
MiRNAs and Cancer: Key Link in Diagnosis and Therapy
by
Yu Shi
1,2,†, 
Zihao Liu
1,2,†,
Qun Lin
1,2,
Qing Luo
1,2,
Yinghuan Cen
1,2,
Juanmei Li
1,2,
Xiaolin Fang
1,2 and
Chang Gong
1,2,* 1
Breast Tumor Center, Sun Yat-Sen Memorial Hospital, Sun Yat-Sen University, Guangzhou 510000, China
2
Guangdong Provincial Key Laboratory of Malignant Tumor Epigenetics and Gene Regulation, Sun Yat-Sen Memorial Hospital, Sun Yat-Sen University, Guangzhou 510120, China
*
Author to whom correspondence should be addressed.
†
These authors contribute equally to this work.
Genes 2021, 12(8), 1289; https://0-doi-org.brum.beds.ac.uk/10.3390/genes12081289
Submission received: 18 July 2021
/
Revised: 18 August 2021
/
Accepted: 20 August 2021
/
Published: 23 August 2021
(This article belongs to the Special Issue The Role of MicroRNA in Cancer)
Abstract
:Since the discovery of the first microRNA (miRNA), the exploration of miRNA biology has come to a new era in recent decades. Monumental studies have proven that miRNAs can be dysregulated in different types of cancers and the roles of miRNAs turn out to function to either tumor promoters or tumor suppressors. The interplay between miRNAs and the development of cancers has grabbed attention of miRNAs as novel tools and targets for therapeutic attempts. Moreover, the development of miRNA delivery system accelerates miRNA preclinical implications. In this review, we depict recent advances of miRNAs in cancer and discuss the potential diagnostic or therapeutic approaches of miRNAs.
1. Introduction
Hippocrates naming the abnormal mass of tissue in stomach and uterus as “cancer” revealed the long-lasting exploration of human malignant tumors [1]. The International Agency for Research on Cancer (IARC) has published the Latest Global Cancer Data, which indicates that in 2020, nearly 19,300,000 people were diagnosed with cancer worldwide. Breast cancer has the highest incidence among 36 types of cancer, while the mortality of lung cancer remains the highest. Due to its forementioned characteristics, cancer has remained the top leading cause of death worldwide for nearly a decade [2]. Still, efforts have been made to protect humans from cancer. The development of diagnostic and therapeutic strategies has come to a new era. The current clinical diagnostic and therapeutic approaches are diverse, and new genomic diagnostic tools as well as therapeutic systems are under development [3,4].
Since the discovery of the first microRNAs (miRNAs) on Caenorhabditis elegans in 1993, the understanding of miRNA biology has grown deeper. The action of mechanisms of miRNAs, how they participate in the regulation of genes in prokaryotes and eukaryotes, have been revealed [5,6]. miRNAs are capable of binding to the 3′UTR of corresponding mRNA, which ultimately leads to the post-transcriptional suppression or degradation or silencing of target genes [7]. It has been revealed that approximately 30% of human genes can be regulated by miRNAs [8]. Moreover, a single miRNA can interact with a dozen of mRNAs, which ultimately influence the expression of many genes simultaneously [9]. The complex regulation function of miRNAs is involved in multiple biological process, including cell proliferation, differentiation, invasion, migration, and apoptosis [10]. Meanwhile, these dysregulated biological processes are bound up with sets of diseases.
As a result, miRNAs relate to miscellaneous human disease including tumors [11,12,13]. The very first discovery of a cancer associated with miRNA was published in 2002 [14]. Since then, growing studies have demonstrated the abnormal expression of miRNAs in several types of tumors [15,16,17,18]. In this review, we mainly focus on the relationship between miRNAs and cancer biogenetic process as well as miRNA-based diagnostic and therapeutic strategies.
2. miRNA Biology
miRNAs are small endogenous RNAs, 19–25 nucleotides in length. In 1993, the very first miRNA lin-4 was found in Caenorhabditis elegans, and it potentially suppresses the expression of lin-14, lin-28, lin-41, lin-42, and daf-12 [19,20,21]. Several decades later, nearly 3000 miRNAs have been found in the human genome. miRNAs have been proven to take part in the normal cell growth as well as the development and progression of diseases. As the fundamental element of RNA-induced silencing complex (RISC), miRNAs modulate target gene expression by the degradation of targets or inhibition of target gene translation [22,23,24].
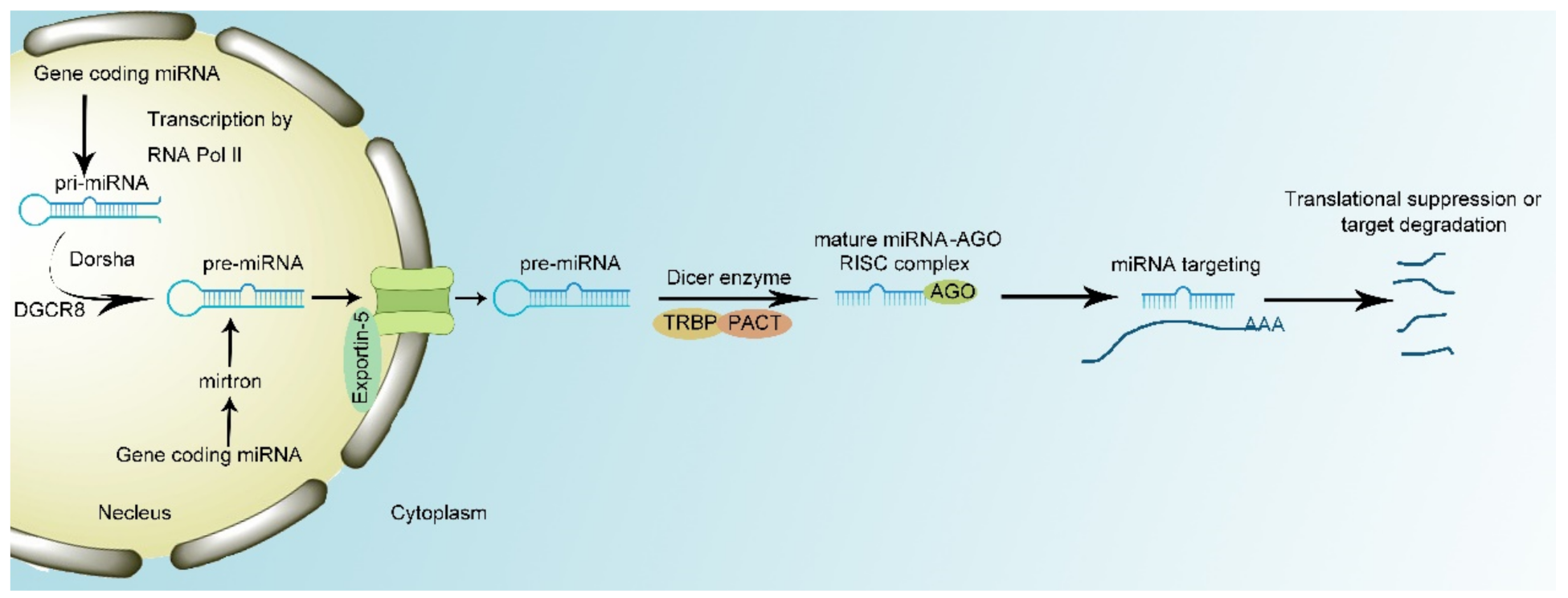
Generally, miRNAs are biosynthesized by two pathways. In the canonical biosynthesis pathway, primary miRNAs (pri-miRNAs) are transcribed from the introns by RNA polymerase II (RNA Pol II). Then, the microprocessor complex, composed of Drosha enzyme and DiGeorge syndrome critical region 8 (DGCR8), interacts with pri-miRNA to cut the specific stem-loop structure to 60–70 nt in size and processes pri-miRNAs into precursor miRNAs (pre-miRNAs) [25]. The transporter protein Exportin-5 transports pre-miRNAs to the cytoplasm. However, knockout of Exportin-5 cannot completely block the transportation of pre-miRNA. This indicates there are other transporting mechanisms to translocate pre-miRNAs into cytoplasm [26,27]. At last, the Dicer enzyme, composed of TAR RNA-binding protein (TRBP) and protein activator of the interferon-induced protein kinase (PACT), can splice pre-miRNA into single-strand mature miRNA and load mature miRNA with AGO protein into RISC [25,28]. In the noncanonical pathway, miRNA biosynthesis is independent of the microprocessor. Mirtrons directly derive from introns and ignore the interplay with the microprocessor complex, while the remaining procedures remain the same [29].
Once biosynthesized, miRNAs complementarily base pair with corresponding mRNA, a process that is initiated by conformational change of the MID and PIWI domains in AGO proteins [30]. miRNA interacts with 3′UTR of target mRNA to manipulate its expression. The seed region of miRNA which is composed by the 2nd–7th of the nucleotides of miRNA sequence conducts the interaction between miRNA and mRNA. mRNAs with an adenine opposite miRNA nucleotide 1 (t1A) ahead of seed-region-target sequences have the maximum binding capacity. Moreover, the 13th–16th nucleotides of 3′UTR of miRNA serve as supplemental pairing region to mediate additional interaction between miRNAs and mRNAs [31,32]. miRNAs can lead to either the degradation or translation repression of target mRNA at specific site. Moreover, RISC complex can recruit polyA-nuclease deadenylation complex subunit 2/3 (PAN2/3), carbon catabolite repressor protein 4 (CCR4), and NOT complex and the recruitment of these proteins result in the degradation of mRNAs conducted by 5′-3′ exonuclease 1 [33]. In addition, miRNAs interact with eukaryotic initiation factor 4 A- I/II in the translation–initiation complex to tamper the translation of target mRNA without influencing mRNA stability [34,35]. However, recent research has proven that miRNAs can act as nuclear activating miRNAs to modulate transcription. For example, when miR-24-1 is transfected into cells, the surrounding genes’ transcription is shown to be enhanced (Figure 1) [36].
