
Overexpression of a Voltage-Dependent Anion-Selective Channel (VDAC) Protein-Encoding Gene, MsVDAC, from Medicago sativa Confers Cold and Drought Tolerance to Transgenic Tobacco


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Growth and Experimental Treatments
2.2. Identification and Conserved Motif Analysis of VDAC Genes in Alfalfa
2.3. Isolation and Cloning of MsVDAC
2.4. Sequence Analysis of MsVDAC
2.5. Subcellular Localization Assays
2.6. Quantitative Real-Time PCR (qPCR) Analysis
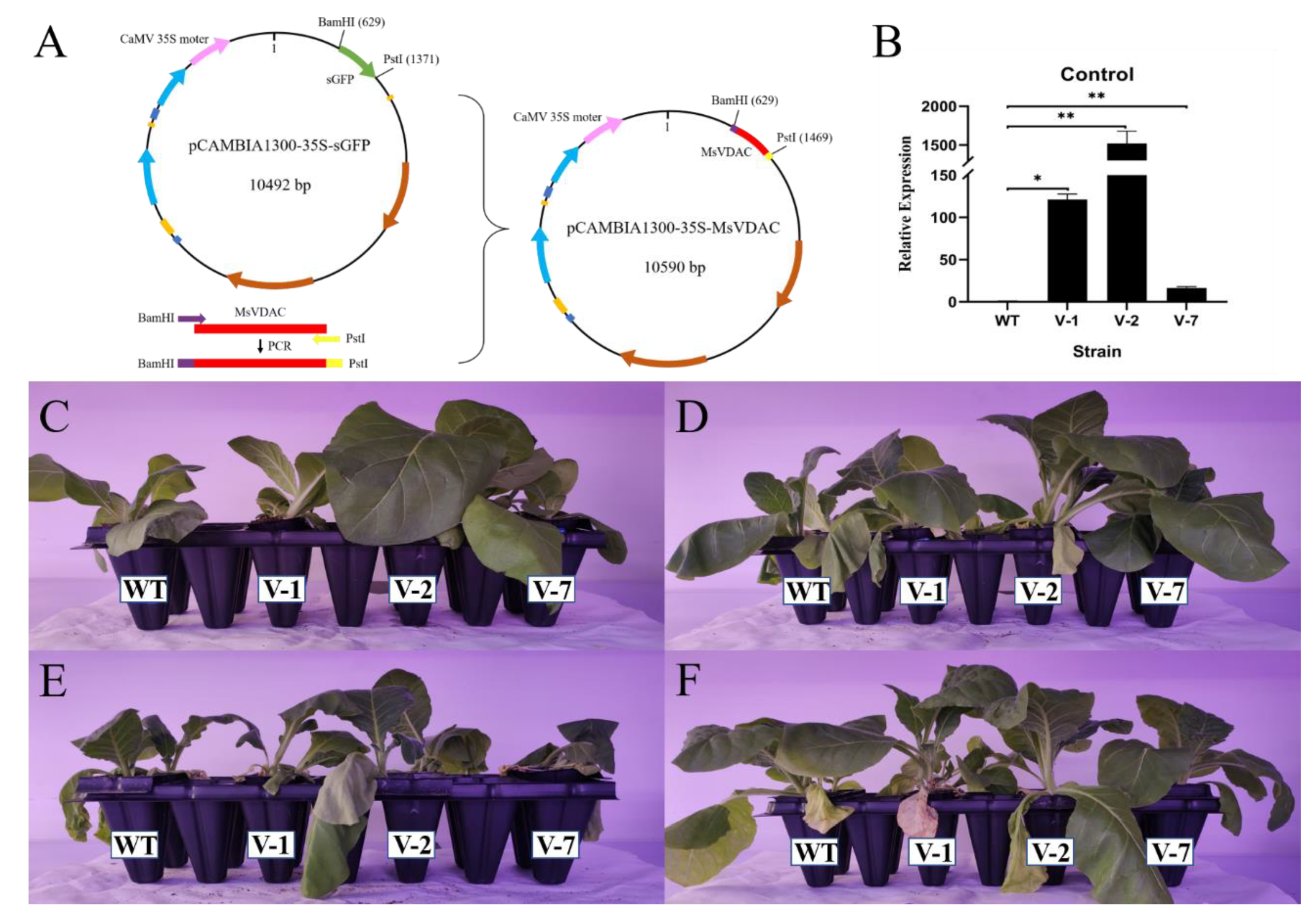
2.7. Plant Transformation and Generation of Transgenic Plants
2.8. Stress Tolerance Assays
2.9. Measurement of Physiological Changes
2.10. Expression Analysis of Stress-Response Genes
2.11. Statistical Analysis
3. Results
3.1. Isolation and Characteristics of MsVDAC
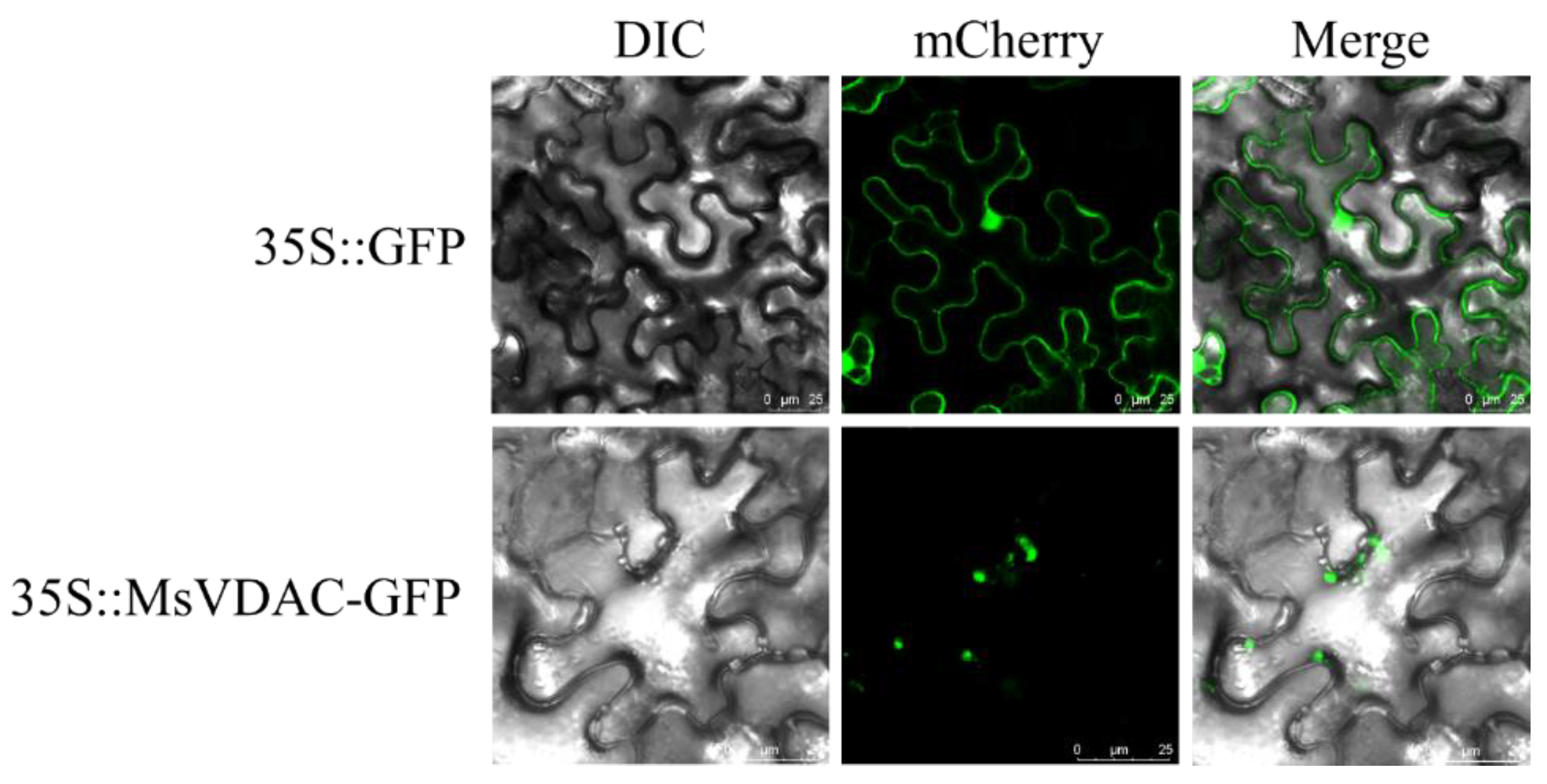
3.2. Subcellular Localization of MsVDAC
3.3. Expression Pattern of MsVDAC
3.4. Overexpression of MsVDAC Improves the Cold, Drought and Salt Tolerance of Transgenic Tobacco
3.5. Physiological Changes in Tobacco Overexpressing MsVDAC under Cold, Drought and Salt Stress
3.6. Expression Analysis of Stress-Response Genes in Transgenic Tobacco Plants Overexpressing MsVDAC under Cold and Drought Stress
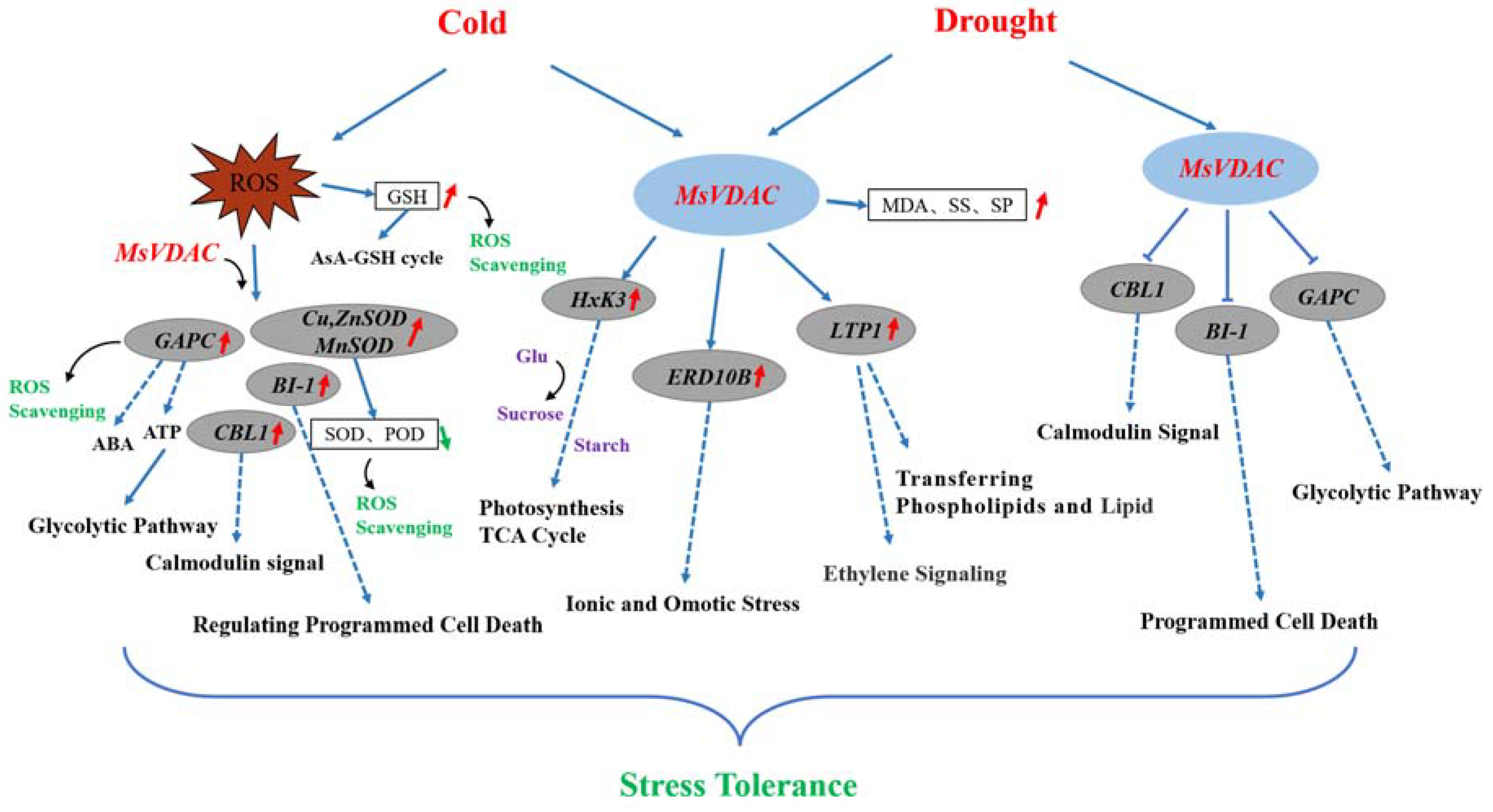
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Zhu, J.-K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Fountain, J.C.; Wang, H.; Ni, X.; Ji, P.; Lee, R.D.; Kemerait, R.C.; Scully, B.T.; Guo, B. Stress sensitivity is associated with differential accumulation of reactive oxygen and nitrogen species in maize genotypes with contrasting levels of drought tolerance. Int. J. Mol. Sci. 2015, 16, 24791–24819. [Google Scholar] [CrossRef] [Green Version]
- Cui, G.; Chai, H.; Yin, H.; Yang, M.; Hu, G.; Guo, M.; Yi, R.; Zhang, P. Full-length transcriptome sequencing reveals the low-temperature-tolerance mechanism of Medicago falcata roots. BMC Plant Biol. 2019, 19, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saijo, Y.; Loo, E.P. Plant immunity in signal integration between biotic and abiotic stress responses. New Phytol. 