
Genome Wide Association Study with Imputed Whole Genome Sequence Data Identifies a 431 kb Risk Haplotype on CFA18 for Congenital Laryngeal Paralysis in Alaskan Sled Dogs
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Phenotype Assignment
2.2. DNA Isolation and Genotyping
2.3. Imputation
2.4. Genome-Wide Association Study
2.5. Gene-Based Association Study
2.6. Whole Genome Sequence Analysis
3. Results
3.1. Phenotype
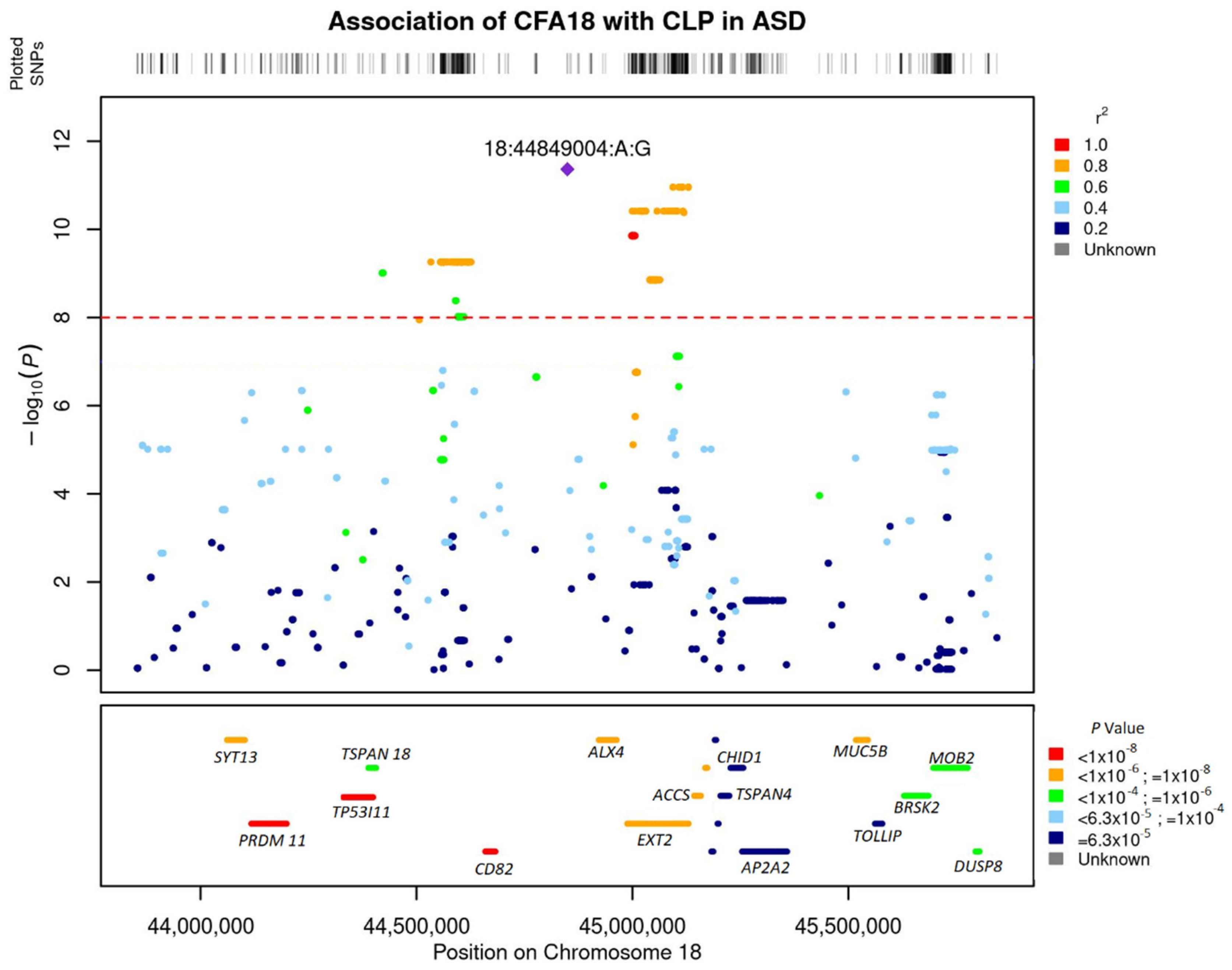
3.2. Genome-Wide Association Study
3.3. Linkage Disequilibrium and Associated Haplotypes
3.4. Annotation of Significant SNPs to Genes and Gene-Based Association Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huson, H.J.; Parker, H.G.; Runstadler, J.; Ostrander, E.A. A genetic dissection of breed composition and performance enhancement in the Alaskan sled dog. BMC Genet. 2010, 11, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huson, H.J.; Vonholdt, B.M.; Rimbault, M.; Byers, A.M.; Runstadler, J.A.; Parker, H.G.; Ostrander, E.A. Breed-specific ancestry studies and genome-wide association analysis highlight an association between the MYH9 gene and heat tolerance in Alaskan sprint racing sled dogs. Mamm. Genome 2012, 23, 178–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tosi, I.; Art, T.; Boemer, F.; Votion, D.-M.; Davis, M.S. Acylcarnitine profile in Alaskan sled dogs during submaximal multiday exercise points out metabolic flexibility and liver role in energy metabolism. PLoS ONE 2021, 16, e0256009. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; Hendriks, J. Inherited laryngeal paralysis. Analysis in the husky cross. Vet. Q. 1986, 8, 301–302. [Google Scholar] [CrossRef]
- Von Pfeil, D.J.; Zellner, E.; Fritz, M.C.; Langohr, I.; Griffitts, C.; Stanley, B.J. Congenital laryngeal paralysis in Alaskan Huskies: 25 cases (2009–2014). J. Am. Vet. Med. Assoc. 2018, 253, 1057–1065. [Google Scholar] [CrossRef]
- Stanley, B.J.; Hauptman, J.G.; Fritz, M.C.; Rosenstein, D.S.; Kinns, J. Esophageal dysfunction in dogs with idiopathic laryngeal paralysis: A controlled cohort study. Vet. Surg. 2010, 39, 139–149. [Google Scholar] [CrossRef]
- Polizopoulou, Z.; Koutinas, A.; Papadopoulos, G.; Saridomichelakis, M. Juvenile laryngeal paralysis in three Siberian husky x Alaskan malamute puppies. Vet. Rec. 2003, 153, 624–627. [Google Scholar] [CrossRef]
- Ubbink, G.; Knol, B.; Bouw, J. The relationship between homozygosity and the occurrence of specific diseases in Bouvier Belge des Flandres dogs in the Netherlands: Inbreeding and disease in the bouvier dog. Vet. Q. 1992, 14, 137–140. [Google Scholar] [CrossRef]
- Venker-van Haagen, A.; Bouw, J.; Hartman, W. Hereditary transmission of laryngeal paralysis in Bouviers. J. Am. Anim. Hosp. Assoc. 1981, 17, 75–76. [Google Scholar]
