
The Molecular Genetics of Dissociative Symptomatology: A Transdiagnostic Literature Review


Department of Psychiatry, Jawaharlal Institute of Postgraduate Medical Education and Research (JIPMER), Puducherry 605 006, India
Genes 2022, 13(5), 843; https://0-doi-org.brum.beds.ac.uk/10.3390/genes13050843
Submission received: 23 April 2022
/
Revised: 5 May 2022
/
Accepted: 6 May 2022
/
Published: 8 May 2022
(This article belongs to the Special Issue Genetic Basis Underlying Neuropsychiatric Disorders)
Abstract
:Dissociative disorders are a common and frequently undiagnosed group of psychiatric disorders, characterized by disruptions in the normal integration of awareness, personality, emotion and behavior. The available evidence suggests that these disorders arise from an interaction between genetic vulnerability and stress, particularly traumatic stress, but the attention paid to the underlying genetic diatheses has been sparse. In this paper, the existing literature on the molecular genetics of dissociative disorders, as well as of clinically significant dissociative symptoms not reaching the threshold of a disorder, is reviewed comprehensively across clinical and non-clinical samples. Association studies suggest a link between dissociative symptoms and genes related to serotonergic, dopaminergic and peptidergic transmission, neural plasticity and cortisol receptor sensitivity, particularly following exposure to childhood trauma. Genome-wide association studies have identified loci of interest related to second messenger signaling and synaptic integration. Though these findings are inconsistent, they suggest biologically plausible mechanisms through which traumatic stress can lead to pathological dissociation. However, methodological concerns related to phenotype definition, study power, and correction for the confounding factors limit the value of these findings, and they require replication and extension in studies with better design.
1. Introduction
Dissociative disorders represent an important group of psychiatric disorders occurring in response to stress or trauma [1]. According to the American Psychiatric Association’s Diagnostic and Statistical Manual for Mental Disorders, Fifth Edition (DSM-5), these disorders are characterized by “a disruption of and/or discontinuity in the normal integration of consciousness, memory, identity, emotion, perception, body representation, motor control, and behavior” [2]. The disorders in this group include depersonalization/derealization disorder, dissociative amnesia, and dissociative identity disorder. The estimated lifetime prevalence of these disorders is around 10% in general population samples [3,4,5], with higher rates being reported in patients seeking treatment for other psychiatric disorders [6,7] and in groups exposed to trauma [8,9,10]. In addition to these discrete syndromes, clinically significant symptoms of dissociation occur in a wide range of other psychiatric disorders, including post-traumatic stress disorder (PTSD), personality disorders, somatic symptom disorders, depression, anxiety disorders, and eating disorders [11]. Dissociative disorders tend to run a chronic course [12], and are associated with elevated levels of disability, impaired quality of life, a high economic cost, and a significantly increased risk of suicide attempts [13,14]. In spite of the substantial burden associated with these conditions, patients suffering from them often go undiagnosed and untreated [7,15]. This is due to several factors, including variations in criteria across diagnostic systems [16], negative attitudes towards these disorders among both the general public and mental health professionals [17,18], and therapeutic pessimism arising from a paucity of evidence-based treatment approaches [19,20].
From a psychological perspective, research into the etiology of dissociative disorders has centered on “post-traumatic” and “socio-cognitive” models; however, more recent evidence suggests that neither model is completely satisfactory. Instead, these disorders should be understood as involving impairments in multiple domains of mental functioning, including self-regulation, reality testing, cognitive attributions, and higher-order cognitive processes [21]. Neurobiological evidence converges with this perspective, suggesting that dissociative disorders are best understood as belonging to a group of trauma-related disorders which share certain key structural and functional abnormalities. These include grey matter volume reductions in key limbic system structures, the dysregulation of prefrontal–limbic circuitry, and the altered functioning of the hypothalamic–pituitary-adrenal (HPA) axis [22]. These structural and functional alterations may underlie the multiple domains of impairment identified in psychological research. A recent meta-analysis identified reductions in the volume of hippocampus, basal ganglia and thalami, increased levels of the neuropeptide transmitters oxytocin and prolactin, and reduced levels of the inflammatory marker tumor necrosis factor alpha (TNF-α) as replicable biological markers of dissociative disorders [23]. Attempts to integrate the results of biological and psychological research into possible models of pathogenesis have already been made in the case of dissociative identity disorders [24] and dissociative amnesia [25,26].
A key finding from empirical research is that dissociation is a frequent, but not universal, response following exposure to a traumatic stressor. For example, around 20–25% of individuals exposed to a disaster may experience transient dissociative symptoms, but only a small proportion of them fulfill criteria for a later dissociative disorder [27]. Similarly, a study of women who had been held captive and subjected to sexual violence by a militant group found that only 41% met criteria for a dissociative disorder [28]. The finding that dissociation is not an invariable result of traumatic stress, even when severe and prolonged, suggests that some of this variation is due to an innate predisposition which is at least partly genetic in origin [29,30]. This hypothesis is supported by the results of twin studies which suggest that dissociation has a heritable component [31,32]. Understanding the contributions of specific genetic factors to dissociative disorders, whether in terms of monogenic, polygenic or epigenetic processes, would deepen our understanding of the stress–diathesis equation in these disorders. The insights obtained from molecular genetic data could potentially open the door to more effective forms of prevention and treatment, including early intervention strategies following a mass traumatic event. The current paper attempts to aid in this understanding by reviewing the scientific literature on the genetics of dissociative symptoms and disorders.
2. Materials and Methods
The current review was conducted in line with the principles outlined in the PRISMA guidelines [33]. A comprehensive search of the PubMed, Scopus and ProQuest literature databases was carried out using the key search terms “dissociative disorder”, “dissociative disorders”, “dissociative identity disorder”, “depersonalization”, “derealization”, and “dissociative amnesia” in association with “genetic”, “gene”, “polymorphism”, “genome”, “genome-wide”, “epigenetic”, “heritability”, and “inheritance”. All articles published up to 31 March 2022 were included. The complete search strategy is provided in Table S1.
