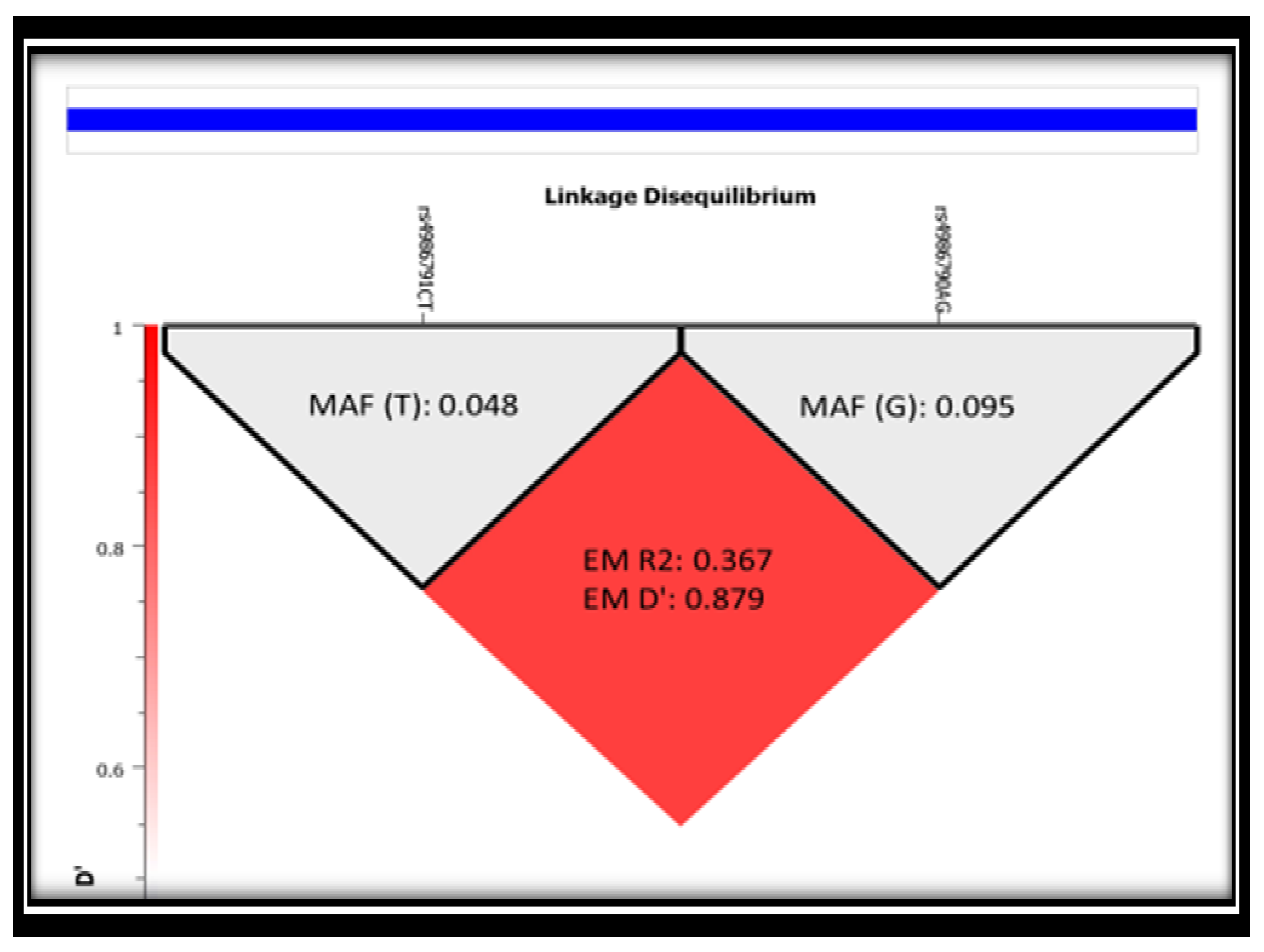
3.3. Linkage Disequilibrium (LD) of the Two SNPs (rs4986791 and rs4986790) in Study Subjects
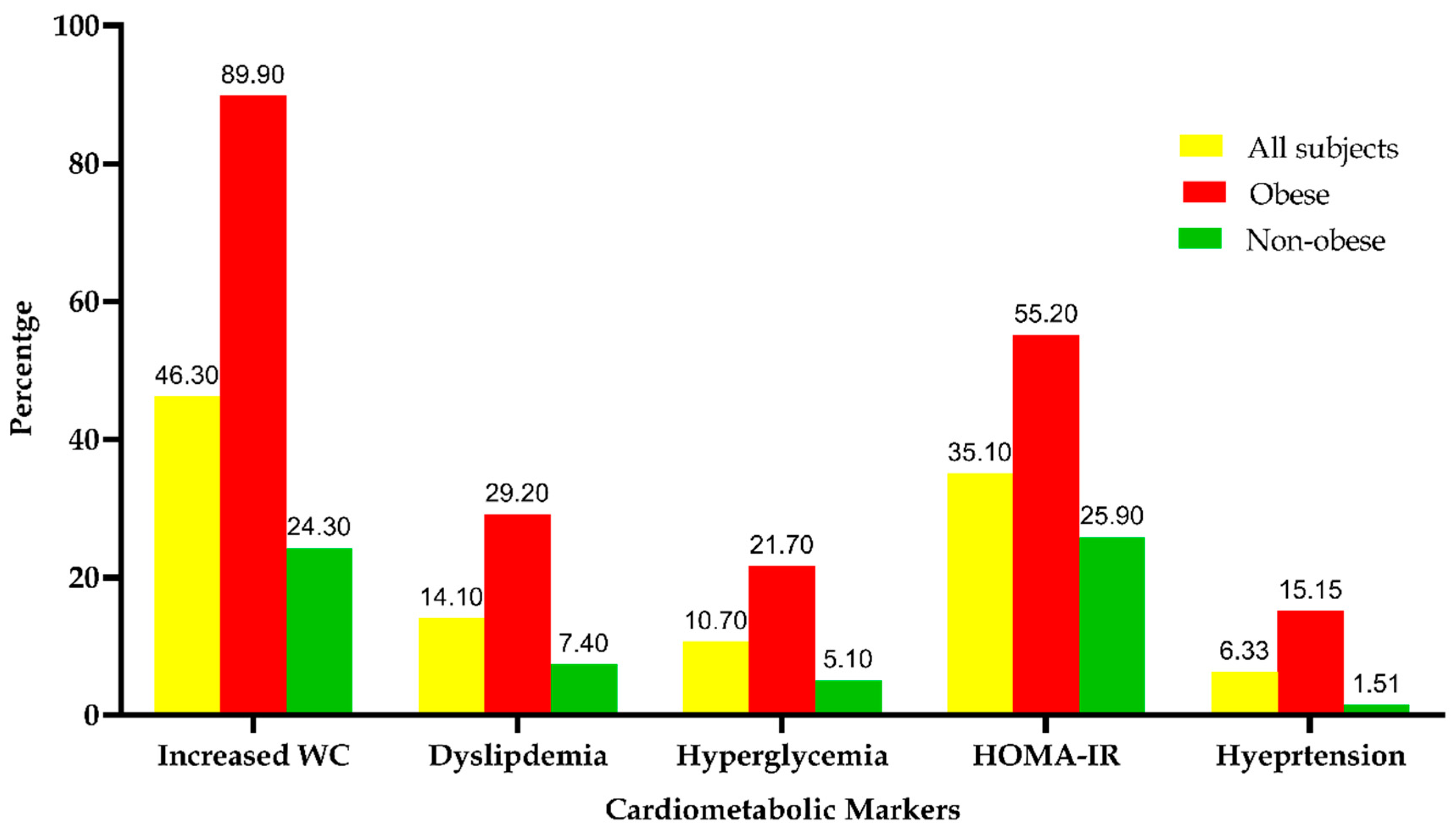
3.4. The Frequency Distribution of the Metabolic Syndrome among the Study Subjects
3.5. Association between Obesity Phenotypes and Dominant Genetic Models of rs4986790 and SNP rs4986791 among the Study Subjects
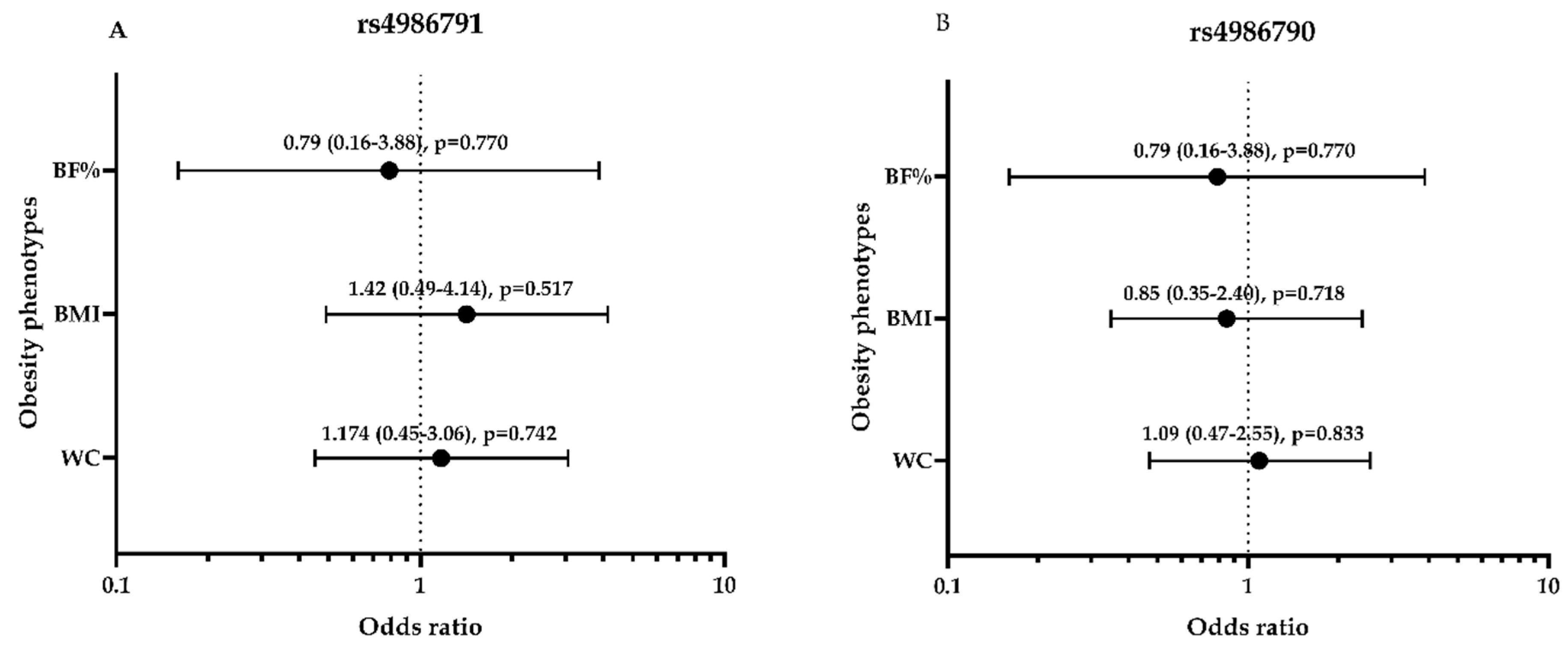
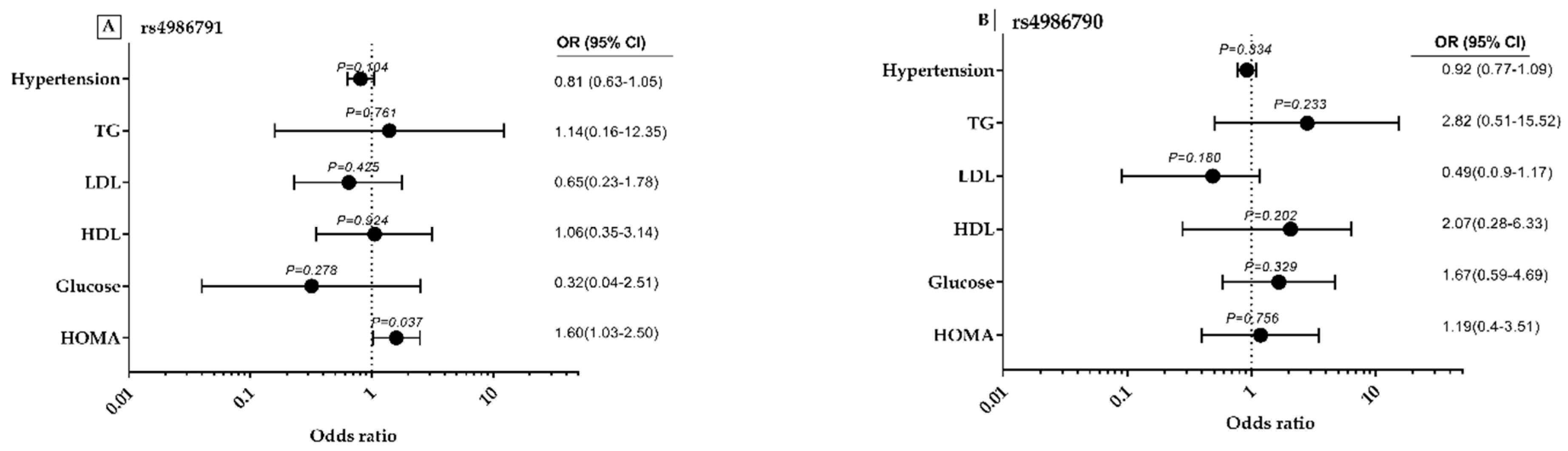
3.6. rs4986791 and rs4986790 Polymorphisms-Related Odds Ratios with MS Components and Lipid Profile
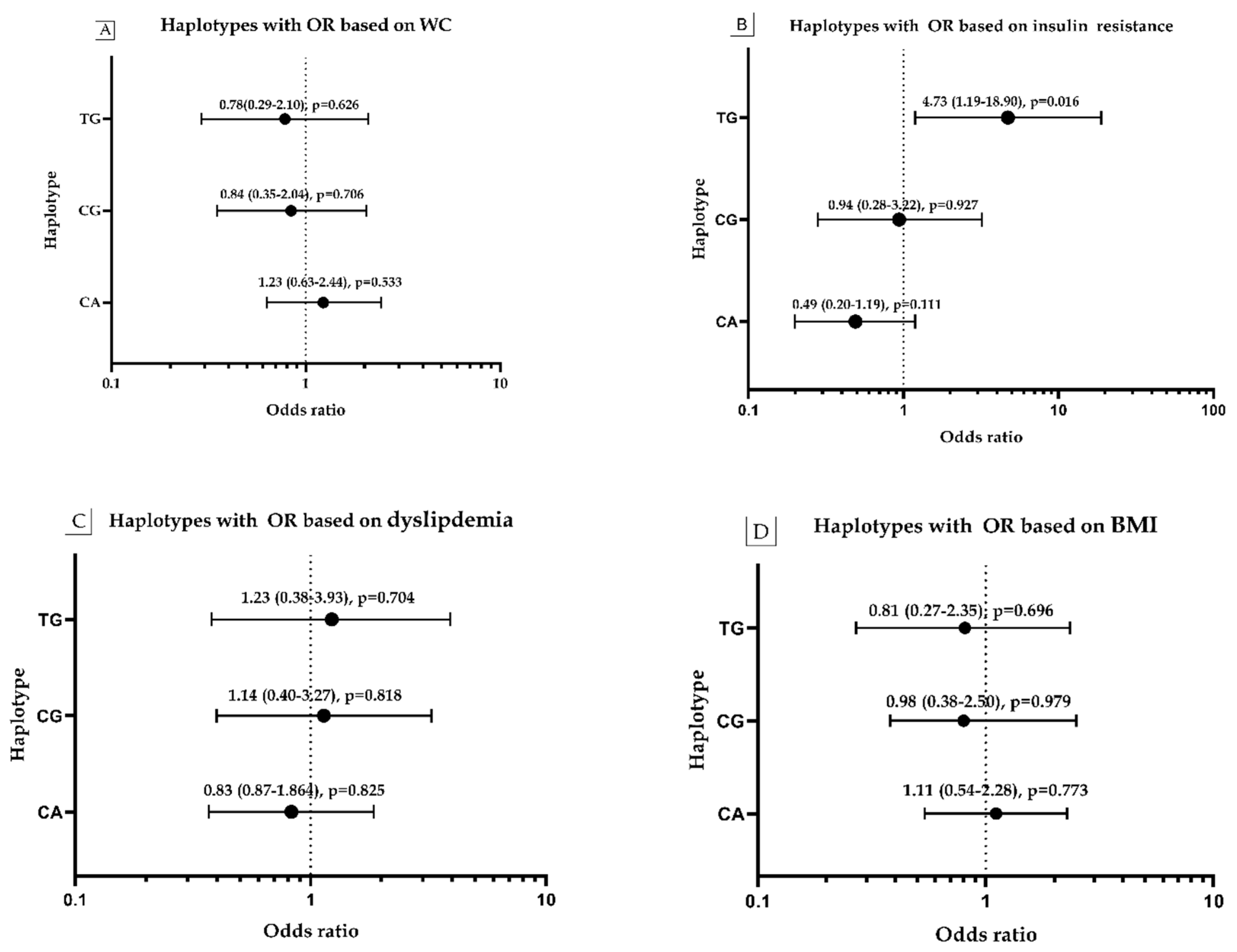
3.7. Haplotype Frequency and Its Association with Obesity, WC, HOMA and Dyslipidemia
3.8. The Expression of CD284 of TLR4 in Study Subjects by Flow Cytometry
3.9. Association between CD284 of TLR4 and the Genotype and Haplotype Distribution of Two SNPs (rs4986791 and rs4986790)
3.10. The Correlation between Haplotype of Two SNPs (rs4986791 and rs4986790) and IL6, IL10, and CD284 of Monocyte TLR4 Expression
4. Discussion
5. Plain Summary
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- WHO. Obesity and Overweight. 2016. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 6 August 2016).
