
Administration of AAV-Alpha Synuclein NAC Antibody Improves Locomotor Behavior in Rats Overexpressing Alpha Synuclein

 ,
,
Abstract
:- Overexpression of αSyn in nigra DA neurons induces bradykinesia and neurodegeneration in rats.
- Injection of AAV-NAC32 produces a selective antibody against αSyn in nigra.
- AAV-NAC32 normalizes the behavior and improves the survival of nigra dopaminergic cells.
- AAV-NAC32 effectively antagonizes αSyn-mediated dopaminergic degeneration in nigra.
1. Introduction
2. Materials and Methods
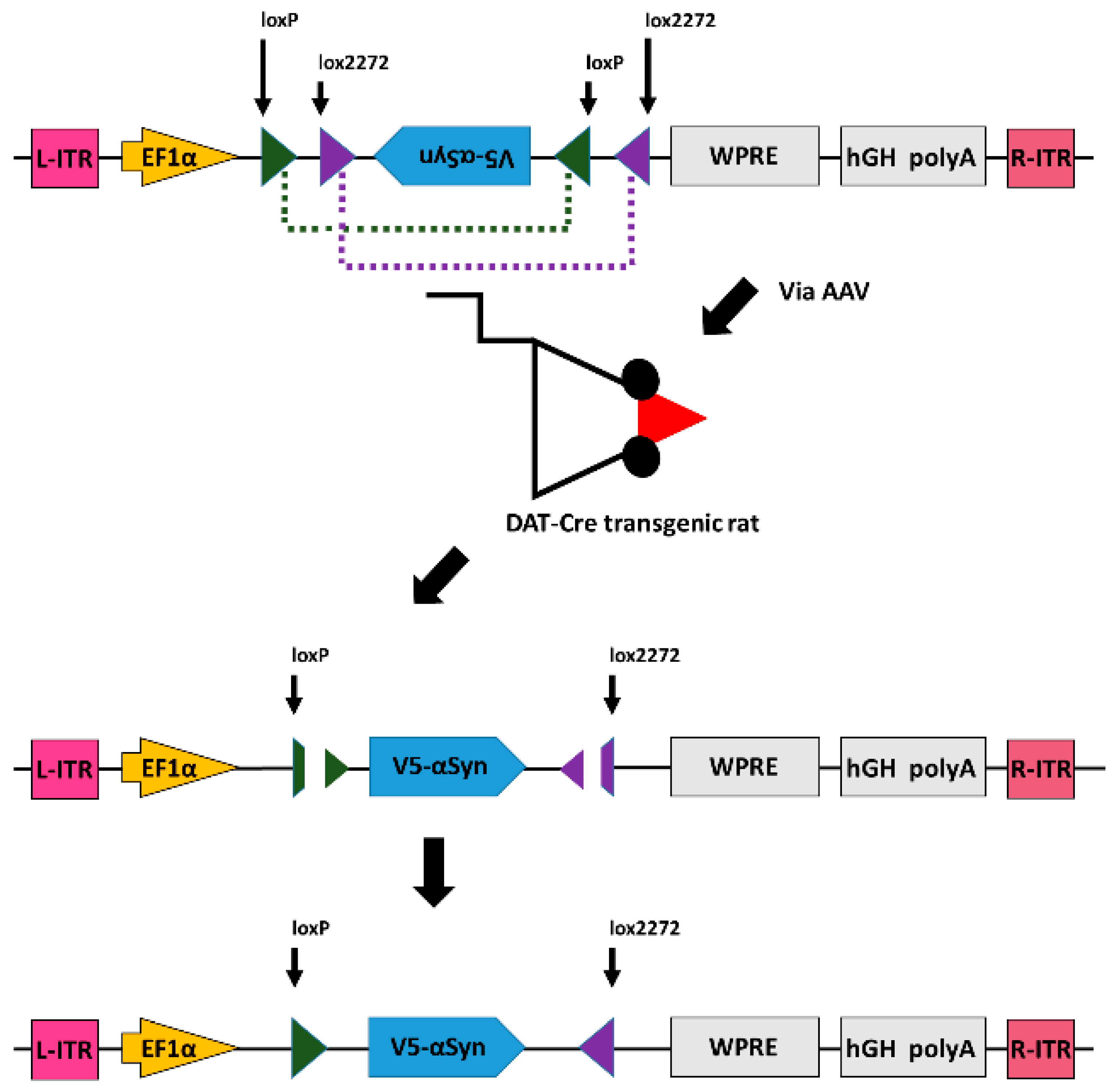
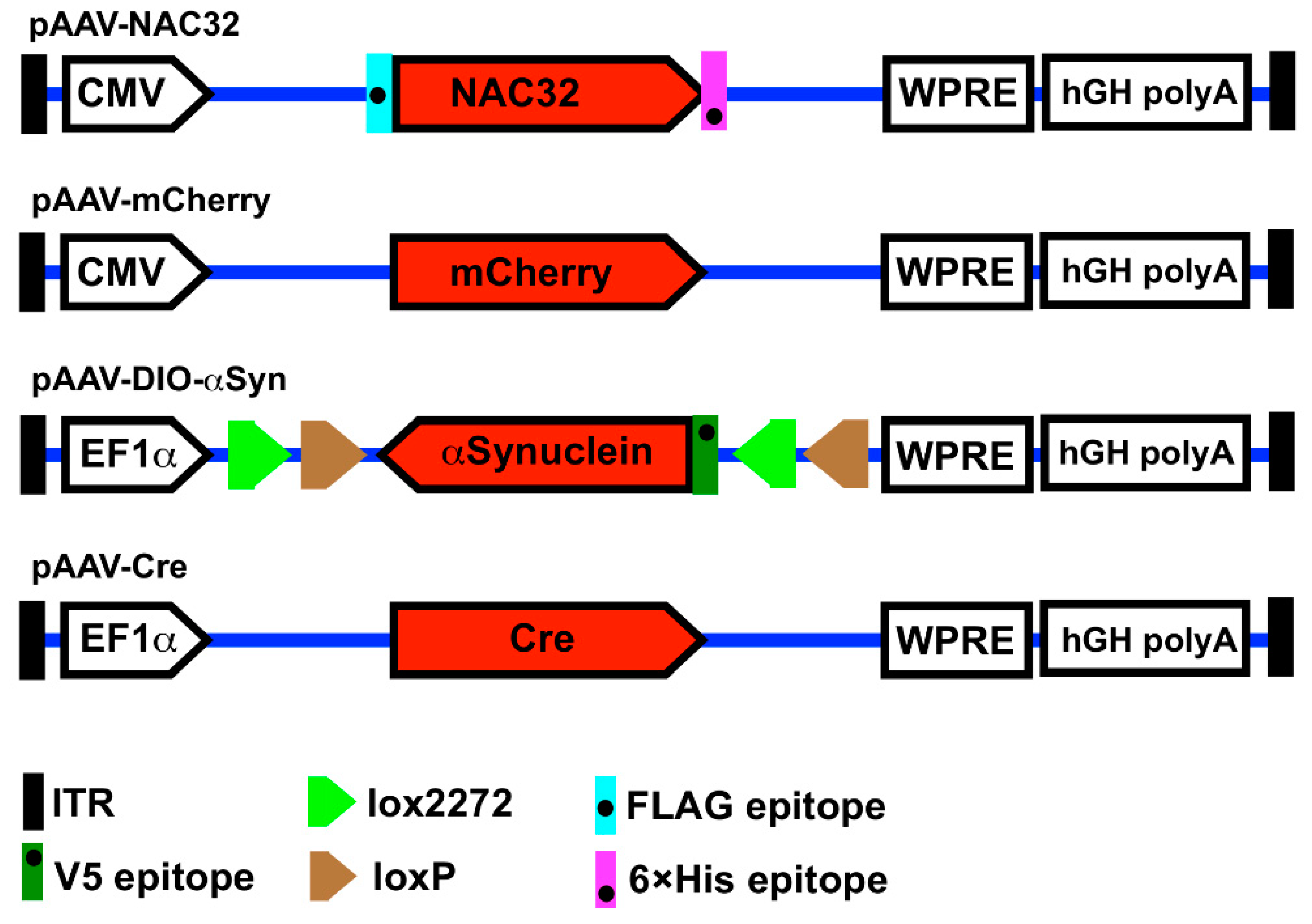
2.1. Plasmid Construction
2.2. Virus Production and Titration
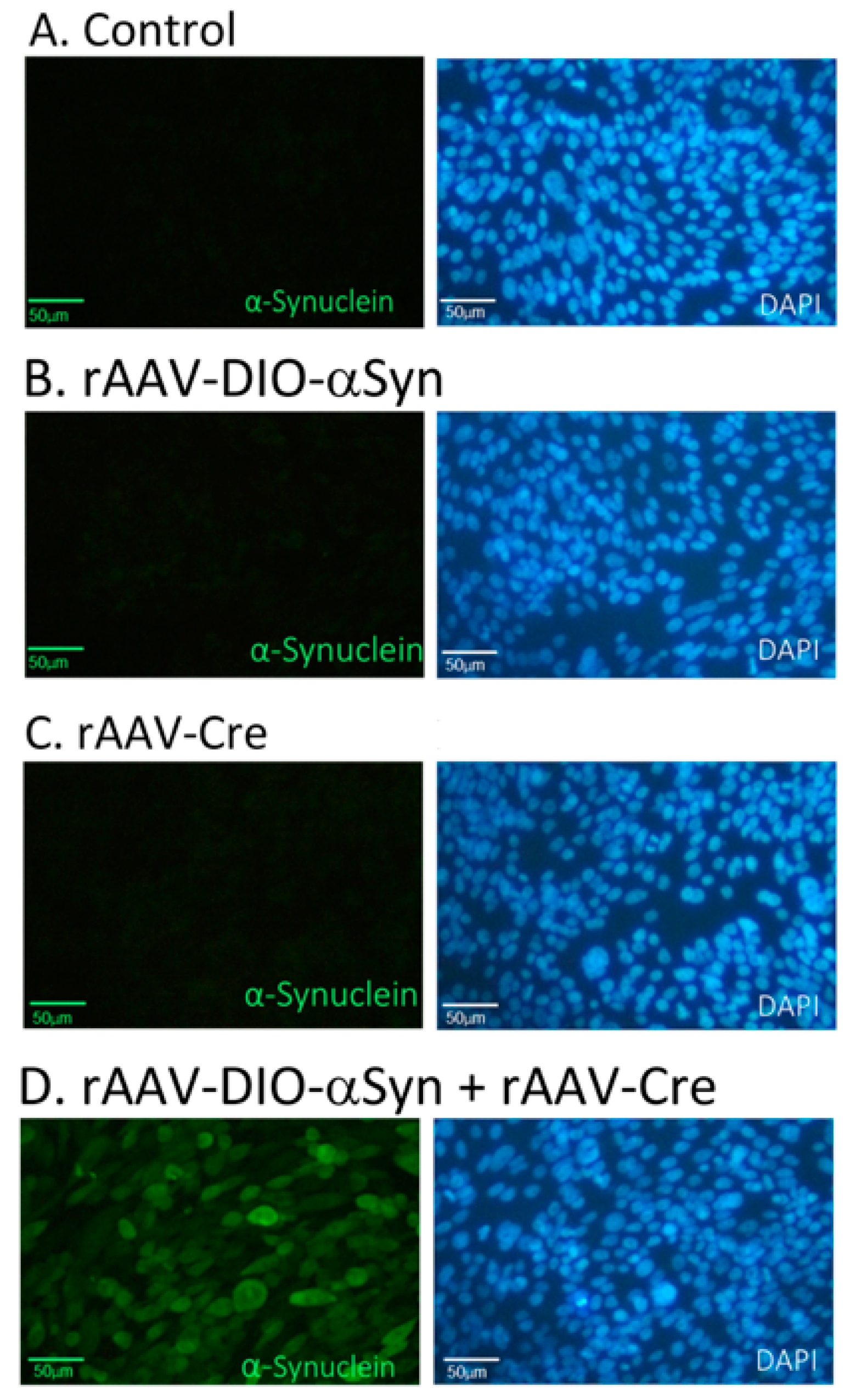
2.3. Cell Culture and Immunocytochemistry
2.4. Animals and Surgery
2.5. Locomotor Behavioral Measurement
2.6. Immunohistochemistry
2.7. Statistical Analysis
3. Results
3.1. Expression of α-Synuclein by Cre-DIO in Cultured CHO Cells
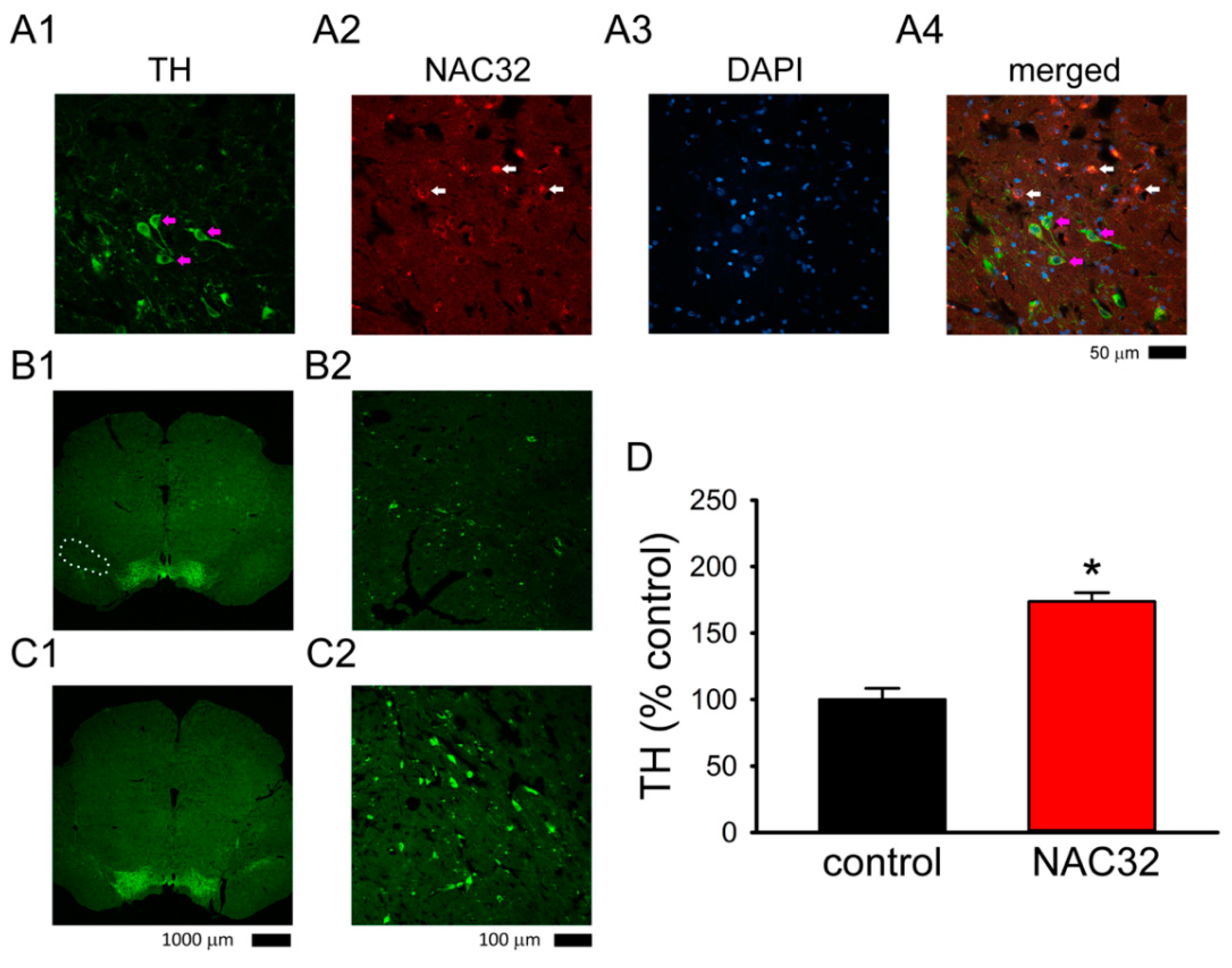
3.2. Expression of αSyn in Nigral Dopaminergic Neurons In Vivo
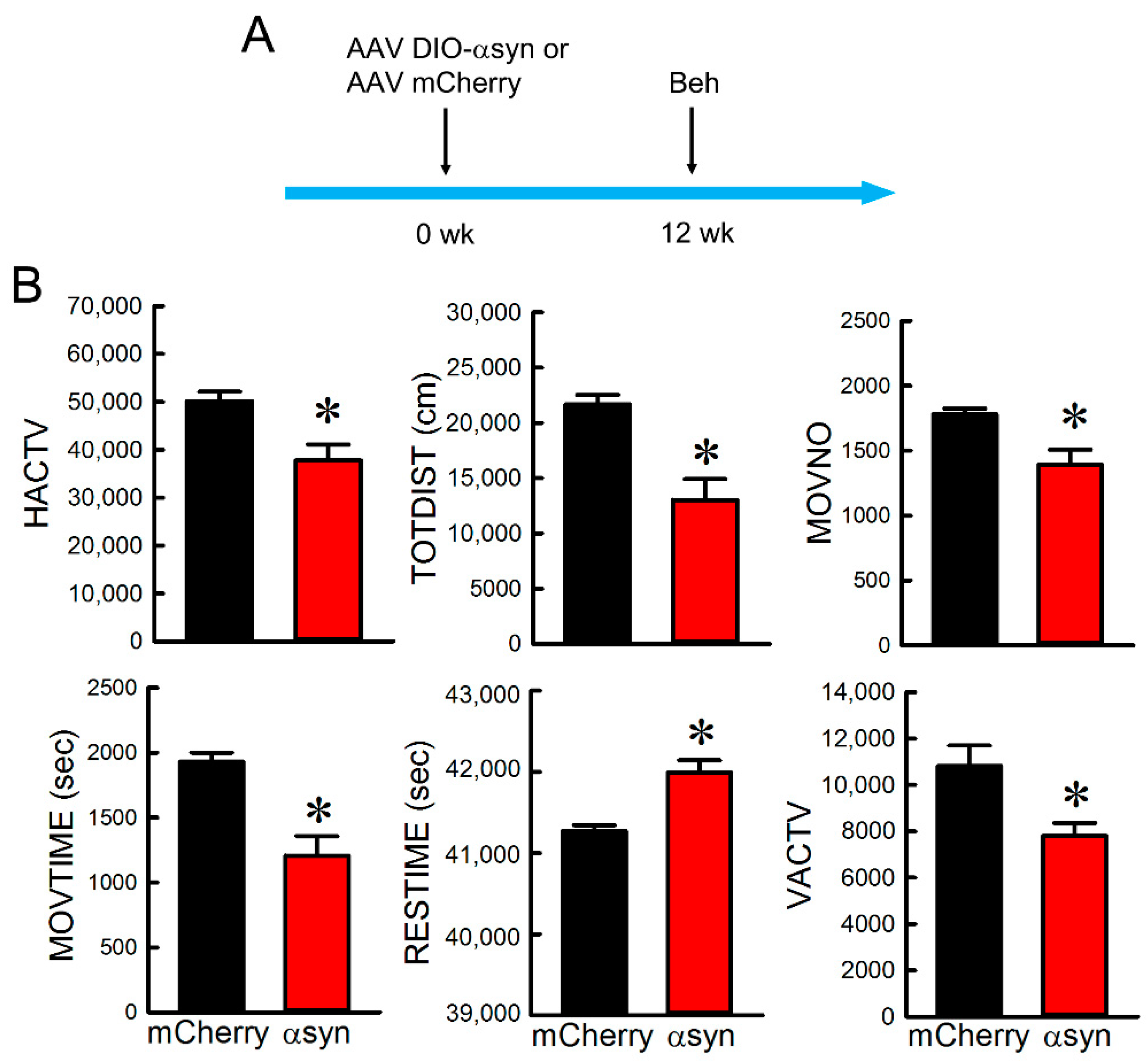
3.3. Overexpression of αSyn in Nigra Dopaminergic Neurons Induced Bradykinesia
3.4. Intranigral Injection of AAV-NAC32 Normalized Locomotor Behavior
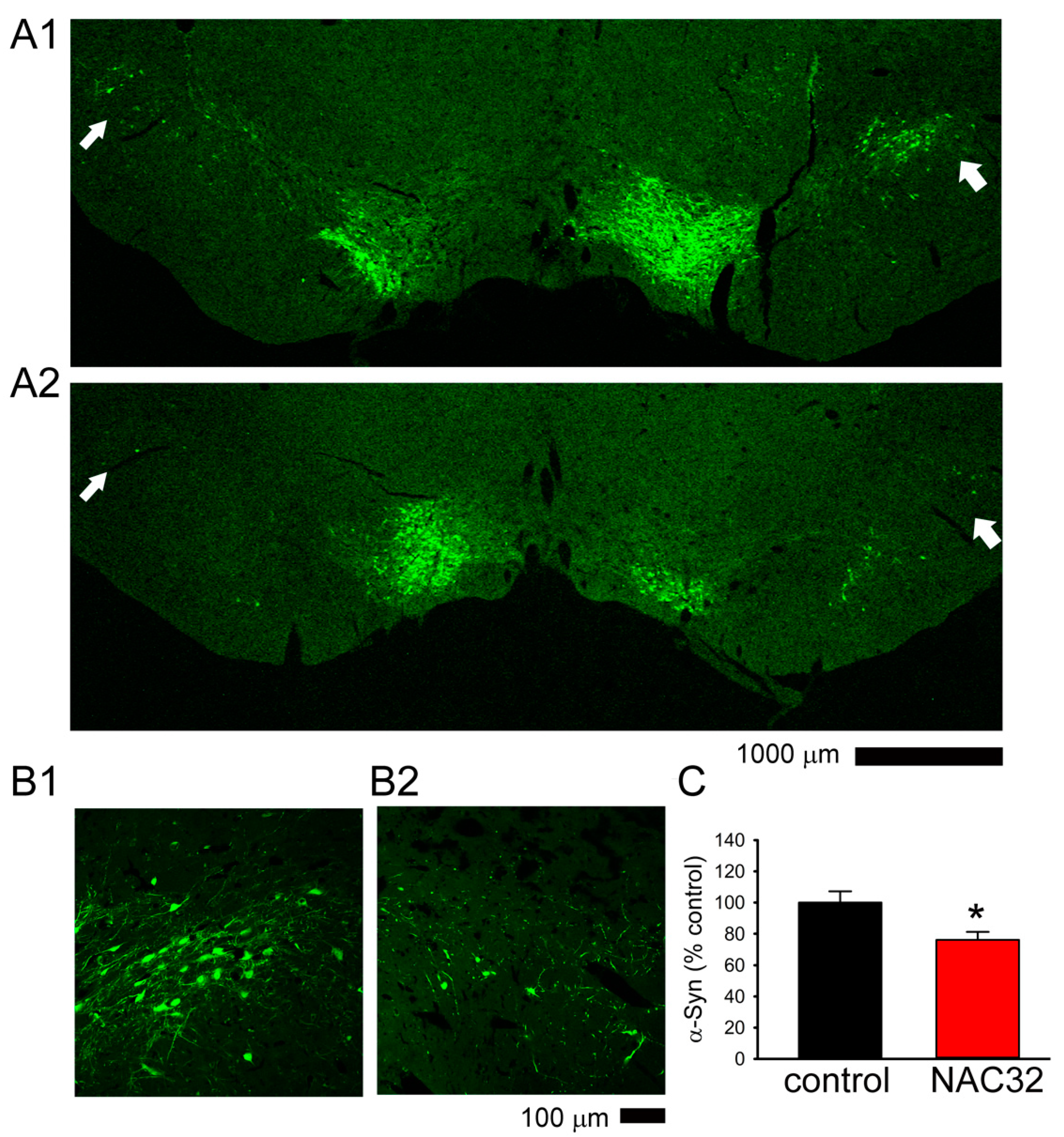
3.5. AAV-NAC32 Increased TH While It Reduced αSyn Immunoreactivity in SNcd
