
Harnessing Crop Wild Diversity for Climate Change Adaptation



1
Corporación Colombiana de Investigación Agropecuaria AGROSAVIA, C.I. La Selva, Km 7 Vía Rionegro, Las Palmas, Rionegro 054048, Colombia
2
Departamento de Ciencias Forestales, Facultad de Ciencias Agrarias, Universidad Nacional de Colombia, Sede Medellín, Medellín 050034, Colombia
*
Author to whom correspondence should be addressed.
Genes 2021, 12(5), 783; https://0-doi-org.brum.beds.ac.uk/10.3390/genes12050783
Submission received: 29 March 2021
/
Revised: 28 April 2021
/
Accepted: 19 May 2021
/
Published: 20 May 2021
(This article belongs to the Special Issue Evolutionary Genetics of Plant Crop-Wild Complexes: From Fundamental to Applied Research)
Abstract
:Warming and drought are reducing global crop production with a potential to substantially worsen global malnutrition. As with the green revolution in the last century, plant genetics may offer concrete opportunities to increase yield and crop adaptability. However, the rate at which the threat is happening requires powering new strategies in order to meet the global food demand. In this review, we highlight major recent ‘big data’ developments from both empirical and theoretical genomics that may speed up the identification, conservation, and breeding of exotic and elite crop varieties with the potential to feed humans. We first emphasize the major bottlenecks to capture and utilize novel sources of variation in abiotic stress (i.e., heat and drought) tolerance. We argue that adaptation of crop wild relatives to dry environments could be informative on how plant phenotypes may react to a drier climate because natural selection has already tested more options than humans ever will. Because isolated pockets of cryptic diversity may still persist in remote semi-arid regions, we encourage new habitat-based population-guided collections for genebanks. We continue discussing how to systematically study abiotic stress tolerance in these crop collections of wild and landraces using geo-referencing and extensive environmental data. By uncovering the genes that underlie the tolerance adaptive trait, natural variation has the potential to be introgressed into elite cultivars. However, unlocking adaptive genetic variation hidden in related wild species and early landraces remains a major challenge for complex traits that, as abiotic stress tolerance, are polygenic (i.e., regulated by many low-effect genes). Therefore, we finish prospecting modern analytical approaches that will serve to overcome this issue. Concretely, genomic prediction, machine learning, and multi-trait gene editing, all offer innovative alternatives to speed up more accurate pre- and breeding efforts toward the increase in crop adaptability and yield, while matching future global food demands in the face of increased heat and drought. In order for these ‘big data’ approaches to succeed, we advocate for a trans-disciplinary approach with open-source data and long-term funding. The recent developments and perspectives discussed throughout this review ultimately aim to contribute to increased crop adaptability and yield in the face of heat waves and drought events.
1. Introduction—The Imminence of the Threat
How plants and crops will respond to a warmer and drier climate is currently one of the most discussed multi-disciplinary questions in the fields of environmental science, ecology, and evolution. It is estimated that climate change effects may limit global crop production by at least 10% in 2050 [1,2], especially in vulnerable regions around the globe where heat, drought, and malnutrition are already substantial. Given that current agricultural resources might not be sufficient to meet future food demand [3], crop wild relatives and landraces historically adapted to dry and semi-arid environments are key sources of yet unexplored diversity with respect to major food crops [4]. Exotics may donate necessary genetic variation to make heat and drought-tolerant cultivars, or may even stand as novel crops by themselves [5,6,7] (e.g., Lupinus mutabilis [8]).
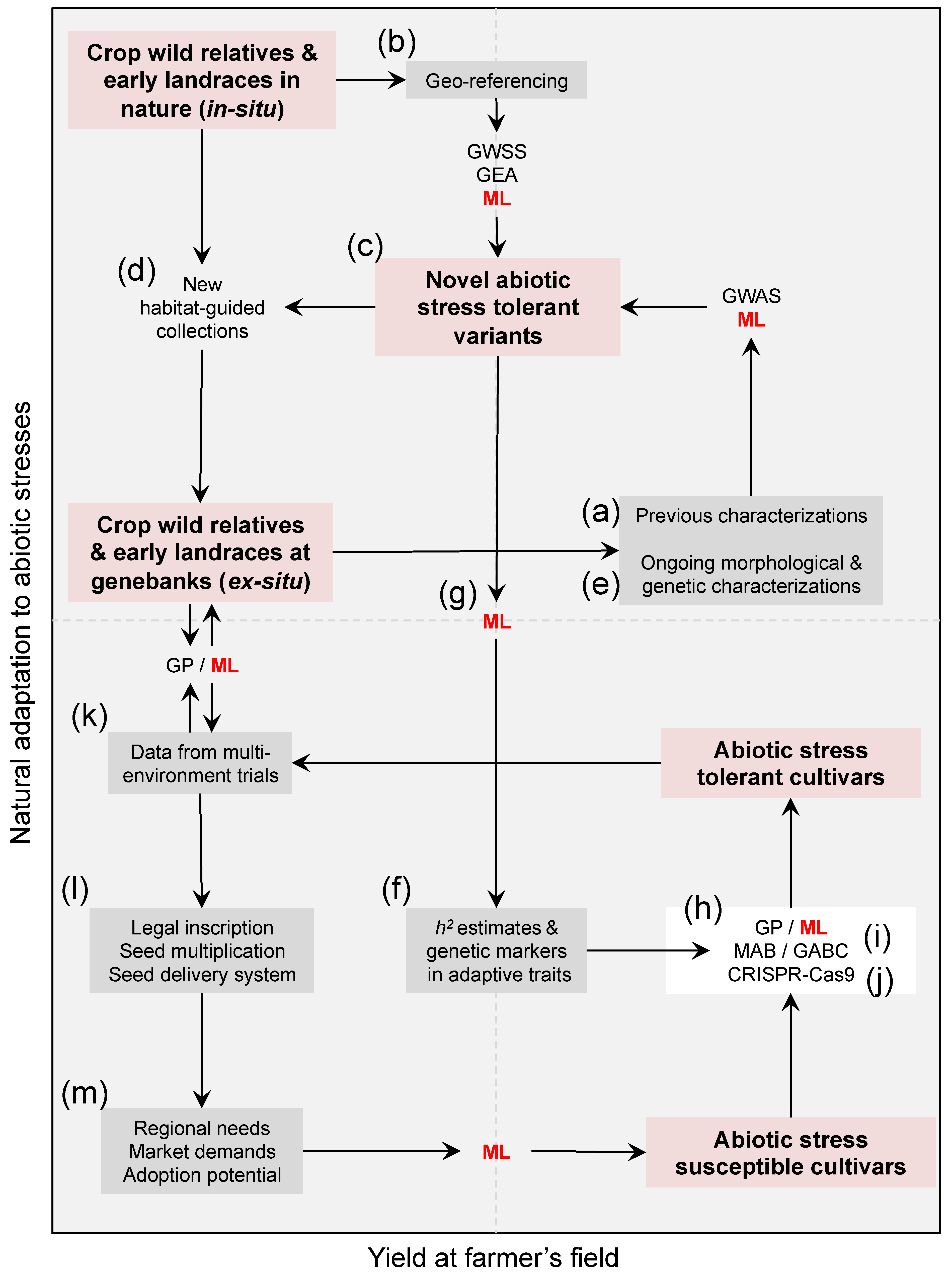
However, efforts to capture and pyramid target tolerant variants from the wild still face major challenges. First, identifying useful variation in wild accessions through field trials (Figure 1a) has been inefficient because of the disparity in growth rates and phenologies [9]. This limitation is reinforced by a complex inheritance of the abiotic stress tolerant phenotypes involving many genes of low effects and several environmental interactions [10]. Second, the domestication syndrome typically neglected heat and drought tolerance in the majority of crop species [11,12] because these adaptations in wild populations (Figure 1b) tend to perpetuate vegetative phases, delay reproductive stages, and therefore compromise overall yield [13]. Due to this, transferring variation from wild exotic donors into elite lines may induce undesired linked trait variation such as flowering delay and reduced crop yields (e.g., in Phaseolus species [14]).
With the advent of the genomic era, heat and drought tolerance molecular pathways have been extensively studied [15,16,17], and numerous candidate genes and genetic marker associations have already been identified and validated (e.g., ABA-dependent or independent pathways, and ASR, DREB, and ERECTA-encoding genes [18,19,20,21]). Coupling these comprehensive datasets with novel analytical tools harbors the potential to identify and unlock useful genetic variation among crop wild relatives and landraces to challenge with abiotic stresses. In the following sections, we outline the main avenues to establish novel sources of abiotic stress tolerance variation from wild crop relatives and semi-domesticated landraces (Figure 1c), and discuss modern genomic-assisted strategies to utilize these variants to obtain heat and drought-tolerant elite crop lines. We finish by emphasizing the need to enable the construction and long-term maintenance of big heterogeneous dataset repositories capable of powering these innovative predictive strategies.
2. Valuing the Wild—Strategies to Identify Naturally Available Exotic Variants
Domestication has been the most ambitious evolutionary experiment humanity has ever embarked on [11,22]. Being the longest running selection trial [23], it has served as a playground for geneticists and biologists to explore the patterns and processes during crop evolution [24,25,26]. For instance, it has allowed humans to test hypothesis such as whether dual domestication syndromes have recruited the same genetic variants in parallel [27], or whether genomic divergence is more prone to harbor signatures of selection due to reduced recombination and increased drift [28]. In this way, domestication has invited researchers to study the repeatability of evolution [29], and the relative role of isolation, migration, and hybridization [30], long standing questions in evolutionary biology [31,32,33]. Similarly, studying crop evolution has proven enlightening, and addressing their wild relatives’ natural adaptation to distinct habitats (Figure 1b) informs on how plant phenotypes may react to a changing climate [34]. After all, natural selection has already tested more options than humans ever will [35].
2.1. On the Necessity of Broadening the Germplasm
Natural selection rarely misses key adaptive improvements. For instance, it has already improved the efficiency of photosynthesis and water use [35], which are major developments to enhance the yield potential that has increased little in recent decades. Therefore, inferring in situ genetic adaptation to heat and drought stresses, usually from ex situ geo-referenced widespread collections of crop wild relatives, has become a prerequisite to capture these naturally available exotic variants. Unfortunately, after a century of collecting and preserving diversity in plants throughout germplasm banks, living seed collections that serve as repositories of genetic and ecological variation [36], ex situ collections do not necessarily fully span the ecological niche of agricultural crop relatives [37,38] as to establish new sources of genes for improving complex adaptive traits. Nowadays, wild relatives are still discovered [39], and further expeditions (Figure 1d) are needed (e.g., in the ecological hyper-variable and species-rich neotropics) [40,41]. Because isolated pockets of cryptic diversity still persist, novel habitat-based population-guided collections for genebanks are paramount, now more than ever.
2.2. Going Global by Adapting Local
Germplasm collections should be used not only to introduce exotic variation but also to avoid genetic erosion and increase long-term adaptability to climate change by making crops more resistant to abiotic stresses such as heat and drought. Local adaptation to abiotic factors can be studied using historical climate at the habitats where geo-referenced germplasm accessions were originally collected (Figure 1b). If an ecological balance [42] between genotypes and environments can be assumed [43,44] (Humboldt’s ‘harmony in nature’), geo-referencing and repositories of in situ climate variables can then be used to compute adaptive capacity. The former condition typically applies for landraces and crop wild relatives because they have occupied local niches long enough as to be shaped by natural selection forces [45]. This approach has typically relied on clustering algorithms to predict (Table 1) not only drought tolerance, water use efficiency [46,47], and thermal tolerance [48], but also resistance to pathogens [49], and aluminum toxicity [50] in a wide range of landraces and wild species.
Genomic analytical tools (Figure 1e) commonly coupled with environmental variables in order to reconstruct the genetic architecture (Figure 1f) of adaptive trait variation to abiotic stresses are genome-wide selection scans (GWSS) [51] and genome–environment associations (GEA) [52]. Both approaches aim to capture the signatures of selection to different environments by retrieving those genomic regions that segregate and are fixed among contrasting habitats (e.g., arid vs. wet regions) [53,54]. The strategies differ in that the former uses outlier tests given a background (‘baseline’) distribution, usually within a Bayesian framework [55], while the latter relies on mixed linear models (MLMs) that explicitly incorporate covariates as random effects [56]. Because these analyses may be misleading [57,58], if confusing factors [33,59] are not appropriately accounted for, MLMs are nowadays the preferred method to describe the genetic basis of local adaptation in germplasm collections [60]. Other major improvements are the use of indices, rather than raw environmental variables, that summarize precise physiological processes (e.g., thermal thresholds and potential evapotranspiration models to infer heat/drought stress) [10,18,19,20], and the collection of spatial high-resolution climate data to make accurate predictions at the regional [61] and micro-habitat [62,63] levels.
3. On the Need to Speed-Up
By studying local adaptation in crop-related wild collections and landraces using available phenotypic data (Figure 1a,e) and geo-referencing (Figure 1b), and uncovering the genes that underlie complex adaptive traits (such as root morphotypes, phenology and fruit/seed set, Figure 1f), natural variation has the potential to meet the global food security demands of a drier world [4]. However, unlocking adaptive genetic variation hidden in early landraces and related wild species (Figure 1c) remains a major challenge. The rate of genetic gain for abiotic stress tolerance in elite lines via traditional backcrossing schemes [64,65,66] may be limited due to high phylogenetic distance [67,68] and low hybridization levels. To overcome the bottleneck of pyramiding allelic variants from the wild into elite lines (Figure 1h), marker-assisted backcrossing (MAB) [69,70,71,72,73], predictive breeding [74], and gene editing [75] are promising alternatives.
3.1. Going Quicker by Selecting (Poly) Genomically
MAB recurrent selection strategies (Figure 1i) have proven to be useful for the introgression, from exotic germplasm, of simple Mendelian traits, which are those regulated by few genes such as resistance to biotic threats (e.g., [73]). However, by definition, MAB may be unable to trace quantitative traits such as heat and drought tolerance that are typically polygenic—regulated by many low-effect genes [76]. A last-generation analytical innovation that arose by merging quantitative genetics theory with genomics is predictive breeding, more commonly known as genomic prediction (GP) [74,77,78].

{kind=link}
{kind=link}
Table 1.
Non-parametric and parametric classification approaches that can assist clustering efforts to differentiate between abiotic stress tolerant and susceptible germplasm accessions. Habitat types and local adaptation to heat and drought stresses can be inferred using climate variables and physiological indices from each accession’s geo-referencing (Figure 1b) because crop wild relatives and landraces have occupied local niches (e.g., arid vs. wet regions) long enough as to be shaped by natural selection. Predicted thermal tolerance and water use efficiency, together with other data types (Figure 1e,f), can then be merged (Figure 1g) in order to identify and unlock novel sources of heat and drought tolerance. The trained classification may also speed-up the utilization of these tolerant variants by genomic-assisted breeding techniques (Figure 1i). ML approaches (Table 2) are also capable of including further data types for more cohesive multi-dimensional predictions (e.g., Figure 1m).
Table 1.