- Camargo, J.F.; Teixeira, L.G.; Trindade-Gerardi, A.B.; Dos Santos, B.S.; da Rosa, M.P.; Paim, M.G.; Contesini, E.A. Laryngeal Paralysis Following Endotracheal Intubation in a Dog: A Case Report. Top. Companion Anim. Med. 2022, 48, 100635. [Google Scholar] [CrossRef]
- MacPhail, C. Laryngeal disease in dogs and cats. Vet. Clin. Small Anim. Pract. 2014, 44, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Letko, A.; Minor, K.M.; Friedenberg, S.G.; Shelton, G.D.; Salvador, J.P.; Mandigers, P.J.; Leegwater, P.A.; Winkler, P.A.; Petersen-Jones, S.M.; Stanley, B.J. A CNTNAP1 Missense Variant Is Associated with Canine Laryngeal Paralysis and Polyneuropathy. Genes 2020, 11, 1426. [Google Scholar] [CrossRef] [PubMed]
- Hadji Rasouliha, S.; Barrientos, L.; Anderegg, L.; Klesty, C.; Lorenz, J.; Chevallier, L.; Jagannathan, V.; Rösch, S.; Leeb, T. A RAPGEF6 variant constitutes a major risk factor for laryngeal paralysis in dogs. PLoS Genet. 2019, 15, e1008416. [Google Scholar] [CrossRef] [PubMed]
- Broome, C.; Burbidge, H.; Pfeiffer, D. Prevalence of laryngeal paresis in dogs undergoing general anaesthesia. Aust. Vet. J. 2000, 78, 769–772. [Google Scholar] [CrossRef] [PubMed]
- Tobias, K.M.; Jackson, A.M.; Harvey, R.C. Effects of doxapram HCl on laryngeal function of normal dogs and dogs with naturally occurring laryngeal paralysis. Vet. Anaesth. Analg. 2004, 31, 258–263. [Google Scholar] [CrossRef]
- Monnet, E. Surgical treatment of laryngeal paralysis. Vet. Clin. Small Anim. Pract. 2016, 46, 709–717. [Google Scholar] [CrossRef]
- von Pfeil, D.J.; Edwards, M.R.; Déjardin, L.M. Less invasive unilateral arytenoid lateralization: A modified technique for treatment of idiopathic laryngeal paralysis in dogs: Technique description and outcome. Vet. Surg. 2014, 43, 704–711. [Google Scholar] [CrossRef]
- Moe, L.; Bjerkås, I. Hereditary polyneuropathy of alaskan malamutes. In Kirks Current Veterinary Therapy XI; Saunders: Philadelphia, PA, USA, 1992; pp. 1038–1039. [Google Scholar]
- Braund, K.; Steinberg, H.; Shores, A.; Steiss, J.; Mehta, J.; Toivio-Kinnucan, M.; Amling, K. Laryngeal paralysis in immature and mature dogs as one sign of a more diffuse polyneuropathy. J. Am. Vet. Med. Assoc. 1989, 194, 1735–1740. [Google Scholar]
- Venker van Haagen, A.; Hartman, W.; Goedegebuure, S. Spontaneous laryngeal paralysis in young bouviers [Dogs]. J. Am. Anim. Hosp. Assoc. 1978, 14, 714–720. [Google Scholar]
- Harvey, C.; O’Brien, J. Upper airway obstruction surgery. 5. Treatment of laryngeal paralysis in dogs by partial laryngectomy. J. Am. Anim. Hosp. Assoc. 1982, 18, 551–556. [Google Scholar]
- Shelton, G.D.; Podell, M.; Poncelet, L.; Schatzberg, S.; Patterson, E.; Powell, H.C.; Mizisin, A.P. Inherited polyneuropathy in Leonberger dogs: A mixed or intermediate form of Charcot-Marie-Tooth disease? Muscle Nerve Off. J. Am. Assoc. Electrodiagn. Med. 2003, 27, 471–477. [Google Scholar] [CrossRef] [PubMed]
