
A Non-Shedding Fruit Elaeis oleifera Palm Reveals Perturbations to Hormone Signaling, ROS Homeostasis, and Hemicellulose Metabolism

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Phenotyping the Abscission Character
2.3. RNA Extraction for Transcriptome Analyses
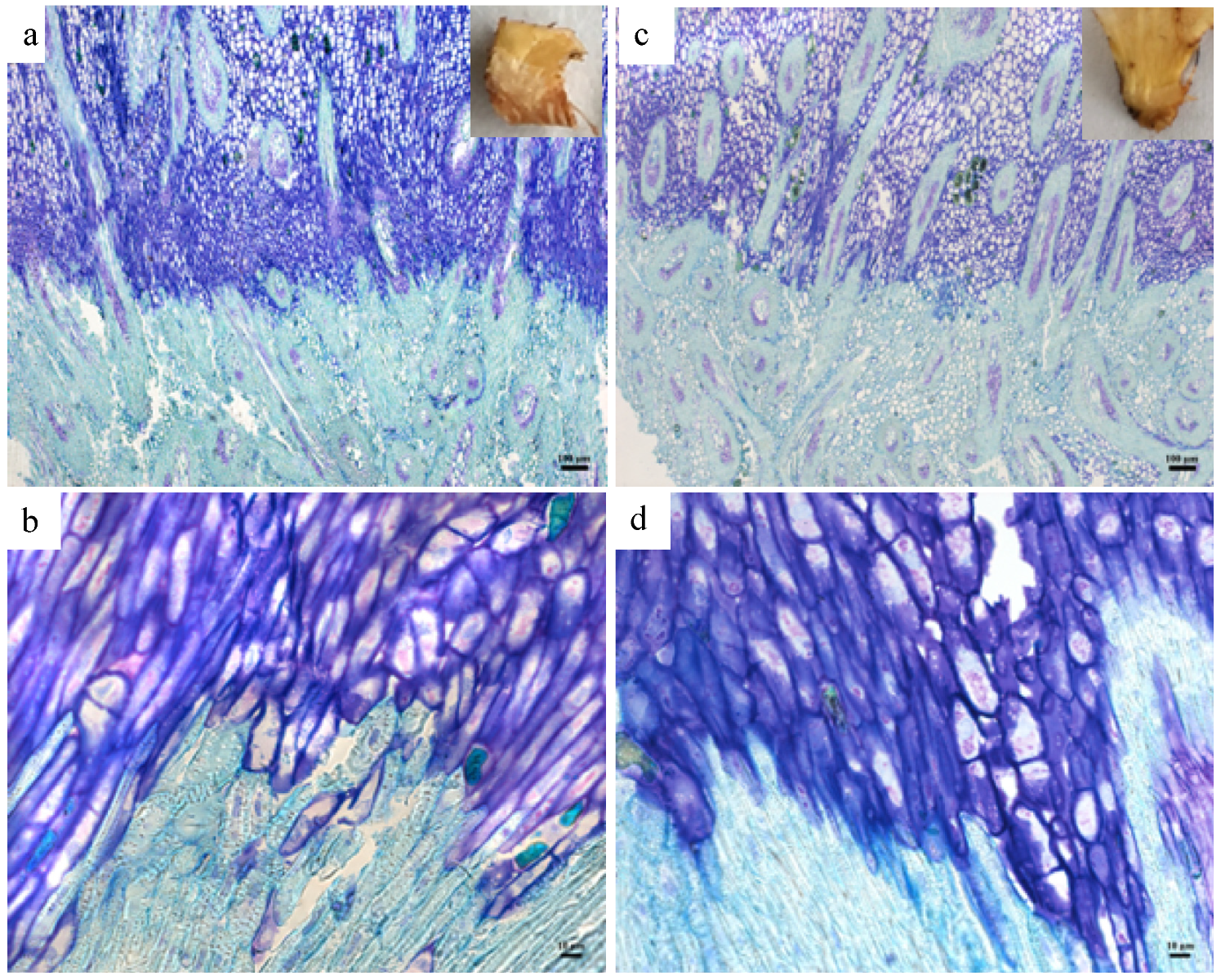
2.4. Histology
2.5. Analysis of Differential Gene Expression
2.6. Functional Annotation of Genes and Identification of Enriched Categories
3. Results
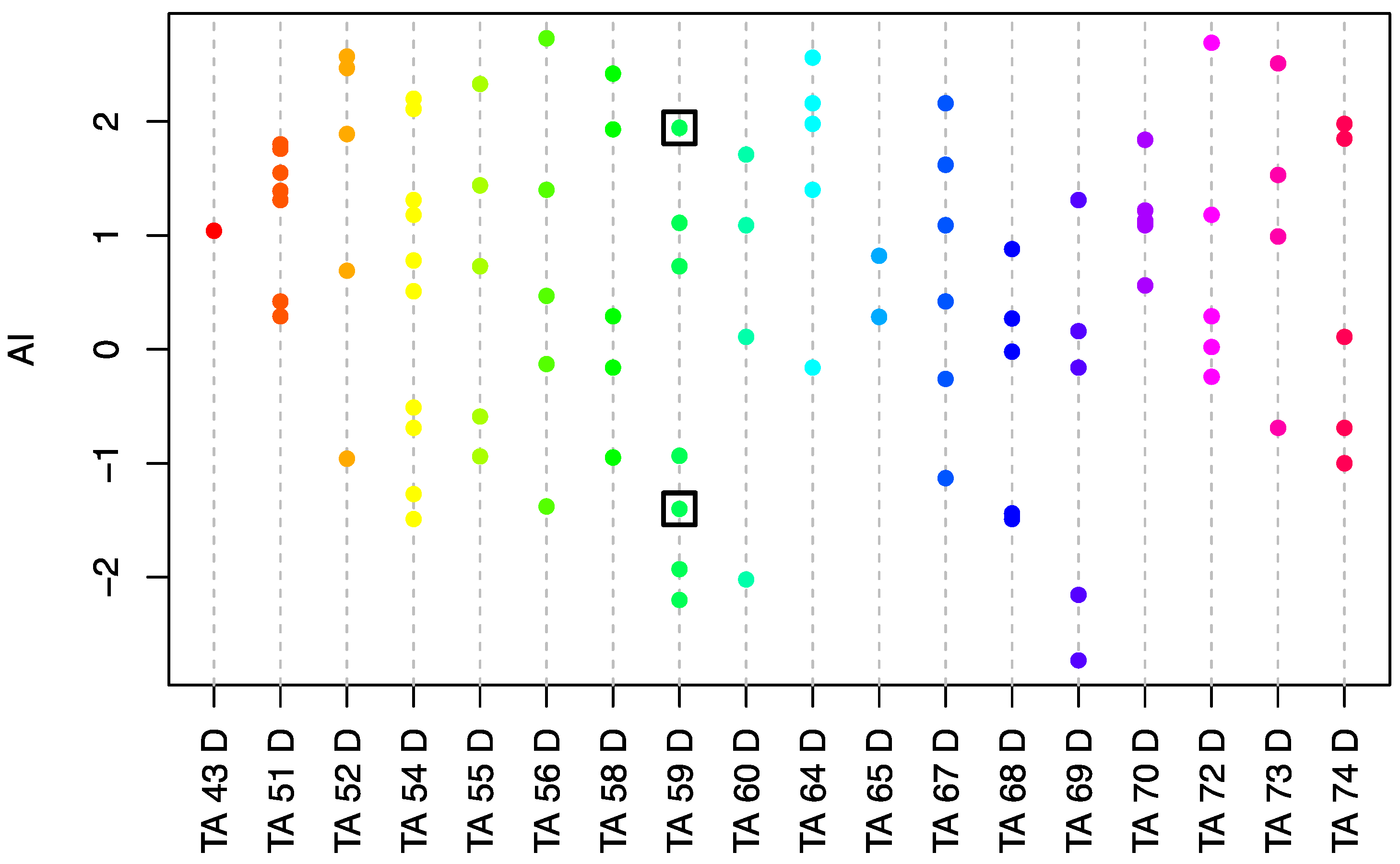
3.1. Abscission Phenotype Analysis of an E. oleifera Population
3.2. The Phenotypic Comparison of Fruit-Shedding and Non-Shedding Oleifera Palms
3.3. Gene Expression Profiling of AZ Samples from Normal Fruit-Shedding and Non-Shedding Variant Oleifera Palm
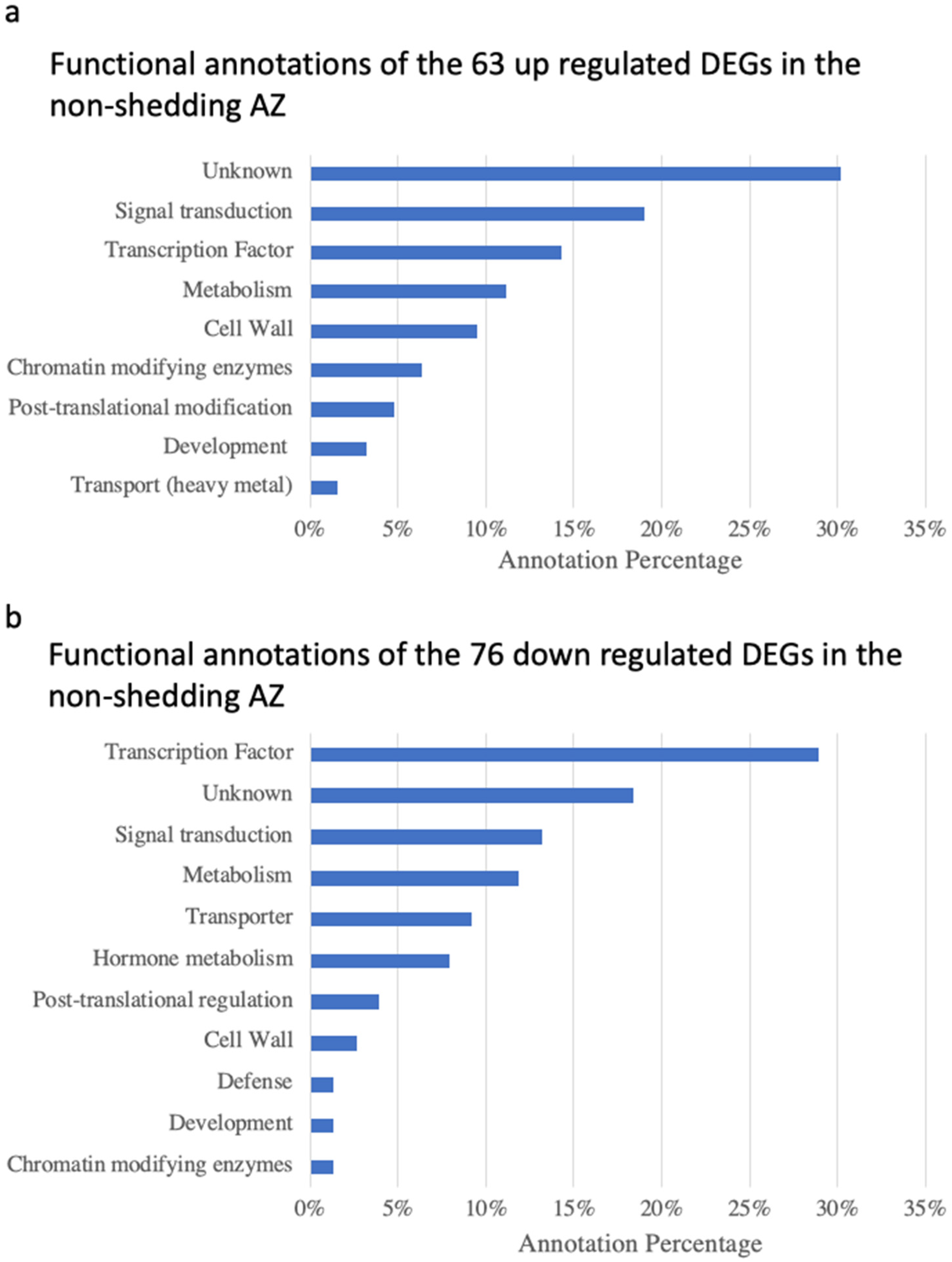
3.4. Functional Annotation of Differentially Expressed Genes and Identification of Enriched Categories
3.5. Comparison with E. guineensis Ripe Fruit AZ Transcriptome
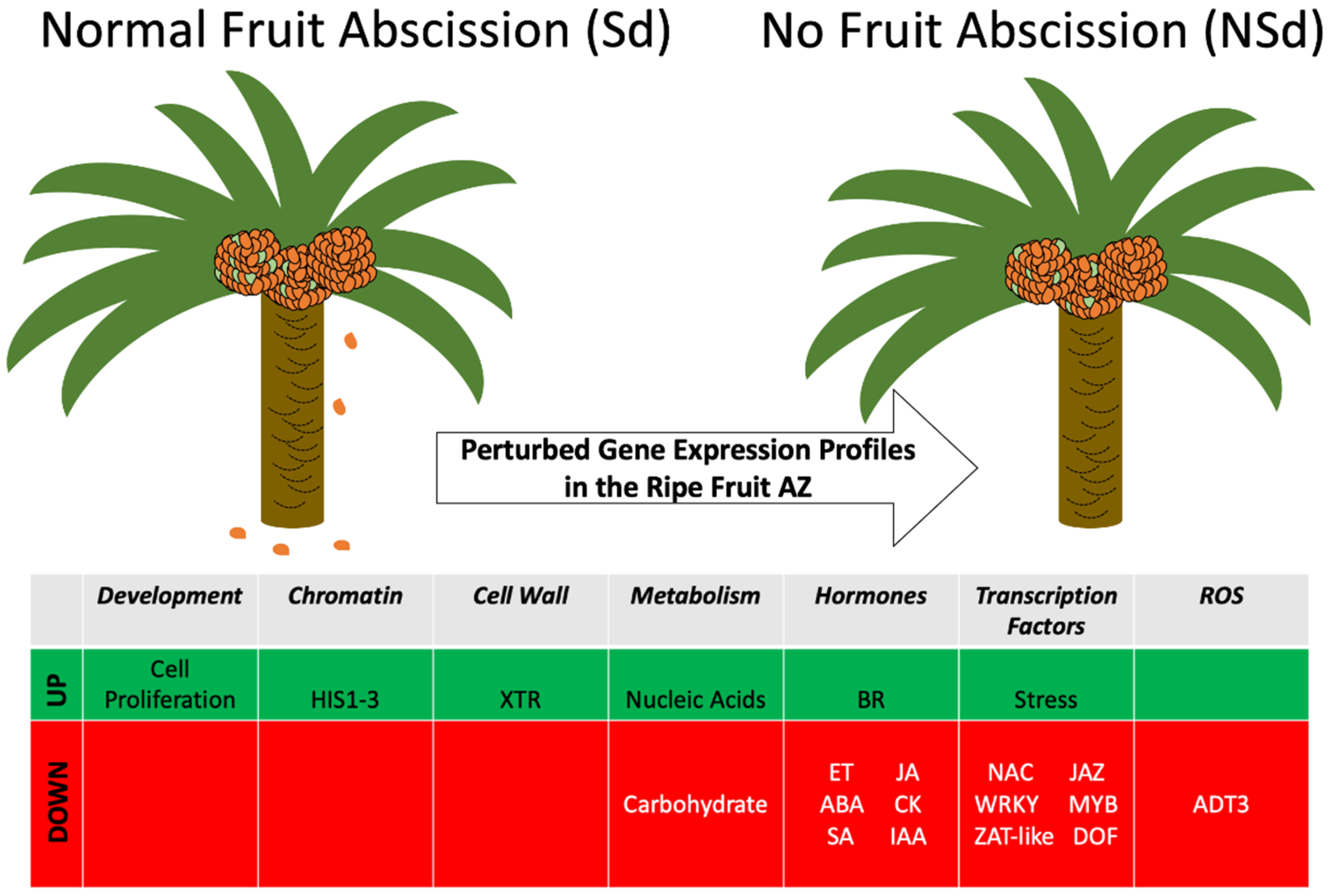
4. Discussion
4.1. Evidence for Transcription Factors and Hormone-Signaling Pathways Common to Organ Abscission Processes That Function during Oil Palm Fruit Abscission
4.2. Evidence That Changes in ROS Homeostasis and Hemicellulose Metabolism Are a Part of the Basis for the NSd Phenotype
4.3. A Comparison of E. oleifera and E. guineensis Suggests Abscission Processes Related to Environmental Stress and Epigenetic Modifications
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tranbarger, T.J.; Tadeo, F.R. Diversity and Functional Dynamics of Fleshy Fruit Abscission Zones. Annu. Plant Rev. Online 2020, 3, 1–64. [Google Scholar] [CrossRef]
- Tranbarger, T.J.; Tucker, M.L.; Roberts, J.A.; Meir, S. Editorial: Plant Organ Abscission: From Models to Crops. Front. Plant Sci. 2017, 8, 196. [Google Scholar] [CrossRef] [Green Version]
- Baker, W.J.; Couvreur, T. Global biogeography and diversification of palms sheds light on the evolution of tropical lineages. I. Historical biogeography. J. Biogeogr. 2012, 40, 274–285. [Google Scholar] [CrossRef]
- Meerow, A.W.; Noblick, L.; Salas-Leiva, D.E.; Sanchez, V.; Francisco-Ortega, J.; Jestrow, B.; Nakamura, K. Phylogeny and historical biogeography of the cocosoid palms (Arecaceae, Arecoideae, Cocoseae) inferred from sequences of six WRKY gene family loci. Cladistics 2015, 31, 509–534. [Google Scholar] [CrossRef]
- Montúfar, R.; Louise, C.; Tranbarger, T. Elaeis oleifera (Kunth) Cortés: A neglected palm from the Ecuadorian Amazon. Rev. Ecuat. Med. Cienc. Biol. 2018, 39, 11–18. [Google Scholar] [CrossRef]
- Fooyontphanich, K.; Morcillo, F.; Amblard, P.; Collin, M.; Jantasuriyarat, C.; Verdeil, J.-L.; Tangphatsornruang, S.; Tranbarger, T.J. A phenotypic test for delay of abscission and non-abscission oil palm fruit and validation by abscission marker gene expression analysis. Acta Hortic. 2016, 1119, 97–104. [Google Scholar] [CrossRef]
- Roongsattham, P.; Morcillo, F.; Jantasuriyarat, C.; Pizot, M.; Moussu, S.; Jayaweera, D.; Collin, M.; Gonzalez-Carranza, Z.H.; Amblard, P.; Tregear, J.W.; et al. Temporal and spatial expression of polygalacturonase gene family members reveals divergent regulation during fleshy fruit ripening and abscission in the monocot species oil palm. BMC Plant Biol. 2012, 12, 150. [Google Scholar] [CrossRef] [Green Version]
