
Revisiting Plant Heterosis—From Field Scale to Molecules

 , , , ,
, , , ,
Abstract
:
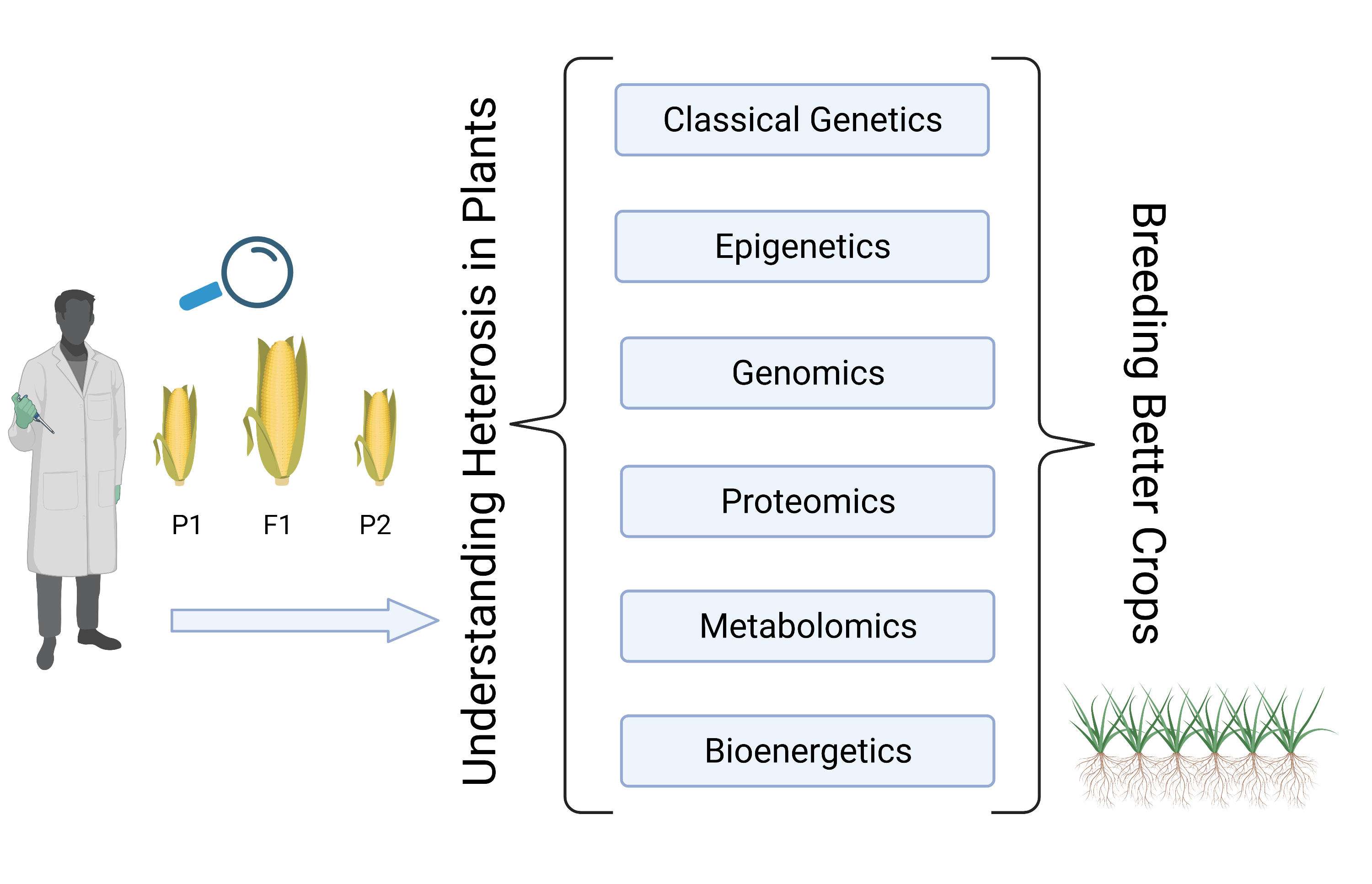
1. Introduction
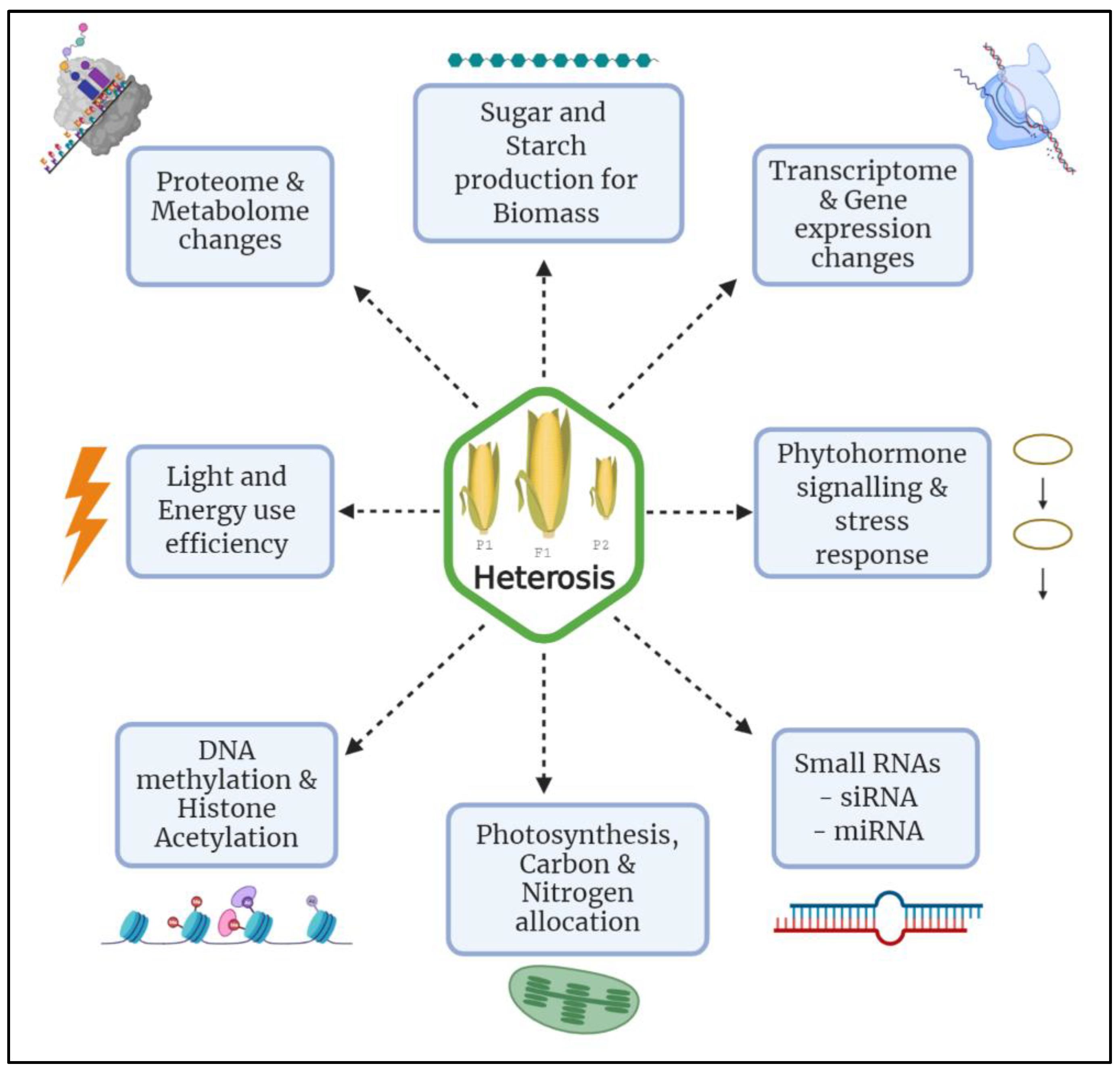
2. Understanding the Molecular Basis of Heterosis
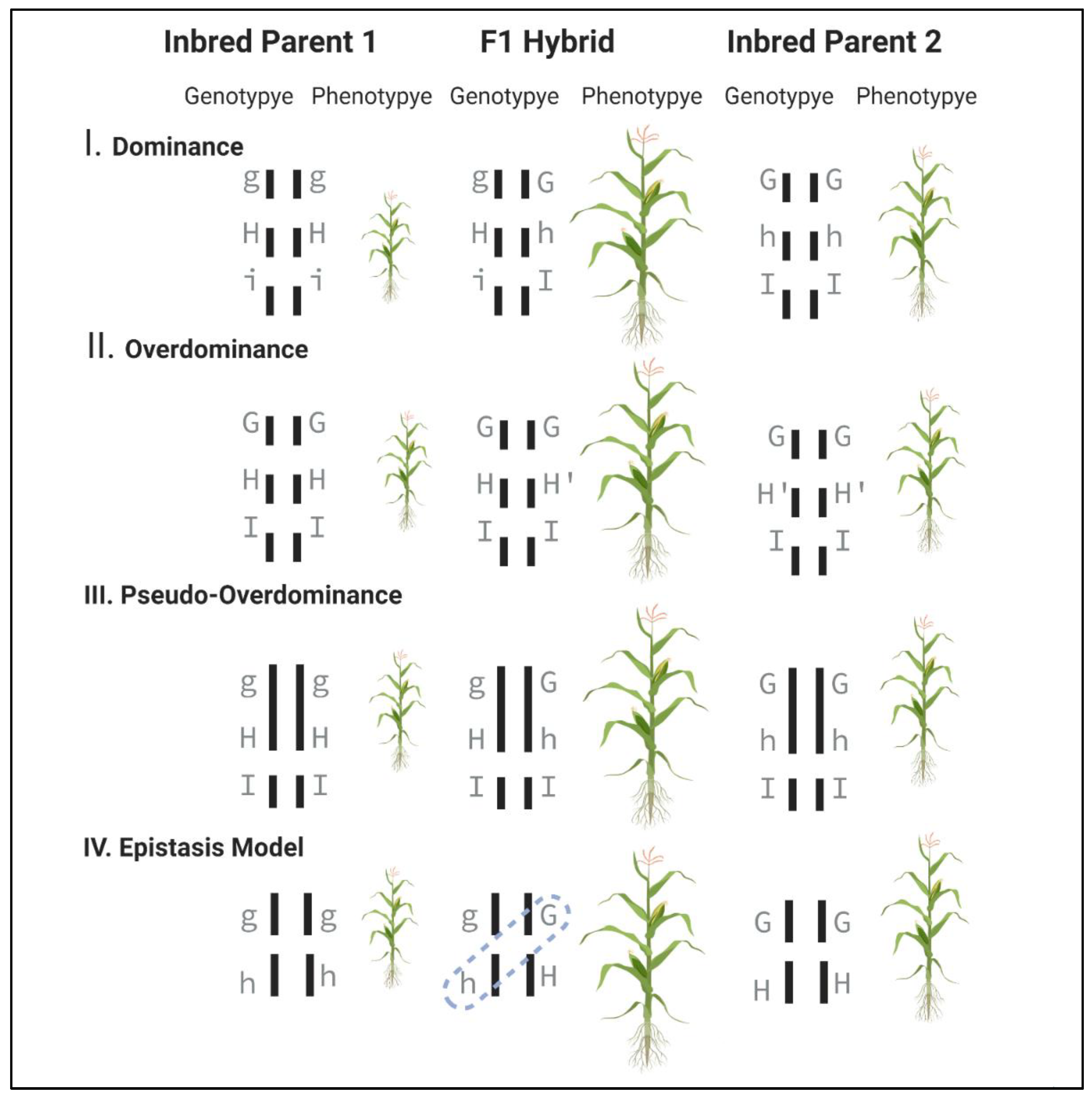
2.1. Genetic Models Explaining the Phenomenon of Heterosis
2.1.1. Dominance Model
2.1.2. Overdominance Model
2.1.3. Pseudo-Overdominance Model
2.1.4. Epistatic Model
2.2. Genomic View of Heterosis
2.3. Epigenetic View of Heterosis
2.3.1. Heterosis and DNA Methylation
2.3.2. Heterosis and Histone Modifications
2.3.3. Small RNA (sRNAs)—Role in Epigenetic Regulation and Heterosis
2.4. Heterosis, Proteomics and Transcriptomics
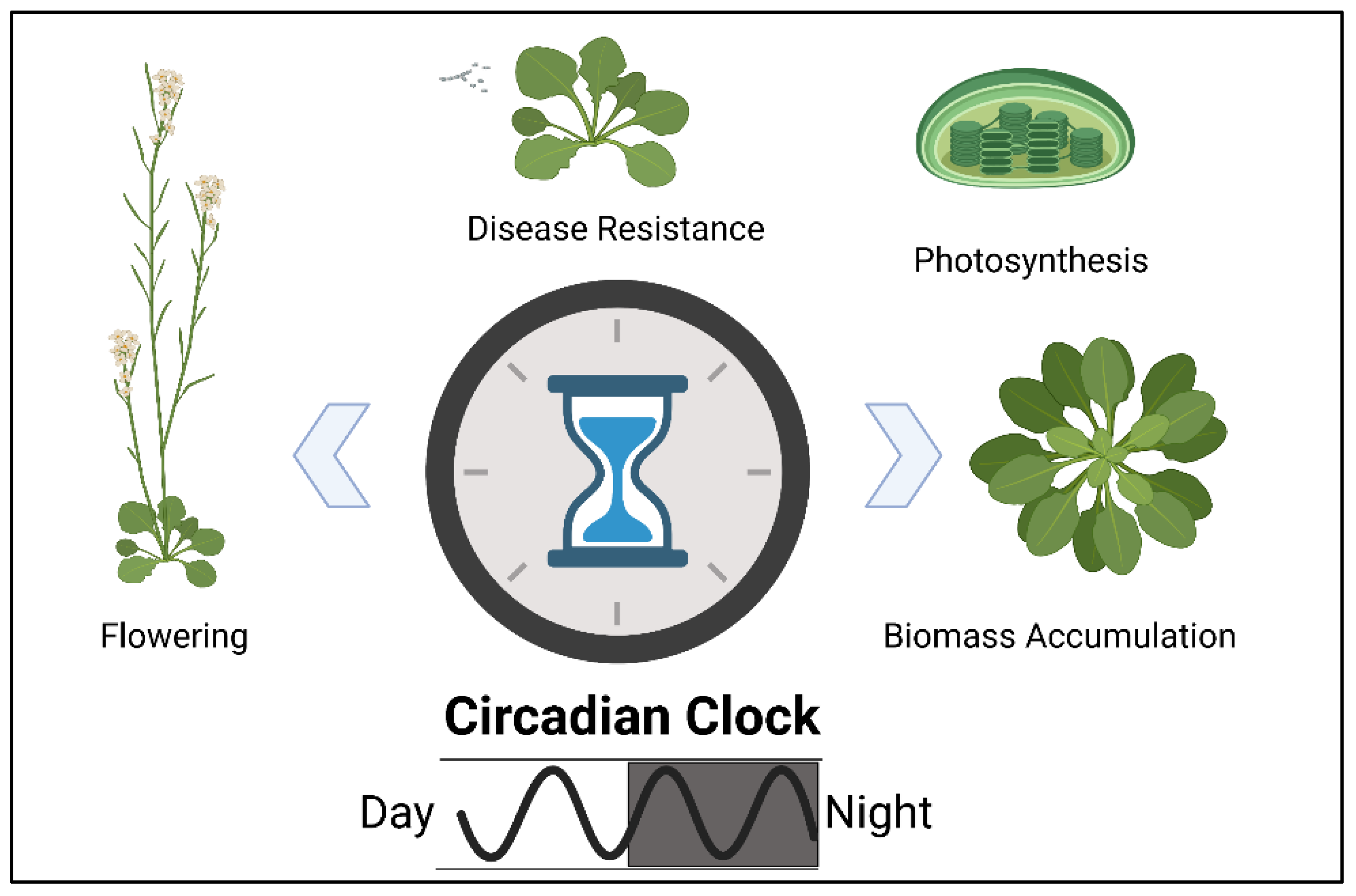
2.5. Intrinsic Biological Processes Contributing to Heterosis
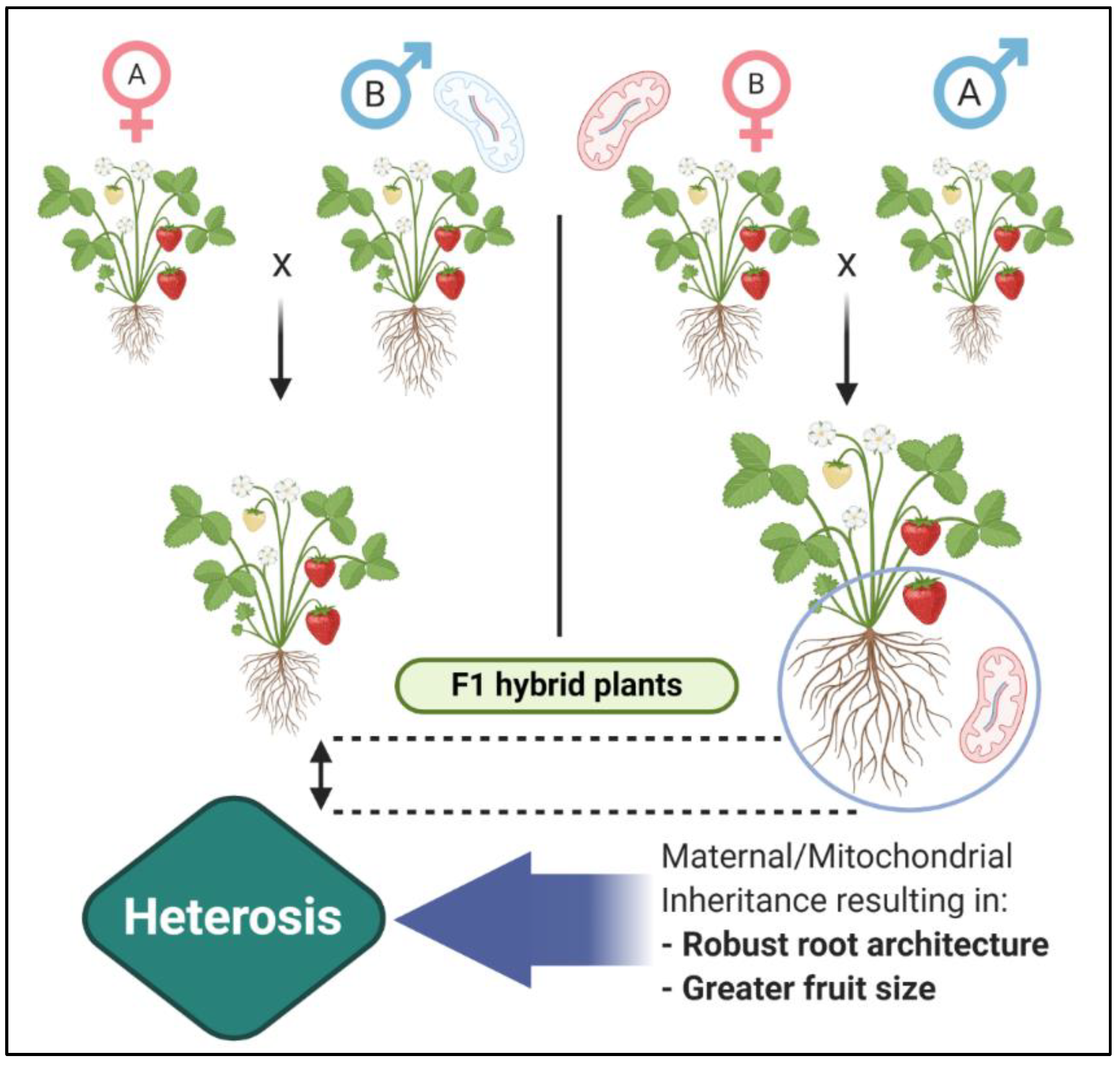
2.6. Mitochondrial Inheritance and Heterosis
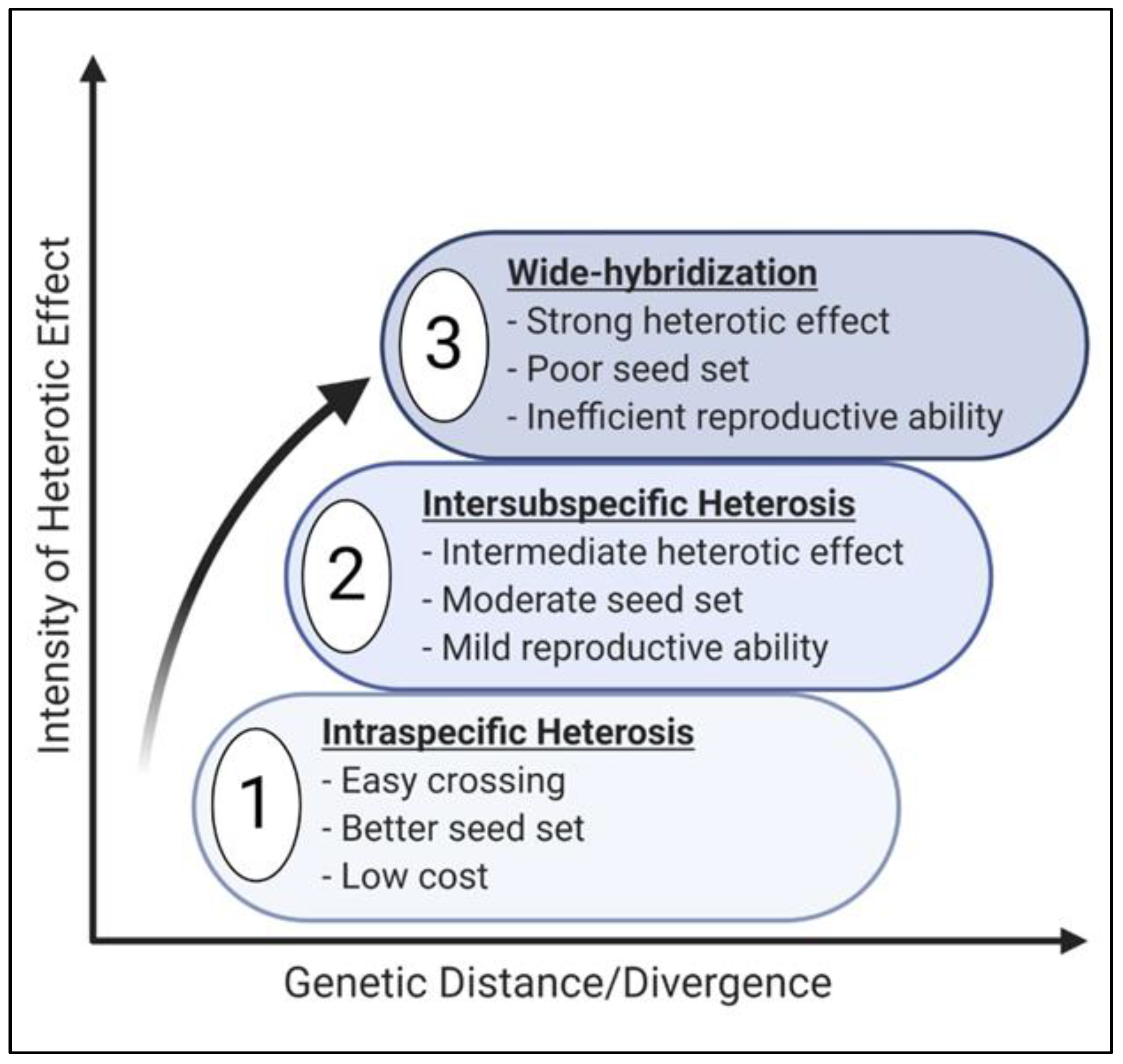
3. Heterosis in Self-Pollinated and Apomictic Plant Species
4. Future Perspective on Understanding and Utilizing Heterosis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fu, D.; Xiao, M.; Hayward, A.; Fu, Y.; Liu, G.; Jiang, G.; Zhang, H. Utilization of crop heterosis: A review. Euphytica 2014, 197, 161–173. [Google Scholar] [CrossRef]
- Hochholdinger, F.; Baldauf, J.A. Heterosis in plants. Curr. Biol. 2018, 28, R1089–R1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, S.-H.; Zhuang, J.-Y.; Fan, Y.-Y.; Du, J.-H.; Cao, L.-Y. Progress in Research and Development on Hybrid Rice: A Super-domesticate in China. Ann. Bot. 2007, 100, 959–966. [Google Scholar] [CrossRef]
- Hochholdinger, F.; Hoecker, N. Towards the molecular basis of heterosis. Trends Plant Sci. 2007, 12, 427–432. [Google Scholar] [CrossRef]
- Chen ZJ Molecular mechanisms of polyploidy and hybrid vigor. Trends Plant Sci. 2010, 15, 57–71. [CrossRef] [Green Version]
- Wehner, T.C. Heterosis in Vegetable crops. In Genetic Exploitation of Heterosis in Crops, 1st ed.; Coors, J.G., Pandey, S., Eds.; American Society of Agronomy: Madison, WI, USA, 1999; pp. 387–397. [Google Scholar]
- East, E.M. Heterosis. Genetics 1936, 21, 375–397. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.J. Genomic and epigenetic insights into the molecular bases of heterosis. Nat. Rev. Genet. 2013, 14, 471–482. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yuan, L. Hybrid Rice: Genetics, Breeding, and Seed Production. Plant Breed. Rev. 1999, 17, 15–158. [Google Scholar] [CrossRef]