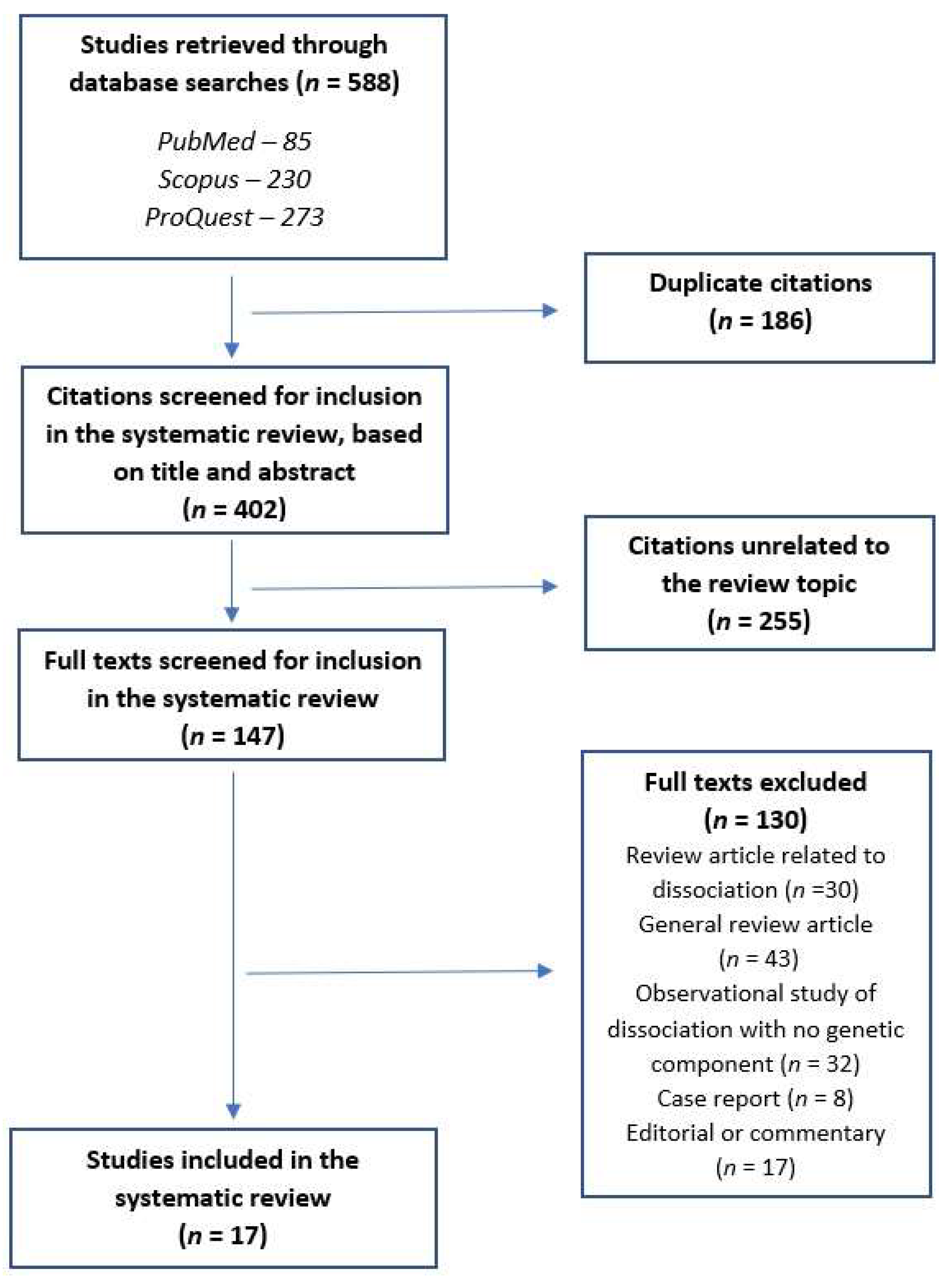
Papers were included in the review if they presented the results of genetic studies of either (a) dissociative disorders, as defined by DSM-5, or (b) clinically significant dissociative symptoms (“pathological dissociation”) either alone or in conjunction with other psychiatric disorders, as measured using a standardized rating scale. A total of 588 citations were retrieved through the above search. After the removal of 186 duplicate citations, 402 citations (title and abstract) were screened. At this stage, 255 citations unrelated to the subject were excluded, and 147 papers were assessed for eligibility. Of these, only 17 met the criteria for inclusion in this review. To ensure that no citations were missed in this process, the following precautions were taken: (a) citation lists from each database were entered into individual data sheets and cross-checked against each other, and (b) the reference lists of all published studies were checked for relevant citations that may have been missed in a standard search. No additional papers were included as a result of (b). An outline of this process is provided in Figure 1 and complete details of reasons for exclusion are provided in Table S2.
When planning this review, a specific problem was posed by studies of “non-pathological” dissociative experiences, which can occur in healthy individuals and do not qualify for a psychiatric diagnosis. While some authors consider these experiences to lie on a continuum with dissociative symptoms or disorders [34,35], the majority of contemporary researchers make a clear distinction between “normal” and “pathological” dissociation based on psychometric data [36,37,38]. As only two studies of this phenomenon were included in this review, their findings are discussed along with the rest of the included studies.
The quality of individual studies was assessed using the Q-Genie tool, which is an 11-item scale specifically developed to assess the quality of genetic association studies for the purpose of a literature review or meta-analysis [39]. These guidelines include an item covering the risk of bias, which is reported separately along with the total score for the purposes of this review. For genetic studies containing a comparison group, study quality was rated based on the total Q-Genie score as poor (≤35), moderate (36–45), or good (>45). For studies without a comparison group, the corresponding cut-offs were: poor (≤32), moderate (33–40), and good (>40). Bias was rated on a seven-point scale, with the following categories: poor (1–2), good (3–4), very good (5–6), and excellent (7). Complete scores for each article are provided in Table S3.
