
Association of Gene Variants for Mechanical and Metabolic Muscle Quality with Cardiorespiratory and Muscular Variables Related to Performance in Skiing Athletes
 , ,
, , 
Abstract
:1. Introduction
2. Material & Methods
2.1. Subjects
2.2. Ethics
2.3. Design
2.4. Peak Performance of Single Contractions
2.5. Aerobic Performance
2.6. Genotyping
2.7. Data Handling
2.8. Statistics
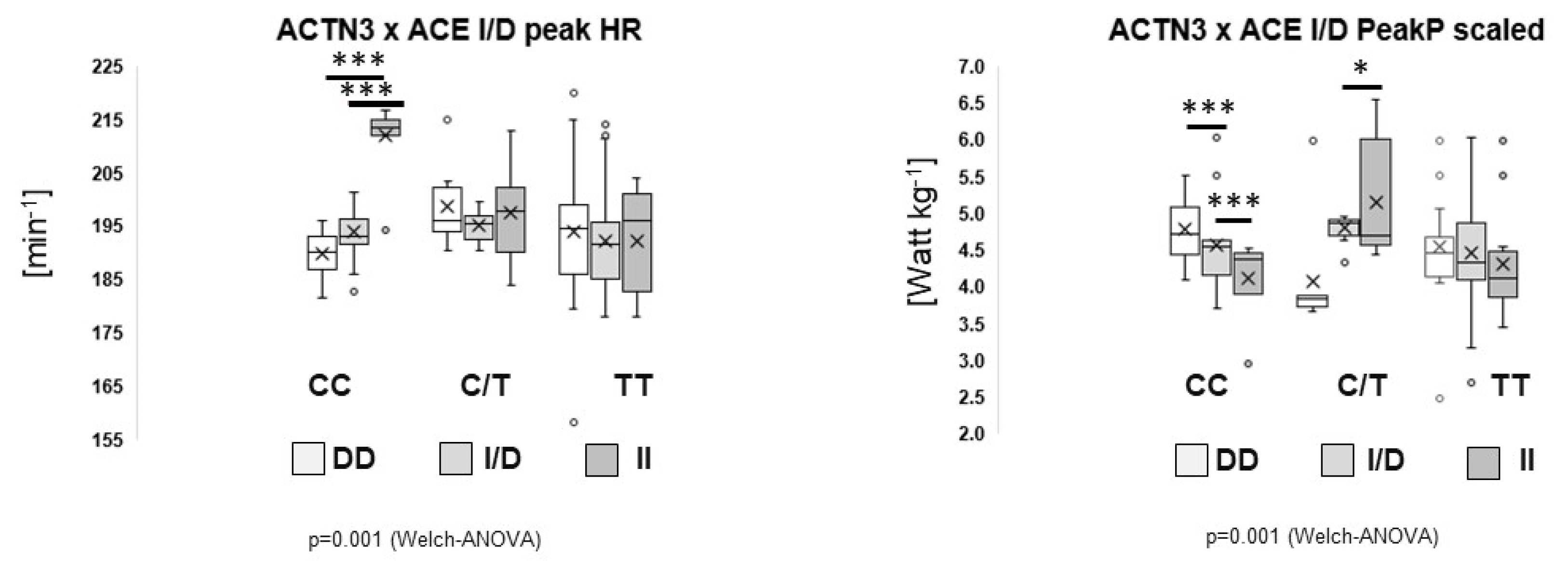
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tesch, P.A. Aspects on muscle properties and use in competitive Alpine skiing. Med. Sci. Sports Exerc. 1995, 27, 310–314. [Google Scholar] [CrossRef] [PubMed]
- Turnbull, J.R.; Kilding, A.E.; Keogh, J.W.L. Physiology of alpine skiing. Scand. J. Med. Sci. Sports 2009, 19, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Gilgien, M.; Reid, R.; Raschner, C.; Supej, M.; Holmberg, H.-C. The Training of Olympic Alpine Ski Racers. Front. Physiol. 2018, 9, 1772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flück, M.; Kramer, M.; Fitze, D.P.; Kasper, S.; Franchi, M.V.; Valdivieso, P. Cellular Aspects of Muscle Specialization Demonstrate Genotype–Phenotype Interaction Effects in Athletes. Front. Physiol. 2019, 10, 526. [Google Scholar] [CrossRef] [Green Version]
- Kröll, J.; Spörri, J.; Kandler, C.; Fasel, B.; Müller, E. Kinetic and kinematic comparison of alpine ski racing disciplines as a base for specific conditioning regimes. ISBS Proc. Arch. 2015, 33, 6526. Available online: https://ojs.ub.uni-konstanz.de/cpa/article/view/6526 (accessed on 1 August 2022).