- Virk, P.S.; Khush, G.S.; Peng, S. Breeding to Enhance Yield Potential of Rice at IRRI: The Ideotype Approach. Int. Rice Res. Notes 2004, 21, 5–9. [Google Scholar]
- Wang, Z.; Ni, Z.; Wu, H.; Nie, X.; Sun, Q. Heterosis in root development and differential gene expression between hybrids and their parental inbreds in wheat (Triticum aestivum L.). Theor. Appl. Genet. 2006, 113, 1283–1294. [Google Scholar] [CrossRef]
- Zeliang, P.K.; Pattanayak, A. Wide Hybridization in the Genus Oryza: Aspects and Prospects. Indian J. Hill Farming 2013, 26, 71–77. [Google Scholar]
- Shewry, P.R. The HEALTHGRAIN programme opens new opportunities for improving wheat for nutrition and health. Nutr. Bull. 2009, 34, 225–231. [Google Scholar] [CrossRef]
- Edger, P.P.; Poorten, T.J.; VanBuren, R.; Hardigan, M.A.; Colle, M.; McKain, M.R.; Smith, R.D.; Teresi, S.J.; Nelson, A.D.L.; Wai, C.M.; et al. Origin and evolution of the octoploid strawberry genome. Nat. Genet. 2019, 51, 541–547. [Google Scholar] [CrossRef] [Green Version]
- Falconer, D.S.; Mackay, T.F.C. Introduction to Quantitative Genetics; Longman Group: Harlow, UK, 1996. [Google Scholar]
- Birchler, J.A.; Auger, D.L.; Riddle, N.C. In Search of the Molecular Basis of Heterosis. Plant Cell 2003, 15, 2236–2239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schnable, P.S.; Springer, N.M. Progress Toward Understanding Heterosis in Crop Plants. Annu. Rev. Plant Biol. 2013, 64, 71–88. [Google Scholar] [CrossRef] [PubMed]
- Ryder, P.; McKeown, P.C.; Fort, A.; Spillane, C. Epigenetics and Heterosis in Crop Plants. In Epigenetics in Plants of Agronomic Importance: Fundamentals and Applications; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2019; pp. 129–147. [Google Scholar]
- Yao, H.; Gray, A.D.; Auger, D.L.; Birchler, J.A. Genomic dosage effects on heterosis in triploid maize. Proc. Natl. Acad. Sci. USA 2013, 110, 2665–2669. [Google Scholar] [CrossRef] [Green Version]
- Birchler, J.A.; Yao, H.; Chudalayandi, S.; Vaiman, D.; Veitia, R.A. Heterosis. Plant Cell 2010, 22, 2105–2112. [Google Scholar] [CrossRef] [Green Version]
- Birchler, J.A.; Yao, H.; Chudalayandi, S. Unraveling the genetic basis of hybrid vigor. Proc. Natl. Acad. Sci. USA 2006, 103, 12957–12958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powers, L. An expansion of jones’s theory for the explanation of heterosis. Genetics 1944, 2, 466–475. [Google Scholar] [CrossRef]
