
Initiator-Directed Transcription: Fission Yeast Nmtl Initiator Directs Preinitiation Complex Formation and Transcriptional Initiation

 ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Purification of GTFs, PC4, RNAPII and Mediator
2.2. Mediator and TAF Depletion
2.3. Electrophoretic Mobility Shift Assays
2.4. In Vitro Transcription Assays
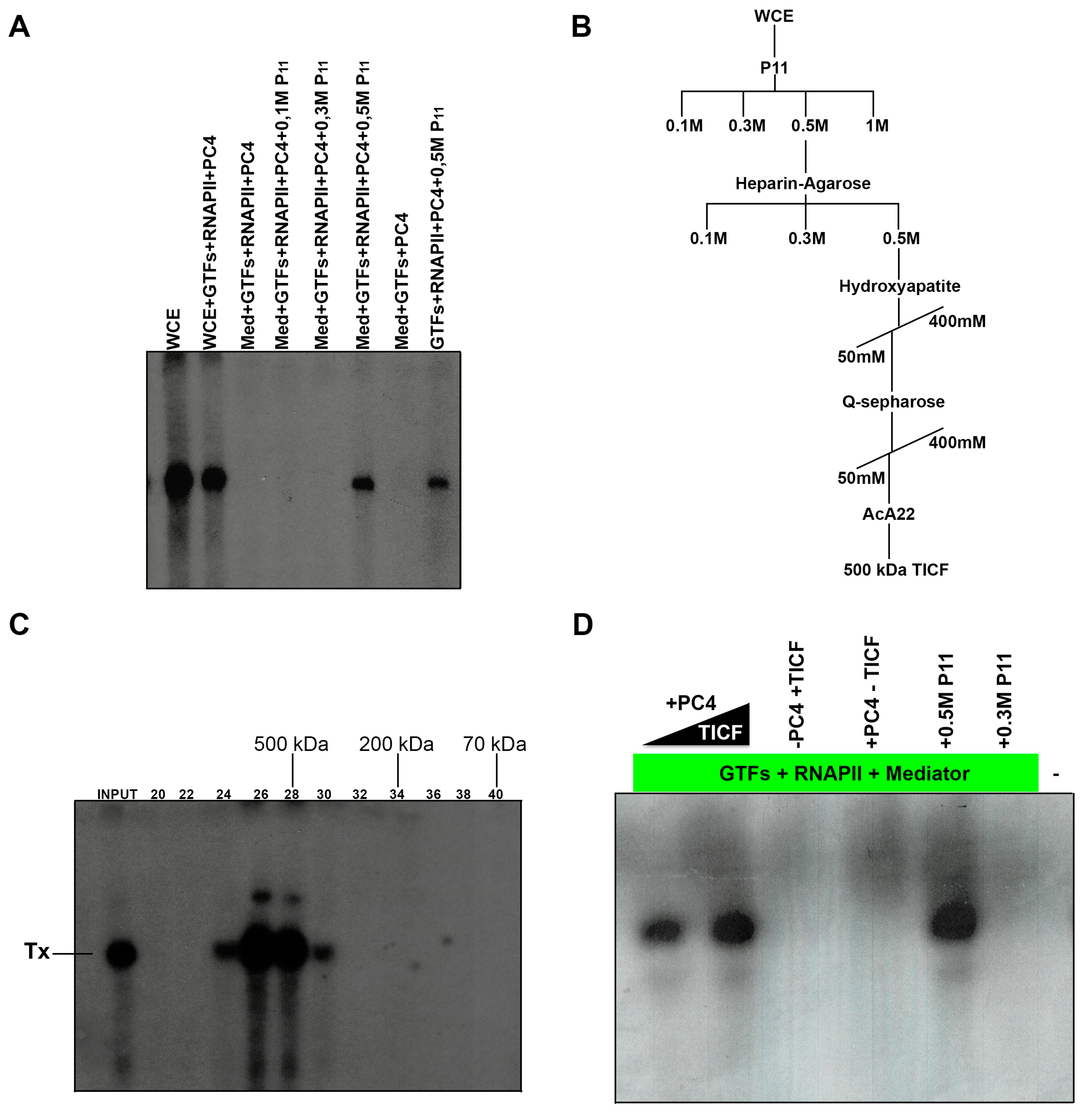
2.5. Purification of TICF
3. Results
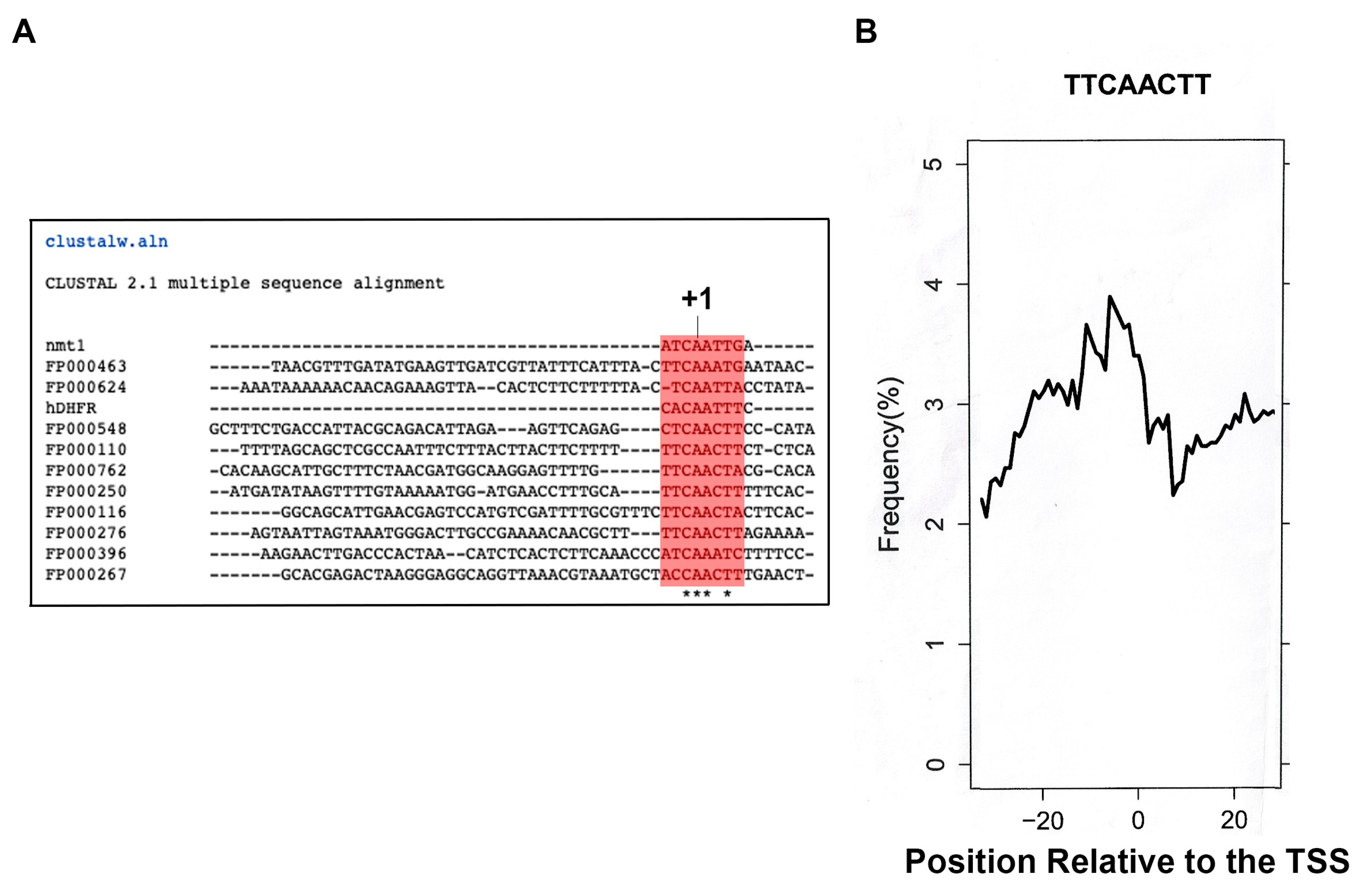
3.1. The Transcription Start Point of the Nmtl Promoter Has Homology to the Human Dihydrofolate Reductase Inr
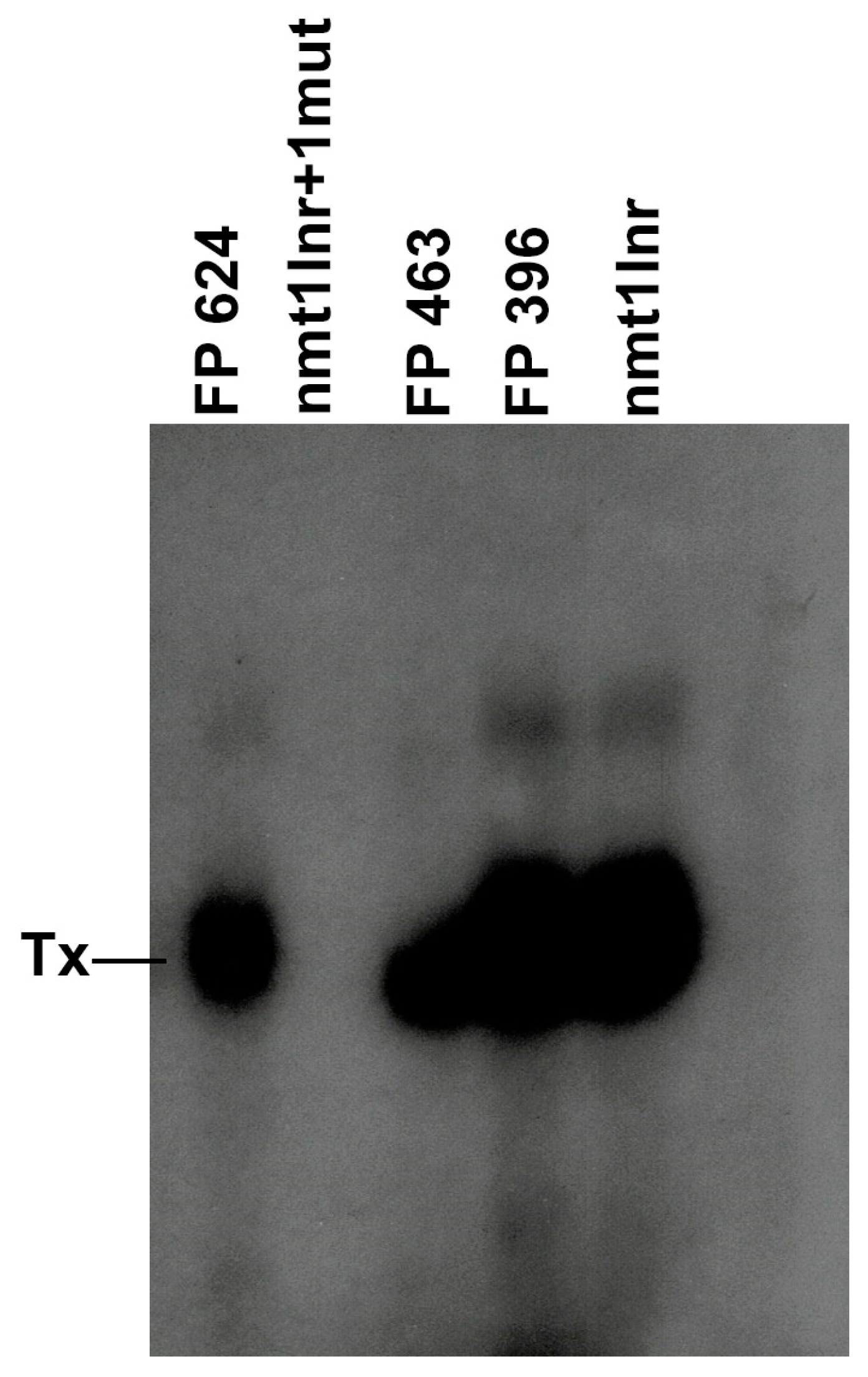
3.2. A TATA Box Mutated Nmtl Promoter Is Transcriptionally Active in S. pombe Whole-Cell Extracts
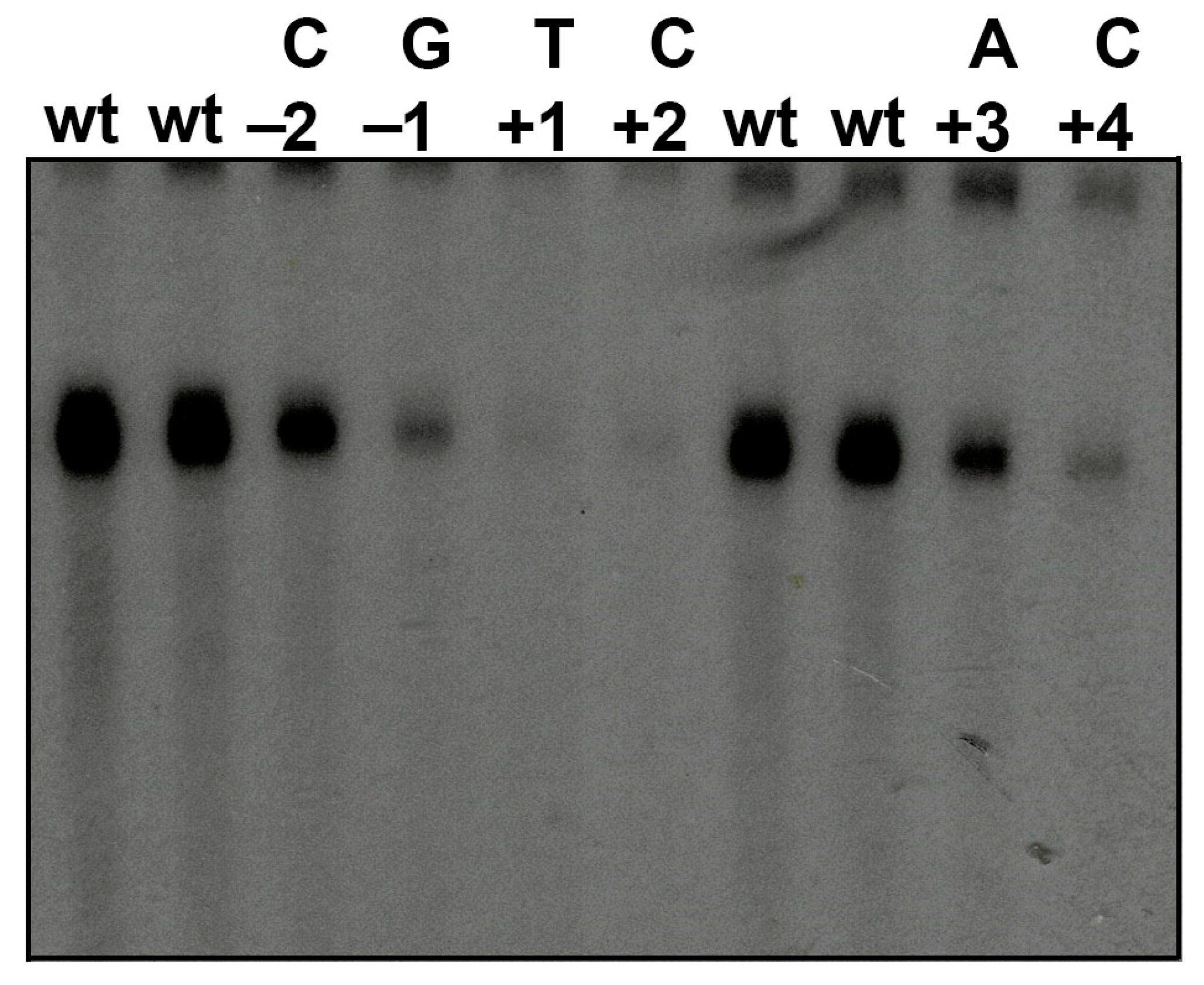
3.3. Mutations around the TSS of nmt1 Promoter Can Impair Transcriptional Initiation
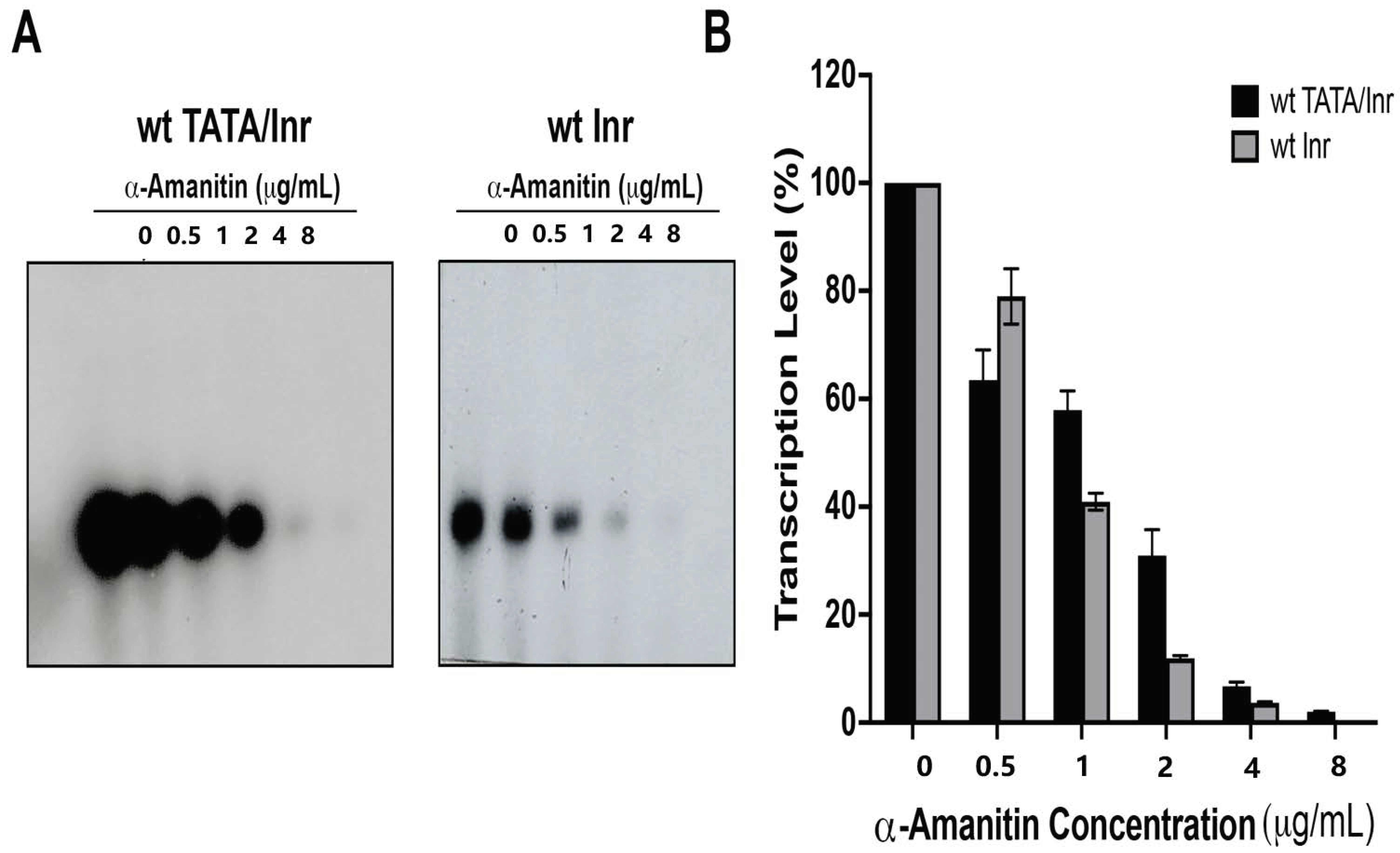
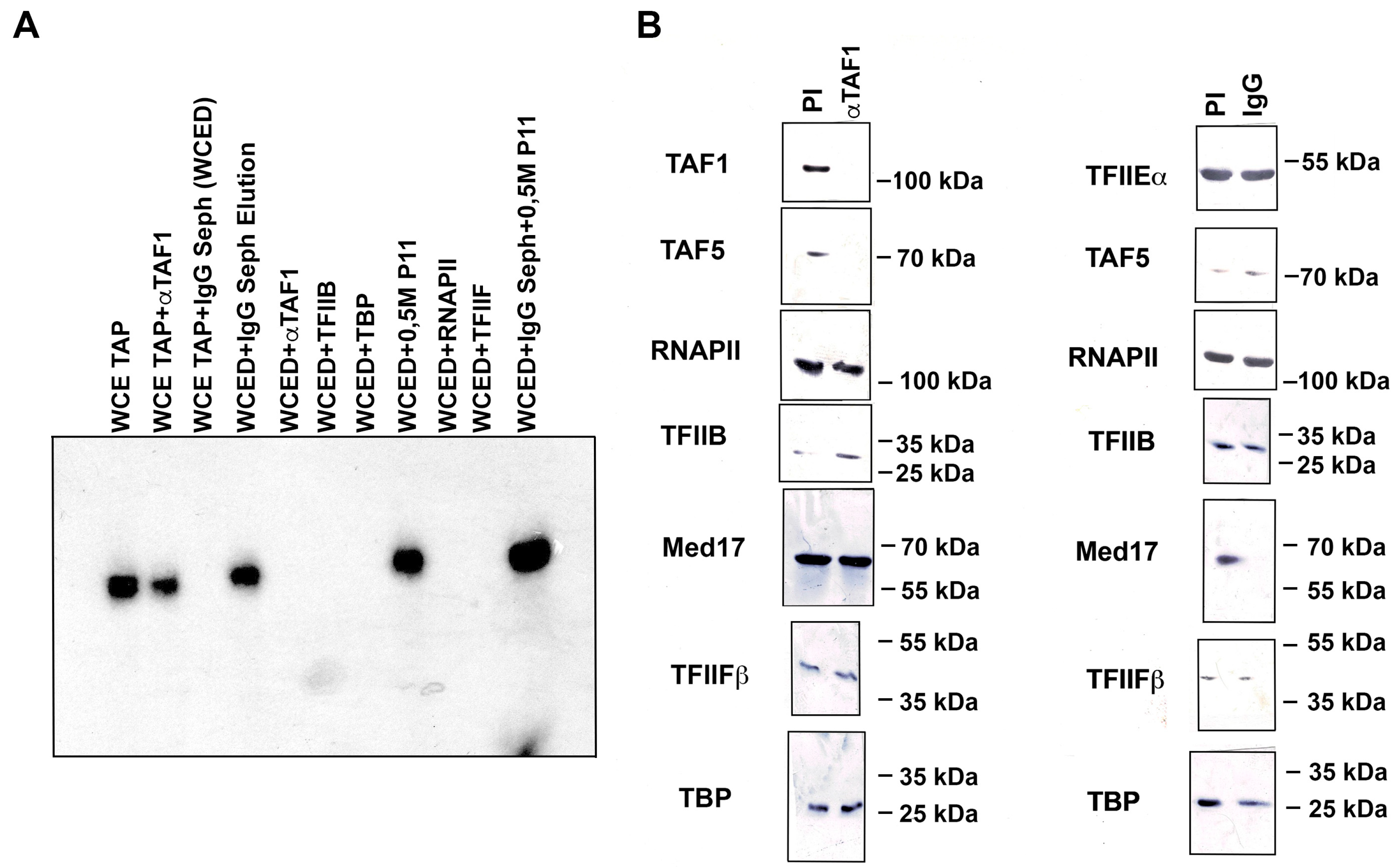
3.4. Nmtl Inr-Directed Transcription Initiation Is Mediator-Dependent but Is TAF-Independent
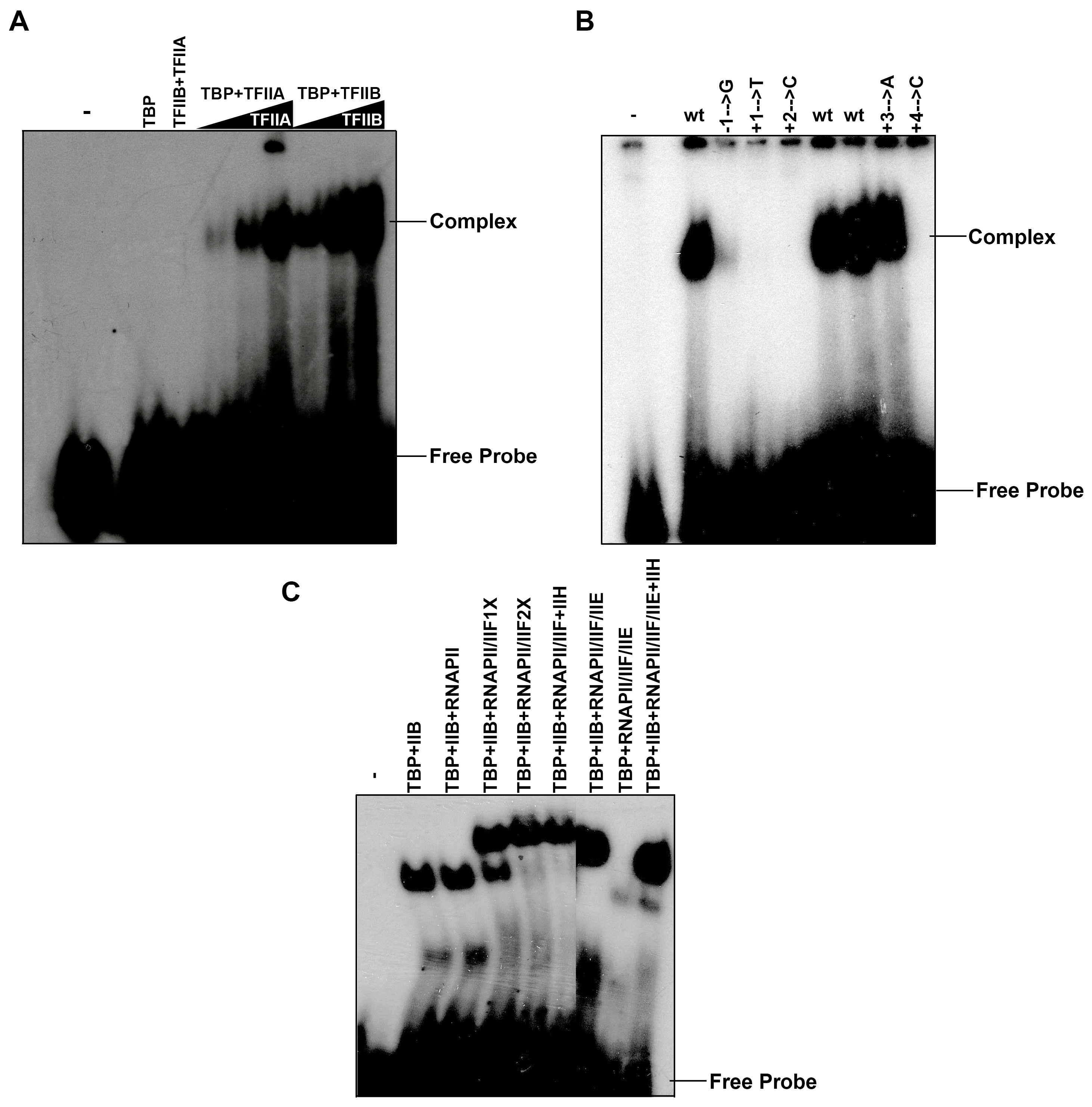
3.5. A Dimeric Complex between TFIIA-TFIIB or TBP-TFIIB Recognizes the Nmtl Inr
3.6. In Vitro Transcription Reconstitution from the Nmtl Inr Promoter
3.7. TATA-Lacking Promoters with a Promoter of the nmt1 Inr/hDHFR Family Can Direct Transcription in Fission Yeast WCE
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Struhl, K. Promoters, activator proteins, and the mechanism of transcriptional initiation in yeast. Cell 1987, 49, 295–297. [Google Scholar] [CrossRef]