Due to the small number of relevant studies and significant methodological heterogeneity, a formal meta-analysis was not carried out.
3. Results
A total of seventeen studies were included in the final review [40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56]. The majority of these were association studies involving a small number of predefined genetic polymorphisms or genotypes, with or without a gene–environment interaction (G×E) component (n = 12); there were three genome-wide association studies, one gene expression study, and one study making use of polygenic risk scores. A complete description of these studies is provided in Table 1.
3.1. Quality of the Included Studies
Q-Genie scores for each study are provided in the Supplementary Material. An examination of the total Q-Genie scores for each study found that nine studies were rated as “moderate” in quality” [40,42,43,45,46,48,50,53,54], seven were rated as “good” [44,47,49,51,52,55,56], and only one was rated as “poor” [41]. The mean Q-Genie score was 39.3 for studies without a control group (n = 11) and 43.3 for studies with a control group (n = 6), indicating an overall “moderate” quality of research in this field.
When evaluating studies specifically in terms of bias, thirteen studies received a rating of “good” and four studies were rated “poor”; no study received a rating of “very good” or “excellent” when evaluated for sources of bias. The mean overall score for bias was 3.1, indicating an overall “good” quality for the included studies.
An additional issue of concern was that several authors, particularly those of association and G×E studies, reported concerns related to study power sample size. Twelve studies received a rating of “poor” (scores 1–2) on this item, and only four studies received a score of “good” or “very good” (scores 3–5). The mean score on this item across all included studies was 2.5, indicating significant concerns related to study power.
3.2. Study Populations
None of the included studies focused on patients with a primary diagnosis of dissociative disorder. Two studies were conducted in “normal” adults, with no medical or psychiatric diagnosis, selected from the general population [46,54]. Five studies were conducted in subjects considered to be “high-risk” or vulnerable, including participants from socially and economically deprived backgrounds [47,52] and victims of trauma [41,45,49]. Three studies were based on samples from participants in biobanking programs who had a wide range of medical or psychiatric diagnoses [51,55,56]. The remaining studies were conducted in patients with a primary psychiatric diagnosis other than a dissociative disorder, including obsessive–compulsive and related disorders (OCD) [40,42], borderline personality disorder [44,48], depression [50,53] and bipolar disorder (BD) [43]. The majority of studies were conducted in adults, with only two studies focusing on children and adolescents [41,52]. None of the studies involved a replication of current or past findings in a separate study population.
Ethnicity and gender are important potential confounding factors in studies of neuropsychiatric genetics [57,58,59,60]. Eight of the seventeen studies were conducted in samples where an attempt was made to ensure a degree of ethnic homogeneity: seven studies of subjects of Caucasian/European descent [40,42,45,49,51,55], and one each of Mexican-American [53] and Japanese [54] descent. Three studies recruited only women in an attempt to correct for the influence of gender [47,48,50], while the remainder included both men and women. In all studies with samples of mixed ethnicity or gender, attempts to correct for these confounders were reported in the study methodology and results.
3.3. Definition of the Phenotype of Interest
A clear definition of the phenotype being studied in relation to genetic variants is essential in psychiatric genetics, where the categorical diagnoses used in clinical practice may not correspond to meaningful subtypes at the biological level [61]. Only three of the studies included in this review used categorical diagnoses as the phenotype of interest, due to their reliance on medical records, and in these studies dissociative disorders were “lumped” together with anxiety and phobic disorders [51,55,56]. This significantly limits the validity of any conclusions that can be drawn from these studies with regard to dissociation alone. In contrast, the majority of studies [40,41,42,43,44,45,46,47,48,52,54] measured total dissociative symptoms using a standardized rating scale, such as the Dissociative Experiences Scale. Such psychometric instruments provide a total score, a “cut-off” value indicating “pathological dissociation” likely to be of clinical significance, and sub-scale scores for distinct symptom dimensions, such as amnesia and depersonalization/derealization. In theory, this should allow for a more “fine-grained” analysis of dissociative symptoms in relation to genotypes; however, such an analysis was carried out only in a few of the included studies [46,54]. The remaining three studies measured only depersonalization/derealization symptoms and did not examine other sets of symptoms; thus, their findings were applicable only to this specific dimension of dissociative symptomatology [49,50,53]. As noted above, no study including patients with dissociative disorders alone was identified for inclusion in this review. A complete description of the rating scales used in each study is provided in Table S4.
3.4. Associations with Specific Polymorphisms or Genotypes
Twelve of the studies included in this review attempted to establish an association between dissociation and either a single nucleotide polymorphism (SNP) or a genotype involving a limited number of polymorphisms or variants.
Six of these studies focused on genetic variants associated with the monoamine neurotransmitters serotonin, dopamine and noradrenaline, which are associated with stress- and trauma-related disorders [62]. The most commonly studied variant (n = 4) was 5-HTTLPR, a functional polymorphism of the promoter region of the serotonin transporter. Two studies found an association between the “short” (s) allele of this polymorphism, particularly the homozygous s/s genotype, and dissociative symptoms [42,46]; however, two other studies failed to find any association between this polymorphism and dissociation [40,43]. An equally frequent subject of study was the COMT gene, encoding the catechol-O-methyltransferase enzyme involved in the catabolism of dopamine and norepinephrine. A study of adults involved in road traffic accidents found an association between a “pain-sensitive” COMT haplotype, involving four functional polymorphisms (rs4633-rs4680-rs4818-rs6269 A_C_C_G), and dissociative symptoms [45], while a study of healthy adults found an association between the COMT rs4680 (Val158Met) SNP and both total dissociative and depersonalization/derealization symptoms [54]. However, as in the case of 5-HTTLPR, negative results were obtained in two studies [40,43]. Studies of other monoamine-related polymorphisms or variants, such as those involving the genes for the dopamine type 4 receptor (DRD4), the dopamine transporter (DAT), the serotonin type 1B and type 2C receptors (HTR1B, HTR2C), and the enzymes tryptophan hydroxylase 1 (TPH1) and monoamine oxidase A (MAOA) did not yield any significant results with reference to dissociation [40,43]
Among other variants studied using an association design, two functional polymorphisms (rs3800373, rs1360870) of the FKBP5 gene, encoding a protein that regulates glucocorticoid receptor sensitivity [63], were associated with dissociation during and after acute physical injury in children [41]. Possible associations between three functional polymorphisms (rs7607967, rs4371369, rs4387806) of the SCN9A gene, encoding a voltage-gated sodium channel expressed in limbic system structures, were observed in women with borderline personality disorder, but these findings were insignificant after statistical correction [44]. The Met allele of the rs6265 polymorphism of the BDNF gene, encoding the brain-derived neurotrophic factor, was associated with lower levels of dissociation in patients with bipolar disorder and their relatives [43]; this variant is associated with a reduced susceptibility to post-traumatic stress disorder [64]. A study of the oxytocin receptor gene (OXTR), which is associated with stress sensitivity, found an association between homozygosity for the G allele of the rs53576 functional polymorphism in this gene and dissociation, but this was not statistically significant.