- Shores, A.; Cochrané ihrFFjâte, S. Laryngeal paralysis-polyneuropathy complex in young Dalmatians. Am. J. Vet. Res. 1994, 55, 534–542. [Google Scholar]
- Gabriel, A.; Poncelet, L.; Van Ham, L.; Clercx, C.; Braund, K.; Bhatti, S.; Detilleux, J.; Peeters, D. Laryngeal paralysis-polyneuropathy complex in young related Pyrenean mountain dogs. J. Small Anim. Pract. 2006, 47, 144–149. [Google Scholar] [CrossRef]
- Mahony, O.M.; Knowles, K.E.; Braund, K.G.; Averill, D.R., Jr.; Frimberger, A.E. Laryngeal paralysis-polyneuropathy complex in young Rottweilers. J. Vet. Intern. Med. 1998, 12, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Salvadori, C.; Tartarelli, C.; Baroni, M.; Mizisin, A.; Cantile, C. Peripheral nerve pathology in two Rottweilers with neuronal vacuolation and spinocerebellar degeneration. Vet. Pathol. 2005, 42, 852–855. [Google Scholar] [CrossRef] [PubMed]
- McKeirnan, K.L.; Gross, M.E.; Rochat, M.; Payton, M. Comparison of propofol and propofol/ketamine anesthesia for evaluation of laryngeal function in healthy dogs. J. Am. Anim. Hosp. Assoc. 2014, 50, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Becker, D.; Minor, K.M.; Letko, A.; Ekenstedt, K.J.; Jagannathan, V.; Leeb, T.; Shelton, G.D.; Mickelson, J.R.; Drögemüller, C. A GJA9 frameshift variant is associated with polyneuropathy in Leonberger dogs. BMC Genom. 2017, 18, 662. [Google Scholar] [CrossRef] [Green Version]
- Ekenstedt, K.J.; Becker, D.; Minor, K.M.; Shelton, G.D.; Patterson, E.E.; Bley, T.; Oevermann, A.; Bilzer, T.; Leeb, T.; Drögemüller, C. An ARHGEF10 deletion is highly associated with a juvenile-onset inherited polyneuropathy in Leonberger and Saint Bernard dogs. PLoS Genet. 2014, 10, e1004635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiedmer, M.; Oevermann, A.; Borer-Germann, S.E.; Gorgas, D.; Shelton, G.D.; Drögemüller, M.; Jagannathan, V.; Henke, D.; Leeb, T. A RAB3GAP1 SINE insertion in Alaskan huskies with polyneuropathy, ocular abnormalities, and neuronal vacuolation (POANV) resembling human Warburg micro syndrome 1 (WARBM1). G3 Genes Genomes Genet. 2016, 6, 255–262. [Google Scholar] [CrossRef] [Green Version]
- Bruun, C.S.; Jäderlund, K.H.; Berendt, M.; Jensen, K.B.; Spodsberg, E.H.; Gredal, H.; Shelton, G.D.; Mickelson, J.R.; Minor, K.M.; Lohi, H. A Gly98Val mutation in the N-Myc downstream regulated gene 1 (NDRG1) in Alaskan Malamutes with polyneuropathy. PLoS ONE 2013, 8, e54547. [Google Scholar] [CrossRef] [Green Version]
- Maurya, R.; Kumar, B.; Sundar, S. Evaluation of salt-out method for the isolation of DNA from whole blood: A pathological approach of DNA based diagnosis. Int. J. Life Sci. Biotechnol. Pharma. Res. 2013, 2, 53–57. [Google Scholar]