- Roeder, R.G. The complexities of eukaryotic transcription initiation: Regulation of preinitiation complex assembly. Trends Biochem. Sci. 1991, 16, 402–408. [Google Scholar] [CrossRef]
- Juven-Gershon, T.; Hsu, J.Y.; Theisen, J.W.; Kadonaga, J.T. The RNA polymerase II core promoter—The gateway to transcription. Curr. Opin. Cell Biol. 2008, 20, 253–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vo Ngoc, L.; Wang, Y.L.; Kassavetis, G.A.; Kadonaga, J.T. The punctilious RNA polymerase II core promoter. Genes Dev. 2017, 31, 1289–1301. [Google Scholar] [CrossRef] [Green Version]
- Urbina, F.; Morales-Pison, S.; Maldonado, E. Enzymatic Protein Biopolymers as a Tool to Synthetize Eukaryotic Messenger Ribonucleic Acid (mRNA) with Uses in Vaccination, Immunotherapy and Nanotechnology. Polymers 2020, 12, 1633. [Google Scholar] [CrossRef]
- Smale, S.T.; Baltimore, D. The “initiator” as a transcription control element. Cell 1989, 57, 103–113. [Google Scholar] [CrossRef]
- Javahery, R.; Khachi, A.; Lo, K.; Zenzie-Gregory, B.; Smale, S.T. DNA sequence requirements for transcriptional initiator activity in mammalian cells. Mol. Cell Biol. 1994, 14, 116–127. [Google Scholar] [CrossRef]
- Smale, S.T. Transcription initiation from TATA-less promoters within eukaryotic protein-coding genes. Biochim. Biophys. Acta 1997, 1351, 73–88. [Google Scholar] [CrossRef]