2020, 225, 87–104. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Zhou, G. Responses of leaf stomatal density to water status and its relationship with photosynthesis in a grass. J. Exp. Bot. 2008, 59, 3317–3325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, H.A.; Hussain, S.; Khaliq, A.; Ashraf, U.; Anjum, S.A.; Men, S.; Wang, L. Chilling and drought stresses in crop plants: Implications, cross talk, and potential management opportunities. Front. Plant Sci. 2018, 9, 393. [Google Scholar] [CrossRef]
- Kusano, T.; Tateda, C.; Berberich, T.; Takahashi, Y. Voltage-dependent anion channels: Their roles in plant defense and cell death. Plant Cell Rep. 2009, 28, 1301–1308. [Google Scholar] [CrossRef]
- Godbole, A.; Dubey, A.K.; Reddy, P.S.; Udayakumar, M.; Mathew, M.K. Mitochondrial VDAC and hexokinase together modulate plant programmed cell death. Protoplasma 2013, 250, 875–884. [Google Scholar] [CrossRef] [PubMed]
- Sanyal, S.K.; Kanwar, P.; Fernandes, J.L.; Mahiwal, S.; Yadav, A.K.; Samtani, H.; Srivastava, A.K.; Suprasanna, P.; Pandey, G.K. Arabidopsis Mitochondrial voltage-dependent Anion channels are involved in maintaining reactive oxygen species homeostasis, Oxidative and salt stress tolerance in yeast. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef]
- Tateda, C.; Watanabe, K.; Kusano, T.; Takahashi, Y. Molecular and genetic characterization of the gene family encoding the voltage-dependent anion channel in Arabidopsis. J. Exp. Bot. 2011, 62, 4773–4785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Zhang, L.-D.; Zuo, K.-J.; Qian, H.-M.; Cao, Y.-F.; Tang, K.-X. Cloning and expressional studies of the voltage-dependent Anion Channel Gene from Brassica rapa L. J. Integr. Plant Biol. 2006, 48, 197–203. [Google Scholar] [CrossRef]
- Clausen, C.; Ilkavets, I.; Thomson, R.; Philippar, K.; Vojta, A.; Neuhaus, E.; Fulgosi, H. Intracellular localization of VDAC proteins in plants. Planta 2004, 220, 30–37. [Google Scholar] [CrossRef]