- Poirier, P.; Giles, T.D.; Bray, G.A.; Hong, Y.; Stern, J.S.; Pi-Sunyer, F.X.; Eckel, R.H. Obesity and Cardiovascular Disease: Pathophysiology, Evaluation, and Effect of Weight Loss. Circulation 2006, 113, 898–918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hotamisligil, G.S. Inflammation and metabolic disorders. Nature 2006, 444, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Ringseis, R.; Eder, K.; Mooren, F.C.; Krüger, K. Metabolic signals and innate immune activation in obesity and exercise. Exerc. Immunol. Rev. 2015, 21, 58–68. [Google Scholar] [PubMed]
- Lorenzo, C.; Williams, K.; Hunt, K.J.; Haffner, S.M. The National Cholesterol Education Program–Adult Treatment Panel III, International Diabetes Federation, and World Health Organization Definitions of the Metabolic Syndrome as Predictors of Incident Cardiovascular Disease and Diabetes. Diabetes Care 2007, 30, 8–13. [Google Scholar] [CrossRef] [Green Version]
- Lteif, A.A.; Han, K.; Mather, K.J. Obesity, Insulin Resistance, and the Metabolic Syndrome. Circulation 2005, 112, 32–38. [Google Scholar] [CrossRef] [Green Version]
- Wilson, P.W.; D’Agostino, R.B.; Parise, H.; Sullivan, L.; Meigs, J.B. Metabolic Syndrome as a Precursor of Cardiovascular Disease and Type 2 Diabetes Mellitus. Circulation 2005, 112, 3066–3072. [Google Scholar] [CrossRef]
- Rock, F.L.; Hardiman, G.; Timans, J.C.; Kastelein, R.A.; Bazan, J.F. A family of human receptors structurally related to Drosophila Toll. Proc. Natl. Acad. Sci. USA 1998, 95, 588–593. [Google Scholar] [CrossRef] [Green Version]