Non-parametric and parametric classification approaches that can assist clustering efforts to differentiate between abiotic stress tolerant and susceptible germplasm accessions. Habitat types and local adaptation to heat and drought stresses can be inferred using climate variables and physiological indices from each accession’s geo-referencing (Figure 1b) because crop wild relatives and landraces have occupied local niches (e.g., arid vs. wet regions) long enough as to be shaped by natural selection. Predicted thermal tolerance and water use efficiency, together with other data types (Figure 1e,f), can then be merged (Figure 1g) in order to identify and unlock novel sources of heat and drought tolerance. The trained classification may also speed-up the utilization of these tolerant variants by genomic-assisted breeding techniques (Figure 1i). ML approaches (Table 2) are also capable of including further data types for more cohesive multi-dimensional predictions (e.g., Figure 1m).
| Approach | Method | Description of the Method | R Package/Tool | Method’s Reference | Example (Accessions × Markers) |
|---|---|---|---|---|---|
| Non-Parametric (Reduction-Based) | K-means | Each observation belongs to the cluster with the nearest mean. It minimizes the distance between points labeled to be in a cluster and a point designated as the center (mean) | Stats & optCluster | [79,80] | Maize—2022 × 65,995 [81] |
| Ryegrass—1757 × 1,005,590 [82] | |||||
| Partitioning Around Medoids (PAM) | It minimizes the distance between points labeled to be in a cluster and a point designated as the center (medoid) of that cluster. PAM chooses data points as centers (medoid) and works with a generalization of the Manhattan Norm to define data points distance | cluster & optCluster | [83] | Maize—260 × 11,296,689 [84] | |
| Clustering large applications (CLARA) | It extracts multiple sample sets from the dataset and uses the best cluster as output. It uses PAM for each sample | cluster & optCluster | [83] | 90 × 5000 [85] | |
| Non-Parametric (Distance-Based) | Hierarchical clustering (Hclust) | It is a method of cluster analysis that seeks to build a hierarchy of clusters | hclust & optCluster | [86] | Barley—1816 × 1416 & Wheat—478 × 219 [87] |
| Oat—131 × 3567 [88] | |||||
| DIANA (Divisive analysis) | It first places all objects in a cluster and then subdivides them into smaller clusters until the desired number of clusters is obtained | cluster & optCluster | [83] | These algorithms were systematically compared, and included K-means, PAM, CLARA, Hclust, DIANA, and AGNES [89] | |
| Agglomerative Nesting (AGNES) | It initially takes each object as a cluster, afterwards the clusters are merged step by step according to certain criteria, using a single-link method | cluster & optCluster | [83,90] | ||
| AWclust | The first step of AWclust is to construct the ASD matrix between all pairs of individuals in the sample. The second step is to apply hierarchical clustering to infer clusters of individuals from the ASD matrix using Ward’s minimum variance algorithm | AWclust | [91] | Olive—94 × 8088 [92] | |
| Pepper—222 × 32,950 [93] | |||||
| Parametric | TESS3 | Geography is one of the most important determinants of genetic variation in natural populations. Using genotypic and geographic data, tess3r provides estimates of landscape population genetic structure | tess3r | [94] | These algorithms have been widely used and compared among them [95], and with other non-parametric methods [96] |
| fast STRUCTURE | STRUCTURE uses the core Bayesian principle of comparing likelihoods. Prior information about study samples can be supplied to further shape the unsupervised clustering | fast STRUCTURE | [97] |
GP uses historical phenotypic data [98] to calibrate marker-based infinitesimal additive predictive models [99,100,101], which provide a more comprehensive representation of a quantitative polygenic trait than traditional genetic mapping [102]. Hence, GP offers a key path to assist the introgression breeding of abiotic stress tolerance from the wild. GP’s predictive ability can be significantly enhanced after performing a priori weighted trait mapping through more conventional methods such as quantitative-trait loci (QTL) mapping [103], or genome-wide association studies (GWAS) [104], even across SNP-genotyped multi-parental populations [105,106,107,108,109]. GP may also go beyond breeding efforts, and feed back on optimizing restoration [102] and genebank characterization [110] (e.g., by predicting seed morphometric traits [111] and yield [112,113]) of thousands of germplasm accessions that could hardly be tested in field trials.
3.2. Fast Editing
MAB is likely to mutate into genomic-assisted parental selection (GAPS) [114] and Backcrossing (GABC) [115] as the preferred strategies for the introgression breeding (Figure 1i) of abiotic stress tolerant genetic alleles from crop wild relatives that have not undergone the domestication syndrome. Even though these scopes tend to underrate GWAS, fine-mapping is still needed for future usage of more target gene editing strategies (Figure 1j) such as CRISPR-Cas9 [75], for which a precise location of (few) major target genes is mandatory. This technique allows editing or disrupting target genes and regulator elements with a proven effect on the phenotypes. It originates from a natural system that provides bacteria with adaptive immunity to virus [75] that is co-opted to shuffle genetic variants already present in nature. Because of this, CRISPR-Cas9 does not strictly lead to genetically modified organisms (GMOs) s.s. As a proof of concept, the feasibility of this technology for drought tolerance has been validated in model plant species such as Arabidopsis [116], and crops with consolidated tissue culture protocols like tomato [117,118]. It has even demonstrated its potential for multi-trait quantitative editing [119] in orphan crops like groundcherry (Physalis pruinosa) [120]. However, its applicability in species with recalcitrant tissue culture is still limited, and most plants with enhanced tolerance phenotypes have displayed a trade-off with reduced crop yield [12]. A promising approach to uncouple abiotic stress tolerance from plant growth is to edit tissue-specific pathways for target drought traits [121] that could be improved to obtain heat and drought tolerant varieties more efficiently [122].
Furthermore, predictive breeding and gene editing should not be conceived as separate, non-overlapping approaches. Integration of molecular genetics and genomic analyses with more quantitative approaches has the potential to speed up various other efforts in addition to understand how plants are naturally adapted to drier climates, aiding in the innovation of biotechnological strategies and breeding programs to generate elite lines that are more productive and more capable of adapting to anticipated heat waves and drought events [123]. This is particularly true for complex traits, because even though CRISPR-Cas9 can quickly transfer major elements responsible for the abiotic stress tolerance response, its omni/polygenic nature demands assistance from other approaches like GP. Merging molecular mechanistic details that are often overlooked by genome-based assisted selection, with GP, breeding simulations [99,124], and speed breeding [125,126], would confer a major opportunity for robust whole-genome forecasting of complex and polygenic drought/heat tolerance phenotypes [124].
4. A Way Forward via Machine Learning
Despite the undertaken GWSS and GEA approaches having demonstrated their utility in identifying naturally available genetic adaptation to heat and drought (Figure 1c), these predictions wait to be contrasted in more systematic ways with GWAS results, GP estimates (Figure 1i), and field trials (Figure 1k), a target task for a trans-disciplinary multi-dimensional synthetic approach such as machine learning (ML).
As GP, ML output focuses on optimizing the predictive ability, or ‘recall’ rate, of a given classification (e.g., tolerant vs. susceptible accessions) (Figure 2 and Table 2), usually relying on a ‘supervised’ training with ‘labeled’ data (e.g., accessions for which drought/heat [127] tolerance are already measured or environmentally-estimated, Table 2). An advantage of ML is that it deals more efficiently with high-dimensional inputs of heterogeneous variables (Figure 1g) (‘features’) without a priori knowledge of their joint probability distribution [128], bypassing the ‘curse of dimensionality’ and a complex systems thinking (Figure 1l,m).
ML has long ago been used for ecological niche modeling [129,130] and functional genomics [131]. However, ML has started permeating, until very recently, other approaches more relevant to this review such as GWSS [128,132] and GP [133,134,135]. In this latter example, ML techniques (i.e., deep learning) outperformed GP’s predictive ability for single traits in multi-environment trials (Figure 1k).
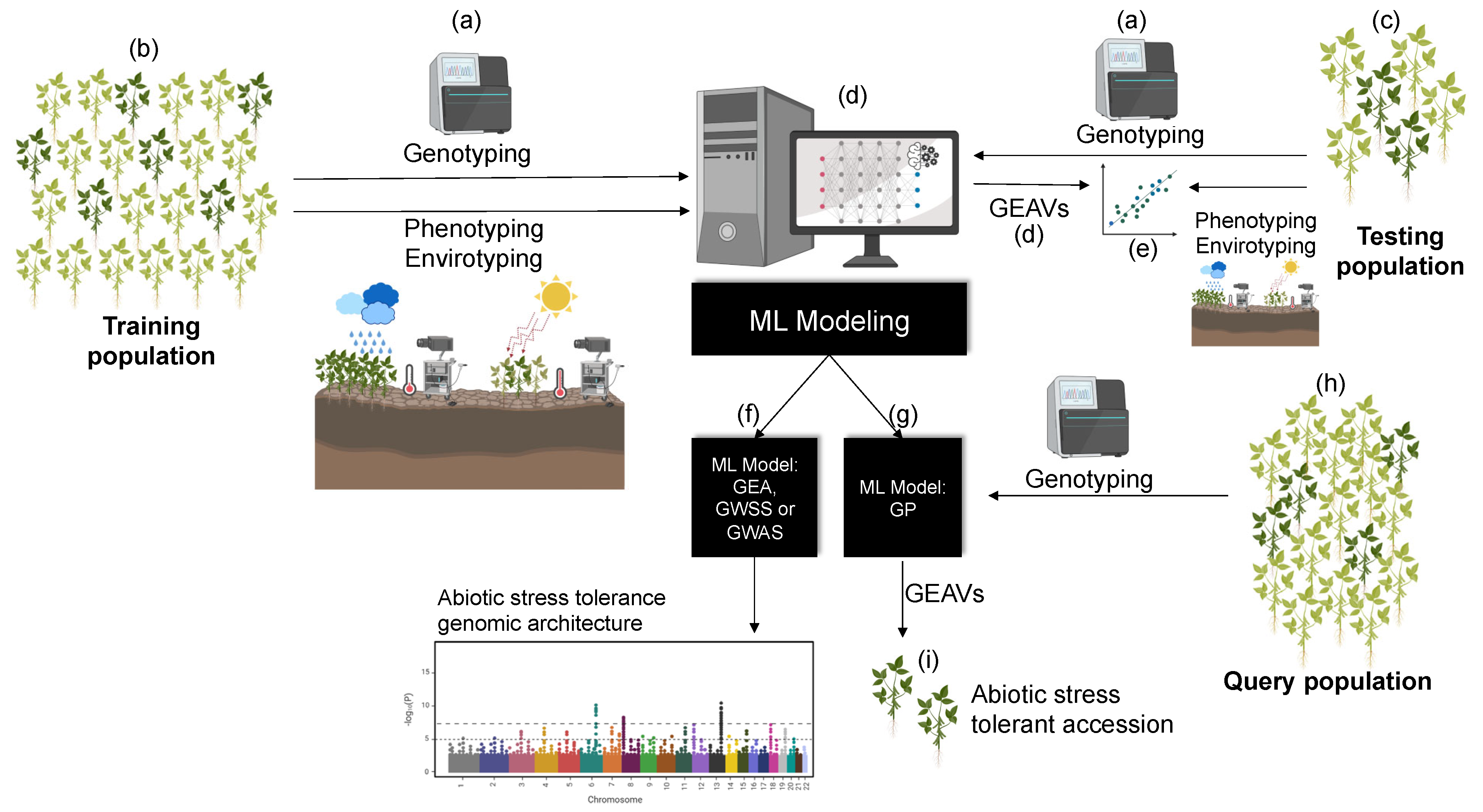
Figure 2.
A pipeline for machine learning (ML) applications capable of predicting abiotic stress tolerant and susceptible germplasm accessions. First, a subset of the germplasm collection is (a) characterizing genomically, phenotypically (whenever possible), and environmentally (i.e., abiotic stress adaptation indices based on geo-referencing). This subset is later on partitioned between (b) training and (c) testing populations. The training population is used to calibrate (d) ML models that aim using genomic information to predict genomic estimated adaptive values (GEAVs, an analogous rank to the polygenic risk score (PGS) and genomic estimated breeding value (GEBV) from the quantitative genomics literature, e.g., [102,136]). The computer screen depicts a hypothetical hidden neural network (HNN) algorithm, which is one among many potential ML tools; the repertoire includes several regressions, classification, and deep learning models, thoughtfully reviewed this year by Sebestyén et al. [137] and Tong and Nikoloski [138]. Meanwhile, the testing population is used to compute the (e) unbiased predictive ability of the model by comparing the GEAVs with the recorded environmental (or phenotypic) abiotic stress tolerant/susceptible indices. Broadly speaking, calibrated and validated ML models can serve two main purposes when applied on germplasm collections. First, (f) they could enhance our knowledge on the genomic architecture (i.e., genetic basis) of abiotic stress tolerance via ML-based genome-wide association studies (GWAS), and on the genomic landscape of adaptation via ML-based genome-wide selection scans (GWSS) and genome–environment associations (GEA). Second, (g) calibrated and validated ML models can be applied on a (h) query population such as extended germplasm samples for which environmental-based indices or phenotyping are not viable, informing GEAVs and (i) abiotic stress tolerance on a wider genepool. Clusters of abiotic stress tolerance and susceptibility based on phenotypic information and/or environmental-based indices can be built using traditional classification tools such as the ones listed in Table 1, or may also leverage ML prediction approaches (Table 2).
Figure 2.
A pipeline for machine learning (ML) applications capable of predicting abiotic stress tolerant and susceptible germplasm accessions. First, a subset of the germplasm collection is (a) characterizing genomically, phenotypically (whenever possible), and environmentally (i.e., abiotic stress adaptation indices based on geo-referencing). This subset is later on partitioned between (b) training and (c) testing populations. The training population is used to calibrate (d) ML models that aim using genomic information to predict genomic estimated adaptive values (GEAVs, an analogous rank to the polygenic risk score (PGS) and genomic estimated breeding value (GEBV) from the quantitative genomics literature, e.g., [102,136]). The computer screen depicts a hypothetical hidden neural network (HNN) algorithm, which is one among many potential ML tools; the repertoire includes several regressions, classification, and deep learning models, thoughtfully reviewed this year by Sebestyén et al. [137] and Tong and Nikoloski [138]. Meanwhile, the testing population is used to compute the (e) unbiased predictive ability of the model by comparing the GEAVs with the recorded environmental (or phenotypic) abiotic stress tolerant/susceptible indices. Broadly speaking, calibrated and validated ML models can serve two main purposes when applied on germplasm collections. First, (f) they could enhance our knowledge on the genomic architecture (i.e., genetic basis) of abiotic stress tolerance via ML-based genome-wide association studies (GWAS), and on the genomic landscape of adaptation via ML-based genome-wide selection scans (GWSS) and genome–environment associations (GEA). Second, (g) calibrated and validated ML models can be applied on a (h) query population such as extended germplasm samples for which environmental-based indices or phenotyping are not viable, informing GEAVs and (i) abiotic stress tolerance on a wider genepool. Clusters of abiotic stress tolerance and susceptibility based on phenotypic information and/or environmental-based indices can be built using traditional classification tools such as the ones listed in Table 1, or may also leverage ML prediction approaches (Table 2).