- Badr, A.; El-Sherif, N.; Aly, S.; Ibrahim, S.D.; Ibrahim, M. Genetic diversity among selected Medicago sativa cultivars using inter-retrotransposon-amplified Polymorphism, Chloroplast DNA barcodes and Morpho-Agronomic trait analyses. Plants 2020, 9, 995. [Google Scholar] [CrossRef]
- Zhou, J.; Zhang, F.; Huo, Y.; Wilson, G.W.T.; Cobb, A.B.; Xu, X.; Xiong, X.; Liu, L.; Zhang, Y. Following legume establishment, microbial and chemical associations facilitate improved productivity in degraded grasslands. Plant Soil 2019, 443, 273–292. [Google Scholar] [CrossRef]
- Shen, C.; Du, H.; Chen, Z.; Lu, H.; Zhu, F.; Chen, H.; Meng, X.; Liu, Q.; Liu, P.; Zheng, L.; et al. The chromosome-level genome sequence of the Autotetraploid Alfalfa and resequencing of core Germplasms provide Genomic resources for Alfalfa research. Mol. Plant 2020, 13, 1250–1261. [Google Scholar] [CrossRef]
- Wei, Z.-H.; Liang, S.-L.; Wang, D.-M.; Liu, H.-Y.; Wanapat, M.; Liu, J.-X. Lactation performance and rumen fermentation in dairy cows fed a diet with alfalfa hay replaced by corn stover and supplemented with molasses. Asian Australas. J. Anim. Sci. 2019, 32, 1122–1127. [Google Scholar] [CrossRef]
- Zhang, C.; Shi, S. Physiological and proteomic responses of contrasting Alfalfa (Medicago sativa L.) varieties to PEG-induced osmotic stress. Front. Plant Sci. 2018, 9, 242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Liu, J.; An, Y.; Cao, Y.; Liu, Y.; Zhang, J.; Geng, J.; Hu, T.; Yang, P.; Tianming, H. MsPIP2;2, a novel aquaporin gene from Medicago sativa, confers salt tolerance in transgenic Arabidopsis. Environ. Exp. Bot. 2019, 165, 39–52. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Chance, B.; Maehly, A. Assay of catalases and peroxidases. Methods Enzym. 1955, 2, 764–775. [Google Scholar] [CrossRef]
- Elstner, E.F.; Heupel, A. Inhibition of nitrite formation from hydroxylammoniumchloride: A simple assay for superoxide dismutase. Anal. Biochem. 1976, 70, 616–620. [Google Scholar] [CrossRef]
- Yang, P.; Zhang, P.; Li, B.; Hu, T. Effect of nodules on dehydration response in alfalfa (Medicago sativa L.). Environ. Exp. Bot. 2013, 86, 29–34. [Google Scholar] [CrossRef]