- Liang, Q.; Shang, L.; Wang, Y.; Hua, J. Partial Dominance, Overdominance and Epistasis as the Genetic Basis of Heterosis in Upland Cotton (Gossypium hirsutum L.). PLoS ONE 2015, 10, e0143548. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Liu, J.; Zhang, J. A review on research and utilization of hybrid vigor of cotton. Acta Gossypii Sin. 1994, 3, 135–139. [Google Scholar]
- Lisec, J.; Steinfath, M.; Meyer, R.C.; Selbig, J.; Melchinger, A.E.; Willmitzer, L.; Altmann, T. Identification of heterotic metabolite QTL in Arabidopsis thaliana RIL and IL populations. Plant J. 2009, 59, 777–788. [Google Scholar] [CrossRef] [PubMed]
- Song, R.; Messing, J. Gene expression of a gene family in maize based on noncollinear haplotypes. Proc. Natl. Acad. Sci. USA 2003, 100, 9055–9060. [Google Scholar] [CrossRef] [Green Version]
- Baranwal, V.K.; Mikkilineni, V.; Zehr, U.B.; Tyagi, A.K.; Kapoor, S. Heterosis: Emerging ideas about hybrid vigour. J. Exp. Bot. 2012, 63, 6309–6314. [Google Scholar] [CrossRef] [Green Version]
- Krieger, U.; Lippman, Z.; Zamir, D. The flowering gene SINGLE FLOWER TRUSS drives heterosis for yield in tomato. Nat. Genet. 2010, 42, 459–463. [Google Scholar] [CrossRef]
- Guo, B.; Chen, Y.; Zhang, G.; Xing, J.; Hu, Z.; Feng, W.; Yao, Y.; Peng, H.; Du, J.; Zhang, Y.; et al. Comparative Proteomic Analysis of Embryos between a Maize Hybrid and Its Parental Lines during Early Stages of Seed Germination. PLoS ONE 2013, 8, e65867. [Google Scholar] [CrossRef] [Green Version]
- Li, A.; Zhou, Y.; Jin, C.; Song, W.; Chen, C.; Wang, C. LaAP2L1, a Heterosis-Associated AP2/EREBP Transcription Factor of Larix, Increases Organ Size and Final Biomass by Affecting Cell Proliferation in Arabidopsis. Plant Cell Physiol. 2013, 54, 1822–1836. [Google Scholar] [CrossRef] [Green Version]
- Edwards, J.R.; Yarychkivska, O.; Boulard, M.; Bestor, T.H. DNA methylation and DNA methyltransferases. Epigenet. Chromatin 2017, 10, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, B.; Li, Y.; Robertson, K.D. DNA Methylation: Superior or Subordinate in the Epigenetic Hierarchy? Genes Cancer 2011, 2, 607–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bannister, A.J.; Kouzarides, T. Regulation of chromatin by histone modifications. Cell Res. 2011, 21, 381–395. [Google Scholar] [CrossRef] [PubMed]
- Demetriadou, C.; Koufaris, C.; Kirmizis, A. Histone N-alpha terminal modifications: Genome regulation at the tip of the tail. Epigenet. Chromatin 2020, 13, 1–13. [Google Scholar] [CrossRef]
