Elevated Expression of SLC6A4 Encoding the Serotonin Transporter (SERT) in Gilles de la Tourette Syndrome

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. GTS and Control Cohort
2.2. Genotyping
2.3. Expression Analysis
2.4. Methylation Analysis
2.5. Statistical Analyses
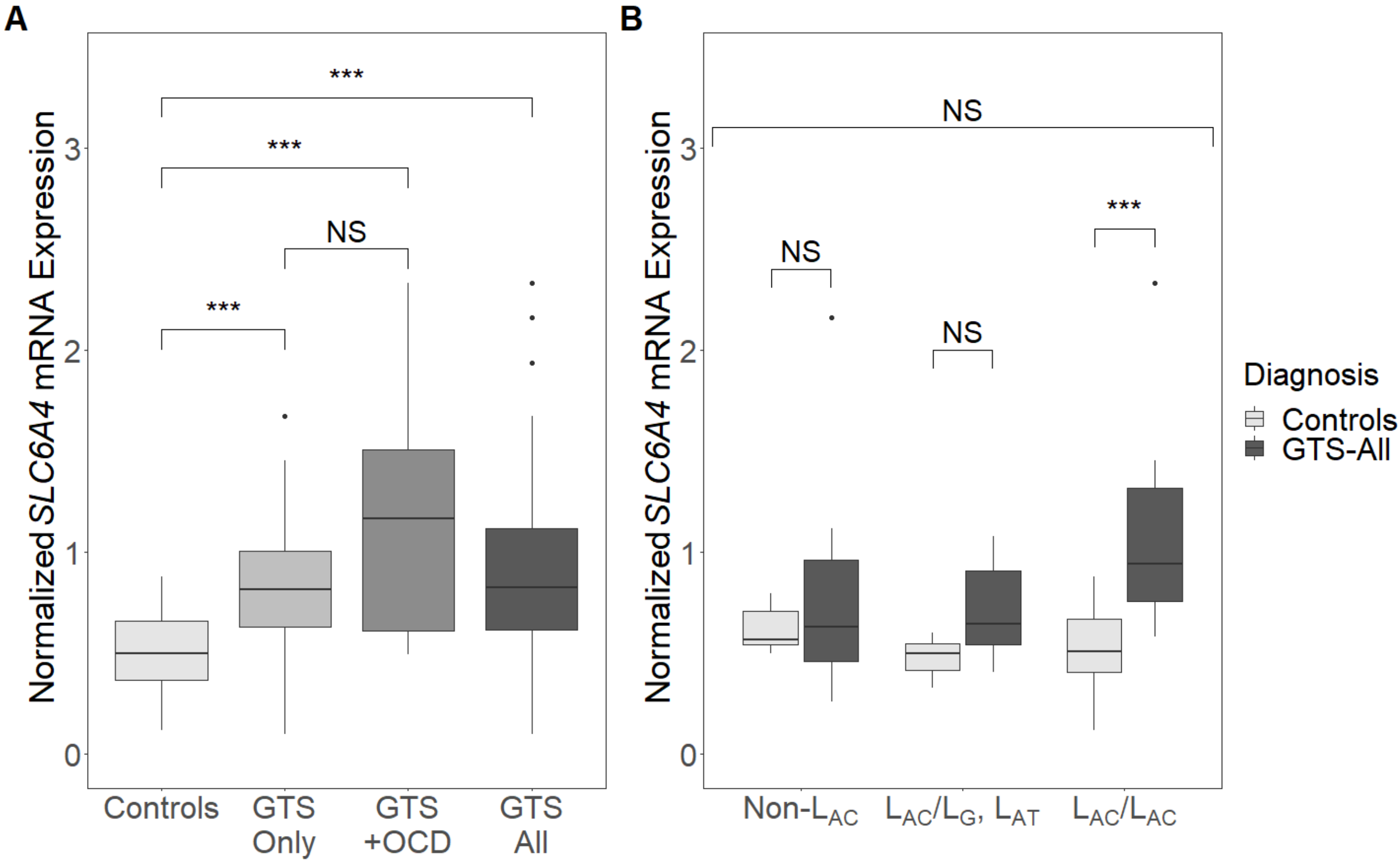
3. Results
3.1. Genotyping
3.2. Expression Analysis
3.3. Methylation Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hirschtritt, M.E.; Lee, P.C.; Pauls, D.L.; Dion, Y.; Grados, M.A.; Illmann, C.; King, R.A.; Sandor, P.; McMahon, W.M.; Lyon, G.J.; et al. Lifetime prevalence, age of risk, and genetic relationships of comorbid psychiatric disorders in tourette syndrome. JAMA Psychiatry 2015. [Google Scholar] [CrossRef]
- Burd, L.; Li, Q.; Kerbeshian, J.; Klug, M.G.; Freeman, R.D. Tourette syndrome and comorbid pervasive developmental disorders. J. Child Neurol. 2009. [Google Scholar] [CrossRef]
- Mataix-Cols, D.; Isomura, K.; Pérez-Vigil, A.; Chang, Z.; Rück, C.; Johan Larsson, K.; Leckman, J.F.; Serlachius, E.; Larsson, H.; Lichtenstein, P. Familial risks of tourette syndrome and chronic tic disorders a population-based cohort study. JAMA Psychiatry 2015. [Google Scholar] [CrossRef] [Green Version]
- Davis, L.K.; Yu, D.; Keenan, C.L.; Gamazon, E.R.; Konkashbaev, A.I.; Derks, E.M.; Neale, B.M.; Yang, J.; Lee, S.H.; Evans, P.; et al. Partitioning the Heritability of Tourette Syndrome and Obsessive Compulsive Disorder Reveals Differences in Genetic Architecture. PLoS Genet. 2013. [Google Scholar] [CrossRef] [Green Version]
- Bertelsen, B.; Stefánsson, H.; Riff Jensen, L.; Melchior, L.; Mol Debes, N.; Groth, C.; Skov, L.; Werge, T.; Karagiannidis, I.; Tarnok, Z.; et al. Association of AADAC Deletion and Gilles de la Tourette Syndrome in a Large European Cohort. Biol. Psychiatry 2016. [Google Scholar] [CrossRef]
- Yu, D.; Sul, J.H.; Tsetsos, F.; Nawaz, M.S.; Huang, A.Y.; Zelaya, I.; Illmann, C.; Osiecki, L.; Darrow, S.M.; Hirschtritt, M.E.; et al. Interrogating the genetic determinants of Tourette’s syndrome and other tiC disorders through genome-wide association studies. Am. J. Psychiatry 2019. [Google Scholar] [CrossRef]
- Huang, A.Y.; Yu, D.; Davis, L.K.; Sul, J.H.; Tsetsos, F.; Ramensky, V.; Zelaya, I.; Ramos, E.M.; Osiecki, L.; Chen, J.A.; et al. Rare Copy Number Variants in NRXN1 and CNTN6 Increase Risk for Tourette Syndrome. Neuron 2017. [Google Scholar] [CrossRef] [Green Version]
- Scharf, J.M.; Yu, D.; Mathews, C.A.; Neale, B.M.; Stewart, S.E.; Fagerness, J.A.; Evans, P.; Gamazon, E.; Edlund, C.K.; Service, S.K.; et al. Genome-wide association study of Tourette’s syndrome. Mol. Psychiatry 2013. [Google Scholar] [CrossRef] [Green Version]