- Kaufmann, J.; Smale, S.T. Direct recognition of initiator elements by a component of the transcription factor IID complex. Genes Dev. 1994, 8, 821–829. [Google Scholar] [CrossRef] [Green Version]
- Purnell, B.A.; Emanuel, P.A.; Gilmour, D.S. TFIID sequence recognition of the initiator and sequences farther downstream in Drosophila class II genes. Genes Dev. 1994, 8, 830–842. [Google Scholar] [CrossRef] [Green Version]
- Martinez, E.; Chiang, C.M.; Ge, H.; Roeder, R.G. TATA-binding protein-associated factor(s) in TFIID function through the initiator to direct basal transcription from a TATA-less class II promoter. EMBO J. 1994, 13, 3115–3126. [Google Scholar] [CrossRef] [PubMed]
- Kugel, J.F.; Goodrich, J.A. Finding the start site: Redefining the human initiator element. Genes Dev. 2017, 31, 1–2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carninci, P.; Sandelin, A.; Lenhard, B.; Katayama, S.; Shimokawa, K.; Ponjavic, J.; Semple, C.A.; Taylor, M.S.; Engström, P.G.; Frith, M.C.; et al. Genome-wide analysis of mammalian promoter architecture and evolution. Nat. Genet. 2006, 38, 626–635. [Google Scholar] [CrossRef] [PubMed]
- Vo Ngoc, L.; Cassidy, C.J.; Huang, C.Y.; Duttke, S.H.; Kadonaga, J.T. The human initiator is a distinct and abundant element that is precisely positioned in focused core promoters. Genes Dev. 2017, 31, 6–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Hou, J.; Bai, L.; Hu, C.; Tong, P.; Kang, Y.; Zhao, X.; Shao, Z. Genome-wide analysis of core promoter