Variation in the Caprine Keratin-Associated Protein 27-1 Gene is Associated with Cashmere Fiber Diameter

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cashmere Goats and Sample Collection
2.2. Polymerase Chain Reaction–Single Strand Conformation Polymorphism (SSCP) Analysis of Caprine KRTAP27-1
2.3. Sequencing of KRTAP27-1 Variants and Sequence Analyses
2.4. Reverse-Transcription Polymerase Chain Reaction (RT-PCR) Analysis
2.5. Statistical Analyses
3. Results
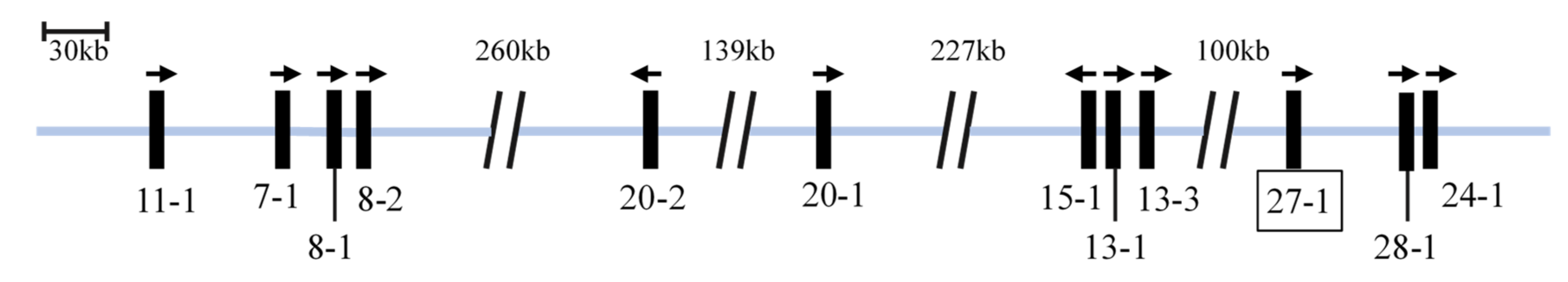
3.1. Identification and Chromosomal Location of Caprine KRTAP27-1
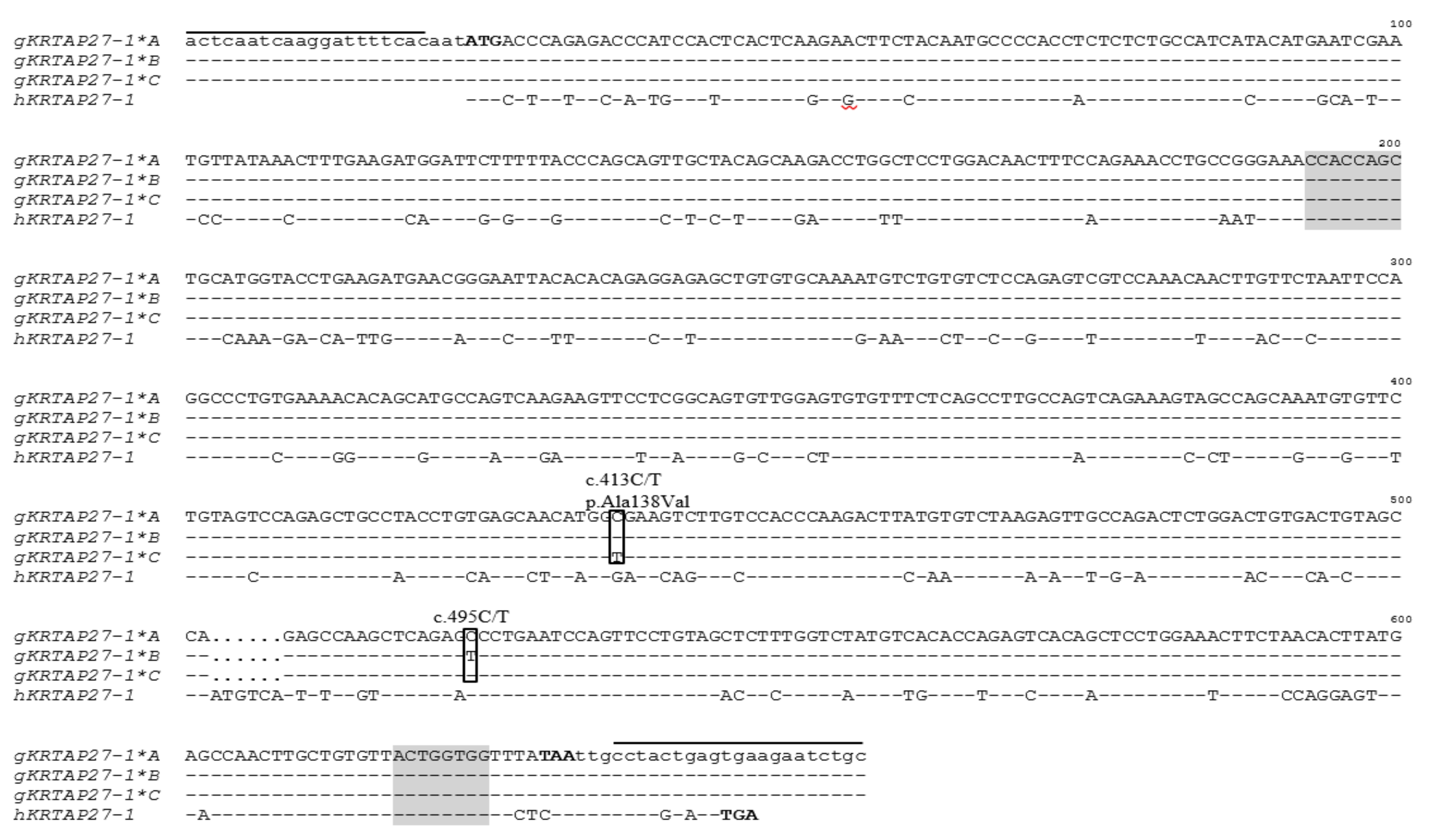
3.2. Sequence Variation in Caprine KRTAP27-1
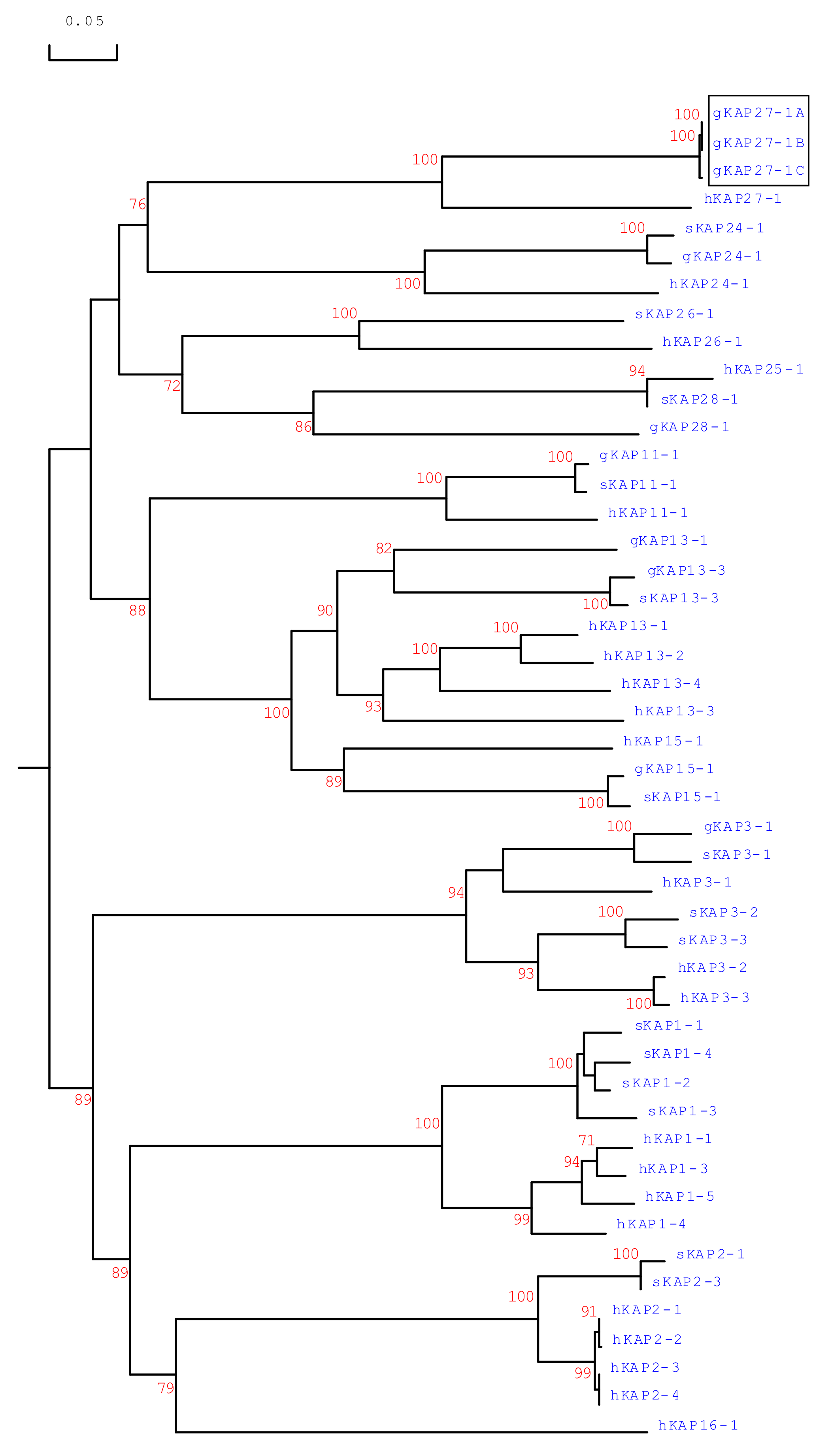
3.3. Phylogenetic Analysis of Caprine KAP27-1
3.4. Expression of Caprine KRTAP27-1
3.5. Correlation between Cashmere Fiber Traits
3.6. Association between KRTAP27-1 Genotypes and Cashmere Fiber Traits
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Waldron, S.; Brown, C.; Komarek, A.M. The Chinese cashmere industry: A global value chain analysis. Dev. Policy Rev. 2014, 32, 589–610. [Google Scholar] [CrossRef]
- Gong, H.; Zhou, H.; Forrest, R.H.; Li, S.; Wang, J.; Dyer, J.M.; Luo, Y.; Hickford, J.G. Wool keratin-associated protein genes in sheep-a review. Genes 2016, 7, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, L.; Wang, J.; Zhou, H.; Gong, H.; Tao, J.; Hickford, J.G. Identification of ovine KRTAP28-1 and its association with wool weight and mean fibre diameter-associated traits. Animals 2019, 9, 142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, H.; Zhou, H.; Wang, J.; Li, S.; Luo, Y.; Hickford, J.G. Characterisation of an ovine keratin associated protein (KAP) gene, which would produce a protein rich in glycine and tyrosine, but lacking in cysteine. Genes 2019, 10, 848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, M.; Zhou, H.; Hickford, J.G.; Gong, H.; Wang, J.; Hu, J.; Liu, X.; Li, S.; Hao, Z.; Luo, Y. Variation in the caprine keratin-associated protein 15-1 (KAP15-1) gene affects cashmere fibre diameter. Arch. Anim. Breed. 