3.5. Gene–Environment Interactions
There is an increasing amount of evidence that trauma-related disorders arise from a dynamic interaction between genes and the environment [65]. In line with this model, six studies have examined the interaction between a specific genotype and exposure to adverse environmental circumstances in relation to dissociation. Two studies, one of normal adults and one of patients with OCD, found a significant interaction between the 5-HTTLPR s/s genotype, childhood trauma, and subsequent dissociative symptoms [42,46]; however, a negative result for an s/s x childhood trauma interaction was noted in bipolar patients and their relatives [43]. A study of patients with bipolar disorder and their relatives found a significant interaction between the COMT rs4680 Val allele, childhood trauma, and dissociative symptoms [43]. A study of depressed and healthy women found that the rs53576 G/G genotype of OXTR was associated with dissociative symptoms in those with a history of unresolved childhood attachment [50]. Finally, two studies examined a haplotype involving four functional polymorphisms (rs3800373, rs9296158, rs1360870, rs9470080 C_A_T_T) of the FKBP5 gene. While this haplotype was associated with an elevated risk of dissociation in low-income women with a history of childhood trauma [47], it appeared to have the opposite effect in low-income adolescents; adolescents with this haplotype had lower dissociative symptoms even when exposed to significant trauma from an early age [52]. No evidence of gene–environment interaction was observed for the functional variants of the DRD4 or DAT genes in one of these studies [43].
3.6. Genome-Wide Association Studies
Three genome-wide association studies were identified in the literature. Only one of these specifically examined dissociative symptoms followed trauma as a phenotype [49]. In this study, no significant associations were found; however, suggestive “peak” associations were reported for the adenylyl cyclase 8 gene (ADCY8) and the dipeptidyl-peptidase 6 gene (DPP6). The authors did not report any association between post-traumatic dissociation and any of the genes identified through association studies, such as 5HTT, COMT or FKBP5, though marginal associations were identified for FKBP5 and COMT which were insignificant after statistical correction.
The two remaining genome-wide studies, based on large samples from biobanking projects, did not study dissociative disorders or symptoms as a primary objective; they examined a large number of groups of disorders affecting various organs and systems, one of which was “anxiety, phobic and dissociative disorders”. One of these identified a potential association between “anxiety, phobic and dissociative disorders” and a region of chromosome 4 containing the amyloid beta precursor protein-binding family member 2 (ABPP2) gene [51]; the other did not report any significant association for this group of disorders [56].
3.7. Other Study Designs
Three additional studies on the genetics of dissociative disorders or symptoms did not fit into any specific category and are described here.
In a study of a small sample of women hospitalized with borderline personality disorder, an association was found between dissociative symptoms and the increased expression of the interleukin-6 (IL6) gene. Subjects with higher levels of these symptoms also had decreased expression of the following genes: interleukin 1-beta (IL1B), mitogen-activated protein kinases-1, -3, and -8 (MAPK1, MAPK3, MAPK8), G-protein subunit alpha-I2 (GNAI2), arrestin beta-1 and -2 (ARRB1, ARRB2), and cyclic AMP responsive element-binding protein 1 (CREB1) [48].
In a study involving only subjects of Mexican-American ancestry, an analysis of nineteen SNPs identified as associated with depression in earlier research were examined in relation to various symptoms of depression. It was found that this specific set of SNPs was associated with lower depersonalization/derealization symptoms in patients with depression; however, this was not the primary objective of the study [53].
Using polygenic risk scores, a study of 10182 subjects of European descent participating in a biobanking program found that the polygenic risk scores for both depression and bipolar disorder were significantly associated with the group of “anxiety, phobic and dissociative disorders”, suggesting a potential genetic association between these groups of disorders [56].
Specific details of the results of individual studies in terms of scores on standardized rating scales are available in Table S4.
4. Discussion
When compared with other psychiatric disorders or symptom domains, dissociative disorders and symptoms have been relatively under-studied from a genetic perspective. When familial patterns were observed for these disorders, they were often explained exclusively on the basis of psychological mechanisms, such as exposure to trauma or learned behavior [66,67]. Family and twin-based genetic studies of dissociative disorder have suggested that this condition has a substantial heritable component [31,32,68], but the results of these studies were not consistent [69,70] due to methodological limitations and the assumptions made when assessing patterns of inheritance. In contrast with these earlier approaches, molecular genetic studies hold the promise of identifying the biological mechanisms associated with vulnerability to dissociative symptoms and disorders with increasing precision, thus allowing for a more accurate approach to diagnosis and management [22] as well as a deeper understanding of the place of dissociation in psychiatric classification [71,72].