- Gilgien, M.; Kröll, J.; Spörri, J.; Crivelli, P.; Müller, E. Application of dGNSS in Alpine Ski Racing: Basis for Evaluating Physical Demands and Safety. Front. Physiol. 2018, 9, 145. [Google Scholar] [CrossRef] [Green Version]
- AlHammoud, M.; Hansen, C.; Meyer, F.; Hautier, C.; Morel, B. On-Field Ski Kinematic According to Leg and Discipline in Elite Alpine Skiers. Front. Sports Act. Living 2020, 2, 56. [Google Scholar] [CrossRef] [PubMed]
- Sandbakk, Ø.; Holmberg, H.-C. Physiological Capacity and Training Routines of Elite Cross-Country Skiers: Approaching the Upper Limits of Human Endurance. Int. J. Sports Physiol. Perform. 2017, 12, 1003–1011. [Google Scholar] [CrossRef]
- Vogt, M.; Däpp, C.; Blatter, J.; Hoppeler, H. Training zur Optimierung der Dosierung exzentrischer Muskelaktivität. Schweiz. Z. Sportmed. Sporttraumatol. 2003, 51, 188–191. [Google Scholar]
- Andersen, R.E.; Montgomery, D.L. Physiology of Alpine Skiing. Sports Med. 1988, 6, 210–221. [Google Scholar] [CrossRef]
- Berg, H.E.; Eiken, O. Muscle control in elite alpine skiing. Med. Sci. Sports Exerc. 1999, 31, 1065–1067. [Google Scholar] [CrossRef] [PubMed]
- Ferland, P.-M.; Comtois, A.S. Athletic Profile of Alpine Ski Racers: A Systematic Review. J. Strength Cond. Res. 2018, 32, 3574–3583. [Google Scholar] [CrossRef] [PubMed]
- Gilgien, M.; Spörri, J.; Kröll, J.; Crivelli, P.; Müller, E. Mechanics of turning and jumping and skier speed are associated with injury risk in men′s World Cup alpine skiing: A comparison between the competition disciplines. Br. J. Sports Med. 2014, 48, 742–747. [Google Scholar] [CrossRef] [Green Version]
- Kröll, J.; Spörri, J.; Fasel, B.; Müller, E.; Schwameder, H. Type of muscle control in elite Alpine skiing: Is it still the same than in 1995? In Proceedings of the Science and Skiing VI: 6th International Congress on Science and Skiing, St. Christoph am Arlberg, Austria, 14–19 December 2013. [Google Scholar]
- Bäurle, L. Sliding Friction of Polyethylene on Snow and Ice. Ph.D. Thesis, ETH, Zürich, Switzerland, 2006. [Google Scholar]
- Alhammoud, M.; Racinais, S.; Dorel, S.; Guilhem, G.; Hautier, C.A.; Morel, B. Muscle-tendon unit length changes in knee extensors and flexors during alpine skiing. Sports Biomech. 2021, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Eisenman, P.A.; Johnson, S.C.; Bainbridge, C.N.; Zupan, M.F. Applied Physiology of Cross-Country Skiing. Sports Med. 1989, 8, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Valdivieso, P.; Vaughan, D.; Laczko, E.; Brogioli, M.; Waldron, S.; Rittweger, J.; Flück, M. The Metabolic Response of Skeletal Muscle to Endurance Exercise Is Modified by the ACE-I/D Gene Polymorphism and Training State. Front. Physiol. 2017, 8, 993. [Google Scholar] [CrossRef] [Green Version]
- Flück, M.; Hoppeler, H. Molecular basis of skeletal muscle plasticity-from gene to form and function. Rev. Physiol. Biochem. Pharmacol. 2003, 146, 159–216. [Google Scholar] [CrossRef]
- MacArthur, D.G.; North, K.N. ACTN3: A genetic influence on muscle function and athletic performance. Exerc. Sport Sci. Rev. 2007, 35, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Gremlich, S.; Cremona, T.P.; Yao, E.; Chabenet, F.; Fytianos, K.; Roth-Kleiner, M.; Schittny, J.C. Tenascin-C: Friend or Foe in Lung Aging? Front. Physiol. 2021, 12, 749776. [Google Scholar] [CrossRef]