- Stø, I.M.; Orr, R.J.S.; Fooyontphanich, K.; Jin, X.; Knutsen, J.M.B.; Fischer, U.; Tranbarger, T.J.; Nordal, I.; Aalen, R.B. Conservation of the abscission signaling peptide IDA during Angiosperm evolution: Withstanding genome duplications and gain and loss of the receptors HAE/HSL. Front. Plant Sci. 2015, 6, 931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roongsattham, P.; Morcillo, F.; Fooyontphanich, K.; Jantasuriyarat, C.; Tragoonrung, S.; Amblard, P.; Collin, M.; Mouille, G.; Verdeil, J.-L.; Tranbarger, T.J. Cellular and Pectin Dynamics during Abscission Zone Development and Ripe Fruit Abscission of the Monocot Oil Palm. Front. Plant Sci. 2016, 7, 540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tranbarger, T.J.; Domonhédo, H.; Cazemajor, M.; Dubreuil, C.; Fischer, U.; Morcillo, F. The PIP Peptide of INFLORESCENCE DEFICIENT IN ABSCISSION Enhances Populus Leaf and Elaeis guineensis Fruit Abscission. Plants 2019, 8, 143. [Google Scholar] [CrossRef] [Green Version]
- Fooyontphanich, K.; Morcillo, F.; Joët, T.; Dussert, S.; Serret, J.; Collin, M.; Amblard, P.; Tangphatsornruang, S.; Roongsattham, P.; Jantasuriyarat, C.; et al. Multi-scale comparative transcriptome analysis reveals key genes and metabolic reprogramming processes associated with oil palm fruit abscission. BMC Plant Biol. 2021, 21, 1–18. [Google Scholar] [CrossRef]
- Estornell, L.H.; Agusti, J.; Merelo, P.; Talon, M.; Tadeo, F. Elucidating mechanisms underlying organ abscission. Plant Sci. 2013, 199–200, 48–60. [Google Scholar] [CrossRef]
- Meir, S.; Philosoph-Hadas, S.; Riov, J.; Tucker, M.L.; Patterson, S.E.; Roberts, J.A. Re-evaluation of the ethylene-dependent and -independent pathways in the regulation of floral and organ abscission. J. Exp. Bot. 2019, 70, 1461–1467. [Google Scholar] [CrossRef]
- Patharkar, O.R.; Walker, J.C. Core Mechanisms Regulating Developmentally Timed and Environmentally Triggered Abscission. Plant Physiol. 2016, 172, 510–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meir, S.; Sundaresan, S.; Riov, J.; Agarwal, I.; Philosoph-Hadas, S. Role of auxin depletion in abscission control. Stewart Postharvest Rev. 2015, 2, 1–15. [Google Scholar]
- Tisné, S.; Denis, M.; Domonhédo, H.; Pallas, B.; Cazemajor, M.; Tranbarger, T.J.; Morcillo, F. Environmental and trophic determinism of fruit abscission and outlook with climate change in tropical regions. Plant Environ. Interact. 2020, 1, 17–28. [Google Scholar] [CrossRef] [Green Version]
- Argout, X.; Fouet, O.; Wincker, P.; Gramacho, K.; Legavre, T.; Sabau, X.; Risterucci, A.M.; Da Silva, C.; Cascardo, J.; Allegre, M.; et al. Towards the understanding of the cocoa transcriptome: Production and analysis of an exhaustive dataset of ESTs of Theobroma cacao L. generated from various tissues and under various conditions. BMC Genom. 2008, 9, 512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H. Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. arXiv 2013, arXiv:1303.3997. [Google Scholar]
- Singh, R.; Ong-Abdullah, M.; Low, E.-T.L.; Manaf, M.A.A.; Rosli, R.; Nookiah, R.; Ooi, L.C.-L.; Ooi, S.; Chan, K.-L.; Halim, M.A.; et al. Oil palm genome sequence reveals divergence of interfertile species in old and new worlds. Nature 2013, 500, 335–339. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Lohse, M.; Nagel, A.; Herter, T.; May, P.; Schroda, M.; Zrenner, R.; Tohge, T.; Fernie, A.R.; Stitt, M.; Usadel, B. Mercator: A fast and simple web server for genome scale functional annotation of plant sequence data. Plant Cell Environ. 2013, 37, 1250–1258. [Google Scholar] [CrossRef] [Green Version]