Table 2.
Machine learning (ML) predictive tools validated within a GP framework that can be extended to assist clustering efforts to differentiate between abiotic stress tolerant and susceptible germplasm accessions. These ML algorithms could be trained to distinguish habitat types and local adaptation to heat and drought stresses by looking into in situ climate variables and physiological indices from each accession’s geo-referencing (Figure 1b). This is possible because crop wild relatives and landraces have occupied local niches (e.g., hot vs. cold regions) for enough time to be shaped by natural selection. Predicted heat and drought tolerance can further harness other data types (Figure 1e,f) in order to identify and unlock novel sources of heat and drought tolerance (Figure 1g). The ML trained classification may also speed-up the utilization of tolerant variants by genomic-assisted breeding techniques (Figure 1i and Figure 2g). Table is sorted by species name and by ML approach.
Table 2.
Machine learning (ML) predictive tools validated within a GP framework that can be extended to assist clustering efforts to differentiate between abiotic stress tolerant and susceptible germplasm accessions. These ML algorithms could be trained to distinguish habitat types and local adaptation to heat and drought stresses by looking into in situ climate variables and physiological indices from each accession’s geo-referencing (Figure 1b). This is possible because crop wild relatives and landraces have occupied local niches (e.g., hot vs. cold regions) for enough time to be shaped by natural selection. Predicted heat and drought tolerance can further harness other data types (Figure 1e,f) in order to identify and unlock novel sources of heat and drought tolerance (Figure 1g). The ML trained classification may also speed-up the utilization of tolerant variants by genomic-assisted breeding techniques (Figure 1i and Figure 2g). Table is sorted by species name and by ML approach.
| ML Approach(es) | Species | Accessions x Genetic Markers | Reference |
|---|---|---|---|
| RF | Barley | 911 × 2146 SNP | Heslot et al., 2012 [139] |
| ANN | Bean | 80 × 384 SNP | Rosado et al., 2020 [140] Ψ |
| SVM | Black tea | 255 × 1421 DArT SNP | Koech et al., 2020 [141] Ψ |
| RF | Chickpea | 315 × 1568 DArT SNP | Roorkiwal et al., 2016 [142] Ψ |
| DT, Bagging, Boosting, RF, ANN | Coffee | 245 × 74 AFLP, 58 SSR, 4 RAPD, and 2 primers | Sousa et al., 2021 [143] |
| RF | Coffee | 96 × 38,106 SNP | Ferrão et al., 2019 [144] |
| SVM | Hybrid Rice | 575 × 116,482 SNP | Xu et al., 2018 [145] |
| ANN | Maize | 300 × 55,000 SNP | González-Camacho, et al., 2012 [146] |
| DL | Maize | 148,452 × 19,465 SNP | Khaki & Wang, 2019 [147] |
| DL | Maize | ~300 × ~1000 SNP | Rachmatia et al., 2017 [148] |
| KNN | Maize | 198 × 75 SSR | Maenhout et al., 2007 [149] |
| MLP, PNN | Maize | ~300 × 46,374 SNP | González-Camacho et al., 2016 [150] Ψ |
| RBFNN, ANN | Maize | ~300 × 46,374 SNP | González-Camacho et al., 2012 [146] Ψ |
| RF | Maize | 240 × 29,619 SNP | Shikha et al., 2017 [151] Ψ |
| RF | Maize | 240 × 56,110 SNP | Shikha et al., 2017 [151] |
| RF, SVM, ANN, Boosting | Maize | 391 × 332,178 SNP | Azodi et al., 2019 [152] |
| SVM | Maize | 4,328 × 564,692 SNP | Zhao et al., 2020 [153] |
| SVM, RF | Maize | 113 × 47,458 SNP | Li et al., 2020 [154] Ψ |
| ZAP-RF | Maize | 115 × 1635 SNP | Montesinos-López et al., 2021 [155] |
| DL | Maize | 309 × 158,281 SNP | Montesinos-López, et al., 2018 [135] |
| RF, SVM | Mice | 1,884 × 9917 SNP | Neves et al., 2012 [156] |
| SVM | Pea | 105 × 7521 SNP | Annicchiarico et al., 2017 [157] |
| RF, Boosting, KNN | Perennial ryegrass | 86 × 1670 SNP | Grinberg et al., 2016 [158] |
| RF, GBM, KNN | Perennial ryegrass | 86 × 1670 SNP | Grinberg et al., 2016 [158] |
| Bagging, RF, SVM | Rice | 363 × 73,147 SNP | Banerjee et al., 2020 [159] Ψ |
| RF | Rice | 110 × 3071 SNP | Onogi et al., 2015 [160] |
| SVM, Boosting | Simulated Dataset | 3226 × 10,031 | Ogutu et al., 2011 [161] |
| DL | Strawberry | 1358 × 9908 SNP | Zingaretti et al., 2020 [162] |
| ANN | Wheat | 599 × 1279 SNP | Gianola, et al., 2011 [163] |
| ANN | Wheat | 306 × 1717 SNP | Pérez-Rodríguez et al., 2012 [164] |
| DL | Wheat | ~500 × 15,744 SNP | Crossa et al., 2019 [133] |
| DL | Wheat | 237 × 27,957 SNP | Guo et al., 2020 [165] Ψ |
| DL | Wheat | 2000 × 33,709 DArT SNP | Ma et al., 2017 [166] |
| GBM, RF, SVM | Wheat | 254 × 33,516 SNP | Grinberg et al., 2020 [167] Ψ |
| MLP, PNN | Wheat | ~300 × 1717 DArT SNP | González-Camacho et al., 2016 [150] Ψ |
| RF | Wheat | 254 × 41,371 SNP | Poland et al., 2012 [168] |
| RF, KNN | Wheat | 273 × 5054 SNP | Arruda et al., 2015 [169] |
| DL, SVM | Wheat | 3486 × 2038 SNP | Montesinos-López et al., 2019 [170] |
ML tool abbreviations as follows: adaptive boosting (AdaBoost), artificial neural networks (ANN), decision tree (DT), deep learning (DL), extreme gradient boosting (XGBoost), gradient boosting machines (GBM), multilayer perceptron neural network (MLP), probabilistic neural network (PNN), radial basis function neural network (RBFNN), random forest (RF), support vector machines (SVM), and zero altered Poisson random forest (ZAP-RF). ML-coupled genomic prediction initiatives explicitly related to abiotic stress tolerance are marked with Ψ under reference.
Nowadays, it is anticipated that ML will overtake current GP models in the near future by incorporating explicit environmental variables and habitat adaptation indices (Figure 1b) as well as estimates of the genome-wide nucleotide diversity (π, expected heterozygosity—He, and runs of homozygosity—ROH). Combining trans-disciplinary datasets from different origins and disciplines will ultimately improve the prediction of the phenotypic variance and the genotype × environment interaction, while sidestepping the restrictions of traditional breeding schemes to increase genetic gain for abiotic stress tolerance per unit of time.
Besides the methodological innovations of ML in terms of data merging and optimized classifications, extensions of this type of research are likely to assist hybrid breeding and polyploid utilization [162], both of which take advantage of dominance and over-dominance effects already present in nature, capable of increasing adaptability to abiotic stresses [31,171]. An antecedent in this direction is that GP can predict untested hybrid genotypes [172], even for perennial crops [125,173,174], by genotyping the potential parental lines and phenotyping a few F1 hybrids. Approximate-Bayesian Computation (ABC) analyses within ML [128,175] may in turn speed up polyploid selection.
In short, ML [138,176,177,178,179] and deep learning approaches [133,170,180,181] promise assisting the conservation [102,182,183,184], managing [185,186], prioritization [187,188,189], and introgression [190,191] of crop wild variation from genebanks (Figure 2). ML may be particularly useful in unexplored isolated pockets of diversity, which contain allelic variants otherwise eroded from modern genotypes [192,193]. These predictive tools may additionally accommodate multi-trait [194] multi-environment [195,196] scores within the growing fields of enviromics [197,198,199], green house speed phenotyping [126,200], simulation-based breeding [99,124], hyperspectral imaging (HSI) [201,202,203], and epigenetics [204], allowing for better resolution and pace to detect and use standing adaptation.
Extending these ultimate analytical approaches will help to resolve major basic questions in the field [205] such as to which scale diversifying selection promotes [206] and gene flow precludes [207,208] local adaptation, and whether large haplotype blocks are pervasive in adaptation signatures due to structural variants [209] such as chromosomal inversions [210,211]. Targeting these issues will allow trans-disciplinary research teams to move into more applied matters like (1) how GP and ML can be intermingled to turbocharge genebanks [212] while disentangling the correlated trade-off complexity in proxy traits as part of speed breeding strategies [125,126], and (2) what is the optimum pathway for pan-genomes [213] and epigenetics [214] to unlock the properties of plants for sustainable development [215]. Approaching these enquiries would enable genomics to cut threats of future global food security from climate change by harnessing crop wild diversity and genebank utilization. After all, a key promise of agriculture genomics [216] is to potentiate genebanks through impact-oriented pre-breeding [217] in the face of the current alimentary and climate emergency [218].
5. Conclusions
Mining the biodiversity in genebanks [4] and the cryptic diversity in nature [219] is a prerequisite to overcome food shortages due to increasing temperatures and drought. While climate data and genomics can speed up the identification (Figure 1c) of novel sources of tolerance, merging these data types more cohesively into cutting edge analytical frameworks like GP and ML will aid unlocking (Figure 1h) these new variants, so that they can realistically be transferred into the cultivated genepools, and ultimately, the farmers’ fields. In order to start moving toward this goal, genebank research should focus on (1) studying the genetic and ecological mechanisms of adaptation and diversification in crop species, while (2) investigating the genetic potential of wild relatives to cope with abiotic stresses, both by examining historical and novel germplasm materials. The research community must pay close attention to the discovery of locally adapted varieties and landraces (Figure 1d).
Genomic signatures of adaptation to dry and semi-arid habitats (Figure 1b) are useful for germplasm characterization, enhancing the identification of novel sources of variation, future genome-assisted selection, and crop improvement for abiotic stress tolerance. Ultimately, these analytical approaches, coupled with ML, promise improving the accuracy of parent selection to be used in pre-breeding and breeding steps, besides reinforcing the relevance of crops and their wild relatives as systems to address classic questions in the fields of quantitative genetics, evolutionary biology, and ecological genetics.
Looking forward, the discipline of quantitative genetics is merging with the field of ‘big data’ analytics, a ground for ML (Figure 1). An expanded view of complex traits is arising, moving from a polygenic framework to a view in which all genes are liable to affect adaptation to abiotic stresses [220], so that most heritability can be explained by the effects of rare variants [221,222] and their interactions with epigenetic factors [223]. However, looking back toward the bases of crop evolution [23,24] still has the potential to improve agriculture because natural selection has tested more options than humans ever will [35]. We envision that mining the molecular footprint of selection and adaptation in wild crop relatives and early landraces for plant breeding [224] and climate adaptation [125,225] will benefit from bridging the gap between phenotyping (Figure 1a) and genotyping (Figure 1e) in genebanks [226,227], and the more classical and deterministic conceptual models from the fields of quantitative and population genetics.
6. Perspectives
In terms of required innovations, it would be desirable to develop high-resolution fitness maps for various types of stresses [228], and to standardize genome-wide polygenetic scores (GPS) [229] by minimizing genomic pseudo-correlation or linkage disequilibrium (LD) [230,231,232], in order to predict novel abiotic stress tolerant sources in concert with other resistances and nutritional quality for the poorest. It is also becoming more essential to assess second-order genomic-context dependent associations (epistatic effects) as a tool to point toward more complex infinitesimal polygenic systems [233] capable of accounting for the missing heritability [58,234,235] in adaptation, while enhancing innovative genomic-assisted selection strategies that do not exclusively rely on MAS and GP, but also on ML.
Meanwhile, ML would be obsolete to speed up the breeding of resilient crops if heterogeneous datasets are not easily accessible and their maintenance is not well funded in the long-term. Germplasm accessions [36,226], previous and ongoing prioritizations [37,38,188,236], climate-based habitat inferences on the adaptive [47] and yield [126,237] potentials, genomic screenings [238] and rankings [239], field trials in contrasting environments [9,240], and agricultural surveys for farmers (Figure 1l), should all be made public access in order to pivot ML [241,242] breeding technologies [243]. Open access resources [244] and data sharing [245,246] are also essential to target new population-based collections [38] and better characterize the isolated pockets of diversity that still persist in the hidden hills and valleys of mega diverse hotspots. Without platforms for biological discovery [110] and predictors of the future’s uncertainty [247], further pre-breeding would exclusively rely on a narrow diversity without sufficient pre-adaptation to abiotic stresses, hence jeopardizing crop adaptation. As a strategy for feeding the future [4], the identification, conservation, and utilization of novel sources of heat and drought tolerance among crop wild relatives, some of them already kept in existing genebanks [226], will enable diversifying selection in plant breeding by offsetting the domestication winnowing effect on natural genetic variation [247].
Author Contributions
A.J.C. conceived this work. F.L.-H. collected literature, and prepared tables and figures with feedback from A.J.C. A.J.C. wrote a first draft later edited by F.L.-H. All authors have read and agreed to the published version of the manuscript.
Funding
Grants 4.1-2016-00418 from Vetenskapsrådet (VR) and BS2017-0036 from Kungliga Vetenskapsakademien (KVA) are acknowledged for supporting A.J.C. during the conception of this work.
Institutional Review Board Statement
Ethical review and approval were waived because this study did not involve humans or animals subjects.
Informed Consent Statement
Not applicable because this study did not involve humans subjects.
Data Availability Statement
Data Availability concerning the literature included in this review can be found in the citations hereinafter.