- Gonzalez, M.; Li, F. DNA replication, RNAi and epigenetic inheritance. Epigenetics 2012, 7, 14–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, R.; Wang, X.; Moazed, D. Epigenetic inheritance mediated by coupling of RNAi and histone H3K9 methylation. Nat. Cell Biol. 2018, 558, 615–619. [Google Scholar] [CrossRef]
- He, G.; Zhu, X.; Elling, A.A.; Chen, L.; Wang, X.; Guo, L.; Liang, M.; He, H.; Zhang, H.; Chen, F.; et al. Global Epigenetic and Transcriptional Trends among Two Rice Subspecies and Their Reciprocal Hybrids. Plant Cell 2010, 22, 17–33. [Google Scholar] [CrossRef] [Green Version]
- Shen, H.; He, H.; Li, J.; Chen, W.; Wang, X.; Guo, L.; Peng, Z.; He, G.; Zhong, S.; Qi, Y.; et al. Genome-Wide Analysis of DNA Methylation and Gene Expression Changes in Two Arabidopsis Ecotypes and Their Reciprocal Hybrids. Plant Cell 2012, 24, 875–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greaves, I.; Groszmann, M.; Ying, H.; Taylor, J.; Peacock, W.J.; Dennis, E.S. Trans Chromosomal Methylation in Arabidopsis hybrids. Proc. Natl. Acad. Sci. USA 2012, 109, 3570–3575. [Google Scholar] [CrossRef] [Green Version]
- Lauss, K.; Wardenaar, R.; Oka, R.; van Hulten, M.H.A.; Guryev, V.; Keurentjes, J.J.B.; Stam, M.; Johannes, F. Parental DNA Methylation States Are Associated with Heterosis in Epigenetic Hybrids. Plant Physiol. 2018, 176, 1627–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, Z.; Kim, E.-D.; Ha, M.; Lackey, E.; Liu, J.; Zhang, Y.; Sun, Q.; Chen, Z.J. Altered circadian rhythms regulate growth vigour in hybrids and allopolyploids. Nat. Cell Biol. 2008, 457, 327–331. [Google Scholar] [CrossRef] [Green Version]
- Kim, A.J.; Kim, H.-S.; Choi, S.-H.; Jang, J.-Y.; Jeong, M.-J.; Lee, S.I. The Importance of the Circadian Clock in Regulating Plant Metabolism. Int. J. Mol. Sci. 2017, 18, 2680. [Google Scholar] [CrossRef] [Green Version]
- Jahnke, S.; Sarholz, B.; Thiemann, A.; Kühr, V.; Gutiérrez-Marcos, J.F.; Geiger, H.H.; Piepho, H.-P.; Scholten, S. Heterosis in early seed development: A comparative study of F1 embryo and endosperm tissues 6 days after fertilization. Theor. Appl. Genet. 2010, 120, 389–400. [Google Scholar] [CrossRef]
- Melnyk, C.W.; Molnar, A.; Bassett, A.; Baulcombe, D.C. Mobile 24 nt small RNAs direct transcriptional gene silencing in the root meristems of Arabidopsis thaliana. Curr. Biol. 2011, 21, 1678–1683. [Google Scholar] [CrossRef] [Green Version]