Moreover, the regulatory function of miRNAs can be influenced by competing endogenous RNAs (ceRNAs), such as circular RNAs (circRNAs), pseudogene transcripts, and long non-coding RNAs (lncRNAs). Accordingly, mounting evidence has indicated that ceRNAs play a critical role in the functional regulation of miRNAs [37,38,39]. Once bound with miRNAs, ceRNAs occupy with individual miRNA and inhibit interaction between miRNAs and target mRNA, ultimately blocking miRNA function [40].
3. miRNAs Regulate the Hallmarks of Cancers
Recent studies have expanded our understanding of the interplay among tumor initiation, progression, recurrence, and the biogenetic markers of angiogenesis, therapy-resistance, invasion, as well as metastasis activation [41]. When compared with normal tissue, miRNAs are often dysregulated in tumors. The abnormally expressed miRNAs have been proven to manipulate stemness, angiogenesis, proliferation, apoptosis process, cell cycle, and the epithelial–mesenchymal transition (EMT) of tumor cells. miRNAs regulate individual target genes directly. Moreover, one miRNA can regulate several target genes, which means a single miRNA modulates a brunch of biological functions.
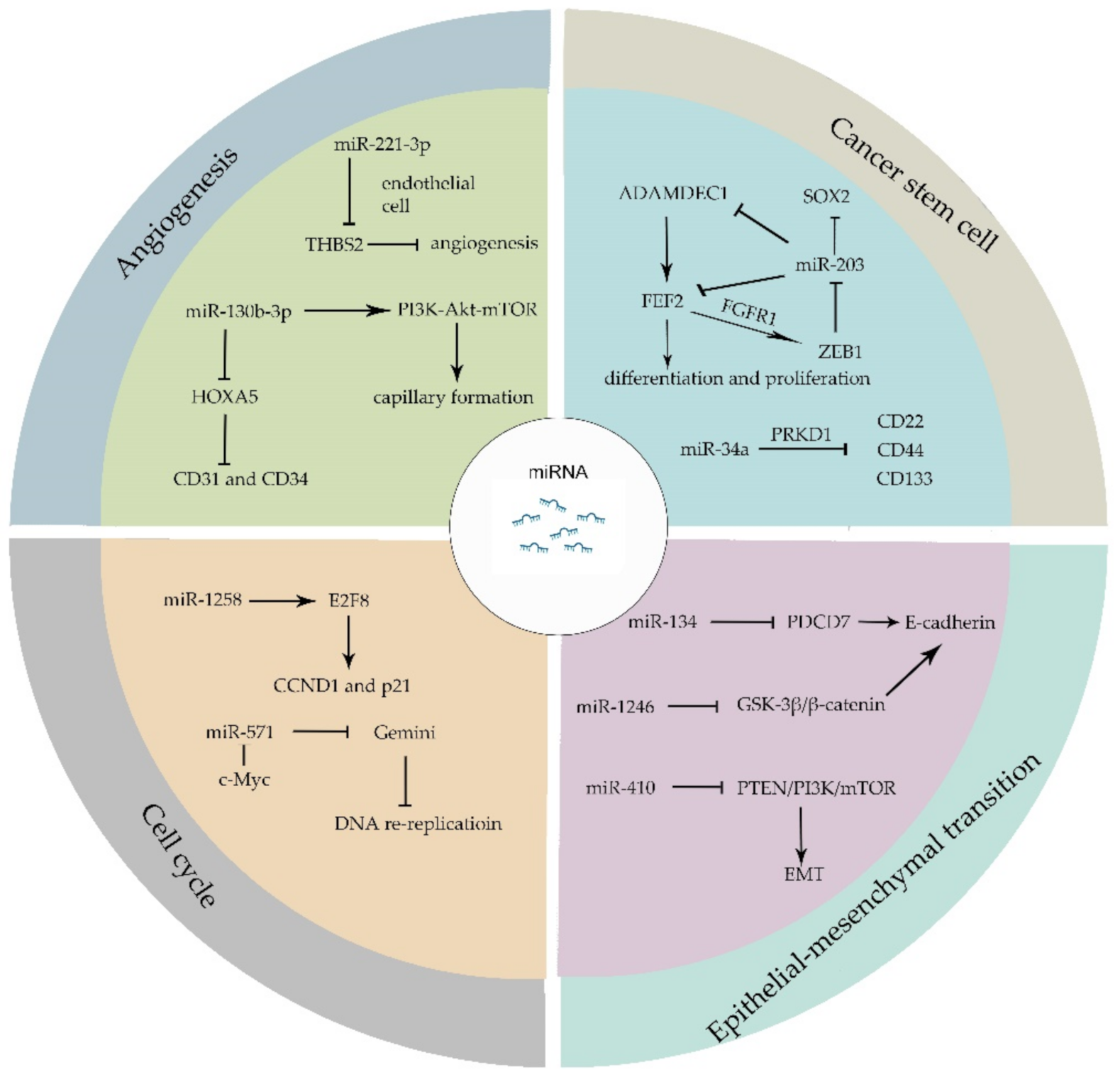
Cancer stem cells (CSCs) share similar characteristics with stem cell and are capable of escaping from conventional cancer therapies. Fibroblast growth factor 2 (FEF2), as one member of the FGF family, responds to injury and tissue repair in normal neural development [42]. Recent studies have proven that FEF2 modulates the differentiation and proliferation of stem cells [43,44]. The existence of glioblastomas cancer stem cells (GSCs) largely contributes to the poor clinical outcome of patients. GSCs secret a disintegrin and metalloproteinase domain-like protein decysin 1 (ADAMDEC1), which in turn accelerate the release of FGF2. FGF2 binds to FGF receptor 1 (FGFR1) and activates Zinc-finger E-box-binding homeobox 1 (ZEB1) which exerts its transcriptional modification through the downregulation of miR-203. The overexpression of miR-203 decreases the expression of ADAMDEC1 and reduces the expression of stem-cell associated transcription factor SOX2 [45,46]. miR-34a binds to serine/threonine-protein kinase D1 (PRKD1) and suppresses the expression of stem cell marker, such as CD24, CD44 and CD133 [47]. Other miRNAs like miR-122 [48], miR-185-3p [49], miR-200c [50], etc., have been reported to mediate cancer stem cell biological processes.
Epithelial–mesenchymal transition (EMT) is a process whereby epithelial-like tumor cells acquire mesenchymal features and tend to invade or migrate. During EMT, several epithelial markers are involved, such as E-cadherin, ZO-1 and mesenchymal markers such as N-cadherin, vimentin, and fibronectin. Signaling pathways including transforming growth factor (TGF), fibroblast growth factor, Wnt/β-catenin, Notch, and human epidermal growth factor receptor have been illustrated to regulate EMT [51]. Programmed Cell Death 7 (PDCD7) can be targeted by miR-134 and it modulates E-cadherin expression through binding to the promotor of E-cadherin. The upregulation of miR-134 reduces PDCD7 and attenuates PDCD7-mediated tumor suppression [52]. Moreover, miR-1246 directly targets GSK-3β/β-catenin pathway which downregulates E-cadherin expression and promotes EMT [53]. Furthermore, miRNAs such as miR-128-3p [54], miR-190 [55], miR-612 [56], and miR-33a-5p [57] have been proven to be involved in the EMT process. The EMT process not only leads to the invasion and migration of tumor cells, but also results in therapy resistance in cancer patients [58]. The overexpression of miR-410 enhances EMT and radiotherapy resistance by increasing DNA damage repair and regulates EMT through PTEN/PI3K/mTOR axis which leads to the radiotherapy resistance in non-small cell lung cancer (NSCLC) [59].
The cell cycle is a complex regulatory process that cells go through in order to exponentially proliferate. It precisely modulates the replication of whole genome and division of genomic replicates into new daughter-cells, which generally contains four stages: Gap1 (G1), Synthesis (S), Gap2 (G2), and Mitosis (M) [60,61,62]. In cancer cells, the continuous proceeding of G1-S-G2-M cycle leads to the ceaseless proliferation of tumors. The abnormal cell-cycle related proteins, such as pRB, p53, cyclin-dependent kinases (CDKs), cyclin-dependent kinase inhibitor (CDKI), and cyclins contribute to cancer initiation and progression. Cancer cells can manipulate cycle-regulatory proteins through miRNAs machinery to thrive and proliferate [61]. A low expression of miR-1258 was observed in colorectal cancer, while overexpression of miR-1258 restrains tumor proliferation. It has been shown that a high expression of miR-1258 arrests cell cycle at G0/G1 phase because miR-1258 binds to the 3′UTR of E2F8 which is a regulator of cyclin D1 (CCND1) and cyclin dependent kinase inhibitor 1A (p21) [63]. Gemini, as a critical regulator of DNA replication, inhibits DNA re-replication during S, G2, and early M phases. miR-571 could downregulate Gemini level independent of anaphase promoting complex (APC/C) in late S phase and downregulation of Gemini leads to DNA re-replication and genomic instability. Moreover, CDK2-dependent c-Myc directly interacts with miR-571 promoter and restrains miR-571 expression [64].