- Hissin, P.J.; Hilf, R. A fluorometric method for determination of oxidized and reduced glutathione in tissues. Anal. Biochem. 1976, 74, 214–226. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Kanwar, P.; Samtani, H.; Sanyal, S.K.; Srivastava, A.K.; Suprasanna, P.; Pandey, G.K. VDAC and its interacting partners in plant and animal systems: An overview. Crit. Rev. Biotechnol. 2020, 40, 715–732. [Google Scholar] [CrossRef]
- Homblé, F.; Krammer, E.-M.; Prévost, M. Plant VDAC: Facts and speculations. Biochim. Biophys. Acta Biomembr. 2012, 1818, 1486–1501. [Google Scholar] [CrossRef]
- Desai, M.; Mishra, R.; Verma, D.; Nair, S.; Sopory, S.; Reddy, M. Structural and functional analysis of a salt stress inducible gene encoding voltage dependent anion channel (VDAC) from pearl millet (Pennisetum glaucum). Plant Physiol. Biochem. 2006, 44, 483–493. [Google Scholar] [CrossRef]
- Gupta, M.; Sharma, G.; Saxena, D.; Budhwar, R.; Vasudevan, M.; Gupta, V.; Gupta, A.; Gupta, R.; Chandran, D. Dual RNA-Seq analysis of Medicago truncatula and the pea powdery mildew Erysiphe pisi uncovers distinct host transcriptional signatures during incompatible and compatible interactions and pathogen effector candidates. Genomics 2020, 112, 2130–2145. [Google Scholar] [CrossRef]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of Malondialdehyde and 4-Hydroxy-2-Nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [Green Version]
- Farooq, M.; Hussain, M.; Wahid, A.; Siddique, K. Drought stress in plants: An overview. In Plant Responses to Drought Stress; Aroca, R., Ed.; Springer: Berlin, Germany, 2012; pp. 1–33. [Google Scholar]
- Guo, Y.; Xu, C.-B.; Sun, X.-J.; Hu, Z.; Fan, S.-J.; Jiang, Q.-Y.; Zhang, H. TaSAUR78 enhances multiple abiotic stress tolerance by regulating the interacting gene TaVDAC1. J. Integr. Agric. 2019, 18, 2682–2690. [Google Scholar] [CrossRef]
- Krasensky-Wrzaczek, J.; Jonak, C. Drought, salt, and temperature stress-induced metabolic rearrangements and regulatory networks. J. Exp. Bot. 2012, 63, 1593–1608. [Google Scholar] [CrossRef] [Green Version]