- Butler, L. Inherited characters in the tomato. II. Jointless pedicel. J. Hered. 1936, 27, 25–26. [Google Scholar]
- McKim, S.; Stenvik, G.-E.; Butenko, M.A.; Kristiansen, W.; Cho, S.K.; Hepworth, S.; Aalen, R.B.; Haughn, G.W. The BLADE-ON-PETIOLE genes are essential for abscission zone formation in Arabidopsis. Developement 2008, 135, 1537–1546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Illgen, S.; Zintl, S.; Zuther, E.; Hincha, D.K.; Schmülling, T. Characterisation of the ERF102 to ERF105 genes of Arabidopsis thaliana and their role in the response to cold stress. Plant Mol. Biol. 2020, 103, 303–320. [Google Scholar] [CrossRef] [Green Version]
- Bolt, S.; Zuther, E.; Zintl, S.; Hincha, D.K.; Schmülling, T. ERF105is a transcription factor gene ofArabidopsis thalianarequired for freezing tolerance and cold acclimation. Plant Cell Environ. 2016, 40, 108–120. [Google Scholar] [CrossRef]
- Ascenzi, R.; Gantt, J.S. A drought-stress-inducible histone gene in Arabidopsis thaliana is a member of a distinct class of plant linker histone variants. Plant Mol. Biol. 1997, 34, 629–641. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.S.; Takahashi, H.; Miyazaki, A.; Yamaguchi, K.; Kusano, T. Identification of the cis-acting elements in Arabidopsis thaliana NHL10 promoter responsible for leaf senescence, the hypersensitive response against Cucumber mosaic virus infection, and spermine treatment. Plant Sci. 2005, 168, 415–422. [Google Scholar] [CrossRef]
- Zheng, M.S.; Takahashi, H.; Miyazaki, A.; Hamamoto, H.; Yamaguchi, I.; Kusano, T.; Shah, J. Up-regulation of Arabidopsis thaliana NHL10 in the hypersensitive response to Cucumber mosaic virus infection and in senescing leaves is controlled by signalling pathways that differ in salicylate involvement. Planta 2004, 218, 740–750. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, S.; Ito, M.; Nishida, I.; Watanabe, A. Isolation and RNA Gel Blot Analysis of Genes that Could Serve as Potential Molecular Markers for Leaf Senescence in Arabidopsis thaliana. Plant Cell Physiol. 2001, 42, 170–178. [Google Scholar] [CrossRef]
- Rubio, M.C.; Calvo-Begueria, L.; Diaz-Mendoza, M.; Elhiti, M.; Moore, M.; Matamoros, M.A.; James, E.K.; Díaz, I.; Pérez-Rontomé, C.; Villar, I.; et al. Phytoglobins in the nuclei, cytoplasm and chloroplasts modulate nitric oxide signaling and interact with abscisic acid. Plant J. 2019, 100, 38–54. [Google Scholar] [CrossRef]
- Shin, K.; Lee, S.; Song, W.-Y.; Lee, R.-A.; Lee, I.; Ha, K.; Koo, J.-C.; Park, S.-K.; Nam, H.-G.; Lee, Y.; et al. Genetic Identification of Acc-Resistant2 Reveals Involvement of Lysine Histidine Transporter1 in the Uptake of 1-Aminocyclopropane-1-Carboxylic Acid in Arabidopsis thaliana. Plant Cell Physiol. 2015, 56, 572–582. [Google Scholar] [CrossRef] [Green Version]
- Glazinska, P.; Wojciechowski, W.; Kulasek, M.; Glinkowski, W.; Marciniak, K.; Klajn, N.; Kęsy, J.; Kopcewicz, J. De novo Transcriptome Profiling of Flowers, Flower Pedicels and Pods of Lupinus luteus (Yellow Lupine) Reveals Complex Expression Changes during Organ Abscission. Front. Plant Sci. 2017, 8, 641. [Google Scholar] [CrossRef] [Green Version]