- Erskine, R.M.; Williams, A.G.; Jones, D.A.; Stewart, C.E.; Degens, H. Do PTK2 gene polymorphisms contribute to the interindividual variability in muscle strength and the response to resistance training? A preliminary report. J. Appl. Physiol. 2012, 112, 1329–1334. [Google Scholar] [CrossRef] [Green Version]
- Flueck, M.; Vaughan, D.; Westerblad, H. Linking genes with exercise: Where is the cut-off? Eur. J. Appl. Physiol. 2010, 110, 1095–1098. [Google Scholar] [CrossRef] [Green Version]
- Baumert, P.; Lake, M.J.; Stewart, C.E.; Drust, B.; Erskine, R.M. Genetic variation and exercise-induced muscle damage: Implications for athletic performance, injury and ageing. Eur. J. Appl. Physiol. 2016, 116, 1595–1625. [Google Scholar] [CrossRef] [PubMed]
- Flück, M.; Ziemiecki, A.; Billeter, R.; Müntener, M. Fibre-type specific concentration of focal adhesion kinase at the sarcolemma: Influence of fibre innervation and regeneration. J. Exp. Biol. 2002, 205, 2337–2348. [Google Scholar] [CrossRef] [PubMed]
- Popper, K.R. Logik der Forschung; Mohr-Siebeck: Wien, Austria, 1969. [Google Scholar]
- Pasqua, L.A.; Bueno, S.; Matsuda, M.; Marquezini, M.V.; Lima-Silva, A.E.; Saldiva, P.H.; Bertuzzi, R. The genetics of human running: ACTN3 polymorphism as an evolutionary tool improving the energy economy during locomotion. Ann. Hum. Biol. 2016, 43, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Musiał, A.D.; Ropka-Molik, K.; Piórkowska, K.; Jaworska, J.; Stefaniuk-Szmukier, M. ACTN3 genotype distribution across horses representing different utility types and breeds. Mol. Biol. Rep. 2019, 46, 5795–5803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whipp, B.J.; Davis, J.A.; Torres, F.; Wasserman, K. A test to determine parameters of aerobic function during exercise. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1981, 50, 217–221. [Google Scholar] [CrossRef] [PubMed]
- Borg, G.A. Psychophysical bases of perceived exertion. Med. Sci. Sports Exerc. 1982, 14, 377–381. [Google Scholar] [CrossRef] [PubMed]
- Valdivieso, P.; Toigo, M.; Hoppeler, H.; Flück, M. T/T homozygosity of the tenascin-C gene polymorphism rs2104772 negatively influences exercise-induced angiogenesis. PLoS ONE 2017, 12, e0174864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaric, S.; Radosavljevic-Jaric, S.; Johansson, H. Muscle force and muscle torque in humans require different methods when adjusting for differences in body size. Eur. J. Appl. Physiol. 2002, 87, 304–307. [Google Scholar] [CrossRef]
- West, S.G.; Finch, J.F.; Curran, P.J. Structural equation models with nonnormal variables: Problems and remedies. In Structural Equation Modeling: Concepts, Issues and Applications; Hoyle, R.H., Ed.; Sage: Newbery Park, CA, USA, 1995; pp. 56–75. [Google Scholar]
- Gasser, B.; Frei, A.; Niederseer, D.; Catuogno, S.; Frey, W.O.; Flück, M. Variability in the Aerobic Fitness-Related Dependence on Respiratory Processes During Muscle Work Is Associated With the ACE-I/D Genotype. Front. Sports Act. Living 2022, 4, 814974. [Google Scholar] [CrossRef] [PubMed]
- Khalid, H. 2022. Available online: https://www.researchgate.net/profile/Khalid_Hassan10/post/Does_anyone_know_how_to_calculate_Hardy_Weinberg_Equilibrium_for_14_different_alleles/attachment/59d64ad979197b80779a50fd/AS:477303651213312@1490809466020/download/Court+lab+-+HW+calculator-+important.xls (accessed on 29 July 2022).