Acknowledgments
M.W. Blair is acknowledged for perceptive discussions with A.J.C. and F.L.-H., concerning drought adaptation in plants, as part of the Fulbright U.S. Specialist Program during the summer of 2019 in Rionegro (Antioquia, Colombia). We thank the Department for Research Capacity Building from AGROSAVIA for subsidizing F.L.-H.’s internship during 2018. Special recognition is given to M.J. Torres-Urrego for assistance during the writing of this review. Two anonymous reviewers and the editor are thanked for their feedback. Finally, we deeply thank MDPI for waiving this review’s APC as part of the Special Issue “Evolutionary Genetics of Plant Crop-Wild Complexes: From Fundamental to Applied Research”.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tai, A.P.K.; Martin, M.V.; Heald, C.L. Threat to future global food security from climate change and ozone air pollution. Nat. Clim. Chang. 2014, 4, 817–821. [Google Scholar] [CrossRef] [Green Version]
- Davis, K.F.; Gephart, J.A.; Emery, K.A.; Leach, A.M.; Galloway, J.N.; D’Odorico, P. Meeting future food demand with current agricultural resources. Glob. Environ. Chang. 2016, 39, 125–132. [Google Scholar] [CrossRef]
- McCouch, S. Feeding the future. Nature 2013, 499, 23–24. [Google Scholar] [CrossRef] [PubMed]
- Blair, M.W.; Pantoja, W.; Carmenza Munoz, L. First use of microsatellite markers in a large collection of cultivated and wild accessions of tepary bean (Phaseolus acutifolius A. Gray). Appl. Genet. 2012, 125, 1137–1147. [Google Scholar] [CrossRef] [PubMed]
- Borelli, T.; Hunter, D.; Powell, B.; Ulian, T.; Mattana, E.; Termote, C.; Pawera, L.; Beltrame, D.; Penafiel, D.; Tan, A.; et al. Born to Eat Wild: An Integrated Conservation Approach to Secure Wild Food Plants for Food Security and Nutrition. Plants 2020, 9, 1299. [Google Scholar] [CrossRef]
- von Wettberg, E.; Davis, T.M.; Smýkal, P. Wild Plants as Source of New Crops. Front. Plant. Sci. 2020, 11. [Google Scholar] [CrossRef]
- Gulisano, A.; Alves, S.; Martins, J.N.; Trindade, L.M. Genetics and Breeding of Lupinus mutabilis: An Emerging Protein Crop. Front. Plant. Sci. 2019, 10, 1385. [Google Scholar] [CrossRef]
- Beebe, S.E.; Rao, I.M.; Blair, M.W.; Acosta-Gallegos, J.A. Phenotyping common beans for adaptation to drought. Front. Physiol. 2013, 4, 35. [Google Scholar] [CrossRef] [Green Version]
- Cortés, A.J.; Blair, M.W. Genotyping by Sequencing and Genome—Environment Associations in Wild Common Bean Predict Widespread Divergent Adaptation to Drought. Front. Plant. Sci. 2018, 9, 128. [Google Scholar] [CrossRef] [Green Version]
- Vavilov, N.I. The law of homologous series in variation. J. Genet. 1922, 12, 47–89. [Google Scholar] [CrossRef]
- Tack, J.; Barkley, A.; Nalley, L.L. Effect of warming temperatures on US wheat yields. Proc. Natl. Acad. Sci. USA 2015, 112, 6931–6936. [Google Scholar] [CrossRef] [Green Version]
- Beebe, S.; Rao, I.M.; Cajiao, C.; Grajales, M. Selection for Drought Resistance in Common Bean Also Improves Yield in Phosphorus Limited and Favorable Environments. Crop. Sci. 2008, 48, 582–592. [Google Scholar] [CrossRef]
- Buitrago-Bitar, M.A.; Cortés, A.J.; López-Hernández, F.; Londoño-Caicedo, J.M.; Muñoz-Florez, J.E.; Muñoz, L.C.; Blair, M.W. Allelic Diversity at Abiotic Stress Responsive Genes in Relationship to Ecological Drought Indices for Cultivated Tepary Bean, Phaseolus acutifolius A. Gray, and Its Wild Relatives. Genes 2021, 12, 556. [Google Scholar] [CrossRef] [PubMed]
- Bartels, D.; Sunkar, R. Drought and salt tolerance in plants. Crit. Rev. Plant Sci. 2005, 24, 23–58. [Google Scholar] [CrossRef]
- Grene, R.; Provart, N.J.; Pardo, J.M. Editorial: Resistance to Salinity and Water Scarcity in Higher Plants. Insights From Extremophiles and Stress-Adapted Plants: Tools, Discoveries and Future Prospects. Front. Plant Sci. 2019, 10, 373. [Google Scholar] [CrossRef]
- Bechtold, U. Plant Life in Extreme Environments: How Do You Improve Drought Tolerance? Front. Plant Sci. 2018, 9, 543. [Google Scholar] [CrossRef] [Green Version]
- Blair, M.W.; Cortés, A.J.; This, D. Identification of an ERECTA gene and its drought adaptation associations with wild and cultivated common bean. Plant. Sci. 2016, 242, 250–259. [Google Scholar] [CrossRef]
- Cortés, A.J.; Chavarro, M.C.; Madriñán, S.; This, D.; Blair, M.W. Molecular ecology and selection in the drought-related Asr gene polymorphisms in wild and cultivated common bean (Phaseolus vulgaris L.). BMC Genet. 2012, 13. [Google Scholar] [CrossRef] [Green Version]
- Cortés, A.J.; This, D.; Chavarro, C.; Madriñán, S.; Blair, M.W. Nucleotide diversity patterns at the drought-related DREB2 encoding genes in wild and cultivated common bean (Phaseolus vulgaris L.). Theor. Appl. Genet. 2012, 125, 1069–1085. [Google Scholar] [CrossRef]
- Asfaw, A.; Ambachew, D.; Shah, T.; Blair, M.W. Trait Associations in Diversity Panels of the Two Common Bean (Phaseolus vulgaris L.) Gene Pools Grown under Well-watered and Water-Stress Conditions. Front. Plant Sci. 2017, 8, 733. [Google Scholar] [CrossRef] [PubMed]
- Darwin, C. The Variation of Animals and Plants under Domestication; John Murray: London, UK, 1868. [Google Scholar]
- Purugganan, M.D.; Fuller, D.Q. The nature of selection during plant domestication. Nature 2009, 457, 843–848. [Google Scholar] [CrossRef] [PubMed]
- Meyer, R.S.; Purugganan, M.D. Evolution of crop species: Genetics of domestication and diversification. Nat. Rev. Genet. 2013, 14, 840–852. [Google Scholar] [CrossRef] [PubMed]
- Meyer, R.S.; DuVal, A.E.; Jensen, H.R. Patterns and processes in crop domestication: An historical review and quantitative analysis of 203 global food crops. New Phytol. 2012, 196, 29–48. [Google Scholar] [CrossRef]
- Khoury, C.K.; Achicanoy, H.A.; Bjorkman, A.D.; Navarro-Racines, C.; Guarino, L.; Flores-Palacios, X.; Engels, J.M.M.; Wiersema, J.H.; Dempewolf, H.; Sotelo, S.; et al. Origins of food crops connect countries worldwide. Proc. R. Soc. B 2016, 283, 1832. [Google Scholar] [CrossRef]
- Schmutz, J.; McClean, P.E.; Mamidi, S.; Wu, G.A.; Cannon, S.B.; Grimwood, J.; Jenkins, J.; Shu, S.; Song, Q.; Chavarro, C.; et al. A reference genome for common bean and genome-wide analysis of dual domestications. Nat. Genet. 2014, 46, 707–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cortés, A.J.; Skeen, P.; Blair, M.W.; Chacón-Sánchez, M.I. Does the genomic landscape of species divergence in Phaseolus beans coerce parallel signatures of adaptation and domestication? Front. Plant Sci. 2018, 9, 1816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stern, D.L.; Orgogozo, V. Is Genetic Evolution Predictable? Science 2009, 323, 746–751. [Google Scholar] [CrossRef] [Green Version]
- Mather, K.A.; Molina, J.; Flowers, J.M.; Rubinstein, S.; Rauh, B.L.; Lawton-Rauh, A.M.Y.; Caicedo, A.L.; McNally, K.L.; Purugganan, M.D. Migration, isolation and hybridization in island crop populations: The case of Madagascar rice. Mol. Ecol. 2010, 19, 4892–4905. [Google Scholar] [CrossRef]
- Marques, D.A.; Meier, J.I.; Seehausen, O. A Combinatorial View on Speciation and Adaptive Radiation. Trends Ecol. Evol. 2019, 34, 531–544. [Google Scholar] [CrossRef] [Green Version]
- Seehausen, O.; Butlin, R.K.; Keller, I.; Wagner, C.E.; Boughman, J.W.; Hohenlohe, P.A.; Peichel, C.L.; Saetre, G.-P.; Bank, C.; Brännström, Å.; et al. Genomics and the origin of species. Nat. Rev. Genet. 2014, 15, 176–192. [Google Scholar] [CrossRef] [Green Version]
- Wolf, J.B.; Ellegren, H. Making sense of genomic islands of differentiation in light of speciation. Nat. Rev. Genet. 2017, 18, 87–100. [Google Scholar] [CrossRef] [PubMed]
- Nicotra, A.B.; Atkin, O.K.; Bonser, S.P.; Davidson, A.M.; Finnegan, E.J.; Mathesius, U.; Poot, P.; Purugganan, M.D.; Richards, C.L.; Valladares, F.; et al. Plant phenotypic plasticity in a changing climate. Trends Plant. Sci. 2010, 15, 684–692. [Google Scholar] [CrossRef] [PubMed]
- Denison, R.F. Darwinian Agriculture: How Understanding Evolution Can Improve Agriculture; Princeton University Press: Princeton, NJ, USA, 2016. [Google Scholar]
- Tanksley, S.D.; McCouch, S.R. Seed Banks and Molecular Maps: Unlocking Genetic Potential from the Wild. Science 1997, 227, 1063–1066. [Google Scholar] [CrossRef] [Green Version]
- Ramirez-Villegas, J.; Khoury, C.K.; Achicanoy, H.A.; Mendez, A.C.; Diaz, M.V.; Sosa, C.C.; Debouck, D.G.; Kehel, Z.; Guarino, L. A gap analysis modelling framework to prioritize collecting for ex situ conservation of crop landraces. Divers. Distrib. 2020, 26, 730–742. [Google Scholar] [CrossRef]
- Castaneda-Alvarez, N.P.; Khoury, C.K.; Achicanoy, H.A.; Bernau, V.; Dempewolf, H.; Eastwood, R.J.; Guarino, L.; Harker, R.H.; Jarvis, A.; Maxted, N.; et al. Global conservation priorities for crop wild relatives. Nat. Plants 2016, 2, 16022. [Google Scholar] [CrossRef]
- Salcedo-Castaño, J.; Araya-Villalobos, R.; Castañeda-Alvarez, N.; Toro-Chica, O.; Debouck, D.G. Phaseolus hygrophilus (Leguminosae-Papilionoideae), a new wild bean species from the wet forests of Costa Rica, with notes about section Brevilegumeni. J. Bot. Res. Inst. Tex. 2011, 5, 53–65. [Google Scholar]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Fonseca, G.A.B.d.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Pimm, S.L.; Jenkins, C.N.; Abell, R.; Brooks, T.M.; Gittleman, J.L.; Joppa, L.N.; Raven, P.H.; Roberts, C.M.; Sexton, J.O. The biodiversity of species and their rates of extinction, distribution, and protection. Science 2014, 344, 6187. [Google Scholar] [CrossRef] [PubMed]
- Berg, J. Gene-environment interplay. Science 2016, 354, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franks, S.J.; Hoffmann, A.A. Genetics of Climate Change Adaptation. Annu. Rev. Genet. 2012, 46, 185–208. [Google Scholar] [CrossRef]
- Stapley, J.; Reger, J.; Feulner, P.G.D.; Smadja, C.; Galindo, J.; Ekblom, R.; Bennison, C.; Ball, A.D.; Beckerman, A.P.; Slate, J. Adaptation genomics: The next generation. Trends Ecol. Evol. 2011, 25, 705–712. [Google Scholar] [CrossRef] [PubMed]
- Hancock, A.M.; Brachi, B.; Faure, N.; Horton, M.W.; Jarymowycz, L.B.; Sperone, F.G.; Toomajian, C.; Roux, F.; Bergelson, J. Adaptation to Climate Across the Arabidopsis thaliana Genome. Science 2011, 334, 83–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lasky, J.R.; Des Marais, D.L.; McKay, J.K.; Richards, J.H.; Juenger, T.E.; Keitt, T.H. Characterizing genomic variation of Arabidopsis thaliana: The roles of geography and climate. Mol. Ecol. 2012, 21, 5512–5529. [Google Scholar] [CrossRef] [PubMed]
- Cortés, A.J.; Monserrate, F.; Ramírez-Villegas, J.; Madriñán, S.; Blair, M.W. Drought Tolerance in Wild Plant Populations: The Case of Common Beans (Phaseolus vulgaris L.). PLoS ONE 2013, 8, e62898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Hernández, F.; Cortés, A.J. Last-Generation Genome–Environment Associations Reveal the Genetic Basis of Heat Tolerance in Common Bean (Phaseolus vulgaris L.). Front. Genet. 2019, 10, 22. [Google Scholar] [CrossRef] [Green Version]
- Yoder, J.B.; Stanton-Geddes, J.; Zhou, P.; Briskine, R.; Young, N.D.; Tiffin, P. Genomic signature of adaptation to climate in Medicago truncatula. Genetics 2014, 196, 1263–1275. [Google Scholar] [CrossRef] [Green Version]
- Lasky, J.R.; Upadhyaya, H.D.; Ramu, P.; Deshpande, S.; Hash, C.T.; Bonnette, J.; Juenger, T.E.; Hyma, K.; Acharya, C.; Mitchell, S.E.; et al. Genome-environment associations in sorghum landraces predict adaptive traits. Sci. Adv. 2015, 1, e1400218. [Google Scholar] [CrossRef] [Green Version]
- Sabeti, P.C.; Varilly, P.; Fry, B.; Lohmueller, J.; Hostetter, E.; Cotsapas, C.; Xie, X.; Byrne, E.H.; McCarroll, S.A.; Gaudet, R.; et al. Genome-wide detection and characterization of positive selection in human populations. Nature 2007, 449, 913–918. [Google Scholar] [CrossRef]
- Forester, B.R.; Jones, M.R.; Joost, S.; Landguth, E.L.; Lasky, J.R. Detecting spatial genetic signatures of local adaptation in heterogeneous landscapes. Mol. Ecol. 2016, 25, 104–120. [Google Scholar] [CrossRef]
- Rellstab, C.; Gugerli, F.; Eckert, A.J.; Hancock, A.M.; Holderegger, R. A practical guide to environmental association analysis in landscape genomics. Mol. Ecol. 2015, 24, 4348–4370. [Google Scholar] [CrossRef] [Green Version]
- Zahn, L.M.; Purnell, B.A. Genes under pressure. Science 2016, 354, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antao, T.; Lopes, A.; Lopes, R.J.; Beja-Pereira, A.; Luikart, G. LOSITAN: A workbench to detect molecular adaptation based on a Fst-outlier method. BMC Bioinform. 2008, 9, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kruglyak, L. The road to genome-wide association studies. Nat. Rev. Genet. 2008, 9, 314–318. [Google Scholar] [CrossRef] [PubMed]
- Pennisi, E. Disputed islands. Science 2014, 345, 611–613. [Google Scholar] [CrossRef] [PubMed]
- Maher, B. The case of the Missing Heritability. Nature 2008, 456, 18–21. [Google Scholar] [CrossRef]