structures in Schizosaccharomyces pombe with DeepCAGE. RNA Biol. 2015, 12, 525–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maundrell, K. nmt1 of fission yeast. A highly transcribed gene completely repressed by thiamine. J. Biol. Chem. 1990, 265, 10857–10864. [Google Scholar] [CrossRef]
- Basi, G.; Schmid, E.; Maundrell, K. TATA box mutations in the Schizosaccharomyces pombe nmt1 promoter affect transcription efficiency but not the transcription start point or thiamine repressibility. Gene 1993, 123, 131–136. [Google Scholar] [CrossRef]
- Maundrell, K. Thiamine-repressible expression vectors pREP and pRIP for fission yeast. Gene 1993, 123, 127–130. [Google Scholar] [CrossRef]
- Montes, M.; Moreira-Ramos, S.; Rojas, D.A.; Urbina, F.; Käufer, N.F.; Maldonado, E. RNA polymerase II components and Rrn7 form a preinitiation complex on the HomolD box to promote ribosomal protein gene expression in Schizosaccharomyces pombe. FEBS J. 2017, 284, 615–633. [Google Scholar] [CrossRef]
- Contreras-Levicoy, J.; Urbina, F.; Maldonado, E. Schizosaccharomyces pombe positive cofactor 4 stimulates basal transcription from TATA-containing and TATA-less promoters through Mediator and transcription factor IIA. FEBS J. 2008, 275, 2873–2883. [Google Scholar] [CrossRef]