Angiogenesis, as a vital process in normal cells growth and development, plays an extremely important role in tumor metastasis and survival [65,66]. Recent research works have revealed that miRNAs enroll in the regulation of angiogenesis [67,68]. The expression of miR-221-3p is related to the microvascular density and leads to tumor progression. miR-221-3p-enriched exosomes secreted by tumor cells are transferred to endothelial cells. In endothelial cells, miR-221-3p binds to the mRNA of Thrombospondin-2 (THBS2) which is regarded as angiogenesis inhibitor and inhibits THBS2 expression [69]. The upregulation of THBS2 can reverse the angiogenic effect of miR-221-3p. HOXA5, which has been proven to associate with angiogenesis biomarker CD31 and CD34, is a direct target of miR-130b-3p. The overexpression of miR-130b-3p leads to the downregulation of HOXA5 and activates PI3K-Akt-mTOR signaling pathway to provoke capillary formation (Figure 2) [70,71].
4. Therapeutic Approaches Targeting miRNA in Cancer
A variety of carcinogenesis-associated miRNAs are abnormally expressed during tumors initiation and progression [72,73]. This raises the possibility that manipulating miRNA expression might be promising for cancer treatment [9,74,75,76,77].
Based on the existing research, attempts to modulate the expression of miRNAs for tumors treatment are in full swing [78,79,80]. A set of exogenous miRNAs which are divided into miRNA mimics and miRNA inhibitors (antimiRs) have been developed for preclinical use. When a tumor suppressor miRNA is diminished in tumors, miRNA mimics can be introduced into tumor cells to replenish the miRNA and abolish tumor progression. miRNA mimics are often transfected to reverse specific tumor-suppressor miRNA in tumor cells. For example, in gastric and colorectal cancer cells, the overexpression of miR-451 can reduce cancer cell proliferation and increase radiotherapy sensitivity [78]. Approaches to antagonize the effects of miRNA include antisense oligonucleotides (ASOs), miRNA antagonists (antagomirs), and locked nucleic acid (LNA) -modified oligonucleotides are also well studied. ASOs are phosphorothioate nucleotides with RNase H activity and it can complement with miRNAs to suppress miRNA function, while antagomirs is a chemically modified single strand oligonucleotides that binds miRNAs to abrogate function of miRNAs [7,12,13,81]. In hepatitis virus infected mice models, an injection of antimiR-122 can significantly downregulate miR-122 level in plasma, liver, and white adipose tissue, which ultimately inhibits hepatitis C virus (HCV) replication [82,83,84,85]. Moreover, LNA-modified antimiRs have remarkable therapeutic response in mouse models [86,87].
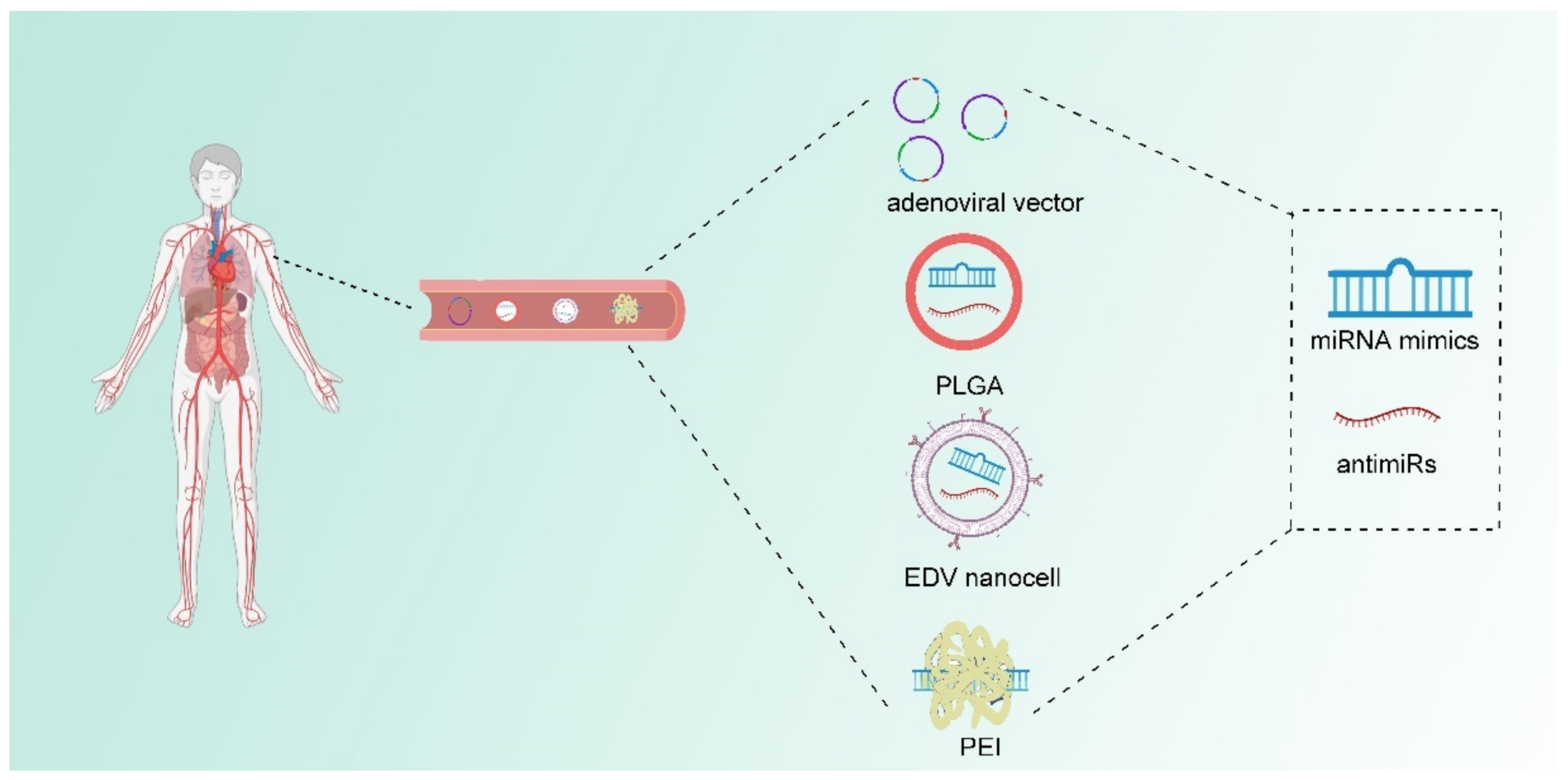
In addition to the chemical modulation of miRNAs, delivery systems that can wrap naked nucleotides can also enhance the efficiency of exogenous miRNA because of the unstable state of naked nucleotides. Huge progress has been made to enhance in vivo delivery. Delivery systems such as adenoviral vectors, Poly (lactide-co-glycolide) (PLGA), EnGeneIC Delivery Vehicle (EDV) nanocells, and polyethylenimine (PEI) have been developed [12]. Adenoviral vectors are capable of encoding miRNA molecules, but the safety of their clinical application remains challenging [88,89]. PLGA which is currently used as biodegradable sutures show advantage for miRNA delivery in vivo. PLGA has low toxicity and can deliver RNA molecules [90,91]. EDV nanocells are 400nm particles coated with lipopolysaccharide and surface-conjugated antibodies [92,93]. A clinical trial [NCT02369198] already uses EDV nanocells to deliver miR-16 mimics in vivo [92,94]. PEI, which can form a complex with exogenous miRNA, can adhere to a negatively charged cell membrane. Studies have indicated that PEI as well as the poly (l-lysine)-modified polyethylenimine (PEI-PLL) have profound delivery efficiency in preclinical models [95,96]. Some of aforementioned miRNA delivery strategies have already implicated in clinical trials and further developments can be expected (Figure 3).
miRNA-based clinical therapy has made huge progress. Miravirsen, as one of the first miRNA-based molecules for HCV treatment clinical trial, is composed of LNA-modified miR-122 complementary sequence. Miravirsen blocks the interaction between miR-122 and HCV RNA. Moreover, Miravirsen targets pri-miR-122 as well as pre-miR-122 and it can also inhibit miR-122 biosynthesis [97]. The success of Miravirsen inspired the development of miRNA-based cancer therapies. miR-142-3p upregulates in breast cancer and leads to hyperproliferation of tumor through Wnt signaling pathway. Exosomes from bone marrow-derived mesenchymal stem cells (MSCs-Exo) were manipulated to deliver anti-miR-142-3p to tumor in vivo which efficiently suppresses cancer stem-like cells proliferation [98,99]. MRX34 is a liposomal-wrapped miR-34a mimic which is under phase I study. miR-34a interacts with programmed cell death ligand 1 (PD-L1) 3′ UTR to suppress PD-L1 expression and it is regarded as a tumor-suppressive miRNA. It has been proven that the overexpression of miR-34a can suppress a set of tumors. MRX34 might be promising for antitumor treatment. It has been validated that delivery of MRX34 in NSCLC mouse model suppresses PD-L1 level by 30%. This leads to the upregulation of CD8+T cell infiltrating rate by 38.6% and the prolonged survival of xenograft mice [100,101,102,103]. Besides, the antitumor activity of MRX34 has been tested in clinical trials which enrolled primary liver cancer, small cell lung cancer (SCLC), lymphoma, melanoma, multiple myeloma, renal cell carcinoma, and non-small cell lung cancer (NSCLC) patients [NCT01820071]. miR-16 mimics based-EDV nanocells have entered phase I trials which enrolls patients with malignant pleural mesothelioma and NSCLC [NCT02369198]. miR-155 antimiR MRG106 (Cobomarsen) have been applied in a phase I clinical trial in patients of lymphoma and leukemia [NCT02580552].