- Groszmann, M.; Greaves, I.K.; Albertyn, Z.I.; Scofield, G.N.; Peacock, W.J.; Dennis, E.S. Changes in 24-nt siRNA levels in Arabidopsis hybrids suggest an epigenetic contribution to hybrid vigor. Proc. Natl. Acad. Sci. USA 2011, 108, 2617–2622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.-M.; Chen, L.-T.; Patel, K.; Li, Y.-H.; Baulcombe, D.; Wu, S.-H. 22-nucleotide RNAs trigger secondary siRNA biogenesis in plants. Proc. Natl. Acad. Sci. USA 2010, 107, 15269–15274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chodavarapu, R.K.; Feng, S.; Ding, B.; Simon, S.A.; Lopez, D.; Jia, Y.; Wang, G.-L.; Meyers, B.; Jacobsen, S.E.; Pellegrini, M. Transcriptome and methylome interactions in rice hybrids. Proc. Natl. Acad. Sci. USA 2012, 109, 12040–12045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenan-Eichler, M.; Leshkowitz, D.; Tal, L.; Noor, E.; Melamed-Bessudo, C.; Feldman, M.; Levy, A.A. Wheat Hybridization and Polyploidization Results in Deregulation of Small RNAs. Genetics 2011, 188, 263–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsaftaris, S.A. Molecular aspects of heterosis in plants. Physiol. Plant. 1995, 94, 362–370. [Google Scholar] [CrossRef]
- Xing, J.; Sun, Q.; Ni, Z. Proteomic patterns associated with heterosis. Biochim. Biophys. Acta–Proteins Proteom. 2016, 1864, 908–915. [Google Scholar] [CrossRef]
- Zhang, C.; Yin, Y.; Zhang, A.; Lu, Q.; Wen, X.; Zhu, Z.; Zhang, L.; Lu, C. Comparative proteomic study reveals dynamic proteome changes between superhybrid rice LYP9 and its parents at different developmental stages. J. Plant Physiol. 2012, 169, 387–398. [Google Scholar] [CrossRef]
- Markert, C.L.; Moller, F. Multiple forms of enzymes: Tissue, ontogenetic, and species specific patterns. Proc. Natl. Acad. Sci. USA 1959, 45, 753–763. [Google Scholar] [CrossRef] [Green Version]
- Tsaftaris, A.S. Isozymes in plant breeding. Isozymes Curr. Top Biol. Med. Res. 1987, 13, 103–125. [Google Scholar]
- Damerval, C.; Hébert, Y.; de Vienne, D. Is the Polymorphism of Protein Amounts Related to Phenotypic Variability: A Comparison of Two-Dimensional Electrophoresis Data with Morphological Traits in Maize. Theor. Appl. Genet. 1987, 74, 194–202. [Google Scholar] [CrossRef]
- Dahal, D.; Mooney, B.P.; Newton, K.J. Specific Changes in Total and Mitochondrial Proteomes Are Associated with Higher Levels of Heterosis in Maize Hybrids. Plant J. 2012, 72, 70–83. [Google Scholar] [CrossRef]