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| αSyn | α-Synuclein |
| AAV | adeno associated virus |
| DA | dopaminergic |
| DAT | dopamine transporter |
| DAT-Cre rats | rats expressing Cre-recombinase under dopamine transporter promotor |
| DIO | double floxed inverted open reading frame |
| DMEM | Dulbecco’s modified Eagle’s medium |
| HACTV | horizontal activity |
| MOVNO | number of movements |
| MOVTIME | movements time |
| NAC | nonamyloidal component |
| NK test | Newman Keuls test |
| PD | Parkinson’s disease |
| RESTIME | rest time |
| SNcd | Substantia nigra pars compacta dorsal tier |
| TH | tyrosine hydroxylase |
| TOTDIST | total distance traveled |
| VACTV | vertical activity |
References
- Kruger, R.; Kuhn, W.; Muller, T.; Woitalla, D.; Graeber, M.; Kosel, S.; Przuntek, H.; Epplen, J.T.; Schols, L.; Riess, O. Ala30Pro mutation in the gene encoding alpha-synuclein in Parkinson’s disease. Nat. Genet. 1998, 18, 106–108. [Google Scholar] [CrossRef]
- Masliah, E.; Rockenstein, E.; Veinbergs, I.; Mallory, M.; Hashimoto, M.; Takeda, A.; Sagara, Y.; Sisk, A.; Mucke, L. Dopaminergic loss and inclusion body formation in alpha-synuclein mice: Implications for neurodegenerative disorders. Science 2000, 287, 1265–1269. [Google Scholar] [CrossRef] [PubMed]