- Tateda, C.; Yamashita, K.; Takahashi, F.; Kusano, T.; Takahashi, Y. Plant voltage-dependent anion channels are involved in host defense against Pseudomonas cichorii and in Bax-induced cell death. Plant Cell Rep. 2009, 28, 41–51. [Google Scholar] [CrossRef]
- Pérez-Díaz, J.; Batista-Silva, W.; Almada, R.; Medeiros, D.B.; Arrivault, S.; Correa, F.; Bastías, A.; Rojas, P.; Beltrán, M.F.; Pozo, M.F.; et al. Prunus Hexokinase 3 genes alter primary C-metabolism and promote drought and salt stress tolerance in Arabidopsis transgenic plants. Sci. Rep. 2021, 11, 1–15. [Google Scholar] [CrossRef]
- Rojas, M.; Jimenez-Bremont, F.; Villicaña, C.; Carreón-Palau, L.; Arredondo-Vega, B.O.; Gómez-Anduro, G. Involvement of OpsLTP1 from Opuntia streptacantha in abiotic stress adaptation and lipid metabolism. Funct. Plant Biol. 2019, 46, 816. [Google Scholar] [CrossRef]
- Tapia, G.; Morales-Quintana, L.; Parra, C.; Berbel, A.; Alcorta, M. Study of nsLTPs in Lotus japonicus genome reveal a specific epidermal cell member (LjLTP10) regulated by drought stress in aerial organs with a putative role in cutin formation. Plant Mol. Biol. 2013, 82, 485–501. [Google Scholar] [CrossRef] [PubMed]
- Golizadeh, F.; Kumleh, H.H. Physiological responses and expression changes of fatty acid metabolism–related genes in Wheat (Triticum aestivum) under cold stress. Plant Mol. Biol. Rep. 2019, 37, 224–236. [Google Scholar] [CrossRef]
- Gupta, K.; Jha, B.; Agarwal, P.K. A Dehydration-Responsive Element Binding (DREB) transcription factor from the succulent halophyte salicornia brachiata enhances abiotic stress tolerance in Transgenic Tobacco. Mar. Biotechnol. 2014, 16, 657–673. [Google Scholar] [CrossRef] [PubMed]
- Close, T.J. Dehydrins: A commonalty in the response of plants to dehydration and low temperature. Physiol. Plant. 2010, 100, 291–296. [Google Scholar] [CrossRef]
- Zelko, I.N.; Mariani, T.J.; Folz, R.J. Superoxide dismutase multigene family: A comparison of the CuZn-SOD (SOD1), Mn-SOD (SOD2), and EC-SOD (SOD3) gene structures, evolution, and expression. Free. Radic. Biol. Med. 2002, 33, 337–349. [Google Scholar] [CrossRef]