In this review, preliminary evidence for an association between dissociative symptomatology and variations in individual genes was identified. The genes implicated are related to monoaminergic transmission (5-HTT, COMT), neural plasticity (BDNF), neuropeptide receptors (OXTR), and the regulation of the hypothalamic–pituitary–adrenal axis (FKBP5). These genes have been associated with other stress- and trauma-related symptoms and disorders [73,74,75,76], though results have not always been consistent. Studies in patients with dissociative disorders or symptoms have also found indirect evidence of dysfunction involving these systems. For example, dissociative symptoms are associated with a poorer response to serotonergic antidepressants in patients being treated for other disorders [77,78]; the pharmacological manipulation of serotonergic transmission can induce dissociative symptoms [79,80]; levels of noradrenaline and dopamine are found to be elevated in certain dissociative states [81]; dissociative disorders are associated with higher levels of the neuropeptides oxytocin and prolactin [23]; and pathological and non-pathological dissociative experiences are associated with distinctive alterations in cortisol secretion [82]. While these findings provide a certain degree of biological plausibility to the results of these single-gene association studies, it is also important to note that these findings have not been replicated consistently, and that these loci were identified as being unrelated to, or only marginally associated with, dissociation in a genome-wide analysis [49]. It is therefore unlikely that these variants account for a substantial proportion of the risk for dissociation, though a case could still be made for the role of gene–environment interactions with childhood adversity in the case of the 5-HTT, COMT, OXTR and FKBP5 functional polymorphisms.
The results of genome-wide association studies suggest a possible association with loci within the ADYC8, DPP6, and APBB2 genes. There is evidence linking variations in these genes with other psychiatric disorders. ADCY8 codes for the enzyme adenylyl cyclase, which catalyzes the conversion of adenosine triphosphate into cyclic adenosine monophosphate (cAMP), a key second messenger for several neurotransmitters. It has been associated with comorbid alcohol dependence and depression in women [83], obsessive–compulsive disorder [84], and avoidance behavior related to mood disorders in an animal model [85]. DPP6, which encodes a potassium channel subunit related to the excitability of neuronal dendrites and synaptic integration, has been associated with tic disorder [86]. Both ADYC8 and DPP6 have also been linked to brain development in childhood [84,87], which may be of relevance to dissociative disorders associated with trauma-related alterations in brain development [88,89]. In contrast, APBB2, which encodes an amyloid beta precursor-binding protein, has been associated with dementia [90]. This finding is of equal interest given the emerging evidence of a prospective association between trauma, traumatic stress-related disorders, and subsequent dementia later in life [91]. Despite this supporting evidence, it must be noted that no consistent or highly significant finding has been reported in the GWAS of dissociative symptoms or dissociative disorder; therefore, these results should be interpreted with caution.
The interpretation of other study designs [48,53,55] is less straightforward. However, evidence for increased IL6 gene expression is consistent with reports of elevated interleukin-6 (IL-6) being associated with dissociative symptoms in depression [92] and with elevated levels of IL-6 in other trauma spectrum disorders [93]. However, IL-6 levels measured immediately after trauma did not appear to predict the course of subsequent trauma-related disorders [94], suggesting that these alterations may appear later in the course of the illness. Likewise, the polygenic association between dissociative disorders and mood disorders is consistent with evidence of high levels of dissociative symptoms in some patients with major depression; some researchers have considered the possibility of a “dissociative depression” subtype associated with childhood abuse and a higher risk of suicide attempts [95], which is consistent with a complex, shared genetic architecture for these disorders.
Several methodological issues should be taken into account when appraising these results. First, though the existing research is of acceptable quality with regard to sources of bias, it is of only “moderate” quality overall, and significant concerns exist with regard to sample size and study power. None of the included studies involved patients with a primary diagnosis of dissociative disorder. Several studies were carried out in patients with another psychiatric diagnosis, introducing a substantial confounding factor. Association studies generally focused on genes related to a limited number of neural or neuroendocrine pathways, many of which are non-specifically associated with a wide range of psychiatric disorders [96,97]. In gene–environment designs, only specific forms of environmental risk, such as childhood abuse by parents or caregivers, were assessed; other relevant forms of traumatic stress, such as bullying by peers, intimate partner violence, or sexual assault after adolescence, were not investigated despite their relevance to dissociation [98,99,100]. In studies measuring dissociative symptoms, there was significant heterogeneity in the instruments used to measure symptom severity, as well as in the specific type(s) of symptoms being studied; in some cases, data on different dimensions of dissociation in relation to the genotype was not analyzed, despite being available. In genome-wide association studies, dissociative disorders were grouped together with anxiety and phobic disorders for analysis in some cases. While this is an inherent limitation of using biobank data in which patient diagnoses are coded using older classificatory systems, it reduces the specificity of any reported findings with reference to dissociation, per se. Not all studies made attempts to correct for confounding factors such as sex and ethnicity. Finally, none of the included studies included a validation of their findings in a separate sample or population. Because of these limitations, it is possible that both false negative (due to low study power) and false positive (due to variations in phenotype, multiple confounding factors, and a lack of replicated associations) results may have been reported in individual studies. In practical terms, this implies that the results reported here are in need of replication before they can truly inform our understanding of the molecular processes underlying dissociative disorders and symptoms.
There are also certain limitations inherent to this review. The retrieval of citations from three specific databases may have led to the inadvertent omission of certain key studies. The marked heterogeneity across studies and the lack of replicated findings for individual associations precluded a formal meta-analysis, limiting the confidence that can be placed in the conclusions drawn above. The current review included only studies of dissociative disorders or symptoms as defined using current criteria; it is possible that conditions such as somatoform and conversion disorders, though placed in separate categories in current systems of classification, may be genetically and neurobiologically linked to dissociation and thus should be studied together [22,101]. As no study included patients with a primary diagnosis of dissociative disorders, as mentioned above, one of the objectives of this review could not be met. Finally, as the review was carried out by a single reviewer, there is a possibility that certain relevant papers may not have been included, though attempts were made to reduce this possibility to the maximum extent possible.