- Anderson, E.N.; Hirpa, D.; Zheng, K.H.; Banerjee, R.; Gunawardena, S. The Non-amyloidal Component Region of alpha-Synuclein Is Important for alpha-Synuclein Transport Within Axons. Front. Cell Neurosci. 2019, 13, 540. [Google Scholar] [CrossRef] [PubMed]
- Bisaglia, M.; Trolio, A.; Bellanda, M.; Bergantino, E.; Bubacco, L.; Mammi, S. Structure and topology of the non-amyloid-beta component fragment of human alpha-synuclein bound to micelles: Implications for the aggregation process. Protein Sci. 2006, 15, 1408–1416. [Google Scholar] [CrossRef] [Green Version]
- Oueslati, A.; Fournier, M.; Lashuel, H.A. Role of post-translational modifications in modulating the structure, function and toxicity of alpha-synuclein: Implications for Parkinson’s disease pathogenesis and therapies. Prog. Brain Res. 2010, 183, 115–145. [Google Scholar] [CrossRef] [PubMed]
- Allison, J.R.; Rivers, R.C.; Christodoulou, J.C.; Vendruscolo, M.; Dobson, C.M. A relationship between the transient structure in the monomeric state and the aggregation propensities of alpha-synuclein and beta-synuclein. Biochemistry 2014, 53, 7170–7183. [Google Scholar] [CrossRef]
- Gupta, V.; Salim, S.; Hmila, I.; Vaikath, N.N.; Sudhakaran, I.P.; Ghanem, S.S.; Majbour, N.K.; Abdulla, S.A.; Emara, M.M.; Abdesselem, H.B.; et al. Fibrillar form of alpha-synuclein-specific scFv antibody inhibits alpha-synuclein seeds induced aggregation and toxicity. Sci. Rep. 2020, 10, 8137. [Google Scholar] [CrossRef]