- Slansky, J.E.; Farnham, P.J. Transcriptional regulation of the dihydrofolate reductase gene. BioEssays 1996, 18, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Schier, A.C.; Taatjes, D.J. Structure and mechanism of the RNA polymerase II transcription machinery. Genes Dev. 2020, 34, 465–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parvin, J.D.; Sharp, P.A. DNA topology and a minimal set of basal factors for transcription by RNA polymerase II. Cell 1993, 73, 533–540. [Google Scholar] [CrossRef]
- Parvin, J.D.; Shykind, B.M.; Meyers, R.E.; Kim, J.; Sharp, P.A. Multiple sets of basal factors initiate transcription by RNA polymerase II. J. Biol. Chem. 1994, 269, 18414–18421. [Google Scholar] [CrossRef]
- Yang, C.; Bolotin, E.; Jiang, T.; Sladek, F.M.; Martinez, E. Prevalence of the initiator over the TATA box in human and yeast genes and identification of DNA motifs enriched in human TATA-less core promoters. Gene 2007, 389, 52–65. [Google Scholar] [CrossRef] [Green Version]
- Kaufmann, J.; Ahrens, K.; Koop, R.; Smale, S.T.; Müller, R. CIF150, a human cofactor for transcription factor IID-dependent initiator function. Mol. Cell Biol. 1998, 18, 233–239. [Google Scholar] [CrossRef] [Green Version]
- Kaufmann, J.; Verrijzer, C.P.; Shao, J.; Smale, S.T. CIF, an essential cofactor for TFIID-dependent initiator function. Genes Dev. 1996, 10, 873–886. [Google Scholar] [CrossRef] [Green Version]
- Chalkley, G.E.; Verrijzer, C.P. DNA binding site selection by RNA polymerase II TAFs: A TAF(II)250-TAF(II)150 complex recognizes the initiator. EMBO J. 1999, 18, 4835–4845. [Google Scholar] [CrossRef]