- Stupar, R.M.; Gardiner, J.M.; Oldre, A.G.; Haun, W.J.; Chandler, V.L.; Springer, N.M. Gene Expression Analyses in Maize Inbreds and Hybrids with Varying Levels of Heterosis. BMC Plant Biol. 2008, 8, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comings, D.E.; MacMurray, J.P. Molecular Heterosis: A Review. Mol Gen. Met. 2000, 71, 19–31. [Google Scholar] [CrossRef]
- Fujimoto, R.; Taylor, J.M.; Sasaki, T.; Kawanabe, T.; Dennis, E.S. Genome wide gene expression in artificially synthesized amphidiploids of Arabidopsis. Plant Mol. Biol. 2011, 77, 419–431. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, R.; Taylor, J.; Shirasawa, S.; Peacock, W.J.; Dennis, E.S. Heterosis of Arabidopsis hybrids between C24 and Col is associated with increased photosynthesis capacity. Proc. Natl. Acad. Sci. USA 2012, 109, 7109–7114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flagel, L.; Udall, J.; Nettleton, D.; Wendel, J. Duplicate gene expression in allopolyploid Gossypium reveals two temporally distinct phases of expression evolution. BMC Biol. 2008, 6, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swanson-Wagner, R.A.; Jia, Y.; DeCook, R.; Borsuk, L.A.; Nettleton, D.; Schnable, P.S. All possible modes of gene action are observed in a global comparison of gene expression in a maize F1 hybrid and its inbred parents. Proc. Natl. Acad. Sci. USA 2006, 103, 6805–6810. [Google Scholar] [CrossRef] [Green Version]
- Song, X.; Ni, Z.; Yao, Y.; Xie, C.; Li, Z.; Wu, H.; Zhang, Y.; Sun, Q. Wheat (Triticum aestivum L.) root proteome and differentially expressed root proteins between hybrid and parents. Proteomics 2007, 7, 3538–3557. [Google Scholar] [CrossRef]
- Marcon, C.; Schützenmeister, A.; Schuütz, W.; Madlung, J.; Piepho, H.-P.; Hochholdinger, F. Nonadditive Protein Accumulation Patterns in Maize (Zea mays L.) Hybrids during Embryo Development. J. Proteome Res. 2010, 9, 6511–6522. [Google Scholar] [CrossRef]
- Kaeppler, S. Heterosis: Many Genes, Many Mechanisms—End the Search for an Undiscovered Unifying Theory. ISRN Bot. 2012, 2012, 682824. [Google Scholar] [CrossRef] [Green Version]
- Goff, S.A. A Unifying Theory for General Multigenic Heterosis: Energy Efficiency, Protein Metabolism, and Implications for Molecular Breeding. New Phytol. 2011, 189, 923–937. [Google Scholar] [CrossRef] [PubMed]