- Masliah, E.; Rockenstein, E.; Mante, M.; Crews, L.; Spencer, B.; Adame, A.; Patrick, C.; Trejo, M.; Ubhi, K.; Rohn, T.T.; et al. Passive immunization reduces behavioral and neuropathological deficits in an alpha-synuclein transgenic model of Lewy body disease. PLoS ONE 2011, 6, e19338. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.H.; Yu, S.J.; Wu, K.J.; Wang, Y.S.; Tsai, H.M.; Liao, L.W.; Chen, S.; Hsieh, W.; Chen, H.; Hsu, S.C.; et al. Downregulation of alpha-Synuclein Protein Levels by an Intracellular Single-Chain Antibody. J. Parkinsons Dis. 2020, 10, 573–590. [Google Scholar] [CrossRef]
- Lynch, S.M.; Zhou, C.; Messer, A. An scFv intrabody against the nonamyloid component of alpha-synuclein reduces intracellular aggregation and toxicity. J. Mol. Biol. 2008, 377, 136–147. [Google Scholar] [CrossRef] [Green Version]
- Schnutgen, F.; Doerflinger, N.; Calleja, C.; Wendling, O.; Chambon, P.; Ghyselinck, N.B. A directional strategy for monitoring Cre-mediated recombination at the cellular level in the mouse. Nat. Biotechnol. 2003, 21, 562–565. [Google Scholar] [CrossRef]
- Xu, J.; Zhu, Y. A rapid in vitro method to flip back the double-floxed inverted open reading frame in a plasmid. BMC Biotechnol. 2018, 18, 52. [Google Scholar] [CrossRef]