Manipulation of miRNAs can also enhance therapy sensitivity in patients. The relatively high expression of miR-621 was found to associate with better paclitaxel plus carboplatin (PTX/CBP) therapeutic effect in breast cancer patients. miR-621 suppresses FBXO11 to promote p53 activity, which consequently increases apoptosis in cancer cells. miR-621 may act as a potential therapeutic target for breast cancer treatment in the future (Table 1) [97].
However, some specific miRNAs have opposite functions in various cancers. For example, miR-106b-5p is upregulated in several cancers and proved to be oncogenic, including gastric cancer, hepatocellular carcinoma, and glioma. Vice versa, it acts as a tumor suppressor in papillary thyroid cancer, breast cancer and epithelial ovarian cancer [104,105]. As aforementioned, these specific miRNAs may likely to have adverse therapeutic effects among different cancers. The precise use of miRNA mimics or antimiRs in different cancers remains to be explored in the future.

{kind=link}
{kind=link}
{kind=link}
Table 1.
Selected miRNAs in cancers and their therapeutic manipulations.
| miRNAs | Disease | Expression of miRNA | Important mRNA Target | Therapeutic Model | Reference |
|---|---|---|---|---|---|
| miR-122 | HCV infection | upregulated | HCV 5′UTR | Miravirsen | [85] |
| miR-142-3p | breast cancer | upregulated | miR-150, APC, and P2X7R | anti-miR-142-3p | [99] |
| miR-34a | liver cancer, lung cancer, lymphoma, melanoma, renal cell carcinoma | downregulated | PD-L1 3′UTR | MRX34 | [100] |
| miR-16 | Mesothelioma, non-small cell lung cancer | downregulated | BCL-2, CDK1, ETS1 and JUN | miR-16 mimic | [106] |
| miR-155 | lymphoma and leukemia | upregulated | SHIP1, WEE1, VHL, TP53INP1 | AntimiR-155 | [107,108,109] |
| miR-621 | breast cancer | downregulated | FBXO11 | N.A. | [97] |
5. Diagnostic Potential of miRNA in Cancer
Since miRNAs are abnormally expressed by tumors, attempts to analyze miRNA expression profiles and detect the relative expression of miRNAs in plasma of tumor patients may predict cancer prognosis in a diagnostic manner. With the development of biotechnology, the sensitivity of detection technology has improved, and the types of detectable samples have expanded from fresh frozen tissue, paraffin tissue fixed with formalin to miRNA in body fluid, which greatly facilitates miRNA clinical applications [110,111,112]. miR-192-5p, miR-194-5p, and miR-215-5p are upregulated in Barret’s esophagus (BE). In a small cohort of 67 BE participates, researchers found that the area under the curve (AUC) to diagnose BE by combining of miR-192-5p, miR-194-5p and miR-215-5p could reach 0.96–0.97 [113]. Moreover, in a prospective study, miRNA expression in bronchial epithelium from 347 smokers were analyzed via small-RNA sequencing. The results indicate that miR-146a-5p, miR-324-5p, miR-223-3p, and miR-223-5p were downregulated in lung cancer patients. In another lung cancer cohort, the combination of miR-146a-5p and existing mRNA biomarkers profoundly improved the prediction accuracy of lung cancer [114].
Circulating miRNAs have proven to bind with argonaut protein or high-density lipoprotein (HDL), which in turn protects circulating miRNAs from degradation by extreme pH or abnormal temperature as well as RNase function. Since circulating miRNAs can be easily detected, circulating miRNAs have become potential diagnostic biomarkers in cancer [7,74]. In a cohort of epithelial ovarian cancer (EOC) patients, ovarian cystadenoma patients, and healthy women, a high level of miR-200b can predict the poor overall survival of patients [80]. Osteosarcoma which shows poor prognosis is susceptible in children and teenagers [115]. In a large training and validation cohort, 29 miRNAs have been identified abnormally expressed in osteosarcoma. Among these abnormally expressed miRNAs, miR-221 and miR-222 have been found to associate with metastasis risk of osteosarcoma and turn out to be promising prognostic markers [116]. Currently, the diagnostic biomarkers of hepatocellular carcinoma (HCC) mainly count on α-fetoprotein (AFP), Lens culinaris agglutinin-reactive AFP (AFP-L3) and des-γ-carboxyprothrombin (DCP). However, a recent study discovered serum miR-16 as a novel marker for HCC. miR-16 itself has a sensitivity of 72.1% and specificity of 88.8% for the detection of HCC. Moreover, when combined with AFP, AFP-L3, and DCP, the sensitivity to diagnose HCC increased to 92.4% [117]. Meanwhile, urinary miRNAs, including miR-618 and miR-650, may also be of great diagnostic value for HCC patients [118]. Plenty of studies have demonstrated that miR-155 can act as a sensitive biomarker for monitoring the change in tumor size of breast cancer, more sensitive than canonical biomarkers such as carbohydrate antigen 15-3 (CA15-3), carcinoembryonic antigen (CEA), and tissue polypeptide specific antigen (TPS) [119,120]. The serum levels of miR-17, miR-34a, miR-155, and miR-373 have proven to be related to progression and metastasis of breast cancer [121]. Based on the existing evidence, two clinical trials to evaluate diagnostic potential of urinary miRNAs as well as blood miRNAs have been carried out [NCT03824613, NCT01391351]. miRNAs could be novel markers of cancer diagnosis and have enormous potential in clinical use in the future (Table 2).
6. Conclusions
miRNAs are key regulators of tumors. A plethora of studies revealed that plenty of miRNAs can regulate hallmarks of cancers including metastasis, therapy resistance, angiogenesis and tumor immunity. A therapeutic approach reversing abnormally expressed miRNAs is promising for cancer treatment and for the detection of abnormally expressed miRNAs might be used as diagnostic tools for cancer. Still, large cohort studies are required to validate the effect of miRNA-based diagnostic tools or therapeutic potential as well as miRNA-based drug safety. For the long haul, it is essential to find cancer specific miRNAs for better specificity as well as accuracy for cancer diagnosis as it is challenging to apply miRNAs-based therapy for cancer treatment.
Author Contributions
Y.S., Z.L.: investigation, data analysis and visualization, writing—original draft preparation; Q.L. (Qun Lin): data analysis and editing; Q.L. (Qing Luo): investigation, data analysis; Y.C.: investigation, writing and editing; J.L. and X.F.: investigation, writing and editing; C.G.: conceptualization, methodology, investigation, supervision, writing and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This work is supported by the National Key R&D Program of China (2017YFC1309103 and 2017YFC1309104); the Natural Science Foundation of China (81672594, 81772836, 81872139 and 82072907); Clinical Innovation Project of Bioland Laboratory (Guangzhou Regenerative Medicine and Health Guangdong Laboratory, 2018GZR0201004); Sun Yat-Sen memorial hospital cultivation project for clinical research (SYS-C-201805); Key Projects of The National Natural Science Foundation of China (51861125203);Project of The Beijing Xisike Clinical Oncology Research Foundation(Y-Roche2019/2-0078); Technology Development Program of Guangdong province (2021A0505030082); Project of The Guangdong Provincial Key Laboratory of Malignant Tumor Epigenetics and Gene Regulation (2020B1212060018OF007).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available upon request from the corresponding authors.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Galmarini, C.M. Lessons from Hippocrates: Time to Change the Cancer Paradigm. Int. J. Chronic Dis. 2020, 2020, 4715426. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Mun, E.J.; Babiker, H.M.; Weinberg, U.; Kirson, E.D.; Von Hoff, D.D. Tumor-treating fields: A fourth modality in cancer treatment. Clin. Cancer Res. 2018, 24, 266–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, J.; Kantoff, P.W.; Wooster, R.; Farokhzad, O.C. Cancer nanomedicine: Progress, challenges and opportunities. Nat. Rev. Cancer 2017, 17, 20–37. [Google Scholar] [CrossRef] [PubMed]