- Hauben, M.; Haesendonckx, B.; Standaert, E.; Van Der Kelen, K.; Azmi, A.; Akpo, H.; Van Breusegem, F.; Guisez, Y.; Bots, M.; Lambert, B.; et al. Energy use efficiency is characterized by an epigenetic component that can be directed through artificial selection to increase yield. Proc. Natl. Acad. Sci. USA 2009, 106, 20109–20114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Idle, J.R.; Gonzalez, F.J. Metabolomics. Cell Metab. 2007, 6, 348–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schauer, N.; Semel, Y.; Roessner, U.; Gur, A.; Balbo, I.; Carrari, F.; Pleban, T. Comprehensive Metabolic Profiling and Phenotyping of Interspecific Introgression Lines for Tomato Improvement. Nat. Biotech. 2006, 24, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Riedelsheimer, C.; Lisec, J.; Czedik-Eysenberg, A.; Sulpice, R.; Flis, A.; Grieder, C.; Altmann, T.; Stitt, M.; Willmitzer, L.; Melchinger, A.E. Genome-Wide Association Mapping of Leaf Metabolic Profiles for Dissecting Complex Traits in Maize. Nat. Acad. Sci. 2012, 109, 8872–8877. [Google Scholar] [CrossRef] [Green Version]
- Dan, Z.; Chen, Y.; Zhao, W.; Wang, Q.; Huang, W. Metabolome-Based Prediction of Yield Heterosis Contributes to the Breeding of Elite Rice. Life Sci. Alliance 2020, 3. [Google Scholar] [CrossRef]
- Wright, S. Physiological and Evolutionary Theories of Dominance. Am. Nat. 1934, 68, 24–53. [Google Scholar] [CrossRef]
- Rosas, U.; Barton, N.H.; Copsey, L.; De Reuille, P.B.; Coen, E. Cryptic Variation between Species and the Basis of Hybrid Performance. PLoS Biol. 2010, 8, e1000429. [Google Scholar] [CrossRef]
- Fiévet, J.B.; Nidelet, T.; Dillmann, C.; De Vienne, D. Heterosis Is a Systemic Property Emerging From Non-linear Genotype-Phenotype Relationships: Evidence From in Vitro Genetics and Computer Simulations. Front. Genet. 2018, 9, 159. [Google Scholar] [CrossRef]
- Vasseur, F.; Fouqueau, L.; De Vienne, D.; Nidelet, T.; Violle, C.; Weigel, D. Nonlinear phenotypic variation uncovers the emergence of heterosis in Arabidopsis thaliana. PLoS Biol. 2019, 17, e3000214. [Google Scholar] [CrossRef]
- Grimwood, B.G.; McDaniel, R.G. Variant Malate Dehydrogenase Isoenzymes in Mitochondrial Populations. Biochim. Biophys. Acta 1970, 220, 410–415. Available online: https://0-www-sciencedirect-com.brum.beds.ac.uk/science/article/pii/000527447090272X (accessed on 20 October 2021). [CrossRef]
- Sinha, S.K.; Khanna, R. Physiological, Biochemical, and Genetic Basis of Heterosis. Adv. Agron. 1975, 27, 123–174. Available online: https://0-www-sciencedirect-com.brum.beds.ac.uk/science/article/pii/S0065211308700099 (accessed on 20 October 2021).