- Niederhuth, C.E.; Patharkar, O.R.; Walker, J.C. Transcriptional profiling of the Arabidopsis abscission mutant hae hsl2by RNA-Seq. BMC Genom. 2013, 14, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, Z.; Wen, Z.; Hou, Q.; Qiao, G.; Yang, K.; Hong, Y.; Wen, X. Cross-talk between transcriptome, phytohormone and HD-ZIP gene family analysis illuminates the molecular mechanism underlying fruitlet abscission in sweet cherry (Prunus avium L). BMC Plant Biol. 2021, 21, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Bharti, N.; Singh, A.P.; Tripathi, S.K.; Pandey, S.P.; Chauhan, A.S.; Kulkarni, A.; Sane, A.P. Petal abscission in fragrant roses is associated with large scale differential regulation of the abscission zone transcriptome. Sci. Rep. 2020, 10, 1–16. [Google Scholar] [CrossRef]
- Sriskantharajah, K.; El Kayal, W.; Torkamaneh, D.; Ayyanath, M.M.; Saxena, P.K.; Sullivan, A.J.; Paliyath, G.; Subramanian, J. Transcriptomics of Improved Fruit Retention by Hexanal in ‘Honeycrisp’ Reveals Hormonal Crosstalk and Reduced Cell Wall Degradation in the Fruit Abscission Zone. Int. J. Mol. Sci. 2021, 22, 8830. [Google Scholar] [CrossRef]
- Whitelaw, C.A.; Lyssenko, N.N.; Chen, L.; Zhou, D.; Mattoo, A.K.; Tucker, M.L. Delayed Abscission and Shorter Internodes Correlate with a Reduction in the Ethylene Receptor LeETR1 Transcript in Transgenic Tomato. Plant Physiol. 2002, 128, 978–987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meir, S.; Hunter, D.; Chen, J.-C.; Halaly, V.; Reid, M.S. Molecular Changes Occurring during Acquisition of Abscission Competence following Auxin Depletion in Mirabilis jalapa. Plant Physiol. 2006, 141, 1604–1616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakano, T.; Fujisawa, M.; Shima, Y.; Ito, Y. Expression profiling of tomato pre-abscission pedicels provides insights into abscission zone properties including competence to respond to abscission signals. BMC Plant Biol. 2013, 13, 40. [Google Scholar] [CrossRef] [Green Version]
- Sundaresan, S.; Philosoph-Hadas, S.; Ma, C.; Jiang, C.; Riov, J.; Kochanek, B.; Salim, S.; Reid, M.S.; Meir, S. Role of the KNOTTED1-LIKE HOMEOBOX protein (KD1) in regulating abscission of tomato flower pedicels at early and late stages of the process. Physiol. Plant. 2021. [Google Scholar] [CrossRef] [PubMed]
- Sundaresan, S.; Philosoph-Hadas, S.; Ma, C.; Jiang, C.-Z.; Riov, J.; Mugasimangalam, R.; Kochanek, B.; Salim, S.; Reid, M.S.; Meir, S. The Tomato Hybrid Proline-rich Protein regulates the abscission zone competence to respond to ethylene signals. Hortic. Res. 2018, 5, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Xu, P.; Chen, H.; Cai, W. Transcription factor CDF4 promotes leaf senescence and floral organ abscission by regulating abscisic acid and reactive oxygen species pathways in Arabidopsis. EMBO Rep. 2020, 21, e48967. [Google Scholar] [CrossRef]