2019, 62, 125–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Zhou, H.; Luo, Y.; Zhao, M.; Gong, H.; Hao, Z.; Hu, J.; Hickford, J.G. Variation in the caprine KAP24-1 gene affects cashmere fibre diameter. Animals 2019, 9, 15. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Zhou, H.; Hickford, J.G.; Zhao, M.; Gong, H.; Hao, Z.; Shen, J.; Hu, J.; Liu, X.; Li, S.; et al. Identification of caprine KRTAP28-1 and its effect on Cashmere fiber diameter. Genes 2020, 11, 121. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Li, N.; Jia, C.; Zhu, X.; Jia, Z. Effect of the polymorphisms of keratin associated protein 8.2 gene on fibre traits in Inner Mongolia cashmere goats. Asian Australas. J. Anim. Sci. 2007, 20, 821–826. [Google Scholar] [CrossRef]
- Wang, J.; Hao, Z.; Zhou, H.; Luo, Y.; Hu, J.; Liu, X.; Li, S.; Hickford, J.G. A keratin-associated protein (KAP) gene that is associated with variation in cashmere goat fleece weight. Small Rumin. Res. 2018, 167, 104–109. [Google Scholar] [CrossRef]
- Wang, J.; Che, L.; Hickford, J.G.; Zhou, H.; Hao, Z.; Luo, Y.; Hu, J.; Liu, X.; Li, S. Identification of the caprine keratin-associated protein 20-2 (KAP20-2) gene and its effect on cashmere traits. Genes 2017, 8, 328. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Zhou, H.; Gong, H.; Zhao, F.; Hu, J.; Luo, Y.; Hickford, J.G. Identification of the ovine keratin-associated protein 26-1 gene and its association with variation in wool traits. Genes 2017, 8, 225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Zhou, H.; Gong, H.; Zhao, F.; Wang, J.; Liu, X.; Luo, Y.; Hickford, J.G. Identification of the ovine keratin-associated protein 22-1 (KAP22-1) gene and its effect on wool traits. Genes 2017, 8, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogers, M.A.; Winter, H.; Langbein, L.; Wollschläger, A.; Praetzel-Wunder, S.; Jave-Suarez, L.F.; Schweizer, J. Characterization of human KAP24.1, a cuticular hair keratin-associated protein with unusual amino-acid composition and repeat structure. J. Invest. Dermatol. 2007, 127, 1197–1204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, H.; Hickford, J.G.; Fang, Q. A two-step procedure for extracting genomic DNA from dried blood spots on filter paper for polymerase chain reaction amplification. Anal. Biochem. 2006, 354, 159–161. [Google Scholar] [CrossRef] [PubMed]