- Draper, A.; Stephenson, M.C.; Jackson, G.M.; Pépés, S.; Morgan, P.S.; Morris, P.G.; Jackson, S.R. Increased GABA contributes to enhanced control over motor excitability in tourette syndrome. Curr. Biol. 2014. [Google Scholar] [CrossRef] [Green Version]
- Nordstrom, E.J.; Bittner, K.C.; McGrath, M.J.; Parks, C.R.; Burton, F.H. Hyperglutamatergic cortico-striato-thalamo-cortical circuit breaker drugs alleviate tics in a transgenic circuit model of Tourette’s syndrome. Brain Res. 2015. [Google Scholar] [CrossRef] [Green Version]
- Kanaan, A.S.; Gerasch, S.; García-García, I.; Lampe, L.; Pampel, A.; Anwander, A.; Near, J.; Möller, H.E.; Müller-Vahl, K. Pathological glutamatergic neurotransmission in Gilles de la Tourette syndrome. Brain 2017. [Google Scholar] [CrossRef] [Green Version]
- Herzberg, I.; Valencia-Duarte, A.V.; Kay, V.A.; White, D.J.; Müller, H.; Rivas, I.C.; Mesa, S.C.; Cuartas, M.; García, J.; Bedoya, G.; et al. Association of DRD2 variants and Gilles de la Tourette syndrome in a family-based sample from a South American population isolate. Psychiatr. Genet. 2010. [Google Scholar] [CrossRef]
- Lee, C.C.; Chou, I.C.; Tsai, C.H.; Wang, T.R.; Li, T.C.; Tsai, F.J. Dopamine receptor D2 gene polymorphisms are associated in Taiwanese children with Tourette syndrome. Pediatr. Neurol. 2005. [Google Scholar] [CrossRef]
- Liu, S.; Cui, J.; Zhang, X.; Wu, W.; Niu, H.; Ma, X.; Xu, H.; Yi, M. Variable number tandem repeats in dopamine receptor D4 in Tourette’s syndrome. Mov. Disord. 2014. [Google Scholar] [CrossRef]
- Yoon, D.Y.; Rippel, C.A.; Kobets, A.J.; Morris, C.M.; Lee, J.E.; Williams, P.N.; Bridges, D.D.; Vandenbergh, D.J.; Shugart, Y.Y.; Singer, H.S. Dopaminergic polymorphisms in Tourette syndrome: Association with the DAT gene (SLC6A3). Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2007. [Google Scholar] [CrossRef]
- Dehning, S.; Müller, N.; Matz, J.; Bender, A.; Kerle, I.; Benninghoff, J.; Musil, R.; Spellmann, I.; Bondy, B.; Möller, H.J.; et al. A genetic variant of HTR2C may play a role in the manifestation of Tourette syndrome. Psychiatr. Genet. 2010. [Google Scholar] [CrossRef]
- Moya, P.R.; Wendland, J.R.; Rubenstein, L.M.; Timpano, K.R.; Heiman, G.A.; Tischfield, J.A.; King, R.A.; Andrews, A.M.; Ramamoorthy, S.; Mcmahon, F.J.; et al. Common and rare alleles of the serotonin transporter gene, SLC6A4, associated with Tourette’s disorder. Mov. Disord. 2013. [Google Scholar] [CrossRef] [Green Version]
- Crane, J.; Fagerness, J.; Osiecki, L.; Gunnell, B.; Stewart, S.E.; Pauls, D.L.; Scharf, J.M.; Cath, D.; Heutink, P.; Grados, M.; et al. Family-based genetic association study of DLGAP3 in Tourette Syndrome. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2011. [Google Scholar] [CrossRef] [Green Version]