- Kadowaki, N.; Ho, S.; Antonenko, S.; Malefyt, R.W.; Kastelein, R.A.; Bazan, F.; Liu, Y.J. Subsets of human dendritic cell precursors express different toll-like receptors and respond to different microbial antigens. J. Exp. Med. 2001, 194, 863–869. [Google Scholar] [CrossRef]
- Huang, S.; Rutkowsky, J.M.; Snodgrass, R.G.; Ono-Moore, K.D.; Schneider, D.A.; Newman, J.W.; Adams, S.H.; Hwang, D.H. Saturated fatty acids activate TLR-mediated proinflammatory signaling pathways. J. Lipid Res. 2012, 53, 2002–2013. [Google Scholar] [CrossRef] [Green Version]
- Rogero, M.M.; Calder, P.C. Obesity, Inflammation, Toll-Like Receptor 4 and Fatty Acids. Nutrients 2018, 10, 432. [Google Scholar] [CrossRef] [Green Version]
- Vaure, C.; Liu, Y.A. Comparative Review of Toll-Like Receptor 4 Expression and Functionality in Different Animal Species. Front. Immunol. 2014, 5, 316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patsouris, D.; Li, P.P.; Thapar, D.; Chapman, J.; Olefsky, J.M.; Neels, J.G. Ablation of CD11c-positive cells normalizes insulin sensitivity in obese insulin resistant animals. Cell Metab. 2008, 8, 301–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arbour, N.C.; Lorenz, E.; Schutte, B.C.; Zabner, J.; Kline, J.N.; Jones, M.; Frees, K.; Watt, J.L.; Schwartz, D.A. TLR4 mutations are associated with endotoxin hyporesponsiveness in humans. Nat. Genet. 