- Lambert, C.G.; Black, L.J. Learning from our GWAS mistakes: From experimental design to scientific method. Biostatistics 2012, 13, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Abebe, T.D.; Naz, A.A.; Leon, J. Landscape genomics reveal signatures of local adaptation in barley (Hordeum vulgare L.). Front. Plant Sci. 2015, 6, 813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pluess, A.R.; Frank, A.; Heiri, C.; Lalague, H.; Vendramin, G.G.; Oddou-Muratorio, S. Genome-environment association study suggests local adaptation to climate at the regional scale in Fagus sylvatica. New Phytol. 2016, 210, 589–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frachon, L.; Bartoli, C.; Carrere, S.; Bouchez, O.; Chaubet, A.; Gautier, M.; Roby, D.; Roux, F. A Genomic Map of Climate Adaptation in Arabidopsis thaliana at a Micro-Geographic Scale. Front. Plant Sci. 2018, 9, 967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cortés, A.J.; Wheeler, J.A.; Sedlacek, J.; Lexer, C.; Karrenberg, S. Genome-wide patterns of microhabitat-driven divergence in the alpine dwarf shrub Salix herbacea L. In On The Big Challenges of a Small Shrub: Ecological Genetics of Salix herbacea L.; Acta Universitatis Upsaliensis: Uppsala, Sweden, 2015. [Google Scholar]
- Muñoz, L.C.; Blair, M.W.; Duque, M.C.; Tohme, J.; Roca, W. Introgression in common bean x tepary bean interspecific congruity-backcross lines as measured by AFLP markers. Crop. Sci. 2003, 44, 637–645. [Google Scholar] [CrossRef]
- Mejía-Jiménez, A.; Muñoz, C.; Jacobsen, H.J.; Roca, W.M.; Singh, S.P. Interspecific hybridization between commonand tepary beans: Increased hybrid embryo growth, fertility, andefficiency of hybridization through recurrent and congruity backcrossing. Theor. Appl. Genet. 1994, 88, 324–331. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.J.; Xu, J.L.; Ismail, A.M.; Fu, B.Y.; Vijaykumar, C.H.M.; Gao, Y.M.; Domingo, J.; Maghirang, R.; Yu, S.B.; Gregorio, G.; et al. Hidden diversity for abiotic and biotic stress tolerances in the primary gene pool of rice revealed by a large backcross breeding program. Field Crop. Res. 2006, 97, 66–76. [Google Scholar] [CrossRef]
- Smýkal, P.; Coyne, C.J.; Ambrose, M.J.; Maxted, N.; Schaefer, H.; Blair, M.W.; Berger, J.; Greene, S.L.; Nelson, M.N.; Besharat, N.; et al. Legume Crops Phylogeny and Genetic Diversity for Science and Breeding. Crit. Rev. Plant Sci. 2014, 34, 43–104. [Google Scholar] [CrossRef] [Green Version]
- Crespo, S.; Rivera-Peña, M.; Rosero-Alpala, D.A.; Muñoz-Florez, J.E.; Rao, I.M.; Muñoz-Florez, L.C. Pollen viability of Tepary bean (Phaseolus acutifolius A. Gray.) mutant lines under water stress conditions and inoculation with rhizobia. Acta Agronómica 2018, 67, 319–325. [Google Scholar] [CrossRef] [Green Version]
- Miklas, P.N.; Kelly, J.D.; Beebe, S.E.; Blair, M.W. Common bean breeding for resistance against biotic and abiotic stresses: From classical to MAS breeding. Euphytica 2006, 147, 105–131. [Google Scholar] [CrossRef]
- Herzog, E.; Frisch, M. Selection strategies for marker-assisted backcrossing with high-throughput marker systems. Theor. Appl. Genet. 2011, 123, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Blair, M.W.; Izquierdo, P. Use of the advanced backcross-QTL method to transfer seed mineral accumulation nutrition traits from wild to Andean cultivated common beans. Appl. Genet. 2012, 125, 1015–1031. [Google Scholar] [CrossRef]
- Oliveira, L.K.; Melo, L.C.; Brondani, C.; Peloso, M.J.D.; Brondani, R.P.V. Backcross assisted by microsatellite markers in common bean. Genet. Mol. Res. 2008, 7, 1000–1010. [Google Scholar] [CrossRef]
- Garzon, L.N.; Ligarreto, G.A.; Blair, M.W. Molecular marker-assisted backcrossing of anthracnose resistance into Andean climbing beans (Phaseolus vulgaris L.). Crop. Sci. 2008, 48, 562–570. [Google Scholar] [CrossRef]
- Crossa, J.; Perez-Rodriguez, P.; Cuevas, J.; Montesinos-Lopez, O.; Jarquin, D.; de Los Campos, G.; Burgueno, J.; Gonzalez-Camacho, J.M.; Perez-Elizalde, S.; Beyene, Y.; et al. Genomic Selection in Plant Breeding: Methods, Models, and Perspectives. Trends Plant. Sci. 2017, 22, 961–975. [Google Scholar] [CrossRef]
- Doudna, J.A.; Charpentier, E. Genome editing. The new frontier of genome engineering with CRISPR-Cas9. Science 2014, 346, 1258096. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.K.; Pickrell, J.K.; Coop, G. The Genetics of Human Adaptation: Hard Sweeps, Soft Sweeps, and Polygenic Adaptation. Curr. Biol. 2010, 20, R208–R215. [Google Scholar] [CrossRef] [Green Version]
- Desta, Z.A.; Ortiz, R. Genomic selection: Genome-wide prediction in plant improvement. Trends Plant Sci. 2014, 19, 592–601. [Google Scholar] [CrossRef]
- Grattapaglia, D.; Silva-Junior, O.B.; Resende, R.T.; Cappa, E.P.; Muller, B.S.F.; Tan, B.; Isik, F.; Ratcliffe, B.; El-Kassaby, Y.A. Quantitative Genetics and Genomics Converge to Accelerate Forest Tree Breeding. Front. Plant Sci. 2018, 9, 1693. [Google Scholar] [CrossRef] [PubMed]
- MacQueen, J.B. Some Methods for Classification and Analysis of Multivariate Observations. Proceedings of 5th Berkeley Symposium on Mathematical Statistics and Probability; University of California Press: Berkeley, CA, USA, 1967. [Google Scholar]
- Lloyd, S.P. Least-Squares Quantization in PCM. IEEE Trans. Inf. Theory 1982, 28, 129–137. [Google Scholar] [CrossRef]
- Edriss, V.; Gao, Y.; Zhang, X.; Jumbo, M.B.; Makumbi, D.; Olsen, M.S.; Crossa, J.; Packard, K.C.; Jannink, J.-L. Genomic Prediction in a Large African Maize Population. Crop. Sci. 2017, 57, 2361–2371. [Google Scholar] [CrossRef] [Green Version]
- Fe, D.; Cericola, F.; Byrne, S.; Lenk, I.; Ashraf, B.H.; Pedersen, M.G.; Roulund, N.; Asp, T.; Janss, L.; Jense, C.S.; et al. Genomic dissection and prediction of heading date in perennial ryegrass. BMC Genom. 2015, 16, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Kaufman, L.; Rousseeuw, P.J. Finding Groups in Data: An Introduction to Cluster Analysis; John Wiley & Sons: New York, NY, USA, 2009; Volume 344. [Google Scholar]
- Coffman, S.M.; Hufford, M.B.; Andorf, C.M.; Lübberstedt, T. Haplotype structure in commercial maize breeding programs in relation to key founder lines. Theor. Appl. Genet. 2020, 133, 547–561. [Google Scholar] [CrossRef] [Green Version]
- Turakulov, R.; Easteal, S. Number of SNPS Loci Needed to Detect Population Structure. Hum. Hered. 2003, 55, 37–45. [Google Scholar] [CrossRef]
- Murtagh, F. Multidimensional Clustering Algorithms; Physika Verlag: Vienna, Austria, 1985. [Google Scholar]
- Newell, M.A.; Cook, D.; Hofmann, H.; Jannink, J.-L. An algorithm for deciding the number of clusters and validation using simulated data with application to exploring crop population structure. Ann. Appl. Stat. 2013, 7, 1898–1916. [Google Scholar] [CrossRef] [Green Version]
- Tumino, G.; Voorrips, R.E.; Rizza, F.; Badeck, F.W.; Morcia, C.; Ghizzoni, R.; Germeier, C.U.; Paulo, M.J.; Terzi, V.; Smulders, M.J. Population structure and genome-wide association analysis for frost tolerance in oat using continuous SNP array signal intensity ratios. Theor. Appl. Genet. 2016, 129, 1711–1724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Li, Y.; Qiao, C.; Liu, X.; Hao, M.; Shugart, Y.Y.; Xiong, M.; Jin, L. Nuclear Norm Clustering: A promising alternative method for clustering tasks. Sci. Rep. 2018, 8, 10873. [Google Scholar] [CrossRef] [PubMed]
- Struyf, A.; Hubert, M.; Rousseeuw, P.J. Integrating robust clustering techniques in S-PLUS. Comput. Stat. Data Anal. 1997, 26, 17–37. [Google Scholar] [CrossRef]
- Gao, X.; Starmer, J.D. AWclust: Point-and-click software for non-parametric population structure analysis. BMC Bioinform. 2008, 9, 77. [Google Scholar] [CrossRef] [Green Version]
- D’Agostino, N.; Taranto, F.; Camposeo, S.; Mangini, G.; Fanelli, V.; Gadaleta, S.; Miazzi, M.M.; Pavan, S.; di Rienzo, V.; Sabetta, W.; et al. GBS-derived SNP catalogue unveiled wide genetic variability and geographical relationships of Italian olive cultivars. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taranto, F.; D’Agostino, N.; Greco, B.; Cardi, T.; Tripodi, P. Genome-wide SNP discovery and population structure analysis in pepper (Capsicum annuum) using genotyping by sequencing. BMC Genom. 2016, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caye, K.; Deist, T.M.; Martins, H.; Michel, O.; Francois, O. TESS3: Fast inference of spatial population structure and genome scans for selection. Mol. Ecol. Resour. 2016, 16, 540–548. [Google Scholar] [CrossRef]
- Stift, M.; Kolář, F.; Meirmans, P.G. STRUCTURE is more robust than other clustering methods in simulated mixed-ploidy populations. Heredity 2019, 123, 429–441. [Google Scholar] [CrossRef]
- Budiarto, A.; Mahesworo, B.; Baurley, J.; Suparyanto, T.; Pardamean, B. Fast and Effective Clustering Method for Ancestry Estimation. Procedia Comput. Sci. 2019, 157, 306–312. [Google Scholar] [CrossRef]
- Raj, A.; Stephens, M.; Pritchard, J.K. fastSTRUCTURE: Variational inference of population structure in large SNP data sets. Genetics 2014, 197, 573–589. [Google Scholar] [CrossRef] [Green Version]
- Reyes-Herrera, P.H.; Muñoz-Baena, L.; Velásquez-Zapata, V.; Patiño, L.; Delgado-Paz, O.A.; Díaz-Diez, C.A.; Navas-Arboleda, A.A.; Cortés, A.J. Inheritance of Rootstock Effects in Avocado (Persea americana Mill.) cv. Hass. Front. Plant Sci. 2020, 11, 555071. [Google Scholar] [CrossRef]
- de los Campos, G.; Hickey, J.M.; Pong-Wong, R.; Daetwyler, H.D.; Calus, M.P. Whole-genome regression and prediction methods applied to plant and animal breeding. Genetics 2013, 193, 327–345. [Google Scholar] [CrossRef] [Green Version]
- Gianola, D.; Fernando, R.L.; Stella, A. Genomic-assisted prediction of genetic value with semiparametric procedures. Genetics 2006, 173, 1761–1776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meuwissen, T.H.E.; Hayes, B.J.; Goddard, M.E. Prediction of Total Genetic Value Using Genome-Wide Dense Marker Maps. Genetics 2001, 157, 1819–1829. [Google Scholar] [CrossRef] [PubMed]
- Arenas, S.; Cortés, A.J.; Mastretta-Yanes, A.; Jaramillo-Correa, J.P. Evaluating the accuracy of genomic prediction for the management and conservation of relictual natural tree populations. Tree Genet. Genomes 2021, 17. [Google Scholar] [CrossRef]
- Mir, R.R.; Reynolds, M.; Pinto, F.; Khan, M.A.; Bhat, M.A. High-throughput phenotyping for crop improvement in the genomics era. Plant. Sci. 2019, 282, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Spindel, J.E.; Begum, H.; Akdemir, D.; Collard, B.; Redona, E.; Jannink, J.L.; McCouch, S. Genome-wide prediction models that incorporate de novo GWAS are a powerful new tool for tropical rice improvement. Hered. Edinb 2016, 116, 395–408. [Google Scholar] [CrossRef] [Green Version]
- Scott, M.F.; Ladejobi, O.; Amer, S.; Bentley, A.R.; Biernaskie, J.; Boden, S.A.; Clark, M.; Dell’Acqua, M.; Dixon, L.E.; Filippi, C.V.; et al. Multi-parent populations in crops: A toolbox integrating genomics and genetic mapping with breeding. Heredity 2020, 125, 396–416. [Google Scholar] [CrossRef]
- Blair, M.W.; Cortés, A.J.; Penmetsa, R.V.; Farmer, A.; Carrasquilla-Garcia, N.; Cook, D.R. A high-throughput SNP marker system for parental polymorphism screening, and diversity analysis in common bean (Phaseolus vulgaris L.). Theor. Appl. Genet. 2013, 126, 535–548. [Google Scholar] [CrossRef]
- Galeano, C.H.; Cortés, A.J.; Fernandez, A.C.; Soler, A.; Franco-Herrera, N.; Makunde, G.; Vanderleyden, J.; Blair, M.W. Gene-Based Single Nucleotide Polymorphism Markers for Genetic and Association Mapping in Common Bean. Bmc Genet. 2012, 13, 48. [Google Scholar] [CrossRef] [PubMed]
- Cortés, A.J.; Chavarro, M.C.; Blair, M.W. SNP marker diversity in common bean (Phaseolus vulgaris L.). Theor. Appl. Genet. 2011, 123, 827–845. [Google Scholar] [CrossRef] [PubMed]
- Keller, B.; Ariza-Suarez, D.; de la Hoz, J.; Aparicio, J.S.; Portilla-Benavides, A.E.; Buendia, H.F.; Mayor, V.M.; Studer, B.; Raatz, B. Genomic Prediction of Agronomic Traits in Common Bean (Phaseolus vulgaris L.) Under Environmental Stress. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Hickey, J.M.; Chiurugwi, T.; Mackay, I.; Powell, W.; Participants, I.G.S.i.C.B.P.W. Genomic prediction unifies animal and plant breeding programs to form platforms for biological discovery. Nat. Genet. 2017, 49, 1297–1303. [Google Scholar] [CrossRef] [PubMed]
- Kehel, Z.; Sanchez-Garcia, M.; El Baouchi, A.; Aberkane, H.; Tsivelikas, A.; Charles, C.; Amri, A. Predictive Characterization for Seed Morphometric Traits for Genebank Accessions Using Genomic Selection. Front. Ecol. Evol. 2020, 8. [Google Scholar] [CrossRef] [Green Version]
- Crossa, J.; Jarquin, D.; Franco, J.; Perez-Rodriguez, P.; Burgueno, J.; Saint-Pierre, C.; Vikram, P.; Sansaloni, C.; Petroli, C.; Akdemir, D.; et al. Genomic Prediction of Gene Bank Wheat Landraces. G3 Bethesda 2016, 6, 1819–1834. [Google Scholar] [CrossRef] [Green Version]
- Crossa, J.; Burgueno, J.; Dreisigacker, S.; Vargas, M.; Herrera-Foessel, S.A.; Lillemo, M.; Singh, R.P.; Trethowan, R.; Warburton, M.; Franco, J.; et al. Association Analysis of Historical Bread Wheat Germplasm Using Additive Genetic Covariance of Relatives and Population Structure. Genetics 2007, 177, 1889–1913. [Google Scholar] [CrossRef] [Green Version]