- Karagiannidis, I.; Dehning, S.; Sandor, P.; Tarnok, Z.; Rizzo, R.; Wolanczyk, T.; Madruga-Garrido, M.; Hebebrand, J.; Nöthen, M.M.; Lehmkuhl, G.; et al. Support of the histaminergic hypothesis in tourette syndrome: Association of the histamine decarboxylase gene in a large sample of families. J. Med. Genet. 2013. [Google Scholar] [CrossRef] [Green Version]
- Buse, J.; Schoenefeld, K.; Münchau, A.; Roessner, V. Neuromodulation in Tourette syndrome: Dopamine and beyond. Neurosci. Biobehav. Rev. 2013, 37, 1069–1084. [Google Scholar]
- Maia, T.V.; Conceição, V.A. Dopaminergic Disturbances in Tourette Syndrome: An Integrative Account. Biol. Psychiatry 2018, 84, 332–344. [Google Scholar]
- Comings, D.E. Blood serotonin and tryptophan in Tourette syndrome. Am. J. Med. Genet. 1990. [Google Scholar] [CrossRef]
- Budman, C.L. The role of atypical antipsychotics for treatment of Tourette’s syndrome: An overview. Drugs 2014, 74, 1177–1193. [Google Scholar]
- Sinopoli, V.M.; Burton, C.L.; Kronenberg, S.; Arnold, P.D. A review of the role of serotonin system genes in obsessive-compulsive disorder. Neurosci. Biobehav. Rev. 2017, 80, 372–381. [Google Scholar]
- Bortolozzi, A.; Díaz-Mataix, L.; Scorza, M.C.; Celada, P.; Artigas, F. The activation of 5-HT2A receptors in prefrontal cortex enhances dopaminergic activity. J. Neurochem. 2005. [Google Scholar] [CrossRef] [Green Version]
- De Deurwaerdère, P.; Navailles, S.; Berg, K.A.; Clarke, W.P.; Spampinato, U. Constitutive Activity of the Serotonin2C Receptor Inhibits In Vivo Dopamine Release in the Rat Striatum and Nucleus Accumbens. J. Neurosci. 2004. [Google Scholar] [CrossRef]
- Esposito, E.; Di Matteo, V.; Di Giovanni, G. Serotonin-dopamine interaction: An overview. Prog. Brain Res. 2008, 172, 3–6. [Google Scholar]
- Sorensen, S.M.; Kehne, J.H.; Fadayel, G.M.; Humphreys, T.M.; Ketteler, H.J.; Sullivan, C.K.; Taylor, V.L.; Schmidt, C.J. Characterization of the 5-HT2 receptor antagonist MDL 100907 as a putative atypical antipsychotic: Behavioral, electrophysiological and neurochemical studies. J. Pharmacol. Exp. Ther. 1993, 266, 684–691. [Google Scholar]
- Larsen, M.B.; Sonders, M.S.; Mortensen, O.V.; Larson, G.A.; Zahniser, N.R.; Amara, S.G. Dopamine transport by the serotonin transporter: A Mechanistically distinct mode of substrate translocation. J. Neurosci. 2011. [Google Scholar] [CrossRef] [Green Version]
- Brivio, P.; Homberg, J.R.; Riva, M.A.; Calabrese, F. Alterations of Glutamatergic Markers in the Prefrontal Cortex of Serotonin Transporter Knockout Rats: A Developmental Timeline. Cell. Mol. Neurobiol. 2019. [Google Scholar] [CrossRef] [Green Version]