2000, 25, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Ferwerda, B.; McCall, M.B.; Verheijen, K.; Kullberg, B.J.; van der Ven, A.J.; Van der Meer, J.W.; Netea, M.G. Functional consequences of toll-like receptor 4 polymorphisms. Mol. Med. 2008, 14, 346–352. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Gu, N.; Gao, Y.; Cen, W. TLR4 Asp299Gly (rs4986790) polymorphism and coronary artery disease: A meta-analysis. Peer J. 2015, 3, e1412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinhardt, A.P.; Aranguren, F.; Tellechea, M.L.; Gómez Rosso, L.A.; Brites, F.D.; Martínez-Larrad, M.T.; Serrano-Ríos, M.; Frechtel, G.D.; Taverna, M.J. A functional nonsynonymous toll-like receptor 4 gene polymorphism is associated with metabolic syndrome, surrogates of insulin resistance, and syndromes of lipid accumulation. Metabolism 2010, 59, 711–717. [Google Scholar] [CrossRef]
- Ferwerda, B.; McCall, M.B.; Alonso, S.; Giamarellos-Bourboulis, E.J.; Mouktaroudi, M.; Izagirre, N.; Syafruddin, D.; Kibiki, G.; Cristea, T.; Hijmans, A.; et al. TLR4 polymorphisms, infectious diseases, and evolutionary pressure during migration of modern humans. Proc. Natl. Acad. Sci. USA 2007, 104, 16645–16650. [Google Scholar] [CrossRef] [Green Version]
- Schneider, S.; Hoppmann, P.; Koch, W.; Kemmner, S.; Schmaderer, C.; Renders, L.; Kastrati, A.; Laugwitz, K.L.; Heemann, U.; Baumann, M. Obesity-associated hypertension is ameliorated in patients with TLR4 single nucleotide polymorphism (SNP) rs4986790. J. Inflamm. (Lond.) 2015, 12, 57. [Google Scholar] [CrossRef] [Green Version]