- Correia de Sousa, M.; Gjorgjieva, M.; Dolicka, D.; Sobolewski, C.; Foti, M. Deciphering mirnas’ action through mirna editing. Int. J. Mol. Sci. 2019, 20, 6249. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, M.A.; Arora, S.; Prakasam, G.; Calin, G.A.; Syed, M.A. MicroRNA in lung cancer: Role, mechanisms, pathways and therapeutic relevance. Mol. Asp. Med. 2019, 70, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Yadav, T.; Rani, V. Exploring miRNA based approaches in cancer diagnostics and therapeutics. Crit. Rev. Oncol. Hematol. 2016, 98, 12–23. [Google Scholar] [CrossRef]
- Lewis, B.P.; Burge, C.B.; Bartel, D.P. Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell 2005, 120, 15–20. [Google Scholar] [CrossRef] [Green Version]
- Shin, V.Y.; Chu, K.M. MiRNA as potential biomarkers and therapeutic targets for gastric cancer. World J. Gastroenterol. 2014, 20, 10432–10439. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Matsuyama, H.; Suzuki, H.I. Systems and Synthetic microRNA Biology: From Biogenesis to Disease Pathogenesis. Int. J. Mol. Sci. 2019, 21, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rupaimoole, R.; Slack, F.J. MicroRNA therapeutics: Towards a new era for the management of cancer and other diseases. Nat. Rev. Drug Discov. 2017, 16, 203–222. [Google Scholar] [CrossRef] [PubMed]
- Saliminejad, K.; Khorram Khorshid, H.R.; Soleymani Fard, S.; Ghaffari, S.H. An overview of microRNAs: Biology, functions, therapeutics, and analysis methods. J. Cell. Physiol. 2019, 234, 5451–5465. [Google Scholar] [CrossRef]
- Calin, G.A.; Dumitru, C.D.; Shimizu, M.; Bichi, R.; Zupo, S.; Noch, E.; Aldler, H.; Rattan, S.; Keating, M.; Rai, K.; et al. Frequent deletions and down-regulation of micro-RNA genes miR15 and miR16 at 13q14 in chronic lymphocytic leukemia. Proc. Natl. Acad. Sci. USA 2002, 99, 15524–15529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abreu, F.B.; Liu, X.; Tsongalis, G.J. miRNA analysis in pancreatic cancer: The Dartmouth experience. Clin. Chem. Lab. Med. 2017, 55, 755–762. [Google Scholar] [CrossRef]
- Du, X.; Zhang, J.; Wang, J.; Lin, X.; Ding, F. Role of miRNA in Lung Cancer-Potential Biomarkers and Therapies. Curr. Pharm. Des. 2018, 23, 5997–6010. [Google Scholar] [CrossRef] [PubMed]
- McGuire, A.; Brown, J.A.; Kerin, M.J. Metastatic breast cancer: The potential of miRNA for diagnosis and treatment monitoring. Cancer Metastasis Rev. 2015, 34, 145–155. [Google Scholar] [CrossRef] [Green Version]
- Qadir, M.I.; Faheem, A. miRNA: A Diagnostic and Therapeutic Tool for Pancreatic Cancer. Crit. Rev. Eukaryot. Gene Expr. 2017, 27, 197–204. [Google Scholar] [CrossRef]
- Wightman, B.; Ha, I.; Ruvkun, G. Posttranscriptional regulation of the heterochronic gene lin-14 by lin-4 mediates temporal pattern formation in C. elegans. Cell 1993, 75, 855–862. [Google Scholar] [CrossRef]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Reinhart, B.J.; Slack, F.J.; Basson, M.; Pasquinelli, A.E.; Bettinger, J.C.; Rougvie, A.E.; Horvitz, H.R.; Ruvkun, G. The 21-nucleotide let-7 RNA regulates developmental timing in Caenorhabditis elegans. Nature 2000, 403, 901–906. [Google Scholar] [CrossRef]
- Beermann, J.; Piccoli, M.T.; Viereck, J.; Thum, T. Non-coding RNAs in Development and Disease: Background, Mechanisms, and Therapeutic Approaches. Physiol. Rev. 2016, 96, 1297–1325. [Google Scholar] [CrossRef] [Green Version]
- Hammond, S.M. An overview of microRNAs. Adv. Drug Deliv. Rev. 2015, 87, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Lu, T.X.; Rothenberg, M.E. MicroRNA. J. Allergy Clin. Immunol. 2018, 141, 1202–1207. [Google Scholar] [CrossRef] [Green Version]
- Achkar, N.P.; Cambiagno, D.A.; Manavella, P.A. miRNA Biogenesis: A Dynamic Pathway. Trends Plant Sci. 2016, 21, 1034–1044. [Google Scholar] [CrossRef]
- Okada, C.; Yamashita, E.; Lee, S.J.; Shibata, S.; Katahira, J.; Nakagawa, A.; Yoneda, Y.; Tsukihara, T. A high-resolution structure of the pre-microRNA nuclear export machinery. Science 2009, 326, 1275–1279. [Google Scholar] [CrossRef]
- Sun, H.L.; Cui, R.; Zhou, J.; Teng, K.Y.; Hsiao, Y.H.; Nakanishi, K.; Fassan, M.; Luo, Z.; Shi, G.; Tili, E.; et al. ERK Activation Globally Downregulates miRNAs through Phosphorylating Exportin-5. Cancer Cell 2016, 30, 723–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef]
- Lee, Y.S.; Dutta, A. MicroRNAs in cancer. Annu. Rev. Pathol. 2009, 4, 199–227. [Google Scholar] [CrossRef] [PubMed]
- Elkayam, E.; Kuhn, C.D.; Tocilj, A.; Haase, A.D.; Greene, E.M.; Hannon, G.J.; Joshua-Tor, L. The structure of human argonaute-2 in complex with miR-20a. Cell 2012, 150, 100–110. [Google Scholar] [CrossRef] [Green Version]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA circles function as efficient microRNA sponges. Nature 2013, 495, 384–388. [Google Scholar] [CrossRef] [PubMed]
- Schirle, N.T.; Sheu-Gruttadauria, J.; MacRae, I.J. Structural basis for microRNA targeting. Science 2014, 346, 608–613. [Google Scholar] [CrossRef]
- Christie, M.; Boland, A.; Huntzinger, E.; Weichenrieder, O.; Izaurralde, E. Structure of the PAN3 pseudokinase reveals the basis for interactions with the PAN2 deadenylase and the GW182 proteins. Mol. Cell 2013, 51, 360–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabian, M.R.; Sonenberg, N. The mechanics of miRNA-mediated gene silencing: A look under the hood of miRISC. Nat. Struct. Mol. Biol. 2012, 19, 586–593. [Google Scholar] [CrossRef] [PubMed]
- Jonas, S.; Izaurralde, E. Towards a molecular understanding of microRNA-mediated gene silencing. Nat. Rev. Genet. 2015, 16, 421–433. [Google Scholar] [CrossRef]
- Xiao, M.; Li, J.; Li, W.; Wang, Y.; Wu, F.; Xi, Y.; Zhang, L.; Ding, C.; Luo, H.; Li, Y.; et al. MicroRNAs activate gene transcription epigenetically as an enhancer trigger. RNA Biol. 2017, 14, 1326–1334. [Google Scholar] [CrossRef]
- Han, T.S.; Hur, K.; Cho, H.S.; Ban, H.S. Epigenetic Associations between lncRNA/circRNA and miRNA in Hepatocellular Carcinoma. Cancers 2020, 12, 2622. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yujiao, W.; Fang, W.; Linhui, Y.; Ziqi, G.; Zhichen, W.; Zirui, W.; Shengwang, W. The roles of miRNA, lncRNA and circRNA in the development of osteoporosis. Biol. Res. 2020, 53, 40. [Google Scholar] [CrossRef]
- Tay, Y.; Rinn, J.; Pandolfi, P.P. The multilayered complexity of ceRNA crosstalk and competition. Nature 2014, 505, 344–352. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Hou, J.; He, D.; Sun, M.; Zhang, P.; Yu, Y.; Chen, Y. The Emerging Function and Mechanism of ceRNAs in Cancer. Trends Genet. 2016, 32, 211–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Ornitz, D.M.; Itoh, N. Fibroblast growth factors. Genome Biol. 2001, 2, reviews3005.1. [Google Scholar] [CrossRef] [Green Version]
- Nandy, S.B.; Mohanty, S.; Singh, M.; Behari, M.; Airan, B. Fibroblast Growth Factor-2 alone as an efficient inducer for differentiation of human bone marrow mesenchymal stem cells into dopaminergic neurons. J. Biomed. Sci. 2014, 21, 83. [Google Scholar] [CrossRef] [Green Version]
- Pizzute, T.; Li, J.; Zhang, Y.; Davis, M.E.; Pei, M. Fibroblast Growth Factor Ligand Dependent Proliferation and Chondrogenic Differentiation of Synovium-Derived Stem Cells and Concomitant Adaptation of Wnt/Mitogen-Activated Protein Kinase Signals. Tissue Eng. Part A 2016, 22, 1036–1046. [Google Scholar] [CrossRef]
- Jimenez-Pascual, A.; Hale, J.S.; Kordowski, A.; Pugh, J.; Silver, D.J.; Bayik, D.; Roversi, G.; Alban, T.J.; Rao, S.; Chen, R.; et al. ADAMDEC1 Maintains a Growth Factor Signaling Loop in Cancer Stem Cells. Cancer Discov. 2019, 9, 1574–1589. [Google Scholar] [CrossRef] [Green Version]
- Zaragosi, L.E.; Ailhaud, G.; Dani, C. Autocrine fibroblast growth factor 2 signaling is critical for self-renewal of human multipotent adipose-derived stem cells. Stem Cells 2006, 24, 2412–2419. [Google Scholar] [CrossRef] [PubMed]