- Atasoy, D.; Aponte, Y.; Su, H.H.; Sternson, S.M. A FLEX switch targets Channelrhodopsin-2 to multiple cell types for imaging and long-range circuit mapping. J. Neurosci. Off. J. Soc. Neurosci. 2008, 28, 7025–7030. [Google Scholar] [CrossRef] [Green Version]
- Shimshek, D.R.; Kim, J.; Hübner, M.R.; Spergel, D.J.; Buchholz, F.; Casanova, E.; Stewart, A.F.; Seeburg, P.H.; Sprengel, R. Codon-improved Cre recombinase (iCre) expression in the mouse. Genesis 2002, 32, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.H.; Wu, K.J.; Wu, K.L.; Wu, K.L.; Tsai, H.M.; Chen, M.L.; Chen, Y.W.; Hsieh, W.; Lin, C.M.; Wang, Y. Recombinant Adeno-Associated Virus-Mediated Expression of Methamphetamine Antibody Attenuates Methamphetamine-Induced Hyperactivity in Mice. Sci. Rep. 2017, 7, 46301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohr, U.P.; Wulf, M.A.; Stahn, S.; Steidl, U.; Haas, R.; Kronenwett, R. Fast and reliable titration of recombinant adeno-associated virus type-2 using quantitative real-time PCR. J. Virol. Methods 2002, 106, 81–88. [Google Scholar] [CrossRef]
- Schapira, A.H. Pathogenesis of Parkinson’s disease. Bailliere’s Clin. Neurol. 1997, 6, 15–36. [Google Scholar]
- Spillantini, M.G.; Schmidt, M.L.; Lee, V.M.; Trojanowski, J.Q.; Jakes, R.; Goedert, M. Alpha-synuclein in Lewy bodies. Nature 1997, 388, 839–840. [Google Scholar] [CrossRef]