- Weyrich, P.; Staiger, H.; Stančáková, A.; Machicao, F.; Machann, J.; Schick, F.; Stefan, N.; Kuusisto, J.; Laakso, M.; Schäfer, S.; et al. The D299G/T399I Toll-like receptor 4 variant associates with body and liver fat: Results from the TULIP and METSIM Studies. PLoS ONE 2010, 5, e13980. [Google Scholar] [CrossRef] [Green Version]
- Rizk, N.M.; Yousef, M. Association of lipid profile and waist circumference as cardiovascular risk factors for overweight and obesity among school children in Qatar. Diabetes Metab. Syndr. Obes. 2012, 5, 425–432. [Google Scholar] [CrossRef] [Green Version]
- El-Menyar, A.; Rizk, N.M.; Asim, M.; Al-Thani, H.; Elgendy, A.; Al-Suwaidi, J. Association of β-Adrenergic Receptor Gene Polymorphisms with Acute Coronary Syndrome and Cardiovascular Risk Factors in an Arab Population. Angiology 2016, 67, 762–771. [Google Scholar] [CrossRef]
- Bocsi, J.; Melzer, S.; Dähnert, I.; Tárnok, A. OMIP-023: 10-Color, 13 antibody panel for in-depth phenotyping of human peripheral blood leukocytes. Cytom. Part A 2014, 85, 781–784. [Google Scholar] [CrossRef] [PubMed]
- Physical Status: The Use and Interpretation of Anthropometry. Report of a Who Expert Committee. World Health Organ. Tech. Rep. Ser. 1995, 854, 1–452.