- Byun, S.O.; Fang, Q.; Zhou, H.; Hickford, J.G. An effective method for silver-staining DNA in large numbers of polyacrylamide gels. Anal. Biochem. 2009, 385, 174–175. [Google Scholar] [CrossRef]
- Gong, H.; Zhou, H.; Hickford, J.G. Diversity of the glycine/tyrosine-rich keratin-associated protein 6 gene (KAP6) family in sheep. Mol. Biol. Rep. 2011, 38, 31–35. [Google Scholar] [CrossRef]
- Gong, H.; Zhou, H.; McKenzie, G.W.; Yu, Z.; Clerens, S.; Dyer, J.M.; Plowman, J.E.; Wright, M.W.; Arora, R.; Bawden, C.S.; et al. An updated nomenclature for keratin-associated proteins (KAPs). Int. J. Biol. Sci. 2012, 8, 258–264. [Google Scholar] [CrossRef] [Green Version]
- Tørresen, O.K.; Star, B.; Mier, P.; Andrade-Navarro, M.A.; Bateman, A.; Jarnot, P.; Gruca, A.; Grynberg, M.; Kajava, A.V.; Promponas, V.J.; et al. Tandem repeats lead to sequence assembly errors and impose multi-level challenges for genome and protein databases. Nucleic Acids Res. 2019, 47, 10994–11006. [Google Scholar] [CrossRef]
- Li, R.; Fu, W.; Su, R.; Tian, X.; Du, D.; Zhao, Y.; Zheng, Z.; Chen, Q.; Gao, S.; Cai, Y.; et al. Towards the complete goat pan-genome by recovering missing genomic segments from the reference genome. Front. Genet. 2019, 10, 1169. [Google Scholar] [CrossRef]
- Gong, H.; Zhou, H.; Dyer, J.M.; Plowman, J.E.; Hickford, J.G. Identification of the keratin-associated protein 13-3 (KAP13-3) gene in sheep. Open J. Genet. 2011, 1, 60–64. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Gong, H.; Yan, W.; Luo, Y.; Hickford, J.G. Identification and sequence analysis of the keratin-associated protein 24-1 (KAP24-1) gene homologue in sheep. Gene 2012, 511, 62–65. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.; Zhou, H.; Dyer, J.M.; Hickford, J.G. Identification of the ovine KAP11-1 gene (KRTAP11-1) and genetic variation in its coding sequence. Mol. Biol. Rep. 2011, 38, 5429–5433. [Google Scholar] [CrossRef] [PubMed]