- Gonzalez-Carranza, Z.H.; Whitelaw, C.A.; Swarup, R.; Roberts, J.A. Temporal and spatial expression of a polygalacturonase during leaf and flower abscission in oilseed rape and Arabidopsis. Plant Physiol. 2002, 128, 534–543. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Xu, Y.-H.; Jiang, S.-C.; Lu, K.; Lu, Y.-F.; Feng, X.-J.; Wu, Z.; Liang, S.; Yu, Y.-T.; Wang, X.-F.; et al. Light-harvesting chlorophyll a/b-binding proteins, positively involved in abscisic acid signalling, require a transcription repressor, WRKY40, to balance their function. J. Exp. Bot. 2013, 64, 5443–5456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, Y.; Yan, L.; Liu, Z.-Q.; Cao, Z.; Mei, C.; Xin, Q.; Wu, F.-Q.; Wang, X.-F.; Du, S.-Y.; Jiang, T.; et al. The Mg-Chelatase H Subunit of Arabidopsis Antagonizes a Group of WRKY Transcription Repressors to Relieve ABA-Responsive Genes of Inhibition. Plant Cell 2010, 22, 1909–1935. [Google Scholar] [CrossRef] [Green Version]
- Kang, J.; Yim, S.; Choi, H.; Kim, A.; Lee, K.P.; Lopez-Molina, L.; Martinoia, E.; Lee, Y. Abscisic acid transporters cooperate to control seed germination. Nat. Commun. 2015, 6, 8113. [Google Scholar] [CrossRef] [Green Version]
- Ghorbel, M.; Brini, F.; Sharma, A.; Landi, M. Role of jasmonic acid in plants: The molecular point of view. Plant Cell Rep. 2021, 40, 1471–1494. [Google Scholar] [CrossRef] [PubMed]
- von Malek, B.; van der Graaff, E.; Schneitz, K.; Keller, B. The Arabidopsis male-sterile mutant dde2-2 is defective in the ALLENE OXIDE SYNTHASE gene encoding one of the key enzymes of the jasmonic acid biosynthesis pathway. Planta 2002, 216, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Kim, J. Four shades of detachment: Regulation of floral organ abscission. Plant Signal. Behav. 2014, 9, e976154. [Google Scholar] [CrossRef] [Green Version]
- Figueroa, P.; Browse, J. The Arabidopsis JAZ2 Promoter Contains a G-Box and Thymidine-Rich Module that are Necessary and Sufficient for Jasmonate-Dependent Activation by MYC Transcription Factors and Repression by JAZ Proteins. Plant Cell Physiol. 2011, 53, 330–343. [Google Scholar] [CrossRef] [Green Version]
- Leone, M.; Keller, M.M.; Cerrudo, I.; Ballaré, C.L. To grow or defend? Low red: Far-red ratios reduce jasmonate sensitivity in Arabidopsis seedlings by promoting DELLA degradation and increasing JAZ10 stability. New Phytol. 2014, 204, 355–367. [Google Scholar] [CrossRef] [PubMed]
- Major, I.T.; Guo, Q.; Zhai, J.; Kapali, G.; Kramer, D.M.; Howe, G.A. A Phytochrome B-Independent Pathway Restricts Growth at High Levels of Jasmonate Defense. Plant Physiol. 2020, 183, 733–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki-Sekimoto, Y.; Jikumaru, Y.; Obayashi, T.; Saito, H.; Masuda, S.; Kamiya, Y.; Ohta, H.; Shirasu, K. Basic Helix-Loop-Helix Transcription Factors JASMONATE-ASSOCIATED MYC2-LIKE1 (JAM1), JAM2, and JAM3 Are Negative Regulators of Jasmonate Responses in Arabidopsis. Plant Physiol. 2013, 163, 291–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoang, N.V.; Choe, G.; Zheng, Y.; Fandino, A.C.A.; Sung, I.; Hur, J.; Kamran, M.; Park, C.; Kim, H.; Ahn, H.; et al. Identification of Conserved Gene-Regulatory Networks that Integrate Environmental Sensing and Growth in the Root Cambium. Curr. Biol. 2020, 30, 2887.e7–2900.e7. [Google Scholar] [CrossRef]
- Butenko, M.A.; Simon, R. Beyond the meristems: Similarities in the CLAVATA3 and INFLORESCENCE DEFICIENT IN ABSCISSION peptide mediated signalling pathways. J. Exp. Bot. 2015, 66, 5195–5203. [Google Scholar] [CrossRef] [Green Version]
- Patharkar, O.R.; Walker, J.C. Connections between abscission, dehiscence, pathogen defense, drought tolerance, and senescence. Plant Sci. 2019, 284, 25–29. [Google Scholar] [CrossRef]