- Kim do, Y.; Park, E.Y.; Chang, E.; Kang, H.G.; Koo, Y.; Lee, E.J.; Ko, J.Y.; Kong, H.K.; Chun, K.H.; Park, J.H. A novel miR-34a target, protein kinase D1, stimulates cancer stemness and drug resistance through GSK3/β-catenin signaling in breast cancer. Oncotarget 2016, 7, 14791–14802. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Liu, J.; Shen, J.; Liu, L.; Wu, J.; Li, W.; Luo, J.; Chen, Q.; Qian, C. Expression of miR-122 mediated by adenoviral vector induces apoptosis and cell cycle arrest of cancer cells. Cancer Biol. Ther. 2010, 9, 554–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, G.; Li, Y.; Ma, Y.; Lu, J.; Chen, Y.; Jiang, Q.; Qin, Q.; Zhao, L.; Huang, Q.; Luo, Z.; et al. Long noncoding RNA LINC00511 contributes to breast cancer tumourigenesis and stemness by inducing the miR-185-3p/E2F1/Nanog axis. J. Exp. Clin. Cancer Res. 2018, 37, 289. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.X.; Yuan, L.; Xue, X.L.; Zhou, M.; Liu, Y.; Zhang, C.; Li, J.P.; Zheng, L.; Hong, M.; Li, X.N. Regulation of colorectal carcinoma stemness, growth, and metastasis by an miR-200c-Sox2-negative feedback loop mechanism. Clin. Cancer Res. 2014, 20, 2631–2642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittal, V. Epithelial Mesenchymal Transition in Tumor Metastasis. Annu. Rev. Pathol. 2018, 13, 395–412. [Google Scholar] [CrossRef]
- Peng, S.Y.; Tu, H.F.; Yang, C.C.; Wu, C.H.; Liu, C.J.; Chang, K.W.; Lin, S.C. miR-134 targets PDCD7 to reduce E-cadherin expression and enhance oral cancer progression. Int. J. Cancer 2018, 143, 2892–2904. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Xiong, H.; Duan, L.; Li, Q.; Li, X.; Zhou, Y. MiR-1246 Promotes Metastasis and Invasion of A549 cells by Targeting GSK-3β‒Mediated Wnt/β-Catenin Pathway. Cancer Res. Treat. 2019, 51, 1420–1429. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Zhang, X.; Du, L.; Wang, Y.; Liu, X.; Tian, H.; Wang, L.; Li, P.; Zhao, Y.; Duan, W.; et al. Exosome-transmitted miR-128-3p increase chemosensitivity of oxaliplatin-resistant colorectal cancer. Mol. Cancer 2019, 18, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Luo, W.; Yang, Z.J.; Chi, J.R.; Li, Y.R.; Ding, Y.; Ge, J.; Wang, X.; Cao, X.C. miR-190 suppresses breast cancer metastasis by regulation of TGF-β-induced epithelial-mesenchymal transition. Mol. Cancer 2018, 17, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Lu, L.L.; Wen, D.; Liu, D.L.; Dong, L.L.; Gao, D.M.; Bian, X.Y.; Zhou, J.; Fan, J.; Wu, W.Z. MiR-612 regulates invadopodia of hepatocellular carcinoma by HADHA-mediated lipid reprogramming. J. Hematol. Oncol. 2020, 13, 12. [Google Scholar] [CrossRef]
- Pan, J.; Fang, S.; Tian, H.; Zhou, C.; Zhao, X.; Tian, H.; He, J.; Shen, W.; Meng, X.; Jin, X.; et al. lncRNA JPX/miR-33a-5p/Twist1 axis regulates tumorigenesis and metastasis of lung cancer by activating Wnt/β-catenin signaling. Mol. Cancer 2020, 19, 9. [Google Scholar] [CrossRef] [PubMed]
- Fischer, K.R.; Durrans, A.; Lee, S.; Sheng, J.; Li, F.; Wong, S.T.; Choi, H.; El Rayes, T.; Ryu, S.; Troeger, J.; et al. Epithelial-to-mesenchymal transition is not required for lung metastasis but contributes to chemoresistance. Nature 2015, 527, 472–476. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Liao, H.; Pu, Q.; Ke, X.; Hu, X.; Ma, Y.; Luo, X.; Jiang, Q.; Gong, Y.; Wu, M.; et al. miR-410 induces both epithelial-mesenchymal transition and radioresistance through activation of the PI3K/mTOR pathway in non-small cell lung cancer. Signal Transduct. Target. Ther. 2020, 5, 85. [Google Scholar] [CrossRef]
- Coffman, J.A. Cell cycle development. Dev. Cell 2004, 6, 321–327. [Google Scholar] [CrossRef] [Green Version]
- Mens, M.M.J.; Ghanbari, M. Cell Cycle Regulation of Stem Cells by MicroRNAs. Stem. Cell Rev. Rep. 2018, 14, 309–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bueno, M.J.; Malumbres, M. MicroRNAs and the cell cycle. Biochim. Biophys. Acta BBA-Mol. Basis Dis. 2011, 1812, 592–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Li, J.; Huang, Y.; Peng, W.; Qian, W.; Gu, J.; Wang, Q.; Hu, T.; Ji, D.; Ji, B.; et al. Upregulated miR-1258 regulates cell cycle and inhibits cell proliferation by directly targeting E2F8 in CRC. Cell Prolif. 2018, 51, e12505. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Li, Z.; Hao, Q.; Tan, W.; Sun, J.; Li, J.; Chen, C.W.; Li, Z.; Meng, Y.; Zhou, Y.; et al. The Cdk2-c-Myc-miR-571 Axis Regulates DNA Replication and Genomic Stability by Targeting Geminin. Cancer Res. 2019, 79, 4896–4910. [Google Scholar] [CrossRef] [Green Version]
- Kir, D.; Schnettler, E.; Modi, S.; Ramakrishnan, S. Regulation of angiogenesis by microRNAs in cardiovascular diseases. Angiogenesis 2018, 21, 699–710. [Google Scholar] [CrossRef]
- Viallard, C.; Larrivée, B. Tumor angiogenesis and vascular normalization: Alternative therapeutic targets. Angiogenesis 2017, 20, 409–426. [Google Scholar] [CrossRef]
- Pencheva, N.; Tran, H.; Buss, C.; Huh, D.; Drobnjak, M.; Busam, K.; Tavazoie, S.F. Convergent multi-miRNA targeting of ApoE drives LRP1/LRP8-dependent melanoma metastasis and angiogenesis. Cell 2012, 151, 1068–1082. [Google Scholar] [CrossRef] [Green Version]
- Roy, S.; Sen, C.K. miRNA in wound inflammation and angiogenesis. Microcirculation 2012, 19, 224–232. [Google Scholar] [CrossRef] [Green Version]
- Sun, R.; Wu, J.; Chen, Y.; Lu, M.; Zhang, S.; Lu, D.; Li, Y. Down regulation of Thrombospondin2 predicts poor prognosis in patients with gastric cancer. Mol. Cancer 2014, 13, 225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, Y.; Wang, C.; Yang, Z.; Liu, W.; Yuan, Y.; Li, K.; Zhang, Y.; Wang, Y.; Shi, Y.; Qiu, Y.; et al. Dysregulated Sp1/miR-130b-3p/HOXA5 axis contributes to tumor angiogenesis and progression of hepatocellular carcinoma. Theranostics 2020, 10, 5209–5224. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.G.; Zhou, C.F.; Zhang, Y.M.; Yan, R.M.; Wei, W.F.; Chen, X.J.; Yi, H.Y.; Liang, L.J.; Fan, L.S.; Liang, L.; et al. Cancer-derived exosomal miR-221-3p promotes angiogenesis by targeting THBS2 in cervical squamous cell carcinoma. Angiogenesis 2019, 22, 397–410. [Google Scholar] [CrossRef]
- Sun, Z.; Shi, K.; Yang, S.; Liu, J.; Zhou, Q.; Wang, G.; Song, J.; Li, Z.; Zhang, Z.; Yuan, W. Effect of exosomal miRNA on cancer biology and clinical applications. Mol. Cancer 2018, 17, 147. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Zhu, C.; Pang, Q.; Liu, H.C. MicroRNA-217: A regulator of human cancer. Biomed. Pharm. 2021, 133, 110943. [Google Scholar] [CrossRef]
- Bertoli, G.; Cava, C.; Castiglioni, I. MicroRNAs: New Biomarkers for Diagnosis, Prognosis, Therapy Prediction and Therapeutic Tools for Breast Cancer. Theranostics 2015, 5, 1122–1143. [Google Scholar] [CrossRef]
- Hayes, J.; Peruzzi, P.P.; Lawler, S. MicroRNAs in cancer: Biomarkers, functions and therapy. Trends Mol. Med. 2014, 20, 460–469. [Google Scholar] [CrossRef]
- Heneghan, H.M.; Miller, N.; Kerin, M.J. MiRNAs as biomarkers and therapeutic targets in cancer. Curr. Opin. Pharmacol. 2010, 10, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Getz, G.; Miska, E.A.; Alvarez-Saavedra, E.; Lamb, J.; Peck, D.; Sweet-Cordero, A.; Ebert, B.L.; Mak, R.H.; Ferrando, A.A.; et al. MicroRNA expression profiles classify human cancers. Nature 2005, 435, 834–838. [Google Scholar] [CrossRef] [PubMed]
- Bandres, E.; Bitarte, N.; Arias, F.; Agorreta, J.; Fortes, P.; Agirre, X.; Zarate, R.; Diaz-Gonzalez, J.A.; Ramirez, N.; Sola, J.J.; et al. microRNA-451 regulates macrophage migration inhibitory factor production and proliferation of gastrointestinal cancer cells. Clin. Cancer Res. 2009, 15, 2281–2290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janssen, H.L.; Reesink, H.W.; Lawitz, E.J.; Zeuzem, S.; Rodriguez-Torres, M.; Patel, K.; van der Meer, A.J.; Patick, A.K.; Chen, A.; Zhou, Y.; et al. Treatment of HCV infection by targeting microRNA. N. Engl. J. Med. 2013, 368, 1685–1694. [Google Scholar] [CrossRef] [Green Version]
- Pan, C.; Stevic, I.; Müller, V.; Ni, Q.; Oliveira-Ferrer, L.; Pantel, K.; Schwarzenbach, H. Exosomal microRNAs as tumor markers in epithelial ovarian cancer. Mol. Oncol. 2018, 12, 1935–1948. [Google Scholar] [CrossRef] [Green Version]