- Ishitani, M.; Rao, I.; Wenzl, P.; Beebe, S.; Tohme, J. Integration of genomics approach with traditional breeding towards improving abiotic stress adaptation: Drought and aluminum toxicity as case studies. Field Crop. Res. 2004, 90, 35–45. [Google Scholar] [CrossRef]
- Ravi, K.; Vadez, V.; Isobe, S.; Mir, R.; Guo, Y.; Nigam, S.; Gowda, M.; Radhakrishnan, T.; Bertioli, D.; Knapp, S.; et al. Identification of several small main-effect QTLs and a large number of epistatic QTLs for drought tolerance related traits in groundnut (Arachis hypogaea L.). Theor. Appl. Genet. 2011, 122, 1119–1132. [Google Scholar] [CrossRef] [Green Version]
- Roca Paixão, J.F.; Gillet, F.X.; Ribeiro, T.P.; Bournaud, C.; Lourenço-Tessutti, I.T.; Noriega, D.D.; Melo, B.P.; de Almeida-Engler, J.; Grossi-de-Sa, M.F. Improved drought stress tolerance in Arabidopsis by CRISPR/dCas9 fusion with a Histone AcetylTransferase. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Chen, L.; Li, R.; Zhao, R.; Yang, M.; Sheng, J.; Shen, L. Reduced Drought Tolerance by CRISPR/Cas9-Mediated SlMAPK3 Mutagenesis in Tomato Plants. J. Agric. Food Chem. 2017, 65, 8674–8682. [Google Scholar] [CrossRef]
- Li, R.; Liu, C.; Zhao, R.; Wang, L.; Chen, L.; Yu, W.; Zhang, S.; Sheng, J.; Shen, L. CRISPR/Cas9-Mediated SlNPR1 mutagenesis reduces tomato plant drought tolerance. BMC Plant. Biol. 2019, 19, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Leal, D.; Lemmon, Z.H.; Man, J.; Bartlett, M.E.; Lippman, Z.B. Engineering Quantitative Trait Variation for Crop Improvement by Genome Editing. Cell 2017, 171, 470–480.e478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemmon, Z.H.; Reem, N.T.; Dalrymple, J.; Soyk, S.; Swartwood, K.E.; Rodriguez-Leal, D.; Eck, J.V.; Lippman, Z.B. Rapid improvement of domestication traits in an orphan crop by genome editing. Nat. Plants 2018, 4, 766–770. [Google Scholar] [CrossRef] [PubMed]
- Uga, Y.; Sugimoto, K.; Ogawa, S.; Rane, J.; Ishitani, M.; Hara, N.; Kitomi, Y.; Inukai, Y.; Ono, K.; Kanno, N.; et al. Control of root system architecture by DEEPER ROOTING 1 increases rice yield under drought conditions. Nat. Genet. 2013, 45, 1097–1102. [Google Scholar] [CrossRef] [PubMed]
- Martignago, D.; Rico-Medina, A.; Blasco-Escamez, D.; Fontanet-Manzaneque, J.B.; Cano-Delgado, A.I. Drought Resistance by Engineering Plant Tissue-Specific Responses. Front. Plant Sci. 2019, 10, 1676. [Google Scholar] [CrossRef]
- Singh, R.K.; Svystun, T.; AlDahmash, B.; Jonsson, A.M.; Bhalerao, R.P. Photoperiod- and temperature-mediated control of phenology in trees—A molecular perspective. New Phytol. 2017, 213, 511–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, E.; Matsunaga, H.; Onogi, A.; Kajiya-Kanegae, H.; Minamikawa, M.; Suzuki, A.; Shirasawa, K.; Hirakawa, H.; Nunome, T.; Yamaguchi, H.; et al. A simulation-based breeding design that uses whole-genome prediction in tomato. Sci. Rep. 2016, 6, 19454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Migicovsky, Z.; Myles, S. Exploiting Wild Relatives for Genomics-assisted Breeding of Perennial Crops. Front. Plant Sci. 2017, 8, 460. [Google Scholar] [CrossRef] [Green Version]
- Watson, A.; Ghosh, S.; Williams, M.J.; Cuddy, W.S.; Simmonds, J.; Rey, M.D.; Asyraf Md Hatta, M.; Hinchliffe, A.; Steed, A.; Reynolds, D.; et al. Speed breeding is a powerful tool to accelerate crop research and breeding. Nat. Plants 2018, 4, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Cortés, A.J.; López-Hernández, F.; Osorio-Rodriguez, D. Predicting thermal adaptation by looking into populations’ genomic past. Front. Genet. 2020, 11, 564515. [Google Scholar] [CrossRef]
- Schrider, D.R.; Kern, A.D. Supervised Machine Learning for Population Genetics: A New Paradigm. Trends Genet. 2018, 34, 301–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, S.; Anderson, R.; Schapire, R. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Valencia, J.B.; Mesa, J.; León, J.G.; Madriñán, S.; Cortés, A.J. Climate Vulnerability Assessment of the Espeletia Complex on Páramo Sky Islands in the Northern Andes. Front. Ecol. Evol. 2020, 8. [Google Scholar] [CrossRef]
- Libbrecht, M.W.; Noble, W.S. Machine learning applications in genetics and genomics. Nat. Rev. Genet. 2015, 16, 321–332. [Google Scholar] [CrossRef] [Green Version]
- Payseur, B.A.; Schrider, D.R.; Kern, A.D. S/HIC: Robust Identification of Soft and Hard Sweeps Using Machine Learning. PLoS Genet. 2016, 12, e1005928. [Google Scholar] [CrossRef] [Green Version]
- Crossa, J.; Martini, J.W.R.; Gianola, D.; Perez-Rodriguez, P.; Jarquin, D.; Juliana, P.; Montesinos-Lopez, O.; Cuevas, J. Deep Kernel and Deep Learning for Genome-Based Prediction of Single Traits in Multienvironment Breeding Trials. Front. Genet. 2019, 10, 1168. [Google Scholar] [CrossRef] [PubMed]
- Abdollahi-Arpanahi, R.; Gianola, D.; Peñagaricano, F. Deep learning versus parametric and ensemble methods for genomic prediction of complex phenotypes. Genet. Sel. Evol. 2020, 52, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montesinos-Lopez, A.; Montesinos-Lopez, O.A.; Gianola, D.; Crossa, J.; Hernandez-Suarez, C.M. Multi-environment Genomic Prediction of Plant Traits Using Deep Learners With Dense Architecture. G3 2018, 8, 3813–3828. [Google Scholar] [CrossRef] [Green Version]
- Capblancq, T.; Fitzpatrick, M.C.; Bay, R.A.; Exposito-Alonso, M.; Keller, S.R. Genomic Prediction of (Mal)Adaptation Across Current and Future Climatic Landscapes. Annu Rev. Ecol. Evol. Syst 2020, 51, 245–269. [Google Scholar] [CrossRef]
- Sebestyén, V.; Czvetkó, T.; Abonyi, J. The Applicability of Big Data in Climate Change Research: The Importance of System of Systems Thinking. Front. Environ. Sci. 2021, 9, 619092. [Google Scholar] [CrossRef]
- Tong, H.; Nikoloski, Z. Machine learning approaches for crop improvement: Leveraging phenotypic and genotypic big data. J. Plant. Physiol. 2021, 257, 153354. [Google Scholar] [CrossRef] [PubMed]
- Heslot, N.; Yang, H.P.; Sorrells, M.E.; Jannink, J.L. Genomic Selection in Plant Breeding: A Comparison of Models. Crop. Sci. 2012, 52, 146–160. [Google Scholar] [CrossRef]
- Rosado, R.D.S.; Cruz, C.D.; Barili, L.D.; de Souza Carneiro, J.E.; Carneiro, P.C.S.; Carneiro, V.Q.; da Silva, J.T.; Nascimento, M. Artificial Neural Networks in the Prediction of Genetic Merit to Flowering Traits in Bean Cultivars. Agriculture 2020, 10, 638. [Google Scholar] [CrossRef]
- Koech, R.K.; Malebe, P.M.; Nyarukowa, C.; Mose, R.; Kamunya, S.M.; Loots, T.; Apostolides, Z.; Léon, J. Genome-enabled prediction models for black tea (Camellia sinensis) quality and drought tolerance traits. Plant. Breed. 2020, 139, 1003–1015. [Google Scholar] [CrossRef]
- Roorkiwal, M.; Rathore, A.; Das, R.R.; Singh, M.K.; Jain, A.; Srinivasan, S.; Gaur, P.M.; Chellapilla, B.; Tripathi, S.; Li, Y.; et al. Genome-Enabled Prediction Models for Yield Related Traits in Chickpea. Front. Plant. Sci. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Sousa, I.C.d.; Nascimento, M.; Silva, G.N.; Nascimento, A.C.C.; Cruz, C.D.; Silva, F.F.; Almeida, D.P.d.; Pestana, K.N.; Azevedo, C.F.; Zambolim, L.; et al. Genomic prediction of leaf rust resistance to Arabica coffee using machine learning algorithms. Sci. Agric. 2021, 78. [Google Scholar] [CrossRef]
- Ferrão, L.-F.V.; Ferrão, R.G.; Ferrão, M.-A.G.; Fonseca, A.; Carbonetto, P.; Stephens, M.; Garcia, A.-A.F. Accurate genomic prediction of Coffea canephora in multiple environments using whole-genome statistical models. Heredity 2019, 122, 261–275. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Wang, X.; Ding, X.; Zheng, X.; Yang, Z.; Xu, C.; Hu, Z. Genomic selection of agronomic traits in hybrid rice using an NCII population. Rice 2018, 11, 1–10. [Google Scholar] [CrossRef] [Green Version]
- González-Camacho, J.M.; de los Campos, G.; Pérez, P.; Gianola, D.; Cairns, J.E.; Mahuku, G.; Babu, R.; Crossa, J. Genome-enabled prediction of genetic values using radial basis function neural networks. Theor. Appl. Genet. 2012, 125, 759–771. [Google Scholar] [CrossRef] [Green Version]
- Khaki, S.; Wang, L. Crop Yield Prediction Using Deep Neural Networks. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Rachmatia, H.; Kusuma, W.A.; Hasibuan, L.S. Prediction of maize phenotype based on whole-genome single nucleotide polymorphisms using deep belief networks. J. Phys. Conf. Ser. 2017, 835, 012003. [Google Scholar] [CrossRef] [Green Version]
- Maenhout, S.; Baets, B.D.; Haesaert, G.; Bockstaele, E.V. Support vector machine regression for the prediction of maize hybrid performance. Theor. Appl. Genet. 2007, 115, 1003–1013. [Google Scholar] [CrossRef] [PubMed]
- González-Camacho, J.M.; Crossa, J.; Pérez-Rodríguez, P.; Ornella, L.; Gianola, D. Genome-enabled prediction using probabilistic neural network classifiers. BMC Genom. 2016, 17, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shikha, M.; Kanika, A.; Rao, A.R.; Mallikarjuna, M.G.; Gupta, H.S.; Nepolean, T. Genomic Selection for Drought Tolerance Using Genome-Wide SNPs in Maize. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef]
- Azodi, C.B.; Bolger, E.; McCarren, A.; Roantree, M.; de los Campos, G.; Shiu, S.-H. Benchmarking Parametric and Machine Learning Models for Genomic Prediction of Complex Traits. G3 2019, 9, 3691–3702. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; Lai, X.; Liu, D.; Zhang, Z.; Ma, P.; Wang, Q.; Zhang, Z.; Pan, Y. Applications of Support Vector Machine in Genomic Prediction in Pig and Maize Populations. Front. Genet. 2020, 11. [Google Scholar] [CrossRef]
- Li, G.; Dong, Y.; Zhao, Y.; Tian, X.; Würschum, T.; Xue, J.; Chen, S.; Reif, J.C.; Xu, S.; Liu, W. Genome-wide prediction in a hybrid maize population adapted to Northwest China. Crop. J. 2020, 8, 830–842. [Google Scholar] [CrossRef]
- Montesinos-López, O.A.; Montesinos-López, A.; Mosqueda-Gonzalez, B.A.; Montesinos-López, J.C.; Crossa, J.; Ramirez, N.L.; Singh, P.; Valladares-Anguiano, F.A.; Akhunov, E. A zero altered Poisson random forest model for genomic-enabled prediction. G3 2021, 11. [Google Scholar] [CrossRef]
- Neves, H.H.; Carvalheiro, R.; Queiroz, S.A. A comparison of statistical methods for genomic selection in a mice population. BMC Genet. 2012, 13, 100. [Google Scholar] [CrossRef] [Green Version]
- Annicchiarico, P.; Nazzicari, N.; Pecetti, L.; Romani, M.; Ferrari, B.; Wei, Y.; Brummer, E.C. GBS-Based Genomic Selection for Pea Grain Yield under Severe Terminal Drought. Plant Genome 2017, 10. [Google Scholar] [CrossRef]
- Grinberg, N.F.; Lovatt, A.; Hegarty, M.; Lovatt, A.; Skøt, K.P.; Kelly, R.; Blackmore, T.; Thorogood, D.; King, R.D.; Armstead, I.; et al. Implementation of Genomic Prediction in Lolium perenne (L.) Breeding Populations. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, R.; Marathi, B.; Singh1, M. Efficient genomic selection using ensemble learning and ensemble feature reduction. J. Crop. Sci. Biotechnol. 2020, 23, 311–323. [Google Scholar] [CrossRef]
- Onogi, A.; Ideta, O.; Inoshita, Y.; Ebana, K.; Yoshioka, T.; Yamasaki, M.; Iwata, H. Exploring the areas of applicability of whole-genome prediction methods for Asian rice (Oryza sativa L.). Theor. Appl. Genet. 2015, 128, 41–53. [Google Scholar] [CrossRef]
- Ogutu, J.O.; Piepho, H.-P.; Schulz-Streeck, T. A comparison of random forests, boosting and support vector machines for genomic selection. BMC Proc. 2011, 5, S11. [Google Scholar] [CrossRef] [Green Version]
- Zingaretti, L.M.; Gezan, S.A.; Ferrão, L.F.V.; Osorio, L.F.; Monfort, A.; Muñoz, P.R.; Whitaker, V.M.; Pérez-Enciso, M. Exploring Deep Learning for Complex Trait Genomic Prediction in Polyploid Outcrossing Species. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gianola, D.; Okut, H.; Weigel, K.A.; Rosa, G.J. Predicting complex quantitative traits with Bayesian neural networks: A case study with Jersey cows and wheat. BMC Genet. 2011, 12, 87. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Rodríguez, P.; Gianola, D.; González-Camacho, J.M.; Crossa, J.; Manès, Y.; Dreisigacker, S. Comparison Between Linear and Non-parametric Regression Models for Genome-Enabled Prediction in Wheat. G3 Genes Genomes Genet. 2012, 2, 1595–1605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; Khan, J.; Pradhan, S.; Shahi, D.; Khan, N.; Avci, M.; McBreen, J.; Harrison, S.; Brown-Guedira, G.; Murphy, J.P.; et al. Multi-Trait Genomic Prediction of Yield-Related Traits in US Soft Wheat under Variable Water Regimes. Genes 2020, 11, 1270. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Qiu, Z.; Song, J.; Cheng, Q.; Ma, C. DeepGS: Predicting phenotypes from geno- types using Deep Learning. bioRxiv 2017. [Google Scholar] [CrossRef] [Green Version]
- Grinberg, N.F.; Orhobor, O.I.; King, R.D. An evaluation of machine-learning for predicting phenotype: Studies in yeast, rice, and wheat. Mach. Learn. 2020, 109, 251–277. [Google Scholar] [CrossRef] [Green Version]