- Volpicelli-Daley, L.A.; Luk, K.C.; Patel, T.P.; Tanik, S.A.; Riddle, D.M.; Stieber, A.; Meaney, D.F.; Trojanowski, J.Q.; Lee, V.M. Exogenous α-synuclein fibrils induce Lewy body pathology leading to synaptic dysfunction and neuron death. Neuron 2011, 72, 57–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taguchi, K.; Watanabe, Y.; Tsujimura, A.; Tanaka, M. Expression of α-synuclein is regulated in a neuronal cell type-dependent manner. Anat. Sci. Int. 2019, 94, 11–22. [Google Scholar] [CrossRef] [Green Version]
- Mori, F.; Nishie, M.; Kakita, A.; Yoshimoto, M.; Takahashi, H.; Wakabayashi, K. Relationship among alpha-synuclein accumulation, dopamine synthesis, and neurodegeneration in Parkinson disease substantia nigra. J. Neuropathol. Exp. Neurol. 2006, 65, 808–815. [Google Scholar] [CrossRef] [Green Version]
- Papp, M.I.; Kahn, J.E.; Lantos, P.L. Glial cytoplasmic inclusions in the CNS of patients with multiple system atrophy (striatonigral degeneration, olivopontocerebellar atrophy and Shy-Drager syndrome). J. Neurol. Sci. 1989, 94, 79–100. [Google Scholar] [CrossRef]
- Inoue, M.; Yagishita, S.; Ryo, M.; Hasegawa, K.; Amano, N.; Matsushita, M. The distribution and dynamic density of oligodendroglial cytoplasmic inclusions (GCIs) in multiple system atrophy: A correlation between the density of GCIs and the degree of involvement of striatonigral and olivopontocerebellar systems. Acta Neuropathol. 1997, 93, 585–591. [Google Scholar] [CrossRef]