- Mellis, D.; Caporali, A. Microrna-based therapeutics in cardiovascular disease: Screening and delivery to the target. Biochem. Soc. Trans. 2018, 46, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Chai, C.; Cox, B.; Yaish, D.; Gross, D.; Rosenberg, N.; Amblard, F.; Shemuelian, Z.; Gefen, M.; Korach, A.; Tirosh, O.; et al. Agonist of RORA Attenuates Nonalcoholic Fatty Liver Progression in Mice via Up-regulation of MicroRNA 122. Gastroenterology 2020, 159, 999–1014. [Google Scholar] [CrossRef]
- Hsu, S.H.; Wang, B.; Kota, J.; Yu, J.; Costinean, S.; Kutay, H.; Yu, L.; Bai, S.; La Perle, K.; Chivukula, R.R.; et al. Essential metabolic, anti-inflammatory, and anti-tumorigenic functions of miR-122 in liver. J. Clin. Investig. 2012, 122, 2871–2883. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.H.; Kim, J.H.; Lee, S.W. The role of microRNAs in hepatitis C virus replication and related liver diseases. J. Microbiol. 2014, 52, 445–451. [Google Scholar] [CrossRef]
- Luna, J.M.; Scheel, T.K.; Danino, T.; Shaw, K.S.; Mele, A.; Fak, J.J.; Nishiuchi, E.; Takacs, C.N.; Catanese, M.T.; de Jong, Y.P.; et al. Hepatitis C virus RNA functionally sequesters miR-122. Cell 2015, 160, 1099–1110. [Google Scholar] [CrossRef] [Green Version]
- Elmén, J.; Lindow, M.; Schütz, S.; Lawrence, M.; Petri, A.; Obad, S.; Lindholm, M.; Hedtjärn, M.; Hansen, H.F.; Berger, U.; et al. LNA-mediated microRNA silencing in non-human primates. Nature 2008, 452, 896–899. [Google Scholar] [CrossRef]
- Elmén, J.; Lindow, M.; Silahtaroglu, A.; Bak, M.; Christensen, M.; Lind-Thomsen, A.; Hedtjärn, M.; Hansen, J.B.; Hansen, H.F.; Straarup, E.M.; et al. Antagonism of microRNA-122 in mice by systemically administered LNA-antimiR leads to up-regulation of a large set of predicted target mRNAs in the liver. Nucleic Acids Res. 2008, 36, 1153–1162. [Google Scholar] [CrossRef] [Green Version]
- Athanasopoulos, T.; Munye, M.M.; Yáñez-Muñoz, R.J. Nonintegrating Gene Therapy Vectors. Hematol. Oncol. Clin. N. Am. 2017, 31, 753–770. [Google Scholar] [CrossRef] [PubMed]
- Crystal, R.G. Adenovirus: The first effective in vivo gene delivery vector. Hum. Gene Ther. 2014, 25, 3–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mir, M.; Ahmed, N.; Rehman, A.U. Recent applications of PLGA based nanostructures in drug delivery. Colloids Surf. B Biointerfaces 2017, 159, 217–231. [Google Scholar] [CrossRef]
- Zhu, S.; Xing, H.; Gordiichuk, P.; Park, J.; Mirkin, C.A. PLGA Spherical Nucleic Acids. Adv. Mater. 2018, 30, e1707113. [Google Scholar] [CrossRef]
- Taylor, K.; Howard, C.B.; Jones, M.L.; Sedliarou, I.; MacDiarmid, J.; Brahmbhatt, H.; Munro, T.P.; Mahler, S.M. Nanocell targeting using engineered bispecific antibodies. MAbs 2015, 7, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Sagnella, S.M.; Yang, L.; Stubbs, G.E.; Boslem, E.; Martino-Echarri, E.; Smolarczyk, K.; Pattison, S.L.; Vanegas, N.; St Clair, E.; Clarke, S.; et al. Cyto-Immuno-Therapy for Cancer: A Pathway Elicited by Tumor-Targeted, Cytotoxic Drug-Packaged Bacterially Derived Nanocells. Cancer Cell 2020, 37, 354–370. [Google Scholar] [CrossRef] [PubMed]
- MacDiarmid, J.A.; Mugridge, N.B.; Weiss, J.C.; Phillips, L.; Burn, A.L.; Paulin, R.P.; Haasdyk, J.E.; Dickson, K.A.; Brahmbhatt, V.N.; Pattison, S.T.; et al. Bacterially derived 400 nm particles for encapsulation and cancer cell targeting of chemotherapeutics. Cancer Cell 2007, 11, 431–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, S.; Tian, H.; Guo, Y.; Li, Y.; Guo, Z.; Zhu, X.; Chen, X. miRNA oligonucleotide and sponge for miRNA-21 inhibition mediated by PEI-PLL in breast cancer therapy. Acta Biomater. 2015, 25, 184–193. [Google Scholar] [CrossRef]
- Ou, L.; Lin, H.; Song, Y.; Tan, G.; Gui, X.; Li, J.; Chen, X.; Deng, Z.; Lin, S. Efficient miRNA Inhibitor with GO-PEI Nanosheets for Osteosarcoma Suppression by Targeting PTEN. Int. J. Nanomed. 2020, 15, 5131–5146. [Google Scholar] [CrossRef]
- Xue, J.; Chi, Y.; Chen, Y.; Huang, S.; Ye, X.; Niu, J.; Wang, W.; Pfeffer, L.M.; Shao, Z.M.; Wu, Z.H.; et al. MiRNA-621 sensitizes breast cancer to chemotherapy by suppressing FBXO11 and enhancing p53 activity. Oncogene 2016, 35, 448–458. [Google Scholar] [CrossRef] [Green Version]
- Naseri, Z.; Oskuee, R.K.; Jaafari, M.R.; Forouzandeh Moghadam, M. Exosome-mediated delivery of functionally active miRNA-142-3p inhibitor reduces tumorigenicity of breast cancer in vitro and in vivo. Int. J. Nanomed. 2018, 13, 7727–7747. [Google Scholar] [CrossRef] [Green Version]
- Naseri, Z.; Oskuee, R.K.; Forouzandeh-Moghadam, M.; Jaafari, M.R. Delivery of LNA-antimiR-142-3p by Mesenchymal Stem Cells-Derived Exosomes to Breast Cancer Stem Cells Reduces Tumorigenicity. Stem Cell Rev. Rep. 2020, 16, 541–556. [Google Scholar] [CrossRef]
- Beg, M.S.; Brenner, A.J.; Sachdev, J.; Borad, M.; Kang, Y.K.; Stoudemire, J.; Smith, S.; Bader, A.G.; Kim, S.; Hong, D.S. Phase I study of MRX34, a liposomal miR-34a mimic, administered twice weekly in patients with advanced solid tumors. Investig. New Drugs 2017, 35, 180–188. [Google Scholar] [CrossRef]
- Hong, D.S.; Kang, Y.K.; Borad, M.; Sachdev, J.; Ejadi, S.; Lim, H.Y.; Brenner, A.J.; Park, K.; Lee, J.L.; Kim, T.Y.; et al. Phase 1 study of MRX34, a liposomal miR-34a mimic, in patients with advanced solid tumours. Br. J. Cancer 2020, 122, 1630–1637. [Google Scholar] [CrossRef]
- Cortez, M.A.; Ivan, C.; Valdecanas, D.; Wang, X.; Peltier, H.J.; Ye, Y.; Araujo, L.; Carbone, D.P.; Shilo, K.; Giri, D.K.; et al. PDL1 Regulation by p53 via miR-34. J. Natl. Cancer Inst. 2016, 108. [Google Scholar] [CrossRef] [Green Version]
- He, L.; He, X.; Lim, L.P.; de Stanchina, E.; Xuan, Z.; Liang, Y.; Xue, W.; Zender, L.; Magnus, J.; Ridzon, D.; et al. A microRNA component of the p53 tumour suppressor network. Nature 2007, 447, 1130–1134. [Google Scholar] [CrossRef] [Green Version]
- Volinia, S.; Calin, G.A.; Liu, C.G.; Ambs, S.; Cimmino, A.; Petrocca, F.; Visone, R.; Iorio, M.; Roldo, C.; Ferracin, M.; et al. A microRNA expression signature of human solid tumors defines cancer gene targets. Proc. Natl. Acad. Sci. USA 2006, 103, 2257–2261. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Dou, R.; Yin, T.; Ding, J. MiRNA-106b-5p in human cancers: Diverse functions and promising biomarker. Biomed. Pharm. 2020, 127, 110211. [Google Scholar] [CrossRef]
- Pekarsky, Y.; Croce, C.M. Role of miR-15/16 in CLL. Cell Death Differ. 2015, 22, 6–11. [Google Scholar] [CrossRef] [Green Version]
- Gironella, M.; Seux, M.; Xie, M.J.; Cano, C.; Tomasini, R.; Gommeaux, J.; Garcia, S.; Nowak, J.; Yeung, M.L.; Jeang, K.T.; et al. Tumor protein 53-induced nuclear protein 1 expression is repressed by miR-155, and its restoration inhibits pancreatic tumor development. Proc. Natl. Acad. Sci. USA 2007, 104, 16170–16175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tili, E.; Croce, C.M.; Michaille, J.J. miR-155: On the crosstalk between inflammation and cancer. Int. Rev. Immunol. 2009, 28, 264–284. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.; He, L.; Richards, E.J.; Challa, S.; Xu, C.X.; Permuth-Wey, J.; Lancaster, J.M.; Coppola, D.; Sellers, T.A.; Djeu, J.Y.; et al. Upregulation of miRNA-155 promotes tumour angiogenesis by targeting VHL and is associated with poor prognosis and triple-negative breast cancer. Oncogene 2014, 33, 679–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Backes, C.; Meese, E.; Keller, A. Specific miRNA Disease Biomarkers in Blood, Serum and Plasma: Challenges and Prospects. Mol. Diagn. Ther. 2016, 20, 509–518. [Google Scholar] [CrossRef]
- Kappel, A.; Keller, A. miRNA assays in the clinical laboratory: Workflow, detection technologies and automation aspects. Clin. Chem. Lab. Med. 2017, 55, 636–647. [Google Scholar] [CrossRef] [PubMed]
- Schulte, C.; Karakas, M.; Zeller, T. microRNAs in cardiovascular disease-Clinical application. Clin. Chem. Lab. Med. 2017, 55, 687–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bansal, A.; Hong, X.; Lee, I.H.; Krishnadath, K.K.; Mathur, S.C.; Gunewardena, S.; Rastogi, A.; Sharma, P.; Christenson, L.K. MicroRNA Expression can be a Promising Strategy for the Detection of Barrett’s Esophagus: A Pilot Study. Clin. Transl. Gastroenterol. 2014, 5, e65. [Google Scholar] [CrossRef]