- Mcdaniel, R.G.; Sarkissian, I.V. Heterosis: Complementation by Mitochondria. Science 1966, 152, 1640–1642. [Google Scholar] [CrossRef]
- Van Gelder, W.M.J.; Miedema, P. Significance of Mitochondrial Complementation for Plant Breeding: Negative Evidence from a Study on Maize. Euphytica 1975, 24, 421–429. [Google Scholar] [CrossRef]
- Cowling, W.A.; Gaynor, R.C.; Antolín, R.; Gorjanc, G.; Edwards, S.M.; Powell, O.; Hickey, J.M. In Silico Simulation of Future Hybrid Performance to Evaluate Heterotic Pool Formation in a Self-Pollinating Crop. Sci. Rep. 2020, 10, 1–8. [Google Scholar] [CrossRef]
- Moll, R.H.; Lindsey, M.F.; Robinson, H.F. Estimates of Genetic Variances and Level of Dominance in Maize. Genetics 1964, 49, 411–423. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/17248201 (accessed on 20 October 2021). [CrossRef] [PubMed]
- Mulualem, T.; Abate, M. Heterotic Response in Major Cereals and Vegetable Crops. Int. J. Plant Breed. Genet. 2016, 10, 69–78. [Google Scholar] [CrossRef] [Green Version]
- Lamkey, K.R.; Edwards, J.W. Quantitative Genetics of Heterosis; John Wiley Sons: Hoboken, NJ, USA, 1999; pp. 31–48. [Google Scholar] [CrossRef] [Green Version]
- Nienhuis, J.; Sills, G. The Potential of Hybrid Varieties in Self-Pollinating Vegetables. Reprod. Biol. Plant Breed. 1992, 387–396. [Google Scholar] [CrossRef]
- Blount, A.R.; Acuña, C.A. Bahiagrass. In Genetic Resources, Chromosome Engineering, and Crop Improvement Series: Forage Crops; Singh, R.J., Ed.; CRC Press: Boca Raton, FL, USA, 2009; Volume 5, pp. 81–101. [Google Scholar]
- Hanna, W.W.; Bashaw, E.C. Apomixis: Its identification and use in plant breeding. Crop Sci. 1987, 27, 1136–1139. [Google Scholar] [CrossRef]
- Evers, G.W.; Burson, B.L. Dallisgrass and other Paspalum species. In Warm-Season (C4) Grasses; Agronomy Monographs; Moser, L.E., Burson, B.L., Sollenberger, L.E., Eds.; ASA, CSSA, SSSA: Madison, WI, USA, 2004; Volume 45, pp. 681–713. [Google Scholar]
- Charlesworth, D. Effects of Inbreeding on the Genetic Diversity of Populations. Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 2003, 358, 1051–1070. [Google Scholar] [CrossRef] [Green Version]
- Volis, S. Adaptive Genetic Differentiation in a Predominantly Self-Pollinating Species Analyzed by Transplanting into Natural Environment, Crossbreeding and Q ST-F ST Test. New Phytol. 2011, 192, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Goff, S.A.; Zhang, Q. Heterosis in Elite Hybrid Rice: Speculation on the Genetic and Biochemical Mechanisms. Curr. Opin. Plant Biol. 2013, 16, 221–227. [Google Scholar] [CrossRef] [PubMed]
- McKeown, P.C.; Fort, A.; Duszynska, D.; Sulpice, R.; Spillane, C. Emerging Molecular Mechanisms for Biotechnological Harnessing of Heterosis in Crops. Trends Biotechnol. 2013, 16, 221–227. [Google Scholar] [CrossRef]
- Shivashankar, G.V. Mechanosignaling to the Cell Nucleus and Gene Regulation. Annu. Rev. Biophys. 2011, 40, 361–378. [Google Scholar] [CrossRef]
- Liu, J.H.; Li, M.; Zhang, Q.; Wei, X.; Huang, X. Exploring the molecular basis of heterosis for plant breeding. J. Integr. Plant Biol. 2020, 62, 287–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labroo, M.R.; Studer, A.J.; Rutkoski, J.E. Heterosis and Hybrid Crop Breeding: A Multidisciplinary Review. Front. Genet. 2021, 12, 643761. [Google Scholar] [CrossRef]
- Nicolas, S.D.; Mignon, G.L.; Eber, F.; Coriton, O.; Monod, H.; Clouet, V.; Huteau, V.; Lostanlen, A.; Delourme, R.; Chalhoub, B.; et al. Homeologous Recombination Plays a Major Role in Chromosome Rearrangements That Occur During Meiosis of Brassica Napus Haploids. Genet. Soc. Am. 2007, 75, 487–503. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Crop/Plant | Arabidopsis Orthologue | Expression State | Impact | Reference |
|---|---|---|---|---|---|
| ZAR1 (Zea mays ARGOS1) | Maize | ARGOS (Auxin Regulated Gene involved in Organ Size) | Overexpression | Increased organ size Increased Yield Improved drought tolerance | [29] |
| CNR1 (Cell Number Regulator 1) | Maize | Silencing | Increased plant size Increased organ size | [29] | |
| SFT (SINGLE FLOWER TRUSS) | Tomato | FT (FLOWERING LOCUS T) | Loss-of-function mutation | Enhanced yield | [28] |
| AP2/EREBP (APETALA 2/ethylene responsive element binding protein) | Arabidopsis | Over-expression | Cell proliferation Enhanced heterotic effect | [30] | |
| CCA1 (CIRCADIAN CLOCK ASSOCIATED 1) | Arabidopsis | Epigenetic modification | Increased vigor in plant development Increased biomass production | [8] | |
| LHY (LATE ELONGATED HYPOCOTYL) | Arabidopsis | Epigenetic modification | Increased vigour in plant development Increased biomass production | [8] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rehman, A.u.; Dang, T.; Qamar, S.; Ilyas, A.; Fatema, R.; Kafle, M.; Hussain, Z.; Masood, S.; Iqbal, S.; Shahzad, K. Revisiting Plant Heterosis—From Field Scale to Molecules. Genes 2021, 12, 1688. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12111688
Rehman Au, Dang T, Qamar S, Ilyas A, Fatema R, Kafle M, Hussain Z, Masood S, Iqbal S, Shahzad K. Revisiting Plant Heterosis—From Field Scale to Molecules. Genes. 2021; 12(11):1688. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12111688
Chicago/Turabian StyleRehman, Attiq ur, Trang Dang, Shanzay Qamar, Amina Ilyas, Reemana Fatema, Madan Kafle, Zawar Hussain, Sara Masood, Shehyar Iqbal, and Khurram Shahzad. 2021. "Revisiting Plant Heterosis—From Field Scale to Molecules" Genes 12, no. 11: 1688. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12111688