- Bethke, G.; Unthan, T.; Uhrig, J.F.; Pöschl, Y.; Gust, A.A.; Scheel, D.; Lee, J. Flg22 regulates the release of an ethylene response factor substrate from MAP kinase 6 in Arabidopsis thaliana via ethylene signaling. Proc. Natl. Acad. Sci. USA 2009, 106, 8067–8072. [Google Scholar] [CrossRef] [Green Version]
- Camehl, I.; Oelmüller, R. Do ethylene response factorS9 and -14 repress PR gene expression in the interaction between Piriformospora indica and Arabidopsis? Plant Signal. Behav. 2010, 5, 932–936. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Meng, Y.; Zhang, K.; Li, Q.; Li, S.; Xu, B.; Georgiev, M.I.; Zhou, M. Jasmonic acid-responsive RRTF1 transcription factor controls DTX18 gene expression in hydroxycinnamic acid amide secretion. Plant Physiol. 2021, 185, 369–384. [Google Scholar] [CrossRef]
- Maruyama, Y.; Yamoto, N.; Suzuki, Y.; Chiba, Y.; Yamazaki, K.-I.; Sato, T.; Yamaguchi, J. The Arabidopsis transcriptional repressor ERF9 participates in resistance against necrotrophic fungi. Plant Sci. 2013, 213, 79–87. [Google Scholar] [CrossRef]
- Nakai, Y.; Nakahira, Y.; Sumida, H.; Takebayashi, K.; Nagasawa, Y.; Yamasaki, K.; Akiyama, M.; Ohme-Takagi, M.; Fujiwara, S.; Shiina, T.; et al. Vascular plant one-zinc-finger protein 1/2 transcription factors regulate abiotic and biotic stress responses in Arabidopsis. Plant J. 2012, 73, 761–775. [Google Scholar] [CrossRef]
- Sharabi-Schwager, M.; Samach, A.; Porat, R. Overexpression of theCBF2transcriptional activator inArabidopsissuppresses the responsiveness of leaf tissue to the stress hormone ethylene. Plant Biol. 2009, 12, 630–638. [Google Scholar] [CrossRef]
- Yin, X.-R.; Xie, X.-L.; Xia, X.-J.; Yu, J.-Q.; Ferguson, I.B.; Giovannoni, J.J.; Chen, K.-S. Involvement of an ethylene response factor in chlorophyll degradation during citrus fruit degreening. Plant J. 2016, 86, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Barbez, E.; Kubeš, M.; Rolčík, J.; Béziat, C.; Pěnčík, A.; Wang, B.; Rosquete, M.R.; Zhu, J.; Dobrev, P.I.; Lee, Y.; et al. A novel putative auxin carrier family regulates intracellular auxin homeostasis in plants. Nat. Cell Biol. 2012, 485, 119–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinoshita, N.; Wang, H.; Kasahara, H.; Liu, J.; MacPherson, C.; Machida, Y.; Kamiya, Y.; Hannah, M.; Chua, N.-H. IAA-Ala Resistant3, an Evolutionarily Conserved Target of miR167, Mediates Arabidopsis Root Architecture Changes during High Osmotic Stress. Plant Cell 2012, 24, 3590–3602. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, V.; Parasuraman, B.; Laxmi, A.; Chattopadhyay, D. CIPK6, a CBL-interacting protein kinase is required for development and salt tolerance in plants. Plant J. 2009, 58, 778–790. [Google Scholar] [CrossRef]
- Benjamins, R.; Ampudia, C.S.G.; Hooykaas, P.; Offringa, R. PINOID-Mediated Signaling Involves Calcium-Binding Proteins. Plant Physiol. 2003, 132, 1623–1630. [Google Scholar] [CrossRef] [Green Version]