- Deane-Coe, P.E.; Chu, E.T.; Slavney, A.; Boyko, A.R.; Sams, A.J. Direct-to-consumer DNA testing of 6,000 dogs reveals 98.6-kb duplication associated with blue eyes and heterochromia in Siberian Huskies. PLoS Genet. 2018, 14, e1007648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.C.; Chow, C.C.; Tellier, L.C.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-generation PLINK: Rising to the challenge of larger and richer datasets. GigaScience 2015, 4, s13742-015. [Google Scholar] [CrossRef]
- Das, S.; Forer, L.; Schönherr, S.; Sidore, C.; Locke, A.E.; Kwong, A.; Vrieze, S.I.; Chew, E.Y.; Levy, S.; McGue, M. Next-generation genotype imputation service and methods. Nat. Genet. 2016, 48, 1284–1287. [Google Scholar] [CrossRef] [Green Version]
- Plassais, J.; Kim, J.; Davis, B.W.; Karyadi, D.M.; Hogan, A.N.; Harris, A.C.; Decker, B.; Parker, H.G.; Ostrander, E.A. Whole genome sequencing of canids reveals genomic regions under selection and variants influencing morphology. Nat. Commun. 2019, 10, 1489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Browning, B.L.; Zhou, Y.; Browning, S.R. A one-penny imputed genome from next-generation reference panels. Am. J. Hum. Genet. 2018, 103, 338–348. [Google Scholar] [CrossRef]
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; DePristo, M.A.; Handsaker, R.E.; Lunter, G.; Marth, G.T.; Sherry, S.T. The variant call format and VCFtools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef] [PubMed]
- Ramnarine, S.; Zhang, J.; Chen, L.-S.; Culverhouse, R.; Duan, W.; Hancock, D.B.; Hartz, S.M.; Johnson, E.O.; Olfson, E.; Schwantes-An, T.-H. When does choice of accuracy measure alter imputation accuracy assessments? PLoS ONE 2015, 10, e0137601. [Google Scholar] [CrossRef]
- Zerbino, D.R.; Achuthan, P.; Akanni, W.; Amode, M.R.; Barrell, D.; Bhai, J.; Billis, K.; Cummins, C.; Gall, A.; Girón, C.G. Ensembl 2018. Nucleic Acids Res. 2018, 46, D754–D761. [Google Scholar] [CrossRef]
- Yang, J.; Lee, S.H.; Goddard, M.E.; Visscher, P.M. GCTA: A tool for genome-wide complex trait analysis. Am. J. Hum. Genet. 2011, 88, 76–82. [Google Scholar] [CrossRef] [Green Version]
- Srikanth, K.; Lee, S.-H.; Chung, K.-Y.; Park, J.-E.; Jang, G.-W.; Park, M.-R.; Kim, N.Y.; Kim, T.-H.; Chai, H.-H.; Park, W.C. A gene-set enrichment and protein–protein interaction network-based GWAS with regulatory SNPs identifies candidate genes and pathways associated with carcass traits in hanwoo cattle. Genes 2020, 11, 316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, L.; Zhang, H.; Tang, Z.; Xu, J.; Yin, D.; Zhang, Z.; Yuan, X.; Zhu, M.; Zhao, S.; Li, X. rMVP: A memory-efficient, visualization-enhanced, and parallel-accelerated tool for genome-wide association study. Genom. Proteom. Bioinform. 2021, 19, 619–628. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: http://www.R-project.org/ (accessed on 17 April 2022).