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and β-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esteghamati, A.; Ashraf, H.; Khalilzadeh, O.; Zandieh, A.; Nakhjavani, M.; Rashidi, A.; Haghazali, M.; Asgari, F. Optimal cut-off of homeostasis model assessment of insulin resistance (HOMA-IR) for the diagnosis of metabolic syndrome: Third national surveillance of risk factors of non-communicable diseases in Iran (SuRFNCD-2007). Nutr. Metab. (Lond.) 2010, 7, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, Y.; Garg, M.K.; Tandon, N.; Marwaha, R.K. A study of insulin resistance by HOMA-IR and its cut-off value to identify metabolic syndrome in urban Indian adolescents. J. Clin. Res. Pediatr. Endocrinol. 2013, 5, 245–251. [Google Scholar] [PubMed]
- Santos, J.L.; Lera, L.; Pérez-Bravo, F.; Albala, C. Adiposity and bone mineral density of Chilean elderly women in relation to toll-like receptor 4 gene polymorphisms. Ann. Hum. Biol. 2006, 33, 585–592. [Google Scholar] [CrossRef]
- Jialal, I.; Kaur, H.; Devaraj, S. Toll-like receptor status in obesity and metabolic syndrome: A translational perspective. J. Clin. Endocrinol. Metab. 2014, 99, 39–48. [Google Scholar] [CrossRef]
- Portela-Cidade, J.P.; Borges-Canha, M.; Leite-Moreira, A.F.; Pimentel-Nunes, P. Systematic Review of the Relation Between Intestinal Microbiota and Toll-Like Receptors in the Metabolic Syndrome: What Do We Know So Far? GE Port. J. Gastroenterol. 2015, 22, 240–258. [Google Scholar] [CrossRef] [Green Version]
- Fu, X.D.; Sun, X.Q.; Wang, H.Y.; Guo, J.F.; Zhang, G.Z.; Xu, Q.H. Genetic polymorphisms of the TLR4 gene and their association with susceptibility to type 2 diabetes mellitus in the Chinese population. Genet. Mol. Res. GMR 2013, 12, 3813–3820. [Google Scholar] [CrossRef]
- Zhu, S.L.; Wu, Q.H.; Tu, J. Study on regulation of NLRP3/SOCS3-TLR4-NF-kappaB inflammatory pathway by wogonoside to improve hepatic insulin resistance. Zhongguo Zhong Yao Za Zhi 2019, 44, 4504–4510. [Google Scholar] [PubMed]
- Ao, R.; Wang, Y.; Zhnag, D.R.; Du, Y.Q. Role of TLR4 rs4986790A>G and rs4986791C>T Polymorphisms in the Risk of Inflammatory Bowel Disease. Gastroenterol. Res. Pract. 2015, 2015, 141070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Li, M.; Haghiac, M.; Catalano, P.M.; O’Tierney-Ginn, P.; Hauguel-de Mouzon, S. Causal relationship between obesity-related traits and TLR4-driven responses at the maternal-fetal interface. Diabetologia 2016, 59, 2459–2466. [Google Scholar] [CrossRef] [Green Version]
- Penas-Steinhardt, A.; Barcos, L.S.; Belforte, F.S.; de Sereday, M.; Vilariño, J.; Gonzalez, C.D.; Martínez-Larrad, M.T.; Tellechea, M.L.; Serrano-Ríos, M. Functional characterization of TLR4 +3725 G/C polymorphism and association with protection against overweight. PLoS ONE 2012, 7, e50992. [Google Scholar] [CrossRef] [PubMed]
- Bagarolli, R.A.; Saad, M.J.A.; Saad, S.T.O. Toll-like receptor 4 and inducible nitric oxide synthase gene polymorphisms are associated with Type 2 diabetes. J. Diabetes Complicat. 2010, 24, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Kolz, M.; Baumert, J.; Müller, M.; Khuseyinova, N.; Klopp, N.; Thorand, B.; Meisinger, C.; Herder, C.; Koenig, W.; Illig, T. Association between variations in the TLR4 gene and incident type 2 diabetes is modified by the ratio of total cholesterol to HDL-cholesterol. BMC Med. Genet. 2008, 9, 9. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, R.; Al-Mass, A.; Atizado, V.; Al-Hubail, A.; Al-Ghimlas, F.; Al-Arouj, M.; Bennakhi, A.; Dermime, S.; Behbehani, K. Elevated expression of the toll like receptors 2 and 4 in obese individuals: Its significance for obesity-induced inflammation. J. Inflamm. 2012, 9, 48. [Google Scholar] [CrossRef] [Green Version]