- Verrijzer, C.P.; Tjian, R. TAFs mediate transcriptional activation and promoter selectivity. Trends Biochem. Sci. 1996, 21, 338–342. [Google Scholar] [CrossRef]
- Martinez, E.; Zhou, Q.; L’Etoile, N.D.; Oelgeschläger, T.; Berk, A.J.; Roeder, R.G. Core promoter-specific function of a mutant transcription factor TFIID defective in TATA-box binding. Proc. Natl. Acad. Sci. USA 1995, 92, 11864–11868. [Google Scholar] [CrossRef] [Green Version]
- Lewis, B.A.; Sims, R.J., 3rd; Lane, W.S.; Reinberg, D. Functional characterization of core promoter elements: DPE-specific transcription requires the protein kinase CK2 and the PC4 coactivator. Mol. Cell 2005, 18, 471–481. [Google Scholar] [CrossRef]
- Aso, T.; Conaway, J.W.; Conaway, R.C. Role of core promoter structure in assembly of the RNA polymerase II preinitiation complex. A common pathway for formation of preinitiation intermediates at many TATA and TATA-less promoters. J. Biol. Chem. 1994, 269, 26575–26583. [Google Scholar] [CrossRef]
- Xu, M.; Sharma, P.; Pan, S.; Malik, S.; Roeder, R.G.; Martinez, E. Core promoter-selective function of HMGA1 and Mediator in Initiator-dependent transcription. Genes Dev. 2011, 25, 2513–2524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hantsche, M.; Cramer, P. Conserved RNA polymerase II initiation complex structure. Curr. Opin. Struct. Biol. 2017, 47, 17–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Il’icheva, I.A.; Khodikov, M.V.; Poptsova, M.S.; Nechipurenko, D.Y.; Nechipurenko, Y.D.; Grokhovsky, S.L. Structural features of DNA that determine RNA polymerase II core promoter. BMC Genom. 2016, 17, 973. [Google Scholar] [CrossRef] [Green Version]