- Heinz, A.; Knable, M.B.; Wolf, S.S.; Jones, D.W.; Gorey, J.G.; Hyde, T.M.; Weinberger, D.R. Tourette’s syndrome: [I-123]β-CIT SPECT correlates of vocal tic severity. Neurology 1998. [Google Scholar] [CrossRef]
- Müller-Vahl, K.R.; Meyer, G.J.; Knapp, W.H.; Emrich, H.M.; Gielow, P.; Brücke, T.; Berding, G. Serotonin transporter binding in Tourette Syndrome. Neurosci. Lett. 2005. [Google Scholar] [CrossRef]
- Wong, D.F.; Brašić, J.R.; Singer, H.S.; Schretlen, D.J.; Kuwabara, H.; Zhou, Y.; Nandi, A.; Maris, M.A.; Alexander, M.; Ye, W.; et al. Mechanisms of dopaminergic and serotonergic neurotransmission in Tourette syndrome: Clues from an in vivo neurochemistry study with PET. Neuropsychopharmacology 2008. [Google Scholar] [CrossRef]
- Müller-Vahl, K.R.; Szejko, N.; Wilke, F.; Jakubovski, E.; Geworski, L.; Bengel, F.; Berding, G. Serotonin transporter binding is increased in Tourette syndrome with Obsessive Compulsive Disorder. Sci. Rep. 2019. [Google Scholar] [CrossRef]
- Gunther, J.; Tian, Y.; Stamova, B.; Lit, L.; Corbett, B.; Ander, B.; Zhan, X.; Jickling, G.; Bos-Veneman, N.; Liu, D.; et al. Catecholamine-related gene expression in blood correlates with tic severity in tourette syndrome. Psychiatry Res. 2012. [Google Scholar] [CrossRef]
- Li, J.; Li, Z.; Li, A.; Wang, S.; Qi, F.; Zhao, L.; Lv, H. Abnormal expression of dopamine and serotonin transporters associated with the pathophysiologic mechanism of Tourette syndrome. Neurol. India 2010. [Google Scholar] [CrossRef]
- Kilic, F.; Murphy, D.L.; Rudnick, G. A human serotonin transporter mutation causes constitutive activation of transport activity. Mol. Pharmacol. 2003. [Google Scholar] [CrossRef]
- Wendland, J.R.; Moya, P.R.; Kruse, M.R.; Ren-Patterson, R.F.; Jensen, C.L.; Timpano, K.R.; Murphy, D.L. A novel, putative gain-of-function haplotype at SLC6A4 associates with obsessive-compulsive disorder. Hum. Mol. Genet. 2008, 17, 717–723. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.Z.; Lipsky, R.H.; Zhu, G.; Akhtar, L.A.; Taubman, J.; Greenberg, B.D.; Xu, K.; Arnold, P.D.; Richter, M.A.; Kennedy, J.L.; et al. Serotonin transporter promoter gain-of-function genotypes are linked to obsessive-compulsive disorder. Am. J. Hum. Genet. 2006. [Google Scholar] [CrossRef] [Green Version]
- Bengel, D.; Greenberg, B.D.; Corá-Locatelli, G.; Altemus, M.; Heils, A.; Li, Q.; Murphy, D.L. Association of the serotonin transporter promoter regulatory region polymorphism and obsessive-compulsive disorder. Mol. Psychiatry 1999. [Google Scholar] [CrossRef] [Green Version]