- Gilman, S.; Wenning, G.K.; Low, P.A.; Brooks, D.J.; Mathias, C.J.; Trojanowski, J.Q.; Wood, N.W.; Colosimo, C.; Dürr, A.; Fowler, C.J.; et al. Second consensus statement on the diagnosis of multiple system atrophy. Neurology 2008, 71, 670–676. [Google Scholar] [CrossRef] [PubMed]
- Jankovic, J.; Goodman, I.; Safirstein, B.; Marmon, T.K.; Schenk, D.B.; Koller, M.; Zago, W.; Ness, D.K.; Griffith, S.G.; Grundman, M.; et al. Safety and Tolerability of Multiple Ascending Doses of PRX002/RG7935, an Anti-alpha-Synuclein Monoclonal Antibody, in Patients With Parkinson Disease: A Randomized Clinical Trial. JAMA Neurol 2018, 75, 1206–1214. [Google Scholar] [CrossRef] [PubMed]
- Southern, J.A.; Young, D.F.; Heaney, F.; Baumgärtner, W.K.; Randall, R.E. Identification of an epitope on the P and V proteins of simian virus 5 that distinguishes between two isolates with different biological characteristics. J. Gen. Virol. 1991, 72 Pt 7, 1551–1557. [Google Scholar] [CrossRef]
- Dunn, C.; O’Dowd, A.; Randall, R.E. Fine mapping of the binding sites of monoclonal antibodies raised against the Pk tag. J. Immunol. Methods 1999, 224, 141–150. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| * Two-Way ANOVA | Posthoc NK Test | ||||