- Ku, N.O.; Liao, J.; Chou, C.F.; Omary, M.B. Implications of intermediate filament protein phosphorylation. Cancer Metastasis Rev. 1996, 15, 429–444. [Google Scholar] [CrossRef] [PubMed]
- Rogers, M.A.; Langbein, L.; Praetzel-Wunder, S.; Giehl, K. Characterization and expression analysis of the hair keratin associated protein KAP26.1. Br. J. Dermatol. 2008, 159, 725–729. [Google Scholar] [CrossRef] [PubMed]
- Kijas, J.W.; Townley, D.; Dalrymple, B.P.; Heaton, M.P.; Maddox, J.F.; McGrath, A.; Wilson, P.; Ingersoll, R.G.; McCulloch, R.; McWilliam, S.; et al. International Sheep Genomics Consortium. A genome wide survey of SNP variation reveals the genetic structure of sheep breeds. PLoS ONE 2009, 4, e4668. [Google Scholar] [CrossRef] [Green Version]
- Itenge-Mweza, T.O.; Forrest, R.H.; McKenzie, G.W.; Hogan, A.; Abbott, J.; Amoafo, O.; Hickford, J.G. Polymorphism of the KAP1.1, KAP1.3 and K33 genes in Merino sheep. Mol. Cell. Probes 2007, 21, 338–342. [Google Scholar] [CrossRef]
- Gong, H.; Zhou, H.; Hickford, J.G. Polymorphism of the ovine keratin-associated protein 1-4 gene (KRTAP1-4). Mol. Biol. Rep. 2010, 37, 3377–3380. [Google Scholar] [CrossRef]
- Rogers, G.R.; Hickford, J.G.; Bickerstaffe, R. Polymorphism in two genes for B2 high sulfur proteins of wool. Anim. Genet. 1994, 25, 407–415. [Google Scholar] [CrossRef]
- Zhou, H.; Visnovska, T.; Gong, H.; Schmeier, S.; Hickford, J.G.; Ganley, A.R. Contrasting patterns of coding and flanking region evolution in mammalian keratin associated protein-1 genes. Mol. Phylogenet. Evol. 2019, 133, 352–361. [Google Scholar] [CrossRef]
- Wang, J.; Zhou, H.; Zhu, J.; Hu, J.; Liu, X.; Li, S.; Luo, Y.; Hickford, J.G. Identification of the ovine keratin-associated protein 15-1 gene (KRTAP15-1) and genetic variation in its coding sequence. Small Rumin. Res. 2017, 153, 131–136. [Google Scholar] [CrossRef]