- Charbonneau, D.E.; Hanson, E.D.; Ludlow, A.T.; Delmonico, M.J.; Hurley, B.F.; Roth, S.M. ACE genotype and the muscle hypertrophic and strength responses to strength training. Med. Sci. Sports Exerc. 2008, 40, 677–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khanal, P.; Morse, C.I.; He, L.; Herbert, A.J.; Onambélé-Pearson, G.L.; Degens, H.; Thomis, M.; Williams, A.G.; Stebbings, G.K. Polygenic Models Partially Predict Muscle Size and Strength but Not Low Muscle Mass in Older Women. Genes 2022, 13, 982. [Google Scholar] [CrossRef] [PubMed]
- Berman, Y.; North, K.N. A Gene for Speed: The Emerging Role of alpha-Actinin-3 in Muscle Metabolism. Physiology 2010, 25, 250–259. [Google Scholar] [CrossRef] [PubMed]
- Nowak, K.J.; Wattanasirichaigoon, D.; Goebel, H.H.; Wilce, M.; Pelin, K.; Donner, K.; Jacob, R.L.; Hubner, C.; Oexle, K.; Anderson, J.R.; et al. Mutations in the skeletal muscle alpha-actin gene in patients with actin myopathy and nemaline myopathy. Nat. Genet. 1999, 23, 208–212. [Google Scholar] [CrossRef] [PubMed]
- MacArthur, D.G.; Seto, J.T.; Chan, S.; Quinlan, K.G.R.; Raftery, J.M.; Turner, N.; Nicholson, M.D.; Kee, A.J.; Hardeman, E.C.; Gunning, P.W.; et al. An Actn3 knockout mouse provides mechanistic insights into the association between alpha-actinin-3 deficiency and human athletic performance. Hum. Mol. Genet. 2008, 17, 1076–1086. [Google Scholar] [CrossRef] [Green Version]
- MacArthur, D.G.; Seto, J.T.; Raftery, J.M.; Quinlan, K.G.; Huttley, G.A.; Hook, J.W.; Lemckert, F.A.; Kee, A.J.; Edwards, M.R.; Berman, Y.; et al. Loss of ACTN3 gene function alters mouse muscle metabolism and shows evidence of positive selection in humans. Nat. Genet. 2007, 39, 1261–1265. [Google Scholar] [CrossRef] [PubMed]
- Eynon, N.; Duarte, J.; Oliveira, J.; Sagiv, M.; Yamin, C.; Meckel, Y.; Sagiv, M.; Goldhammer, E. ACTN3 R577X polymorphism and Israeli top-level athletes. Int. J. Sports Med. 2009, 30, 695–698. [Google Scholar] [CrossRef]
- Yang, N.; MacArthur, D.G.; Gulbin, J.P.; Hahn, A.G.; Beggs, A.H.; Easteal, S.; Yang, N.K.N.; MacArthur, D.G.; Gulbin, J.P.; Hahn, A.G.; et al. ACTN3 genotype is associated with human elite athletic performance. Am. J. Hum. Genet. 2003, 73, 627–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmetov, I.I.; Druzhevskaya, A.M.; Astratenkova, I.V.; Popov, D.V.; Vinogradova, O.L.; Rogozkin, V.A. The ACTN3 R577X polymorphism in Russian endurance athletes. Br. J. Sports Med. 2010, 44, 649–652. [Google Scholar] [CrossRef]
- Lucia, A.; Gomez-Gallego, F.; Santiago, C.; Bandres, F.; Earnest, C.; Rabadan, M.; Alonso, J.M.; Hoyos, J.; Cordova, A.; Villa, G.; et al. ACTN3 genotype in professional endurance cyclists. Int. J. Sports Med. 2006, 27, 880–884. [Google Scholar] [CrossRef]
- Niemi, A.K.; Majamaa, K. Mitochondrial DNA and ACTN3 genotypes in Finnish elite endurance and sprint athletes. Eur. J. Hum. Genet. 2005, 13, 965–969. [Google Scholar] [CrossRef]