Despite these limitations, the existing evidence suggests the possibility of links between dissociative symptoms and specific genes related to monoamine transmission, neural plasticity, hypothalamic–pituitary–adrenal axis functioning, peptidergic neurotransmission, second messenger signaling, and synaptic integration. Dissociative symptoms may arise from an interaction between functional variants in these genes and early life adversity, particularly childhood abuse. These findings require replication in larger and more homogenous samples, as do the more tentative findings linking dissociation to genes involved in immune functioning. From a clinical perspective, it is possible that the pharmacological manipulation of these pathways may result in the development of better treatments for dissociative disorders. From a more fundamental perspective, there is a need to examine the genetics of dissociation from the point of view of other biomarkers identified in the literature. Potential genetic variants of interest that have not yet been studied include the prolactin receptor [23], opioid peptide receptors [81], genes involved in cytokine signaling [94], and genes involved in lipid metabolism [102]. Studies of gene–environment interactions should move beyond the examination of childhood trauma to include both subtler disruptions in attachment bonds [50] and the effect of traumatic stress later in life, both at an individual level and in survivors of disasters or mass casualties. There is also a need for enhanced genome-wide association studies involving either a more precise definition of the “dissociative” phenotype, or an exploration of shared, “common” genetic factors underlying the co-occurrence of dissociative and other disorders. Epigenetic studies could lead to a more dynamic model of alterations in gene expression and their downstream consequences in patients with clinically significant dissociation [103]. Finally, studies of the genetics of dissociation in relation to other trauma spectrum disorders, such as PTSD and borderline personality disorder, would aid the identification of shared and unique genetic vulnerabilities for this group of disorders.
5. Conclusions
Despite the relatively small number of studies in this area, research on the molecular genetics of dissociative symptoms and disorders has yielded clues pointing both towards conventional, “stress-related” neural mechanisms and novel genetic loci of interest. These results should be interpreted with caution in view of the methodological limitations discussed. However, these findings provide a foundation that can be built upon by further studies with more complex designs, and it is hoped that this will lead to a better understanding of the pathogenesis and treatment of this group of trauma-related disorders.
Supplementary Materials
The following supporting information can be downloaded at: https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/genes13050843/s1, Table S1, search strategy for the literature review; Table S2, articles excluded and reasons for exclusion; Table S3, quality assessments of each study using the Q-Genie tool; Table S4, details of measures of dissociation and scores on standard scales for each study, where applicable.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No original data was reported in this study. Data pertinent to the current review is provided in the text, table and Supplementary Material.
Conflicts of Interest
The author declares no conflict of interest.
References
- Spiegel, D.; Loewenstein, R.J.; Lewis-Fernandez, R.; Sar, V.; Simeon, D.; Vermetten, E.; Cardena, E.; Dell, P.F. Dissociative disorders in DSM-5. Depress. Anxiety 2011, 28, E17–E45. [Google Scholar] [CrossRef] [PubMed]
- Stein, D.J.; Craske, M.A.; Friedman, M.J.; Phillips, K.A. Anxiety disorders, obsessive-compulsive and related disorders, trauma- and stressor-related disorders, and dissociative disorders in DSM-5. Am. J. Psychiatry 2014, 171, 611–613. [Google Scholar] [CrossRef] [PubMed]
- Elmore, J.L. Dissociative spectrum disorders in the primary care setting. Prim. Care Companion J. Clin. Psychiatry 2000, 2, 37–41. [Google Scholar] [CrossRef] [Green Version]
- Kate, M.-A.; Hopwood, T.; Jamieson, G. The prevalence of dissociative disorders and dissociative experiences in college populations: A meta-analysis of 98 studies. J. Trauma Dissociation 2020, 21, 16–61. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.G.; Cohen, P.; Kasen, S.; Brook, J.S. Dissociative disorders among adults in the community, impaired functioning, and axis I and II comorbidity. J. Psychiatry Res. 2006, 40, 131–140. [Google Scholar] [CrossRef]
- Horen, S.A.; Leicher, P.P.; Lawson, J.S. Prevalence of dissociative symptoms and disorders in an adult psychiatric inpatient population in Canada. Can. J. Psychiatry 1995, 40, 185–191. [Google Scholar]
- Foote, B.; Smolin, Y.; Kaplan, M.; Legatt, M.E.; Lipschitz, D. Prevalence of dissociative disorders in psychiatric outpatients. Am. J. Psychiatry 2006, 163, 623–629. [Google Scholar] [CrossRef]
- Roca, V.; Hart, J.; Kimbrell, T.; Freeman, T. Cognitive function and dissociative disorder status among veteran subjects with chronic posttraumatic stress disorder: A preliminary study. J. Neuropsychiatry Clin. Neurosci. 2006, 18, 226–230. [Google Scholar] [CrossRef]