- De Leeuw, C.A.; Mooij, J.M.; Heskes, T.; Posthuma, D. MAGMA: Generalized gene-set analysis of GWAS data. PLoS Comput. Biol. 2015, 11, e1004219. [Google Scholar] [CrossRef] [PubMed]
- Bakshi, A.; Zhu, Z.; Vinkhuyzen, A.A.; Hill, W.D.; McRae, A.F.; Visscher, P.M.; Yang, J. Fast set-based association analysis using summary data from GWAS identifies novel gene loci for human complex traits. Sci. Rep. 2016, 6, 32894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Srikanth, K.; Kim, N.-Y.; Park, W.; Kim, J.-M.; Kim, K.-D.; Lee, K.-T.; Son, J.-H.; Chai, H.-H.; Choi, J.-W.; Jang, G.-W. Comprehensive genome and transcriptome analyses reveal genetic relationship, selection signature, and transcriptome landscape of small-sized Korean native Jeju horse. Sci. Rep. 2019, 9, 16672. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows–Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The sequence alignment/map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Van der Auwera, G.A.; Carneiro, M.O.; Hartl, C.; Poplin, R.; Del Angel, G.; Levy-Moonshine, A.; Jordan, T.; Shakir, K.; Roazen, D.; Thibault, J. From FastQ data to high-confidence variant calls: The genome analysis toolkit best practices pipeline. Curr. Protoc. Bioinform. 2013, 43, 11.10.1–11.10.33. [Google Scholar]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef] [Green Version]
- Robinson, J.T.; Thorvaldsdóttir, H.; Winckler, W.; Guttman, M.; Lander, E.S.; Getz, G.; Mesirov, J.P. Integrative genomics viewer. Nat. Biotechnol. 2011, 29, 24–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallarino, R.; Henegar, C.; Mirasierra, M.; Manceau, M.; Schradin, C.; Vallejo, M.; Beronja, S.; Barsh, G.S.; Hoekstra, H.E. Developmental mechanisms of stripe patterns in rodents. Nature 2016, 539, 518–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beverdam, A.; Brouwer, A.; Reijnen, M.; Korving, J.; Meijlink, F. Severe nasal clefting and abnormal embryonic apoptosis in Alx3/Alx4 double mutant mice. Development 2001, 128, 3975–3986. [Google Scholar] [CrossRef]
- Curtain, M.; Heffner, C.S.; Maddox, D.M.; Gudis, P.; Donahue, L.R.; Murray, S.A. A novel allele of Alx4 results in reduced Fgf10 expression and failure of eyelid fusion in mice. Mamm. Genome 2015, 26, 173–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romeike, B.F.; Shen, Y.; Nishimoto, H.K.; Morton, C.C.; Layman, L.C.; Kim, H.-G. Spectrum of genes involved in a unique case of Potocki Schaffer syndrome with a large chromosome 11 deletion. Clin. Neuropathol. 2014, 33, 238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyagi, R.; Shenoy, A.R.; Visweswariah, S.S. Characterization of an evolutionarily conserved metallophosphoesterase that is expressed in the fetal brain and associated with the WAGR syndrome. J. Biol. Chem. 2009, 284, 5217–5228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brémond-Gignac, D.; Crolla, J.A.; Copin, H.; Guichet, A.; Bonneau, D.; Taine, L.; Lacombe, D.; Baumann, C.; Benzacken, B.; Verloes, A. Combination of WAGR and Potocki–Shaffer contiguous deletion syndromes in a patient with an 11p11. 2–p14 deletion. Eur. J. Hum. Genet. 2005, 13, 409–413. [Google Scholar] [CrossRef]
- Liu, J.; Li, M.; Su, B. GWAS-identified schizophrenia risk SNPs at TSPAN18 are highly diverged between Europeans and East Asians. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2016, 171, 1032–1040. [Google Scholar] [CrossRef]