- Paparini, A.; Ripani, M.; Giordano, G.D.; Santoni, D.; Pigozzi, F.; Romano-Spica, V. ACTN3 genotyping by real-time PCR in the Italian population and athletes. Med. Sci. Sports Exerc. 2007, 39, 810–815. [Google Scholar] [CrossRef]
- Saunders, C.J.; September, A.V.; Xenophontos, S.L.; Cariolou, M.A.; Anastassiades, L.C.; Noakes, T.D.; Collins, M. No association of the ACTN3 gene R577X polymorphism with endurance performance in Ironman Triathlons. Ann. Hum. Genet. 2007, 71, 777–781. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.; MacArthur, D.G.; Wolde, B.; Onywera, V.O.; Boit, M.K.; Lau, S.Y.; Wilson, R.H.; Scott, R.A.; Pitsiladis, Y.P.; North, K. The ACTN3 R577X polymorphism in East and West African athletes. Med. Sci. Sports Exerc. 2007, 39, 1985–1988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mägi, A.; Unt, E.; Prans, E.; Raus, L.; Eha, J. The Association Analysis between ACE and ACTN3 Genes Polymorphisms and Endurance Capacity in Young Cross-Country Skiers: Longitudinal Study. J. Sports Sci. Med. 2016, 15, 287–294. [Google Scholar]
- Orysiak, J.; Zmijewski, P.; Klusiewicz, A.; Kaliszewski, P.; Malczewska-Lenczowska, J.; Gajewski, J.; Pokrywka, A. The association between ACE Gene variation and aerobic capacity in winter endurance disciplines. Biol. Sport 2013, 30, 4. [Google Scholar] [CrossRef] [PubMed]
- Kaynar, O.; Bilic, M.F.; Sercan, C.; Ulucan, K. Determination of HIF1-Ars11549465 Polymorphism in Elite Skiers. Turk. J. Sport Exerc. 2019, 21, 328–331. [Google Scholar] [CrossRef]
- Däpp, C.; Gassmann, M.; Hoppeler, H.; Flück, M. Hypoxia-induced gene activity in disused oxidative muscle. Adv. Exp. Med. Biol. 2006, 588, 171–188. [Google Scholar] [CrossRef]
- Döring, F.; Onur, S.; Fischer, A.; Boulay, M.R.; Pérusse, L.; Rankinen, T.; Rauramaa, R.; Wolfarth, B.; Bouchard, C. A common haplotype and the Pro582Ser polymorphism of the hypoxia-inducible factor-1alpha (HIF1A) gene in elite endurance athletes. J. Appl. Physiol. 2010, 108, 1497–1500. [Google Scholar] [CrossRef]
- Stephen, M.R.; Sean, W.; Dongmei, L.E.; Jeffrey, M.; Luigi, F.; Ben, F.H. The ACTN3 R577X nonsense allele is under-represented in elite-level strength athletes. Eur. J. Hum. Genet. 2009, 16, 391–394. [Google Scholar]
- Coso, J.D.; Hiam, D.; Houweling, P.; Pérez, L.M.; Eynon, N.; Lucía, A. More than a „speed gene“: ACTN3 R577X genotype, trainability, muscle damage, and the risk for injuries. Eur. J. Appl. Physiol. 2019, 119, 49–60. [Google Scholar] [CrossRef]
- Bottura, R.M.; Lima, G.H.P.; Hipolide, D.C.; Pesquero, J.B. Association between ACTN3 and acute mountain sickness. Genes Environ. 2019, 41, 18. [Google Scholar] [CrossRef] [PubMed]
- Hoppeler, H.; Baum, O.; Mueller, M.; Lurman, G. Molekulare Mechanismen der Anpassungsfähigkeit der Skelettmuskulatur. Schweiz. Z. Sportmed. Sporttraumatol. 2011, 59, 6–13. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | ACE-I/D | Tenascin C | ACTN3 | PTK2 | PTK2 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SNP | rs1799752 | rs2104772 | rs1815739 | rs7843014 | rs7460 | ||||||||||
| n | percent | n | percent | n | percent | n | percent | n | percent | ||||||