- Poland, J.; Endelman, J.; Dawson, J.; Rutkoski, J.; Wu, S.; Manes, Y.; Dreisigacker, S.; Crossa, J.; Sánchez-Villeda, H.; Sorrells, M.; et al. Genomic Selection in Wheat Breeding using Genotyping-by-Sequencing. Plant. Genome 2012, 5. [Google Scholar] [CrossRef] [Green Version]
- Arruda, M.P.; Brown, P.J.; Lipka, A.E.; Krill, A.M.; Thurber, C.; Kolb, F.L. Genomic Selection for Predicting Fusarium Head Blight Resistance in a Wheat Breeding Program. Plant. Genome 2015, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montesinos-Lopez, O.A.; Martin-Vallejo, J.; Crossa, J.; Gianola, D.; Hernandez-Suarez, C.M.; Montesinos-Lopez, A.; Juliana, P.; Singh, R. A Benchmarking Between Deep Learning, Support Vector Machine and Bayesian Threshold Best Linear Unbiased Prediction for Predicting Ordinal Traits in Plant Breeding. G3 2019, 9, 601–618. [Google Scholar] [CrossRef] [Green Version]
- Labroo, M.R.; Studer, A.J.; Rutkoski, J.E. Heterosis and Hybrid Crop Breeding: A Multidisciplinary Review. Front. Genet. 2021, 12. [Google Scholar] [CrossRef]
- Technow, F.; Schrag, T.A.; Schipprack, W.; Bauer, E.; Simianer, H.; Melchinger, A.E. Genome properties and prospects of genomic prediction of hybrid performance in a breeding program of maize. Genetics 2014, 197, 1343–1355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, B.; Grattapaglia, D.; Martins, G.S.; Ferreira, K.Z.; Sundberg, B.; Ingvarsson, P.r.K. Evaluating the accuracy of genomic prediction of growth and wood traits in two Eucalyptus species and their F1 hybrids. BMC Plant Biol. 2017, 17, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holliday, J.A.; Aitken, S.N.; Cooke, J.E.; Fady, B.; González-Martínez, S.C.; Heuertz, M.; Jaramillo-Correa, J.P.; Lexer, C.; Staton, M.; Whetten, R.W.; et al. Advances in ecological genomics in forest trees and applications to genetic resources conservation and breeding. Mol. Ecol. 2017, 26, 706–717. [Google Scholar] [CrossRef] [PubMed]
- Robert, C.P.; Gautier, M.; Cornuet, J.-M.; Estoup, A.; Marin, J.-M.; Pudlo, P. Reliable ABC model choice via random forests. Bioinformatics 2016, 32, 859–866. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Z.; Cheng, Q.; Song, J.; Tang, Y.; Ma, C. Application of Machine Learning-Based Classification to Genomic Selection and Performance Improvement. Comput. Sci. 2016, 9771, 412–421. [Google Scholar] [CrossRef]
- Park, D.S.; Willis, C.G.; Xi, Z.; Kartesz, J.T.; Davis, C.C.; Worthington, S. Machine Learning Predicts Large Scale Declines in Native Plant Phylogenetic Diversity. New Phytol. 2020. [Google Scholar] [CrossRef]
- Ma, C.; Zhang, H.H.; Wang, X. Machine learning for Big Data analytics in plants. Trends Plant. Sci. 2014, 19, 798–808. [Google Scholar] [CrossRef]
- Ma, C.; Xin, M.; Feldmann, K.A.; Wang, X. Machine Learning–Based Differential Network Analysis: A Study of Stress-Responsive Transcriptomes in Arabidopsis. Plant. Cell 2014, 26, 520–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Cimen, E.; Singh, N.; Buckler, E. Deep learning for plant genomics and crop improvement. Curr. Opin. Plant. Biol. 2020, 54, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Montesinos-López, O.A.; Montesinos-López, A.; Pérez-Rodríguez, P.; Barrón-López, J.A.; Martini, J.W.R.; Fajardo-Flores, S.B.; Gaytan-Lugo, L.S.; Santana-Mancilla, P.C.; Crossa, J. A review of deep learning applications for genomic selection. BMC Genom. 2021, 22, 19. [Google Scholar] [CrossRef] [PubMed]
- Khoury, C.K.; Carver, D.; Greene, S.L.; Williams, K.A.; Achicanoy, H.A.; Schori, M.; León, B.; Wiersema, J.H.; Frances, A. Crop wild relatives of the United States require urgent conservation action. Proc. Natl. Acad. Sci. USA 2020, 117, 33351–33357. [Google Scholar] [CrossRef] [PubMed]
- Li, D.-Z.; Pritchard, H.W. The science and economics of ex situ plant conservation. Trends Plant. Sci. 2009, 14, 614–621. [Google Scholar] [CrossRef] [PubMed]
- Tyack, N.; Dempewolf, H.; Khoury, C.K. The Potential of Payment for Ecosystem Services for Crop Wild Relative Conservation. Plants 2020, 9, 1305. [Google Scholar] [CrossRef]
- Meuwissen, T.H.E.; Sonesson, A.K.; Gebregiwergis, G.; Woolliams, J.A. Management of Genetic Diversity in the Era of Genomics. Front. Genet. 2020, 11. [Google Scholar] [CrossRef]
- Girma, G.; Nida, H.; Tirfessa, A.; Lule, D.; Bejiga, T.; Seyoum, A.; Mekonen, M.; Nega, A.; Dessalegn, K.; Birhanu, C.; et al. A comprehensive phenotypic and genomic characterization of Ethiopian sorghum germplasm defines core collection and reveals rich genetic potential in adaptive traits. Plant. Genome 2020. [Google Scholar] [CrossRef]
- Scherer, L.; Svenning, J.C.; Huang, J.; Seymour, C.L.; Sandel, B.; Mueller, N.; Kummu, M.; Bekunda, M.; Bruelheide, H.; Hochman, Z.; et al. Global priorities of environmental issues to combat food insecurity and biodiversity loss. Sci. Total Environ. 2020, 730, 139096. [Google Scholar] [CrossRef]
- Vincent, H.; Wiersema, J.; Kell, S.; Fielder, H.; Dobbie, S.; Castañeda-Álvarez, N.P.; Guarino, L.; Eastwood, R.; Leόn, B.; Maxted, N. A prioritized crop wild relative inventory to help underpin global food security. Biol. Conserv. 2013, 167, 265–275. [Google Scholar] [CrossRef]
- Pironon, S.; Borrell, J.S.; Ondo, I.; Douglas, R.; Phillips, C.; Khoury, C.K.; Kantar, M.B.; Fumia, N.; Soto Gomez, M.; Viruel, J.; et al. Toward Unifying Global Hotspots of Wild and Domesticated Biodiversity. Plants 2020, 9, 1128. [Google Scholar] [CrossRef] [PubMed]
- Burgarella, C.; Barnaud, A.; Kane, N.A.; Jankowski, F.; Scarcelli, N.; Billot, C.; Vigouroux, Y.; Berthouly-Salazar, C. Adaptive Introgression: An Untapped Evolutionary Mechanism for Crop Adaptation. Front. Plant Sci. 2019, 10, 4. [Google Scholar] [CrossRef] [PubMed]
- Santantonio, N.; Atanda, S.A.; Beyene, Y.; Varshney, R.K.; Olsen, M.; Jones, E.; Roorkiwal, M.; Gowda, M.; Bharadwaj, C.; Gaur, P.M.; et al. Strategies for Effective Use of Genomic Information in Crop Breeding Programs Serving Africa and South Asia. Front. Plant. Sci. 2020, 11. [Google Scholar] [CrossRef] [Green Version]
- Trucchi, E.; Benazzo, A.; Lari, M.; Iob, A.; Vai, S.; Nanni, L.; Bellucci, E.; Bitocchi, E.; Raffini, F.; Xu, C.; et al. Ancient genomes reveal early Andean farmers selected common beans while preserving diversity. Nat. Plants 2021. [Google Scholar] [CrossRef]
- Ulian, T.; Diazgranados, M.; Pironon, S.; Padulosi, S.; Liu, U.; Davies, L.; Howes, M.J.R.; Borrell, J.S.; Ondo, I.; Pérez-Escobar, O.A.; et al. Unlocking plant resources to support food security and promote sustainable agriculture. Plants People Planet 2020, 2, 421–445. [Google Scholar] [CrossRef]
- Runcie, D.E.; Qu, J.; Cheng, H.; Crawford, L. Mega-scale linear mixed models for genomic predictions with thousands of traits. bioRxiv 2020. [Google Scholar] [CrossRef]
- Cross, J.; Cerón-Rojas, J. Multi-trait Multi-environment Genome-wide Molecular Marker Selection Indices. J. Indian Soc. Agric. Stat. 2011, 65, 125–142. [Google Scholar]
- Juliana, P.; Singh, R.P.; Braun, H.-J.; Huerta-Espino, J.; Crespo-Herrera, L.; Payne, T.; Poland, J.; Shrestha, S.; Kumar, U.; Joshi, A.K.; et al. Retrospective Quantitative Genetic Analysis and Genomic Prediction of Global Wheat Yields. Front. Plant. Sci. 2020, 11. [Google Scholar] [CrossRef]
- Resende, R.T.; Piepho, H.P.; Rosa, G.J.M.; Silva-Junior, O.B.; Silva, F.F.e.; Resende, M.D.V.d.; Grattapaglia, D. Enviromics in breeding: Applications and perspectives on envirotypic-assisted selection. Theor. Appl. Genet. 2020. [Google Scholar] [CrossRef]
- Costa-Neto, G.; Fritsche-Neto, R.; Crossa, J. Nonlinear kernels, dominance, and envirotyping data increase the accuracy of genome-based prediction in multi-environment trials. Heredity 2020. [Google Scholar] [CrossRef]
- Graham, L.J.; Spake, R.; Gillings, S.; Watts, K.; Eigenbrod, F.; Isaac, N. Incorporating fine-scale environmental heterogeneity into broad-extent models. Methods Ecol. Evol. 2019, 10, 767–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alves, F.C.; Balmant, K.M.; Resende, M.F.R.; Kirst, M.; los Campos, G. Accelerating forest tree breeding by integrating genomic selection and greenhouse phenotyping. Plant. Genome 2020, 13. [Google Scholar] [CrossRef] [PubMed]
- Volpato, L.; Pinto, F.; González-Pérez, L.; Thompson, I.G.; Borém, A.; Reynolds, M.; Gérard, B.; Molero, G.; Rodrigues, F.A. High Throughput Field Phenotyping for Plant Height Using UAV-Based RGB Imagery in Wheat Breeding Lines: Feasibility and Validation. Front. Plant. Sci. 2021, 12. [Google Scholar] [CrossRef]
- Krause, M.R.; González-Pérez, L.; Crossa, J.; Pérez-Rodríguez, P.; Montesinos-López, O.; Singh, R.P.; Dreisigacker, S.; Poland, J.; Rutkoski, J.; Sorrells, M.; et al. Hyperspectral Reflectance-Derived Relationship Matrices for Genomic Prediction of Grain Yield in Wheat. G3 Genes Genomes Genet. 2019. [Google Scholar] [CrossRef] [Green Version]
- Lozada, D.N.; Godoy, J.V.; Ward, B.P.; Carter, A.H. Genomic Prediction and Indirect Selection for Grain Yield in US Pacific Northwest Winter Wheat Using Spectral Reflectance Indices from High-Throughput Phenotyping. Int. J. Mol. Sci. 2019, 21, 165. [Google Scholar] [CrossRef] [Green Version]
- Roudbar, M.A.; Momen, M.; Mousavi, S.F.; Ardestani, S.S.; Lopes, F.B.; Gianola, D.; Khatib, H. Genome-wide methylation prediction of biological age using reproducing kernel Hilbert spaces and Bayesian ridge regressions. bioRxiv 2020. [Google Scholar] [CrossRef]
- Cortés, A.J.; Restrepo-Montoya, M.; Bedoya-Canas, L.E. Modern Strategies to Assess and Breed Forest Tree Adaptation to Changing Climate. Front. Plant. Sci. 2020, 11, 583323. [Google Scholar] [CrossRef]
- Cortinovis, G.; Di Vittori, V.; Bellucci, E.; Bitocchi, E.; Papa, R. Adaptation to novel environments during crop diversification. Curr. Opin. Plant. Biol. 2020. [Google Scholar] [CrossRef]
- Aguirre-Liguori, J.A.; Gaut, B.S.; Jaramillo-Correa, J.P.; Tenaillon, M.I.; Montes-Hernández, S.; García-Oliva, F.; Hearne, S.J.; Eguiarte, L.E. Divergence with gene flow is driven by local adaptation to temperature and soil phosphorus concentration in teosinte subspecies (Zea mays parviglumis and Zea mays mexicana). Mol. Ecol. 2019, 28, 2814–2830. [Google Scholar] [CrossRef]
- Ellstrand, N.C.; Prentice, H.C.; Hancock, J.F. Gene Flow and Introgression from Domesticated Plants into their Wild Relatives. Annu. Rev. Ecol. Evol. Syst. 1999, 30, 539–563. [Google Scholar] [CrossRef]
- Todesco, M.; Owens, G.L.; Bercovich, N.; Légaré, J.S.; Soudi, S.; Burge, D.O.; Huang, K.; Ostevik, K.L.; Drummond, E.; Imerovski, I.; et al. Massive haplotypes underlie ecotypic differentiation in sunflowers. Nature 2020, 602–607. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.; Andrew, R.L.; Owens, G.L.; Ostevik, K.L.; Rieseberg1, L.H. Multiple chromosomal inversions contribute to adaptive divergence of a dune sunflower ecotype. Mol. Ecol. 2020, 2020, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.; Rieseberg, L.H. Frequency, Origins, and Evolutionary Role of Chromosomal Inversions in Plants. Front. Plant Sci. 2020, 11, 296. [Google Scholar] [CrossRef]
- Yu, X.; Li, X.; Guo, T.; Zhu, C.; Wu, Y.; Mitchell, S.E.; Roozeboom, K.L.; Wang, D.; Wang, M.L.; Pederson, G.A.; et al. Genomic prediction contributing to a promising global strategy to turbocharge gene banks. Nat. Plants 2016, 2, 16150. [Google Scholar] [CrossRef]
- Bayer, P.E.; Golicz, A.A.; Scheben, A.; Batley, J.; Edwards, D. Plant pan-genomes are the new reference. Nat. Plants 2020, 6, 914–920. [Google Scholar] [CrossRef] [PubMed]
- Barrera-Redondo, J.; Pinero, D.; Eguiarte, L.E. Genomic, Transcriptomic and Epigenomic Tools to Study the Domestication of Plants and Animals: A Field Guide for Beginners. Front. Genet. 2020, 11, 742. [Google Scholar] [CrossRef]
- Antonelli, A.; Smith, R.J.; Simmonds, M.S.J. Unlocking the properties of plants and fungi for sustainable development. Nat. Plants 2019, 5, 1100–1102. [Google Scholar] [CrossRef]
- Wang, W.; Cao, X.H.; Miclăuș, M.; Xu, J.; Xiong, W. The Promise of Agriculture Genomics. Int. J. Genom. 2017, 2017, 1–3. [Google Scholar] [CrossRef]
- Singh, S.; Vikram, P.; Sehgal, D.; Burgueño, J.; Sharma, A.; Singh, S.K.; Sansaloni, C.P.; Joynson, R.; Brabbs, T.; Ortiz, C.; et al. Harnessing genetic potential of wheat germplasm banks through impact-oriented-prebreeding for future food and nutritional security. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Ripple, W.J.; Wolf, C.; Newsome, T.M.; Barnard, P.; Moomaw, W.R. World Scientists’ Warning of a Climate Emergency. BioScience 2020, 70, 8–12. [Google Scholar] [CrossRef]