- Haberle, V.; Stark, A. Eukaryotic core promoters and the functional basis of transcription initiation. Nat. Rev. Mol. Cell Biol. 2018, 19, 621–637. [Google Scholar] [CrossRef]
- Liston, D.R.; Johnson, P.J. Analysis of a ubiquitous promoter element in a primitive eukaryote: Early evolution of the initiator element. Mol. Cell Biol. 1999, 19, 2380–2388. [Google Scholar] [CrossRef] [Green Version]
- Liston, D.R.; Lau, A.O.; Ortiz, D.; Smale, S.T.; Johnson, P.J. Initiator recognition in a primitive eukaryote: IBP39, an initiator-binding protein from Trichomonas vaginalis. Mol. Cell Biol. 2001, 21, 7872–7882. [Google Scholar] [CrossRef] [Green Version]
- Lau, A.O.; Smith, A.J.; Brown, M.T.; Johnson, P.J. Trichomonas vaginalis initiator binding protein (IBP39) and RNA polymerase II large subunit carboxy terminal domain interaction. Mol. Biochem. Parasitol. 2006, 150, 56–62. [Google Scholar] [CrossRef]
- Parra-Marín, O.; Rosas-Hernández, L.; López-Pacheco, K.; Franco, B.; Ibáñez-Escribano, A.; Hernández, R.; López-Villaseñor, I. An in vitro characterisation of the Trichomonas vaginalis TATA box-binding proteins (TBPs). Parasitol. Res. 2019, 118, 3019–3031. [Google Scholar] [CrossRef]
- Mittler, G.; Kremmer, E.; Timmers, H.T.; Meisterernst, M. Novel critical role of a human Mediator complex for basal RNA polymerase II transcription. EMBO Rep. 2001, 2, 808–813. [Google Scholar] [CrossRef] [PubMed]