- Qin, G.; Meng, X.; Wang, Q.; Tian, S. Oxidative damage of mitochondrial proteins contributes to fruit senescence: A redox proteomics analysis. J. Proteome Res. 2009, 8, 2449–2462. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, Z.; Peng, Y.; Wang, X.; Peng, D.; Li, Y.; McPhee, D.J.; Zhang, X.; Ma, X.; Huang, L.; et al. Clones of FeSOD, MDHAR, DHAR Genes from white clover and gene expression analysis of ROS-Scavenging enzymes during abiotic stress and hormone treatments. Molecules 2015, 20, 20939–20954. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Anjum, N.A.; Hasanuzzaman, M.; Gill, R.; Trivedi, D.K.; Ahmad, I.; Pereira, E.; Tuteja, N. Glutathione and glutathione reductase: A boon in disguise for plant abiotic stress defense operations. Plant Physiol. Biochem. 2013, 70, 204–212. [Google Scholar] [CrossRef]
- Cheong, Y.H.; Kim, K.-N.; Pandey, G.K.; Gupta, R.; Grant, J.J.; Luan, S. CBL1, a calcium sensor that differentially regulates salt, drought, and cold responses in Arabidopsis. Plant Cell 2003, 15, 1833–1845. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.-Y.; Xu, Z.-S.; He, G.-Y.; Yang, G.-X.; Chen, M.; Li, L.-C.; Ma, Y. The voltage-dependent Anion Channel 1 (AtVDAC1) negatively regulates plant cold responses during germination and seedling development in Arabidopsis and interacts with calcium sensor CBL1. Int. J. Mol. Sci. 2013, 14, 701–713. [Google Scholar] [CrossRef] [Green Version]
- Wojtera-Kwiczor, J.; Gross, F.; Leffers, H.-M.; Kang, M.; Schneider, M.; Scheibe, R. Transfer of a Redox-signal through the Cytosol by Redox-dependent Microcompartmentation of glycolytic enzymes at mitochondria and actin cytoskeleton. Front. Plant Sci. 2013, 3, 284. [Google Scholar] [CrossRef] [Green Version]
- Danshina, P.V.; Schmalhausen, E.V.; Avetisyan, A.V.; Muronetz, V.I. Mildly oxidized glyceraldehyde-3-phosphate dehydrogenase as a possible regulator of Glycolysis. IUBMB Life 2001, 51, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, H.; Yang, S. Cytosolic TaGAPC2 enhances tolerance to drought stress in transgenic arabidopsis plants. Int. J. Mol. Sci. 2020, 21, 7499. [Google Scholar] [CrossRef]