- Pavel, A.B.; Campbell, J.D.; Liu, G.; Elashoff, D.; Dubinett, S.; Smith, K.; Whitney, D.; Lenburg, M.E.; Spira, A. Alterations in Bronchial Airway miRNA Expression for Lung Cancer Detection. Cancer Prev. Res. 2017, 10, 651–659. [Google Scholar] [CrossRef] [Green Version]
- Botti, G.; Giordano, A.; Feroce, F.; De Chiara, A.R.; Cantile, M. Noncoding RNAs as circulating biomarkers in osteosarcoma patients. J. Cell Physiol. 2019, 234, 19249–19255. [Google Scholar] [CrossRef]
- Andersen, G.B.; Knudsen, A.; Hager, H.; Hansen, L.L.; Tost, J. miRNA profiling identifies deregulated miRNAs associated with osteosarcoma development and time to metastasis in two large cohorts. Mol. Oncol. 2018, 12, 114–131. [Google Scholar] [CrossRef]
- Qu, K.Z.; Zhang, K.; Li, H.; Afdhal, N.H.; Albitar, M. Circulating microRNAs as biomarkers for hepatocellular carcinoma. J. Clin. Gastroenterol. 2011, 45, 355–360. [Google Scholar] [CrossRef]
- Abdalla, M.A.; Haj-Ahmad, Y. Promising Candidate Urinary MicroRNA Biomarkers for the Early Detection of Hepatocellular Carcinoma among High-Risk Hepatitis C Virus Egyptian Patients. J. Cancer 2012, 3, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Wang, M.; Lin, G.; Sun, S.; Li, X.; Qi, J.; Li, J. Serum microRNA-155 as a potential biomarker to track disease in breast cancer. PLoS ONE 2012, 7, e47003. [Google Scholar] [CrossRef]
- Cheng, G. Circulating miRNAs: Roles in cancer diagnosis, prognosis and therapy. Adv. Drug Deliv. Rev. 2015, 81, 75–93. [Google Scholar] [CrossRef]
- Eichelser, C.; Flesch-Janys, D.; Chang-Claude, J.; Pantel, K.; Schwarzenbach, H. Deregulated serum concentrations of circulating cell-free microRNAs miR-17, miR-34a, miR-155, and miR-373 in human breast cancer development and progression. Clin. Chem. 2013, 59, 1489–1496. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
miRNA biogenesis and mechanism of action. miRNA production remains conserved across species. RNA polymerase II transcribes pri-miRNA from genome sequence. Drosha and its cofactor protein bind to primary miRNAs (pri-miRNA) leading to the excision of the loop structure to generate precursor miRNA (pre-miRNA). Then Exportin-5 transports pre-miRNA from nucleus to cytosol. Dicer complex, composed of TAR RNA-binding protein (TRBP) and protein activator of the interferon-induced protein kinase (PACT), manipulates maturation of miRNA and formation of RNA-induced silencing complex (RISC) complex. Mature RISC complex binds to target mRNA with complementary sites, resulting in the translational suppression or target degradation.
Figure 1.
miRNA biogenesis and mechanism of action. miRNA production remains conserved across species. RNA polymerase II transcribes pri-miRNA from genome sequence. Drosha and its cofactor protein bind to primary miRNAs (pri-miRNA) leading to the excision of the loop structure to generate precursor miRNA (pre-miRNA). Then Exportin-5 transports pre-miRNA from nucleus to cytosol. Dicer complex, composed of TAR RNA-binding protein (TRBP) and protein activator of the interferon-induced protein kinase (PACT), manipulates maturation of miRNA and formation of RNA-induced silencing complex (RISC) complex. Mature RISC complex binds to target mRNA with complementary sites, resulting in the translational suppression or target degradation.

Figure 2.
miRNAs modulate different hallmarks of cancer. miRNAs manipulate a set of biological processes that ultimately influences the proliferation and migration of cancer cells. ADAMDEC1, a disintegrin and metalloproteinase domain-like protein decysin 1; FEF2, Fibroblast growth factor 2; FGFR1, FGF receptor 1; ZEB1, Zinc-finger E-box-binding homeobox 1; PRKD1, serine/threonine-protein kinase D1; EMT, Epithelial–mesenchymal transition; PDCD7, Programmed Cell Death 7; CCND1, cyclin D1; p21, cyclin dependent kinase inhibitor 1A; THBS2, Thrombospondin-2.
Figure 2.
miRNAs modulate different hallmarks of cancer. miRNAs manipulate a set of biological processes that ultimately influences the proliferation and migration of cancer cells. ADAMDEC1, a disintegrin and metalloproteinase domain-like protein decysin 1; FEF2, Fibroblast growth factor 2; FGFR1, FGF receptor 1; ZEB1, Zinc-finger E-box-binding homeobox 1; PRKD1, serine/threonine-protein kinase D1; EMT, Epithelial–mesenchymal transition; PDCD7, Programmed Cell Death 7; CCND1, cyclin D1; p21, cyclin dependent kinase inhibitor 1A; THBS2, Thrombospondin-2.

Figure 3.
Chemical modifications and delivery system of miRNA in vivo application. The chemical modifications of miRNA mimics and antimiRs increase their stability to ensure the integrality of miRNA-based therapeutic particles, while development of delivery systems facilitates the efficiency of miRNA therapy in vivo. Some of the commonly used delivery vehicles includes adenoviral vector, Poly (lactide-co-glycolide) (PGLA), EnGeneIC Delivery Vehicle (EDV) nanocells and polyethylenimine (PEI) molecules. Safety issue as well as tumor-specific delivery systems are still tested in animal models and clinical trials.
Figure 3.
Chemical modifications and delivery system of miRNA in vivo application. The chemical modifications of miRNA mimics and antimiRs increase their stability to ensure the integrality of miRNA-based therapeutic particles, while development of delivery systems facilitates the efficiency of miRNA therapy in vivo. Some of the commonly used delivery vehicles includes adenoviral vector, Poly (lactide-co-glycolide) (PGLA), EnGeneIC Delivery Vehicle (EDV) nanocells and polyethylenimine (PEI) molecules. Safety issue as well as tumor-specific delivery systems are still tested in animal models and clinical trials.

Table 2.
miRNAs with diagnostic potential in cancers.
| Disease | miRNAs | Samples | Significances | Ref. |
|---|---|---|---|---|
| Barret’s esophagus | miR-192-5p, miR-194-5p and miR-215-5p | serum | The area under the curve (AUC) to diagnose BE by combining of these three miRNAs can reach 0.96–0.97. | [113] |
| Lung cancer | miR-146a-5p, miR-324-5p, miR-223-3p and miR-223-5p | bronchial epithelium | These four miRNAs are remarkably downregulated in lung cancer patients. | [114] |
| epithelial ovarian cancer | miR-200b | serum | High level of miR-200b can predict poor overall survival of patients. | [80] |
| Osteosarcoma | miR-221 and miR-222 | serum | The upregulation of these two miRNAs is related with higher metastasis risk and poor prognosis. | [116] |
| Hepatocellular carcinoma (HCC) | miR-16 | serum | The different expression of miR-16 can significantly increase the sensitivity of diagnosis. | [117] |
| Hepatocellular carcinoma (HCC) | miR-618 and miR-650 | urine | The upregulation of these two miRNAs can increase the sensitivity of the diagnosis of HCC patients. | [118] |
| Breast cancer | miR-155 | serum | miR-155 can act as a sensitive biomarker for monitoring the change in tumor size of breast cancer. | [119] |
| Breast cancer | miR-17, miR-34a, miR-155 and miR-373 | serum | These four miRNAs showed significantly different expressions in serum of breast cancer patients. | [121] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Shi, Y.; Liu, Z.; Lin, Q.; Luo, Q.; Cen, Y.; Li, J.; Fang, X.; Gong, C. MiRNAs and Cancer: Key Link in Diagnosis and Therapy. Genes 2021, 12, 1289. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12081289
AMA Style
Shi Y, Liu Z, Lin Q, Luo Q, Cen Y, Li J, Fang X, Gong C. MiRNAs and Cancer: Key Link in Diagnosis and Therapy. Genes. 2021; 12(8):1289. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12081289
Chicago/Turabian StyleShi, Yu, Zihao Liu, Qun Lin, Qing Luo, Yinghuan Cen, Juanmei Li, Xiaolin Fang, and Chang Gong. 2021. "MiRNAs and Cancer: Key Link in Diagnosis and Therapy" Genes 12, no. 8: 1289. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12081289
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.