- Kim, H.; Jung, J.; Singh, N.; Greenberg, A.; Doyle, J.J.; Tyagi, W.; Chung, J.W.; Kimball, J.; Hamilton, R.S.; McCouch, S.R. Population Dynamics Among six Major Groups of the Oryza rufipogon Species Complex, Wild Relative of Cultivated Asian Rice. Rice 2016, 9, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyle, E.A.; Li, Y.I.; Pritchard, J.K. An Expanded View of Complex Traits: From Polygenic to Omnigenic. Cell 2017, 169, 1177–1186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miska, E.A.; Ferguson-Smith, A.C. Transgenerational inheritance: Models and mechanisms of non–DNA sequence–based inheritance. Science 2016, 354, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Piot, A.; Prunier, J.; Isabel, N.; Klapste, J.; El-Kassaby, Y.A.; Villarreal Aguilar, J.C.; Porth, I. Genomic Diversity Evaluation of Populus trichocarpa Germplasm for Rare Variant Genetic Association Studies. Front. Genet. 2019, 10, 1384. [Google Scholar] [CrossRef] [PubMed]
- Olatoye, M.O.; Hu, Z.; Aikpokpodion, P.O. Epistasis Detection and Modeling for Genomic Selection in Cowpea (Vigna unguiculata L. Walp.). Front. Genet. 2019, 10, 677. [Google Scholar] [CrossRef] [Green Version]
- Dwivedi, S.L.; Ceccarelli, S.; Blair, M.W.; Upadhyaya, H.D.; Are, A.K.; Ortiz, R. Landrace Germplasm for Improving Yield and Abiotic Stress Adaptation. Trends Plant. Sci. 2016, 21, 31–42. [Google Scholar] [CrossRef]
- Coyne, C.J.; Kumar, S.; von Wettberg, E.J.B.; Marques, E.; Berger, J.D.; Redden, R.J.; Ellis, T.H.N.; Brus, J.; Zablatzká, L.; Smýkal, P. Potential and limits of exploitation of crop wild relatives for pea, lentil, and chickpea improvement. Legume Sci. 2020, e36. [Google Scholar] [CrossRef] [Green Version]
- Langridge, P.; Robbie, W. Harnessing the potential of germplasm collections. Nat. Genet. 2019, 51, 200–201. [Google Scholar] [CrossRef]
- Dempewolf, H.; Baute, G.; Anderson, J.; Kilian, B.; Smith, C.; Guarino, L. Past and Future Use of Wild Relatives in Crop Breeding. Crop. Sci. 2017, 57, 1070–1082. [Google Scholar] [CrossRef]
- Joly-Lopez, Z.; Flowers, J.M.; Purugganan, M.D. Developing maps of fitness consequences for plant genomes. Curr. Opin. Plant. Biol. 2016, 30, 101–107. [Google Scholar] [CrossRef] [Green Version]
- Timpson, N.J.; Greenwood, C.M.T.; Soranzo, N.; Lawson, D.J.; Richards, J.B. Genetic architecture: The shape of the genetic contribution to human traits and disease. Nat. Rev. Genet. 2018, 19, 110–124. [Google Scholar] [CrossRef] [PubMed]
- Blair, M.W.; Cortes, A.J.; Farmer, A.D.; Huang, W.; Ambachew, D.; Penmetsa, R.V.; Carrasquilla-Garcia, N.; Assefa, T.; Cannon, S.B. Uneven recombination rate and linkage disequilibrium across a reference SNP map for common bean (Phaseolus vulgaris L.). PLoS ONE 2018, 13, e0189597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slatkin, M. Linkage disequilibrium—Understanding the evolutionary past and mapping the medical future. Nat. Rev. Genet. 2008, 9, 477–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelleher, C.T.; Wilkin, J.; Zhuang, J.; Cortés, A.J.; Quintero, Á.L.P.; Gallagher, T.F.; Bohlmann, J.; Douglas, C.J.; Ellis, B.E.; Ritland, K. SNP discovery, gene diversity, and linkage disequilibrium in wild populations of Populus tremuloides. Tree Genet. Genomes 2012, 8, 821–829. [Google Scholar] [CrossRef]
- Barghi, N.; Hermisson, J.; Schloötterer, C. Polygenic adaptation: A unifying framework to understand positive selection. Nat. Rev. Genet. 2020, 21, 769–781. [Google Scholar] [CrossRef]
- Resende, M.D.V.; Resende, M.F.R., Jr.; Sansaloni, C.P.; Petroli, C.D.; Missiaggia, A.A.; Aguiar, A.M.; Abad, J.M.; Takahashi, E.K.; Rosado, A.M.; Faria, D.A.; et al. Genomic selection for growth and wood quality in Eucalyptus: Capturing the missing heritability and accelerating breeding for complex traits in forest trees. New Phytol. 2012, 194, 116–128. [Google Scholar] [CrossRef]
- de los Campos, G.; Sorensen, D.; Gianola, D. Genomic heritability: What is it? PLoS Genet. 2015, 11, e1005048. [Google Scholar] [CrossRef] [Green Version]
- Reyes-Valdes, M.H.; Burgueno, J.; Singh, S.; Martinez, O.; Sansaloni, C.P. An informational view of accession rarity and allele specificity in germplasm banks for management and conservation. PLoS ONE 2018, 13, e0193346. [Google Scholar] [CrossRef] [Green Version]
- Tollenaar, M.; Fridgen, J.; Tyagi, P.; Stackhouse, P.W., Jr.; Kumudini, S. The contribution of solar brightening to the US maize yield trend. Nat. Clim. Chang. 2017, 7, 275–278. [Google Scholar] [CrossRef]
- Abberton, M.; Batley, J.; Bentley, A.; Bryant, J.; Cai, H.; Cockram, J.; de Oliveira, A.C.; Cseke, L.J.; Dempewolf, H.; De Pace, C.; et al. Global agricultural intensification during climate change: A role for genomics. Plant. Biotechnol. J. 2016, 14, 1095–1098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, R.; Iwata, H. Bayesian optimization for genomic selection: A method for discovering the best genotype among a large number of candidates. Theor. Appl. Genet. 2018, 131, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Lascoux, M.; Glémin, S.; Savolainen, O. Local Adaptation in Plants. Encycl. Life Sci. 2016, 0025270, 1–7. [Google Scholar] [CrossRef]
- Rippke, U.; Ramirez-Villegas, J.; Jarvis, A.; Vermeulen, S.J.; Parker, L.; Mer, F.; Diekkrüger, B.; Challinor, A.J.; Howden, M. Timescales of transformational climate change adaptation in sub-Saharan African agriculture. Nat. Clim. Chang. 2016, 6, 605–609. [Google Scholar] [CrossRef]
- Zhang, W.; Cao, G.; Li, X.; Zhang, H.; Wang, C.; Liu, Q.; Chen, X.; Cui, Z.; Shen, J.; Jiang, R.; et al. Closing yield gaps in China by empowering smallholder farmers. Nature 2016, 537, 671–674. [Google Scholar] [CrossRef] [PubMed]
- Tester, M.; Langridge, P. Breeding Technologies to Increase Crop Production in a Changing World. Science 2010, 327, 818–822. [Google Scholar] [CrossRef]
- McCouch, S.R.; Wright, M.H.; Tung, C.W.; Maron, L.G.; McNally, K.L.; Fitzgerald, M.; Singh, N.; DeClerck, G.; Agosto-Perez, F.; Korniliev, P.; et al. Open access resources for genome-wide association mapping in rice. Nat. Commun. 2016, 7, 10532. [Google Scholar] [CrossRef] [Green Version]
- Spindel, J.E.; McCouch, S.R. When more is better: How data sharing would accelerate genomic selection of crop plants. New Phytol. 2016, 212, 814–826. [Google Scholar] [CrossRef]
- McCouch, S.; Navabi, K.; Abberton, M.; Anglin, N.L.; Barbieri, R.L.; Baum, M.; Bett, K.; Booker, H.; Brown, G.L.; Bryan, G.J.; et al. Mobilizing Crop Biodiversity. Mol. Plant 2020, 13, 1341–1344. [Google Scholar] [CrossRef]
- Spiegelhalter, D.; Pearson, M.; Short, I. Visualizing Uncertainty About the Future. Science 2011, 333, 1393–1400. [Google Scholar] [CrossRef] [Green Version]
- McCouch, S. Diversifying Selection in Plant Breeding. PLoS Biol. 2004, 2, 1507–1512. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
A roadmap of trans-disciplinary approaches aiming at harnessing genebank utilization for climate change research in the face of heat, and water scarcity. Compiling (a) previous characterizations and (b) geo-referencing-derived climate data/indices of available genetic resources in genebanks is a starting point to (c) assess the extent of abiotic stress tolerance among existing accessions, and the need of (d) new habitat-based population-guided collections targeting isolated pockets of cryptic diversity in dry and semi-arid regions. Planning question-oriented collecting trips of crop wild relatives and hidden landraces across contrasting environments/agro-ecologies is needed now more than ever, despite a century of gathering and preserving diversity in plants throughout genebanks. Coupling ex situ agro-ecological screenings together with (e) ongoing in situ genebanks characterizations for morphological and genetic variation is essential to define (c) putative tolerant reference collections, while understanding the (f) heritability (h2) of adaptive traits and their genetic architecture (i.e., underlying genes) via genome-wide selection scans (GWSS), genome–environment associations (GEA), and genome-wide association studies (GWAS). Since identifying these novel sources of heat and drought tolerance demands merging heterogeneous datasets, (g) machine learning (ML, in red letters) promises speeding up genebank characterization. The distinction that clustering (Table 1) and ML (Figure 2 and Table 2) strategies can provide between abiotic stress tolerant and susceptible accessions is essential to (h) transfer useful genetic variation from wild crop donors and early landraces into elite cultivated lines, either by designing (i) genomic-assisted breeding programs such as genomic prediction (GP) and inter-specific marker- and genomic-assisted backcrossing (MAB and GABC) schemes, or by envisioning (j) multi-trait gene editing strategies (e.g., CRISPR-Cas9). Once (k) abiotic stress tolerant varieties are validated across different environments, (l) legal inscription, seed multiplication, seed delivery system to farmers’ associations, and (m) follow-up given the regional needs, market demands, and adoption potential, are necessary downstream validation steps. These heterogeneous datasets are also likely to be inputted into ML, and in turn feedback new needs beyond heat and drought tolerance such as other types of resistances and nutritional quality. For ML to succeed speeding up the breeding of heat and drought-tolerant crops, there must be long-term funding to generate and maintain an assortment of datasets at each step, which in turn need to be publicly available through open access repositories from various geographic locations. Red boxes highlight different reservoirs of wild and cultivated diversity within the Cartesian space, gray boxes are mixed datasets built around these collections, and connectors are methodological approaches.
Figure 1.
A roadmap of trans-disciplinary approaches aiming at harnessing genebank utilization for climate change research in the face of heat, and water scarcity. Compiling (a) previous characterizations and (b) geo-referencing-derived climate data/indices of available genetic resources in genebanks is a starting point to (c) assess the extent of abiotic stress tolerance among existing accessions, and the need of (d) new habitat-based population-guided collections targeting isolated pockets of cryptic diversity in dry and semi-arid regions. Planning question-oriented collecting trips of crop wild relatives and hidden landraces across contrasting environments/agro-ecologies is needed now more than ever, despite a century of gathering and preserving diversity in plants throughout genebanks. Coupling ex situ agro-ecological screenings together with (e) ongoing in situ genebanks characterizations for morphological and genetic variation is essential to define (c) putative tolerant reference collections, while understanding the (f) heritability (h2) of adaptive traits and their genetic architecture (i.e., underlying genes) via genome-wide selection scans (GWSS), genome–environment associations (GEA), and genome-wide association studies (GWAS). Since identifying these novel sources of heat and drought tolerance demands merging heterogeneous datasets, (g) machine learning (ML, in red letters) promises speeding up genebank characterization. The distinction that clustering (Table 1) and ML (Figure 2 and Table 2) strategies can provide between abiotic stress tolerant and susceptible accessions is essential to (h) transfer useful genetic variation from wild crop donors and early landraces into elite cultivated lines, either by designing (i) genomic-assisted breeding programs such as genomic prediction (GP) and inter-specific marker- and genomic-assisted backcrossing (MAB and GABC) schemes, or by envisioning (j) multi-trait gene editing strategies (e.g., CRISPR-Cas9). Once (k) abiotic stress tolerant varieties are validated across different environments, (l) legal inscription, seed multiplication, seed delivery system to farmers’ associations, and (m) follow-up given the regional needs, market demands, and adoption potential, are necessary downstream validation steps. These heterogeneous datasets are also likely to be inputted into ML, and in turn feedback new needs beyond heat and drought tolerance such as other types of resistances and nutritional quality. For ML to succeed speeding up the breeding of heat and drought-tolerant crops, there must be long-term funding to generate and maintain an assortment of datasets at each step, which in turn need to be publicly available through open access repositories from various geographic locations. Red boxes highlight different reservoirs of wild and cultivated diversity within the Cartesian space, gray boxes are mixed datasets built around these collections, and connectors are methodological approaches.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Cortés, A.J.; López-Hernández, F. Harnessing Crop Wild Diversity for Climate Change Adaptation. Genes 2021, 12, 783. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12050783
AMA Style
Cortés AJ, López-Hernández F. Harnessing Crop Wild Diversity for Climate Change Adaptation. Genes. 2021; 12(5):783. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12050783
Chicago/Turabian StyleCortés, Andrés J., and Felipe López-Hernández. 2021. "Harnessing Crop Wild Diversity for Climate Change Adaptation" Genes 12, no. 5: 783. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12050783
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.