- Somer, E.; Ross, C.; Kirshberg, R.; Bakri, R.S.; Ismail, S. Dissociative disorders and possession experiences in Israel: A comparison of opiate use disorder patients, Arab women subjected to domestic violence, and a nonclinical group. Transcult. Psychiatry 2015, 52, 58–73. [Google Scholar] [CrossRef] [Green Version]
- Manzano-Mojica, J.; Martinez-Taboas, A.; Sayers-Montalvo, S.K.; Cabiya, J.J.; Alicea-Rodriguez, L.E. Dissociation in sexually abused Puerto Rican children: A replication utilizing social workers as informers. J. Trauma Dissociation 2012, 13, 330–344. [Google Scholar] [CrossRef]
- Lyssenko, L.; Schmahl, C.; Bockhacker, L.; Vonderlin, R.; Bohus, M.; Kleindienst, N. Dissociation in psychiatric disorders: A meta-analysis of studies using the Dissociative Experiences Scale. Am. J. Psychiatry 2018, 175, 37–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jans, T.; Schneck-Seif, S.; Weigand, T.; Schneider, W.; Ellgring, H.; Wewetzer, C.; Warnke, A. Long-term outcome and prognosis of dissociative disorder with onset in childhood or adolescence. Child Adolesc. Psychiatry Ment. Health 2008, 2, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langeland, W.; Jepsen, E.K.K.; Brand, B.L.; Kleven, L.; Loewenstein, R.J.; Putnam, F.W.; Schielke, H.J.; Myrick, A.; Lanius, R.A.; Heir, T. The economic burden of dissociative disorders: A qualitative systematic review of empirical studies. Psychol. Trauma 2020, 12, 730–738. [Google Scholar] [CrossRef] [PubMed]
- Calati, R.; Bensassi, I.; Courtet, P. The link between dissociation and both suicide attempts and non-suicidal self-injury: Meta-analyses. Psychiatry Res. 2017, 251, 103–114. [Google Scholar] [CrossRef]
- Coons, M.P. The dissociative disorders: Rarely considered and underdiagnosed. Psychiatr. Clin. North. Am. 1998, 21, 637–648. [Google Scholar] [CrossRef]
- Brown, R.J.; Cardena, E.; Nijenhuis, E.; Sar, V.; van der Hart, O. Should conversion disorder be reclassified as a dissociative disorder in DSM V? Psychosomatics 2007, 48, 369–378. [Google Scholar] [CrossRef]
- Loewenstein, R.J. Dissociation debates: Everything you know is wrong. Dialogues Clin. Neurosci. 2018, 20, 229–242. [Google Scholar] [CrossRef]
- Lalonde, J.K.; Hudson, J.I.; Gigante, R.A.; Pope, H.G. Canadian and American psychiatrists’ attitudes towards dissociative disorders diagnoses. Can. J. Psychiatry 2001, 46, 407–412. [Google Scholar] [CrossRef] [Green Version]
- Sutar, R.; Sahu, S. Pharmacotherapy for dissociative disorders: A systematic review. Psychiatry Res. 2019, 281, 112529. [Google Scholar] [CrossRef]
- Ganslev, C.A.; Storebo, O.J.; Callesen, H.E.; Ruddy, R.; Sogaard, U. Psychosocial interventions for conversion and dissociative disorders in adults. Cochrane Database Syst. Rev. 2020, 7, CD005331. [Google Scholar]
- Lynn, S.J.; Polizzi, C.; Merckelbach, H.; Chiu, C.D.; Maxwell, R.; van Heugten, D.; Lilienfeld, S.O. Dissociation and Dissociative Disorders Reconsidered: Beyond Sociocognitive and Trauma Models Toward a Transtheoretical Framework. Annu. Rev. Clin. Psychol. 2022. (Advance online publication). [Google Scholar] [CrossRef] [PubMed]
- Bremner, J.D.; Wittbrodt, M.T. Stress, the brain, and trauma spectrum disorders. Int. Rev. Neurobiol. 2020, 152, 1–22. [Google Scholar] [PubMed]
- Roydeva, M.I.; Reinders, A.A.T.S. Biomarkers of pathological dissociation: A systematic review. Neurosci. Biobehav. Rev. 2021, 123, 120–202. [Google Scholar] [CrossRef] [PubMed]
- Reinders, A.; Chalavi, S.; Schlumpf, Y.R.; Vissia, E.M.; Nijenhuis, E.; Jäncke, L.; Veltman, D.J.; Ecker, C. Neurodevelopmental origins of abnormal cortical morphology in dissociative identity disorder. Acta Psychiatr. Scand. 2018, 137, 157–170. [Google Scholar] [CrossRef] [Green Version]
- Radulovic, J.; Lee, R.; Ortony, A. State-dependent memory: Neurobiological advances and prospects for translation to dissociative Amnesia. Front. Behav. Neurosci. 2018, 12, 259. [Google Scholar] [CrossRef] [Green Version]
- Radulovic, J. Using new Approaches in neurobiology to rethink stress-induced amnesia. Curr. Behav. Neurosci. Rep. 2017, 4, 49–58. [Google Scholar] [CrossRef] [Green Version]
- Canan, F.; North, C.S. Dissociation and disasters: A systematic review. World J. Psychiatry. 2019, 9, 83–98. [Google Scholar] [CrossRef]
- Kizilhan, J.I.; Steger, F.; Noll-Hussong, M. Shame, dissociative seizures and their correlation among traumatised female Yazidi with experience of sexual violence. Br. J. Psychiatry. 2020, 216, 138–143. [Google Scholar] [CrossRef]
- Mai, F.M. “Hysteria” in clinical neurology. Can. J. Neurol. Sci. 1995, 22, 101–110. [Google Scholar] [CrossRef] [Green Version]
- Palomo, T.; Kostrzewa, R.M.; Beninger, R.J.; Archer, T. Genetic variation and shared biological susceptibility underlying comorbidity in neuropsychiatry. Neurotox. Res. 2007, 12, 29–42. [Google Scholar] [CrossRef]
- Jang, K.L.; Paris, J.; Zweig-Frank, H.; Livesley, W.J. Twin study of dissociative experience. J. Nerv. Ment. Dis. 1998, 186, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Becker-Blease, K.A.; Deater-Deckard, K.; Eley, T.; Freyd, J.J.; Stevenson, J.; Plomin, R. A genetic analysis of individual differences in dissociative behaviors in childhood and adolescence. J. Child Psychol. Psychiatry 2004, 45, 522–532. [Google Scholar] [CrossRef] [PubMed]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. PLoS Med. 2021, 18, e1003583. [Google Scholar] [CrossRef] [PubMed]