- Pannier, S.; Legeai-Mallet, L. Hereditary multiple exostoses and enchondromatosis. Best Pract. Res. Clin. Rheumatol. 2008, 22, 45–54. [Google Scholar] [CrossRef]
- Shokraeian, P.; Daneshmandpour, Y.; Jamshidi, J.; Emamalizadeh, B.; Darvish, H. Genetic analysis of rs11038167, rs11038172 and rs835784 polymorphisms of the TSPAN18 gene in Iranian schizophrenia patients. Meta Gene 2019, 22, 100609. [Google Scholar] [CrossRef]
- Garay, P.M.; Wallner, M.A.; Iwase, S. Yin–yang actions of histone methylation regulatory complexes in the brain. Epigenomics 2016, 8, 1689–1708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, J.-Y.; Aromolaran, K.A.; Zukin, R.S. The emerging field of epigenetics in neurodegeneration and neuroprotection. Nat. Rev. Neurosci. 2017, 18, 347–361. [Google Scholar] [CrossRef] [PubMed]
- Südhof, T.C. Synaptotagmins: Why so many? J. Biol. Chem. 2002, 277, 7629–7632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nizzardo, M.; Taiana, M.; Rizzo, F.; Aguila Benitez, J.; Nijssen, J.; Allodi, I.; Melzi, V.; Bresolin, N.; Comi, G.; Hedlund, E. Synaptotagmin 13 is neuroprotective across motor neuron diseases. Acta Neuropathol. 2020, 139, 837–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abad-Rodríguez, J.; Díez-Revuelta, N. Axon glycoprotein routing in nerve polarity, function, and repair. Trends Biochem. Sci. 2015, 40, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Esko, J.D.; Selleck, S.B. Order out of chaos: Assembly of ligand binding sites in heparan sulfate. Annu. Rev. Biochem. 2002, 71, 435–471. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Putative Risk Haplotype (CFA 18:44502007–45132398) | ||||
|---|---|---|---|---|
| CLP Status | N | Homozygous Alternative | Heterozygous | Homozygous Risk |
| Affected | 20 | 2/20 | 0/20 | 18/20 |
| Unaffected | 205 | 9/40 | 21/40 | 0/40 |
| Term/Pathway | Q-Value 1 | Genes |
|---|---|---|
| Cholinergic synapse | 0.002 | GNG4, CREB3L1, KCNQ1 |
| Glycosaminoglycan biosynthesis | 0.010 | EXT2, CHST1 |
| Glycerolipid metabolism | 0.010 | DGKZ, PNPLA2 |
| Glycoprotein | 0.010 | EXT2, CD82, CREB3L1, TSPAN18, PNPLA2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Srikanth, K.; von Pfeil, D.J.F.; Stanley, B.J.; Griffitts, C.; Huson, H.J. Genome Wide Association Study with Imputed Whole Genome Sequence Data Identifies a 431 kb Risk Haplotype on CFA18 for Congenital Laryngeal Paralysis in Alaskan Sled Dogs. Genes 2022, 13, 1808. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13101808
Srikanth K, von Pfeil DJF, Stanley BJ, Griffitts C, Huson HJ. Genome Wide Association Study with Imputed Whole Genome Sequence Data Identifies a 431 kb Risk Haplotype on CFA18 for Congenital Laryngeal Paralysis in Alaskan Sled Dogs. Genes. 2022; 13(10):1808. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13101808
Chicago/Turabian StyleSrikanth, Krishnamoorthy, Dirsko J. F. von Pfeil, Bryden J. Stanley, Caroline Griffitts, and Heather J. Huson. 2022. "Genome Wide Association Study with Imputed Whole Genome Sequence Data Identifies a 431 kb Risk Haplotype on CFA18 for Congenital Laryngeal Paralysis in Alaskan Sled Dogs" Genes 13, no. 10: 1808. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13101808