- Takagi, Y.; Kornberg, R.D. Mediator as a general transcription factor. J. Biol. Chem. 2006, 281, 80–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baek, H.J.; Kang, Y.K.; Roeder, R.G. Human Mediator enhances basal transcription by facilitating recruitment of transcription factor IIB during preinitiation complex assembly. J. Biol. Chem. 2006, 281, 15172–15181. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rojas, D.A.; Urbina, F.; Valenzuela-Pérez, L.; Leiva, L.; Miralles, V.J.; Maldonado, E. Initiator-Directed Transcription: Fission Yeast Nmtl Initiator Directs Preinitiation Complex Formation and Transcriptional Initiation. Genes 2022, 13, 256. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13020256
Rojas DA, Urbina F, Valenzuela-Pérez L, Leiva L, Miralles VJ, Maldonado E. Initiator-Directed Transcription: Fission Yeast Nmtl Initiator Directs Preinitiation Complex Formation and Transcriptional Initiation. Genes. 2022; 13(2):256. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13020256
Chicago/Turabian StyleRojas, Diego A., Fabiola Urbina, Lucía Valenzuela-Pérez, Lorenzo Leiva, Vicente J. Miralles, and Edio Maldonado. 2022. "Initiator-Directed Transcription: Fission Yeast Nmtl Initiator Directs Preinitiation Complex Formation and Transcriptional Initiation" Genes 13, no. 2: 256. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13020256