- Lesch, K.P.; Bengel, D.; Heils, A.; Sabol, S.Z.; Greenberg, B.D.; Petri, S.; Benjamin, J.; Müller, C.R.; Hamer, D.H.; Murphy, D.L. Association of anxiety-related traits with a polymorphism in the serotonin transporter gene regulatory region. Science (80-.) 1996. [Google Scholar] [CrossRef]
- Iga, J.I.; Watanabe, S.Y.; Numata, S.; Umehara, H.; Nishi, A.; Kinoshita, M.; Inoshita, M.; Shimodera, S.; Fujita, H.; Ohmori, T. Association study of polymorphism in the serotonin transporter gene promoter, methylation profiles, and expression in patients with major depressive disorder. Hum. Psychopharmacol. 2016. [Google Scholar] [CrossRef]
- Wang, D.; Szyf, M.; Benkelfat, C.; Provençal, N.; Turecki, G.; Caramaschi, D.; Côté, S.M.; Vitaro, F.; Tremblay, R.E.; Booij, L. Peripheral SLC6A4 DNA methylation is associated with in vivo measures of human brain serotonin synthesis and childhood physical aggression. PLoS ONE 2012. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Lee, J.M.; Kim, J.W.; Cho, D.Y.; Yun, H.J.; Han, D.H.; Cheong, J.H.; Kim, B.N. Associations between serotonin transporter gene (SLC6A4) methylation and clinical characteristics and cortical thickness in children with ADHD. Psychol. Med. 2015. [Google Scholar] [CrossRef]
- Grünblatt, E.; Marinova, Z.; Roth, A.; Gardini, E.; Ball, J.; Geissler, J.; Wojdacz, T.K.; Romanos, M.; Walitza, S. Combining genetic and epigenetic parameters of the serotonin transporter gene in obsessive-compulsive disorder. J. Psychiatr. Res. 2018. [Google Scholar] [CrossRef] [Green Version]
- Müller-Vahl, K.R.; Loeber, G.; Kotsiari, A.; Müller-Engling, L.; Frieling, H. Gilles de la Tourette syndrome is associated with hypermethylation of the dopamine D2 receptor gene. J. Psychiatr. Res. 2017. [Google Scholar] [CrossRef] [Green Version]
- Palma-Gudiel, H.; Peralta, V.; Deuschle, M.; Navarro, V.; Fañanás, L. Epigenetics-by-sex interaction for somatization conferred by methylation at the promoter region of SLC6A4 gene. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2019. [Google Scholar] [CrossRef]
- Mol Debes, N.M.M.; Hjalgrim, H.; Skov, L. Validation of the presence of comorbidities in a danish clinical cohort of children with Tourette syndrome. J. Child Neurol. 2008. [Google Scholar] [CrossRef]
- Rstudio, T. RStudio: Integrated Development for R; Rstudio Team, PBC: Boston, MA, USA, 2020; Available online: http//www.rstudio.com (accessed on 8 January 2021).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24275-0. [Google Scholar]
- Kassambara, A. ggpubr: ‘ggplot2’ Based Publication Ready Plots. R Package Version 0.4.0. 2020. Available online: https://CRAN.R-project.org/package=ggpubr (accessed on 8 January 2021).