- Song, M.J.; Kim, K.H.; Yoon, J.M.; Kim, J.B. Activation of Toll-like receptor 4 is associated with insulin resistance in adipocytes. Biochem. Biophys. Res. Commun. 2006, 346, 739–745. [Google Scholar] [CrossRef]
- Zhang, H.M.; Chen, L.L.; Wang, L.; Xu, S.; Wang, X.; Yi, L.L.; Chen, D.; Wu, Z.H.; Zhang, J.Y.; Liao, Y.F.; et al. Macrophage infiltrates with high levels of Toll-like receptor 4 expression in white adipose tissues of male Chinese. Nutr. Metab. Cardiovasc. Dis. 2009, 19, 736–743. [Google Scholar] [CrossRef]
- Radin, M.S.; Sinha, S.; Bhatt, B.A.; Dedousis, N.; O’Doherty, R.M. Inhibition or deletion of the lipopolysaccharide receptor Toll-like receptor-4 confers partial protection against lipid-induced insulin resistance in rodent skeletal muscle. Diabetologia 2008, 51, 336–346. [Google Scholar] [CrossRef] [Green Version]
- Davis, J.E.; Gabler, N.K.; Walker-Daniels, J.; Spurlock, M.E. Tlr-4 deficiency selectively protects against obesity induced by diets high in saturated fat. Obesity (Silver Spring) 2008, 16, 1248–1255. [Google Scholar] [CrossRef] [PubMed]
- Velloso, L.A.; Folli, F.; Saad, M.J. TLR4 at the Crossroads of Nutrients, Gut Microbiota, and Metabolic Inflammation. Endocr. Rev. 2015, 36, 245–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitseva, O.I.; Tanriverdi, K.; Tchkonia, T.T.; Kirkland, J.L.; McDonnell, M.E.; Apovian, C.M.; Freedman, J.; Gokce, N. Inducible Toll-like receptor and NF-kappaB regulatory pathway expression in human adipose tissue. Obesity (Silver Spring) 2008, 16, 932–937. [Google Scholar] [CrossRef]
- De Loera-Rodriguez, C.O.; Delgado-Rizo, V.; Alvarado-Navarro, A.; Agraz-Cibrian, J.M.; Segura-Ortega, J.E.; Fafutis-Morris, M. Over-expression of TLR4-CD14, pro-inflammatory cytokines, metabolic markers and NEFAs in obese non-diabetic Mexicans. J. Inflamm. (Lond.) 2014, 11, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimabukuro, M. Leptin Resistance and Lipolysis of White Adipose Tissue: An Implication to Ectopic Fat Disposition and Its Consequences. J. Atheroscler. Thromb. 2017, 24, 1088–1089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, J.; Peng, Y.; Wu, J.; Wang, Y.; Yao, L. Toll-like receptor 2/4 links to free fatty acid-induced inflammation and β-cell dysfunction. J. Leukoc. Biol. 2014, 95, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Long, H.; O’Connor, B.P.; Zemans, R.L.; Zhou, X.; Yang, I.V.; Schwartz, D.A. The Toll-Like Receptor 4 Polymorphism Asp299Gly but Not Thr399Ile Influences TLR4 Signaling and Function. PLoS ONE 2014, 9, e93550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jialal, I.; Huet, B.A.; Kaur, H.; Chien, A.; Devaraj, S. Increased toll-like receptor activity in patients with metabolic syndrome. Diabetes Care 2012, 35, 900–904. [Google Scholar] [CrossRef] [Green Version]
- Martínez-García, M.Á.; Ojeda-Ojeda, M.; Rodríguez-Martín, E.; Insenser, M.; Moncayo, S.; Álvarez-Blasco, F.; Luque-Ramírez, M.; Escobar-Morreale, H.F. TLR2 and TLR4 Surface and Gene Expression in White Blood Cells after Fasting and Oral Glucose, Lipid and Protein Challenges: Influence of Obesity and Sex Hormones. Biomolecules 2020, 10, 111. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | NON-Obese (n = 132) | Obese (n = 66) | p-Value |
|---|---|---|---|
| Age (years) | 22.04 ± 3.39 | 21.45 ± 2.67 | 0.215 |
| BMI (kg/m2) | 22.72 ± 3.60 | 35.65 ± 4.01 | <0.0001 |
| WC (cm) | 80.26 ± 9.01 | 102.88 ± 8.22 | <0.0001 |
| %BF | 33.91 ± 7.04 | 48.06 ± 3.43 | <0.0001 |
| Systolic blood pressure (mmHg) | 114.24 ± 6.10 | 121.27 ± 8.40 | 0.785 |
| Diastolic blood pressure (mmHg) | 72.25 ± 4.45 | 78.55 ± 7.85 | 0.425 |
| Glucose (mmol/L) | 4.95 ± 0.43 | 5.06 ± 0.71 | 0.251 |
| TC (mmol/L) | 3.95 ± 0.26 | 4.03 ± 0.93 | 0.032 |
| TG (mmol/L) | 0.79 ± 0.37 | 0.93 ± 0.68 | 0.121 |
| HDL-C (mmol/L) | 1.37 ± 0.36 | 0.90 ± 0.39 | <0.0001 |