- Woo, H.-H.; Jeong, B.R.; Hirsch, A.M.; Hawes, M.C. Characterization of Arabidopsis AtUGT85A and AtGUS gene families and their expression in rapidly dividing tissues. Genomics 2007, 90, 143–153. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhao, L.; Zhao, J.; Li, Y.; Wang, J.; Guo, R.; Gan, S.; Liu, C.-J.; Zhang, K. S5H/DMR6 Encodes a Salicylic Acid 5-Hydroxylase That Fine-Tunes Salicylic Acid Homeostasis. Plant Physiol. 2017, 175, 1082–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schröder, F.; Lisso, J.; Lange, P.; Müssig, C. The extracellular EXO protein mediates cell expansion in Arabidopsis leaves. BMC Plant Biol. 2009, 9, 20. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Yuan, Y.; Li, C.; Wu, Q.; He, Z.; Li, J.; Zhao, M. Brassinosteroids suppress ethylene-induced fruitlet abscission through LcBZR1/2-mediated transcriptional repression of LcACS1/4 and LcACO2/3 in litchi. Hortic. Res. 2021, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Jin, D.; Wang, X.; Xu, Y.; Gui, H.; Zhang, H.; Dong, Q.; Sikder, R.K.; Yang, G.; Song, M. Chemical Defoliant Promotes Leaf Abscission by Altering ROS Metabolism and Photosynthetic Efficiency in Gossypium hirsutum. Int. J. Mol. Sci. 2020, 21, 2738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y. More than cell wall hydrolysis: Orchestration of cellular dynamics for organ separation. Curr. Opin. Plant Biol. 2019, 51, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Yoon, T.H.; Lee, J.; Jeon, S.Y.; Lee, J.H.; Lee, M.K.; Chen, H.; Yun, J.; Oh, S.Y.; Wen, X.; et al. A Lignin Molecular Brace Controls Precision Processing of Cell Walls Critical for Surface Integrity in Arabidopsis. Cell 2018, 173, 1468–1480.e9. [Google Scholar] [CrossRef] [Green Version]
- Liao, W.; Wang, G.; Li, Y.; Wang, B.; Zhang, P.; Peng, M. Reactive oxygen species regulate leaf pulvinus abscission zone cell separation in response to water-deficit stress in cassava. Sci. Rep. 2016, 6, 21542. [Google Scholar] [CrossRef] [Green Version]
- Wilmowicz, E.; Kućko, A.; Pokora, W.; Kapusta, M.; Jasieniecka-Gazarkiewicz, K.; Tranbarger, T.; Wolska, M.; Panek, K. EPIP-Evoked Modifications of Redox, Lipid, and Pectin Homeostasis in the Abscission Zone of Lupine Flowers. Int. J. Mol. Sci. 2021, 22, 3001. [Google Scholar] [CrossRef]
- Yang, Z.; Zhong, X.; Fan, Y.; Wang, H.; Li, J.; Huang, X. Burst of reactive oxygen species in pedicel-mediated fruit abscission after carbohydrate supply was cut off in longan (Dimocarpus longan). Front. Plant Sci. 2015, 6, 360. [Google Scholar] [CrossRef] [Green Version]
- Mhamdi, A.; Van Breusegem, F. Reactive oxygen species in plant development. Development 2018, 145, dev164376. [Google Scholar] [CrossRef] [Green Version]
- Para, A.; Muhammad, D.; Orozco-Nunnelly, D.A.; Memishi, R.; Alvarez, S.; Naldrett, M.J.; Warpeha, K.M. The dehydratase ADT3 affects ROS homeostasis and cotyledon development. Plant Physiol. 2016, 172, 1045–1060. [Google Scholar] [CrossRef] [Green Version]
- Dong, J.; Kim, S.T.; Lord, E.M. Plantacyanin Plays a Role in Reproduction in Arabidopsis. Plant Physiol. 2005, 138, 778–789. [Google Scholar] [CrossRef] [Green Version]
- Mahmood, K.; El-Kereamy, A.; Kim, S.-H.; Nambara, E.; Rothstein, S.J. ANAC032 Positively Regulates Age-Dependent and Stress-Induced Senescence inArabidopsis thaliana. Plant Cell Physiol. 2016, 57, 2029–2046. [Google Scholar] [CrossRef] [Green Version]
- Eklöf, J.M.; Brumer, H. The XTH Gene Family: An Update on Enzyme Structure, Function, and Phylogeny in Xyloglucan Remodeling. Plant Physiol. 2010, 153, 456–466. [Google Scholar] [CrossRef] [Green Version]