- Zhang, L.; Xu, Z.; Ji, H.; Zhou, Y.; Yang, S. TaWRKY40 transcription factor positively regulate the expression of TaGAPC1 to enhance drought tolerance. BMC Genom. 2019, 20, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Fang, H.; Liu, J.; Reid, S.; Hou, J.; Zhou, T.; Tian, Z.; Song, B.; Xie, C. Cytosolic glyceraldehyde-3-phosphate dehydrogenases play crucial roles in controlling cold-induced sweetening and apical dominance of potato (Solanum tuberosum L.) tubers. Plant Cell Environ. 2017, 40, 3043–3054. [Google Scholar] [CrossRef] [PubMed]
- Godlewski, M.; Kobylińska, A. Bax Inhibitor 1 (BI-1) as a conservative regulator of Programmed Cell Death. Postępy Hig. I Med. Doświadczalnej 2019, 73, 681–702. [Google Scholar] [CrossRef]
- Nakashima, K.; Yamaguchi-Shinozaki, K. ABA signaling in stress-response and seed development. Plant Cell Rep. 2013, 32, 959–970. [Google Scholar] [CrossRef] [PubMed]
- Tian, S.; Mao, X.; Zhang, H.; Chen, S.; Zhai, C.; Yang, S.; Jing, R. Cloning and characterization of TaSnRK2.3, a novel SnRK2 gene in common wheat. J. Exp. Bot. 2013, 64, 2063–2080. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, M.; Duan, X.; Wang, Z.; Yin, H.; Zang, J.; Zhu, K.; Wang, Y.; Zhang, P. Overexpression of a Voltage-Dependent Anion-Selective Channel (VDAC) Protein-Encoding Gene, MsVDAC, from Medicago sativa Confers Cold and Drought Tolerance to Transgenic Tobacco. Genes 2021, 12, 1706. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12111706
Yang M, Duan X, Wang Z, Yin H, Zang J, Zhu K, Wang Y, Zhang P. Overexpression of a Voltage-Dependent Anion-Selective Channel (VDAC) Protein-Encoding Gene, MsVDAC, from Medicago sativa Confers Cold and Drought Tolerance to Transgenic Tobacco. Genes. 2021; 12(11):1706. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12111706
Chicago/Turabian StyleYang, Mei, Xinhang Duan, Zhaoyu Wang, Hang Yin, Junrui Zang, Kai Zhu, Yumeng Wang, and Pan Zhang. 2021. "Overexpression of a Voltage-Dependent Anion-Selective Channel (VDAC) Protein-Encoding Gene, MsVDAC, from Medicago sativa Confers Cold and Drought Tolerance to Transgenic Tobacco" Genes 12, no. 11: 1706. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12111706