| skiers | D/D | 8 | 18.6 | A/A | 16 | 37.2 | C/C | 12 | 27.9 | C/C | 7 | 16.3 | A/A | 15 | 34.9 |
| D/I | 24 | 55.8 | A/T | 23 | 53.5 | C/T | 6 | 14.0 | C/T | 20 | 46.5 | A/T | 23 | 53.5 | |
| I/I | 11 | 25.6 | T/T | 4 | 9.3 | T/T | 25 | 58.1 | T/T | 16 | 37.2 | T/T | 5 | 11.6 | |
| 43 | 100 | 43 | 100 | 43 | 100 | 43 | 100 | 43 | 100 | ||||||
| p-value | 0.425 | 0.294 | <0.0001 | 0.859 | 0.391 | ||||||||||
| n | percent | n | percent | n | percent | n | percent | n | percent | ||||||
| data points | D/D | 65 | 28.3 | A/A | 77 | 33.5 | C/C | 59 | 25.7 | C/C | 38 | 16.5 | A/A | 85 | 37.0 |
| D/I | 117 | 50.9 | A/T | 126 | 54.8 | C/T | 34 | 14.8 | C/T | 113 | 49.1 | A/T | 122 | 53.0 | |
| I/I | 48 | 20.9 | T/T | 27 | 11.7 | T/T | 137 | 59.6 | T/T | 79 | 34.3 | T/T | 23 | 10.0 | |
| 230 | 100 | 230 | 100 | 230 | 100 | 230 | 100 | 230 | 100 | ||||||
| p-value | 0.728 | 0.023 | <0.0001 | 0.822 | 0.029 |
| Parameter | Unit | Mean | SD | [Min–Max] | N | |
|---|---|---|---|---|---|---|
| age | [years] | 21.5 | ± | 3.02 | [16.22–31.58] | 230 |
| body mas | [kg] | [kg] | ± | 10.9 | [52.10–106.10] | 230 |
| body size | [cm] | 173.99 | ± | 8.69 | [155.00–202.00] | 230 |
| peakVO2 | [L(O2) min−1] | 3.69 | ± | 0.79 | [2.35–5.62] | 230 |
| peakVO2_scaled | [L(O2) min−1 kg−1] | 0.05 | ± | 0.01 | [0.03–0.08] | 230 |
| peakRER | [L(CO2) × L(O2)−1] | 1.52 | ± | 0.18 | [1.11–2.05] | 230 |
| peakP | [Watt] | 320.99 | ± | 61.74 | [190.00–460.00] | 230 |
| peakP_scaled | [Watt × kg−1] | 4.54 | ± | 0.68 | [2.48–6.54] | 230 |
| peakVE | [L min−1] | 145.54 | ± | 32.44 | [66.09–227.45] | 230 |
| peakVT | [L] | 2.65 | ± | 0.53 | [1.52–4.01] | 230 |
| peakHF | [beats min−1] | 194.03 | ± | 9.24 | [158.20–220.00] | 230 |
| max_P_ext_R360 | [Watt] | 697.94 | ± | 198.25 | [412.40–1349.90] | 141 |
| max_P_ext_L360 | [Watt] | 673.81 | ± | 169.75 | [374.10–1167.80] | 141 |
| max_P_flex_R360 | [Watt] | 464.15 | ± | 170.98 | [217.60–1117.70] | 141 |
| max_P_flex_L360 | [Watt] | 450.5 | ± | 148.9 | [226.80–938.90] | 141 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gasser, B.; Flück, M.; Frey, W.O.; Valdivieso, P.; Spörri, J. Association of Gene Variants for Mechanical and Metabolic Muscle Quality with Cardiorespiratory and Muscular Variables Related to Performance in Skiing Athletes. Genes 2022, 13, 1798. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13101798
Gasser B, Flück M, Frey WO, Valdivieso P, Spörri J. Association of Gene Variants for Mechanical and Metabolic Muscle Quality with Cardiorespiratory and Muscular Variables Related to Performance in Skiing Athletes. Genes. 2022; 13(10):1798. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13101798
Chicago/Turabian StyleGasser, Benedikt, Martin Flück, Walter O. Frey, Paola Valdivieso, and Jörg Spörri. 2022. "Association of Gene Variants for Mechanical and Metabolic Muscle Quality with Cardiorespiratory and Muscular Variables Related to Performance in Skiing Athletes" Genes 13, no. 10: 1798. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13101798