- Stratilová, B.; Kozmon, S.; Stratilová, E.; Hrmova, M. Plant Xyloglucan Xyloglucosyl Transferases and the Cell Wall Structure: Subtle but Significant. Molecules 2020, 25, 5619. [Google Scholar] [CrossRef]
- Le Gall, H.; Philippe, F.; Domon, J.-M.; Gillet, F.; Pelloux, J.; Rayon, C. Cell Wall Metabolism in Response to Abiotic Stress. Plants 2015, 4, 112–166. [Google Scholar] [CrossRef]
- Ma, X.; Li, C.; Yuan, Y.; Zhao, M.; Li, J. Xyloglucan endotransglucosylase/hydrolase genes LcXTH4 /7/19 are involved in fruitlet abscission and are activated by LcEIL2 /3 in litchi. Physiol. Plant. 2021, 173, 1136–1146. [Google Scholar] [CrossRef]
- Xu, P.; Fang, S.; Chen, H.; Cai, W. The brassinosteroid-responsive xyloglucan endotransglucosylase/hydrolase 19 (XTH19) and XTH23 genes are involved in lateral root development under salt stress in Arabidopsis. Plant J. 2020, 104, 59–75. [Google Scholar] [CrossRef] [PubMed]
- Rutowicz, K.; Puzio, M.; Halibart-Puzio, J.; Lirski, M.; Kroteń, M.A.; Kotliński, M.; Knizewski, L.; Lange, B.; Muszewska, A.; Śniegowska-Świerk, K.; et al. A specialized histone H1 variant is required for adaptive responses to complex abiotic stress and related DNA methylation in Arabidopsis. Plant Physiol. 2015, 169, 2080–2101. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Eg Locus | TAIR9 Accession | Best Manual Annotation | Gene Name | References | Related Function |
|---|---|---|---|---|---|
| LOC105043733 | AT5G51190 | ERF (ethylene response factor 105) subfamily B-3 of ERF/AP2 transcription factor family | ERF104 | [25,26] | Response to cold stress |
| LOC105059363 | AT2G18050 | Histone H1-3 | HIS1-3 | [27] | Response to drought stress; senescent leaves |
| LOC105060420 | AT2G35980 | Late embryogenesis abundant (LEA)/YELLOW-LEAF-SPECIFIC GENE 9 | YLS9/NHL10 | [28,29,30] | Response to salt stress; senescent leaves; hypersensitive response |
| LOC105033708 | AT2G16060 | Nonsymbiotic hemoglobin 1 | HB1/GLB1 | [31] | Nitric oxide (NO) homeostasis and interacts with ABA processes of plants (e.g., response to desiccation) |
| LOC105044389 | AT1G07540 | Telomere-binding protein | TRFL2 | - | - |
| LOC105053311 | AT5G40780 | Lysine histidine transporter 1 | LHT1 | [32] | Transport of ACC and ACC-induced ethylene responses in A. thaliana |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morcillo, F.; Serret, J.; Beckers, A.; Collin, M.; Tisné, S.; George, S.; Poveda, R.; Louise, C.; Tranbarger, T.J. A Non-Shedding Fruit Elaeis oleifera Palm Reveals Perturbations to Hormone Signaling, ROS Homeostasis, and Hemicellulose Metabolism. Genes 2021, 12, 1724. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12111724
Morcillo F, Serret J, Beckers A, Collin M, Tisné S, George S, Poveda R, Louise C, Tranbarger TJ. A Non-Shedding Fruit Elaeis oleifera Palm Reveals Perturbations to Hormone Signaling, ROS Homeostasis, and Hemicellulose Metabolism. Genes. 2021; 12(11):1724. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12111724
Chicago/Turabian StyleMorcillo, Fabienne, Julien Serret, Antoine Beckers, Myriam Collin, Sebastien Tisné, Simon George, Roberto Poveda, Claude Louise, and Timothy John Tranbarger. 2021. "A Non-Shedding Fruit Elaeis oleifera Palm Reveals Perturbations to Hormone Signaling, ROS Homeostasis, and Hemicellulose Metabolism" Genes 12, no. 11: 1724. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12111724