- Duan, J.; Wainwright, M.S.; Comeron, J.M.; Saitou, N.; Sanders, A.R.; Gelernter, J.; Gejman, P.V. Synonymous mutations in the human dopamine receptor D2 (DRD2) affect mRNA stability and synthesis of the receptor. Hum. Mol. Genet. 2003, 12, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Nackley, A.G.; Shabalina, S.A.; Tchivileva, I.E.; Satterfield, K.; Korchynskyi, O.; Makarov, S.S.; Maixner, W.; Diatchenko, L. Human catechol-O-methyltransferase haplotypes modulate protein expression by altering mRNA secondary structure. Science 2006, 314, 1930–1933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bigham, M.L.; Morris, C.A.; Southey, B.R.; Baker, R.L. Heritabilities and genetic correlations for liveweight and fibre traits in New Zealand Cashmere goats. Lives. Prod. Sci. 1993, 33, 91–104. [Google Scholar] [CrossRef]
- Gifford, D.R.; Ponzoni, R.W. Recent developments in cashmere breeding in Australia and New Zealand. Proc. Aust. Soc. Anim. Prod. 1990, 18, 77–79. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fiber Trait | Raw Mean ± SE (n = 248) | (Mean ± SE) 1 | p Value | ||
|---|---|---|---|---|---|
| AA (n = 129) | AB (n = 81) | BB (n = 14) | |||
| Cashmere fiber yield (g) | 397.5 ± 3.18 | 420.4 ± 4.88 | 417.0 ± 5.75 | 412.3 ± 11.45 | 0.721 |
| Mean fiber diameter (μm) | 13.3 ± 0.03 | 13.4 ± 0.05 a | 13.5 ± 0.06 b | 13.7 ± 0.11 b | 0.026 |
| Curly fiber length (cm) | 4.1 ± 0.03 | 4.4 ± 0.05 | 4.3 ± 0.06 | 4.2 ± 0.12 | 0.127 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, M.; Zhou, H.; Luo, Y.; Wang, J.; Hu, J.; Liu, X.; Li, S.; Hao, Z.; Jin, X.; Song, Y.; et al. Variation in the Caprine Keratin-Associated Protein 27-1 Gene is Associated with Cashmere Fiber Diameter. Genes 2020, 11, 934. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11080934
Zhao M, Zhou H, Luo Y, Wang J, Hu J, Liu X, Li S, Hao Z, Jin X, Song Y, et al. Variation in the Caprine Keratin-Associated Protein 27-1 Gene is Associated with Cashmere Fiber Diameter. Genes. 2020; 11(8):934. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11080934
Chicago/Turabian StyleZhao, Mengli, Huitong Zhou, Yuzhu Luo, Jiqing Wang, Jiang Hu, Xiu Liu, Shaobin Li, Zhiyun Hao, Xiayang Jin, Yize Song, and et al. 2020. "Variation in the Caprine Keratin-Associated Protein 27-1 Gene is Associated with Cashmere Fiber Diameter" Genes 11, no. 8: 934. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11080934