- Steeves, T.D.L.; Fox, S.H. Neurobiological basis of serotonin-dopamine antagonists in the treatment of Gilles de la Tourette syndrome. Prog. Brain Res. 2008, 172, 495–513. [Google Scholar]
- Cavallini, M.C.; Di Bella, D.; Catalano, M.; Bellodi, L. An association study between 5-HTTLPR polymorphism, COMT polymorphism, and Tourette’s syndrome. Psychiatry Res. 2000. [Google Scholar] [CrossRef]
- Rashidi, F.S.; Ahmadipour, E.; Shiravand, S.; Ahmadiani, A.; Asadi, S.; Shams, J. Association of the functional serotonin transporter haplotype with familial form of obsessive compulsive disorder in Iranian patients. Int. J. Psychiatry Clin. Pract. 2018. [Google Scholar] [CrossRef]
- Voyiaziakis, E.; Evgrafov, O.; Li, D.; Yoon, H.J.; Tabares, P.; Samuels, J.; Wang, Y.; Riddle, M.A.; Grados, M.A.; Bienvenu, O.J.; et al. Association of SLC6A4 variants with obsessive-compulsive disorder in a large multicenter US family study. Mol. Psychiatry 2011. [Google Scholar] [CrossRef] [Green Version]
- Wendland, J.R.; Kruse, M.R.; Cromer, K.C.; Murphy, D.L. A large case-control study of common functional SLC6A4 and BDNF variants in obsessive-compulsive disorder. Neuropsychopharmacology 2007. [Google Scholar] [CrossRef] [Green Version]
- Makki, M.I.; Behen, M.; Bhatt, A.; Wilson, B.; Chugani, H.T. Microstructural abnormalities of striatum and thalamus in children with tourette syndrome. Mov. Disord. 2008. [Google Scholar] [CrossRef]
- Draganski, B.; Martino, D.; Cavanna, A.E.; Hutton, C.; Orth, M.; Robertson, M.M.; Critchley, H.D.; Frackowiak, R.S. Multispectral brain morphometry in Tourette syndrome persisting into adulthood. Brain 2010. [Google Scholar] [CrossRef] [Green Version]
- Müller-Vahl, K.R.; Kaufmann, J.; Grosskreutz, J.; Dengler, R.; Emrich, H.M.; Peschel, T. Prefrontal and anterior cingulate cortex abnormalities in Tourette Syndrome: Evidence from voxel-based morphometry and magnetization transfer imaging. BMC Neurosci. 2009. [Google Scholar] [CrossRef] [Green Version]
- Müller-Vahl, K.R.; Grosskreutz, J.; Prell, T.; Kaufmann, J.; Bodammer, N.; Peschel, T. Tics are caused by alterations in prefrontal areas, thalamus and putamen, while changes in the cingulate gyrus reflect secondary compensatory mechanisms. BMC Neurosci. 2014. [Google Scholar] [CrossRef] [Green Version]
- Segura, B.; Strafella, A.P. Functional imaging of dopaminergic neurotransmission in tourette syndrome. In International Review of Neurobiology; Academic Press: Cambridge, MA, USA, 2013. [Google Scholar]


{kind=link}
| Variable | GTS Individuals | Controls | chi2 | p-Value | OR | |
|---|---|---|---|---|---|---|
| Genotype: no. (%) | ||||||
| Total number of individuals | 72 | 87 | ||||
| L/L | 28 (38.9) | 23 (26.4) | ||||
| S/L | 31 (43.1) | 46 (52.9) | ||||
| S/S | 13 (18.1) | 18 (20.7) | 2.829 | 0.243 | ||
| Allele: no. (%) | ||||||
| L | 87 (60.4) | 92 (52.9) | ||||
| S | 57 (39.6) | 82 (47.1) | 1.822 | 0.177 | 1.360 | |
| Haplotype: no. (%) | ||||||
| Total number of individuals | 41 | 40 | ||||
| LAC | 47 (57) | 42 (52.5) | ||||
| LAT, LG, S | 35 (43) | 38 (47.5) | 0.380 | 0.538 | 1.2150 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hildonen, M.; Levy, A.M.; Dahl, C.; Bjerregaard, V.A.; Birk Møller, L.; Guldberg, P.; Debes, N.M.; Tümer, Z. Elevated Expression of SLC6A4 Encoding the Serotonin Transporter (SERT) in Gilles de la Tourette Syndrome. Genes 2021, 12, 86. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12010086
Hildonen M, Levy AM, Dahl C, Bjerregaard VA, Birk Møller L, Guldberg P, Debes NM, Tümer Z. Elevated Expression of SLC6A4 Encoding the Serotonin Transporter (SERT) in Gilles de la Tourette Syndrome. Genes. 2021; 12(1):86. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12010086
Chicago/Turabian StyleHildonen, Mathis, Amanda M. Levy, Christina Dahl, Victoria A. Bjerregaard, Lisbeth Birk Møller, Per Guldberg, Nanette M. Debes, and Zeynep Tümer. 2021. "Elevated Expression of SLC6A4 Encoding the Serotonin Transporter (SERT) in Gilles de la Tourette Syndrome" Genes 12, no. 1: 86. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12010086