| LDL-C (mmol/L) | 2.35 ± 0.80 | 2.19 ± 1.59 | 0.386 |
| hsCRP (pg/mL) | 1.19 (0.21–4.01) | 4.94 (1.66–7.78) | 0.001 |
| Insulin (μU/L) | 10.68 (4.71–15.54) | 16.05 (5.50–19.25) | <0.0001 |
| HOMA (insulin resistance) (IR) | 1.83 (1.08–3.53) | 3.67 (1.87–4.74) | <0.0001 |
| Leptin (ng/mL) | 8.19 (3.36–13.12) | 20.25 (17.18–37.82) | 0.038 |
| SNP | Genotype | All Subjects (n = 197) | NON-Obese (n = 131) | Obese (n = 66) | p-Value |
| rs4986791 (+1196 C > T) Thr399Ile | CC | 178 (90.4%) | 117 (89.3%) | 61 (92.4%) | 0.485 |
| CT | 19 (9.6%) | 14 (10.7%) | 5 (7.6%) | ||
| Minor allele frequency (T) | 0.048 | 0.053 | 0.038 | 0.493 | |
| SNP | Genotype | All Subjects (n = 198) | NON-Obese (n = 132) | Obese (n = 66) | p-Value |
| rs4986790 (+896 A > G) Asp299Gly | AA | 173 (87.4%) | 117 (88.4%) | 56 (86.4%) | 0.452 |
| AG | 12 (6%) | 4 (4.2%) | 8 (7.8%) | ||
| GG | 13 (6.6%) | 11(7.4%) | 2 (5.8%) | ||
| Minor allele frequency (G) | 0.096 | 0.098 | 0.090 | 0.809 | |
| Variables | Non-Obese (n = 21) | Obese (n = 21) | p-Value |
|---|---|---|---|
| TLR4 Mean Fluorescent Intensity (MFI) | 2536.00 ± 641.60 | 3444.00 ± 461.30 | <0.0001 |
| IL-6 (pg/mL) | 1.31 ± 0.35 | 12.42 ± 3.41 | <0.0001 |
| IL-10 (pg/mL) | 3.91 ± 0.64 | 2.56 ± 0.52 | 0.045 |
| SNP | Genotype | All Subjects (n = 42) | Non-Obese (n = 21) | Obese (n = 21) | p-Value |
| rs4986791 (+1196C > T) Thr399Ile | CC | 38(90.5%) | 19(90.5%) | 19 (90.5%) | 1.000 |
| CT | 4 (9.5%) | 2 (10.5%) | 2 (10.5%) | ||
| SNP | Genotype | All Subjects (n = 42) | Non-Obese (n = 21) | Obese (n = 21) | p-Value |
| rs4986790 (+896A > G) Asp299Gly | AA | 35 (83.3%) | 17 (80.9%) | 18 (85.7%) | 0.506 |
| AG | 3 (7.1%) | 1 (4.8%) | 2 (9.5%) | ||
| GG | 4 (9.5%) | 3 (14.3%) | 1 (4.8%) | ||
| Haplotype | All Subjects (n = 42) | Non-Obese (n = 21) | Obese (n = 21) | p-Value | |
| rs4986790 (+896A > G) Asp299Gly | CA | 35 (83.3%) | 17 (80.9%) | 18 (85.7%) | 0.834 |
| CG | 3 (7.1%) | 2 (9.5%) | 1(4.8%) | ||
| TG | 4 (9.5%) | 2 (9.5%) | 2(9.5%) |
| Genotype | Monocyte CDTLR4 Expression | ||
|---|---|---|---|
| rs4986791 | Low expression group (≥25%) (n = 11) | High expression group (≥75%) (n = 10) | p-Value |
| CC | 10 (90.9%) | 8 (80.0%) | 0.486 |
| CT | 1(9.1%) | 2 (20.0%) | |
| rs4986790 | |||
| AA | 9 (81.8%) | 7 (70.0%) | 0.281 |
| AG | 0 (0.0%) | 2 (20.0%) | |
| GG | 2 (18.2%) | 1 (10.0%) | |
| Haplotype rs4986790 & rs4986790 | Low expression group (≥25%) (n = 11) | High expression group (≥75%) (n = 10) | p-Value |
| CA | 9 (81.8%) | 7 (70.0%) | 0.765 |
| AG | 1 (9.1%) | 1 (10.0%) | |
| TG | 1 (9.1%) | 2 (20.0%) | |
| IL-6 | IL-10 | TLR4 | Haplotype | |
|---|---|---|---|---|
| Variables | r p | r p | r p | r p |
| IL-6 | 1 | −0.675 <0.0001 | 0.595 <0.0001 | 0.036 0.819 |
| IL-10 | −0.675 <0.0001 | 1 | −0.654 <0.0001 | 0.009 0.095 |
| TLR4 | 0.595 <0.0001 | −0.654 <0.0001 | 1 | 0.324 0.0598 |
| Haplotype | 0.036 0.819 | 0.009 0.095 | 0.324 0.0598 | 1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharif, E.; Al-Wakeel, M.; Mohamed, A.; kerkadi, A.; Rizk, N. TLR4 Receptor D299G/T399I Haplotype Polymorphism Is Associated with Insulin Resistance in Obese Female Subjects. Genes 2020, 11, 814. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11070814
Sharif E, Al-Wakeel M, Mohamed A, kerkadi A, Rizk N. TLR4 Receptor D299G/T399I Haplotype Polymorphism Is Associated with Insulin Resistance in Obese Female Subjects. Genes. 2020; 11(7):814. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11070814
Chicago/Turabian StyleSharif, Elham, Mariam Al-Wakeel, Afnan Mohamed, Abdelhamid kerkadi, and Nasser Rizk. 2020. "TLR4 Receptor D299G/T399I Haplotype Polymorphism Is Associated with Insulin Resistance in Obese Female Subjects" Genes 11, no. 7: 814. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11070814