|---|---|---|---|---|---|
| p-Value | F Value | p at 4 wks | p at 8 wks | p at 12 wks | |
| HACTV | <0.001 | 16.415 | 0.163 | 0.032 | 0.004 |
| TOTDIST | 0.002 | 12.18 | 0.259 | 0.059 | 0.009 |
| MOVNO | <0.001 | 20.136 | 0.091 | 0.015 | 0.003 |
| MOVTIME | 0.001 | 14.088 | 0.254 | 0.038 | 0.005 |
| RESTIME | 0.001 | 14.09 | 0.254 | 0.038 | 0.005 |
| VACTV | 0.009 | 8.362 | 0.276 | 0.032 | 0.127 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.-H.; Wu, K.-J.; Hsieh, W.; Harvey, B.K.; Hoffer, B.J.; Wang, Y.; Yu, S.-J. Administration of AAV-Alpha Synuclein NAC Antibody Improves Locomotor Behavior in Rats Overexpressing Alpha Synuclein. Genes 2021, 12, 948. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12060948
Chen Y-H, Wu K-J, Hsieh W, Harvey BK, Hoffer BJ, Wang Y, Yu S-J. Administration of AAV-Alpha Synuclein NAC Antibody Improves Locomotor Behavior in Rats Overexpressing Alpha Synuclein. Genes. 2021; 12(6):948. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12060948
Chicago/Turabian StyleChen, Yun-Hsiang, Kuo-Jen Wu, Wei Hsieh, Brandon K. Harvey, Barry J. Hoffer, Yun Wang, and Seong-Jin Yu. 2021. "Administration of AAV-Alpha Synuclein NAC Antibody Improves Locomotor Behavior in Rats Overexpressing Alpha Synuclein" Genes 12, no. 6: 948. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12060948