- Giesbrecht, T.; Merckelbach, H.; Geraerts, E. The dissociative experiences taxon is related to fantasy proneness. J. Nerv. Ment. Dis. 2007, 195, 769–772. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, J.R.; Sroufe, L.A.; Weinfield, N.S.; Carlson, E.A.; Egeland, B. Development and the fragmented self: Longitudinal study of dissociative symptomatology in a nonclinical sample. Dev. Psychopathol. 1997, 9, 855–879. [Google Scholar] [CrossRef] [Green Version]
- Espirito Santo, H.; Pio-Abreu, J.L. Demographic and mental health factors associated with pathological dissociation in a Portuguese sample. J. Trauma Dissociation 2008, 9, 369–387. [Google Scholar] [CrossRef]
- Spitzer, C.; Barnow, S.; Freyberger, H.J.; Grabe, H.J. Recent developments in the theory of dissociation. World Psychiatry 2006, 5, 82–86. [Google Scholar]
- Rodewald, F.; Dell, P.F.; Wilhelm-Gossling, C.; Gast, U. Are major dissociative disorders characterized by a qualitatively different kind of dissociation? J. Trauma Dissociation 2011, 12, 9–24. [Google Scholar] [CrossRef]
- Sohani, Z.N.; Meyre, D.; de Souza, R.J.; Joseph, P.G.; Gandhi, M.; Dennis, B.B.; Norman, G.; Anand, S.S. Assessing the quality of published genetic association studies in meta-analyses: The quality of genetic studies (Q-Genie) tool. BMC Genet. 2015, 16, 50. [Google Scholar] [CrossRef] [Green Version]
- Lochner, C.; Seedat, S.; Hemmings, S.M.; Kinnear, C.J.; Corfield, V.A.; Niehaus, D.J.; Moolman-Smook, J.C.; Stein, D.J. Dissociative experiences in obsessive-compulsive disorder and trichotillomania: Clinical and genetic findings. Compr. Psychiatry 2004, 45, 384–391. [Google Scholar] [CrossRef]
- Koenen, K.C.; Saxe, G.; Purcell, S.; Smoller, J.W.; Bartholomew, D.; Miller, A.; Hall, E.; Kaplow, J.; Bosquet, M.; Moulton, S.; et al. Polymorphisms in FKBP5 are associated with peritraumatic dissociation in medically injured children. Mol. Psychiatry 2005, 10, 1058–1059. [Google Scholar] [CrossRef]
- Lochner, C.; Seedat, S.; Hemmings, S.M.; Moolman-Smook, J.C.; Kidd, M.; Stein, D.J. Investigating the possible effects of trauma experiences and 5-HTT on the dissociative experiences of patients with OCD using path analysis and multiple regression. Neuropsychobiology 2007, 56, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Savitz, J.B.; van der Merwe, L.; Newman, T.K.; Solms, M.; Stein, D.J.; Ramesar, R.S. The relationship between childhood abuse and dissociation. Is it influenced by catechol-O-methyltransferase (COMT) activity? Int. J. Neuropsychopharmacol. 2008, 11, 149–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tadić, A.; Baskaya, O.; Victor, A.; Lieb, K.; Höppner, W.; Dahmen, N. Association analysis of SCN9A gene variants with borderline personality disorder. J. Psychiatr. Res. 2008, 43, 155–163. [Google Scholar] [CrossRef] [PubMed]
- McLean, S.A.; Diatchenko, L.; Lee, Y.M.; Swor, R.A.; Domeier, R.M.; Jones, J.S.; Jones, C.W.; Reed, C.; Harris, R.E.; Maixner, W.; et al. Catechol O-methyltransferase haplotype predicts immediate musculoskeletal neck pain and psychological symptoms after motor vehicle collision. J. Pain 2011, 12, 101–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pieper, S.; Out, D.; Bakermans-Kranenburg, M.J.; van Ijzendoorn, M.H. Behavioral and molecular genetics of dissociation: The role of the serotonin transporter gene promoter polymorphism (5-HTTLPR). J. Trauma. Stress 2011, 24, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Dackis, M.N.; Rogosch, F.A.; Oshri, A.; Cicchetti, D. The role of limbic system irritability in linking history of childhood maltreatment and psychiatric outcomes in low-income, high-risk women: Moderation by FK506 binding protein 5 haplotype. Dev. Psychopathol. 2012, 24, 1237–1252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmahl, C.; Arvastson, L.; Tamm, J.A.; Bohus, M.; Abdourahman, A.; Antonijevic, I. Gene expression profiles in relation to tension and dissociation in borderline personality disorder. PLoS ONE 2013, 8, e70787. [Google Scholar] [CrossRef]
- Wolf, E.J.; Rasmusson, A.M.; Mitchell, K.S.; Logue, M.W.; Baldwin, C.T.; Miller, M.W. A genome-wide association study of clinical symptoms of dissociation in a trauma-exposed sample. Depress. Anxiety 2014, 31, 352–360. [Google Scholar] [CrossRef] [Green Version]
- Reiner, I.; Frieling, H.; Beutel, M.; Michal, M. Gene-environment interaction of the oxytocin receptor gene polymorphism (rs53576) and unresolved attachment status predict depersonalization symptoms: An exploratory study. Psychol. Stud. 2016, 61, 295–300. [Google Scholar] [CrossRef]
- McCoy, T.H.; Castro, V.M.; Snapper, L.A.; Hart, K.L.; Perlis, R.H. Efficient genome-wide association in biobanks using topic modeling identifies multiple novel disease loci. Mol. Med. 2017, 23, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Yaylaci, F.T.; Cicchetti, D.; Rogosch, F.A.; Bulut, O.; Hetzel, S.R. The interactive effects of child maltreatment and the FK506 binding protein 5 gene (FKBP5) on dissociative symptoms in adolescence. Dev. Psychopathol. 2017, 29, 1105–1117. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Arcos-Burgos, M.; Licinio, J.; Wong, M.L. A latent genetic subtype of major depression identified by whole-exome genotyping data in a Mexican-American cohort. Transl. Psychiatry 2017, 7, e1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honma, M.; Yoshiike, T.; Ikeda, H.; Kuriyama, K. COMT genotype is associated with plasticity in sense of body ownership: A pilot study. Psychol. Res. 2018, 82, 634–644. [Google Scholar] [CrossRef] [PubMed]
- Kember, R.L.; Merikangas, A.K.; Verma, S.S.; Verma, A.; Judy, R.; Regeneron Genetics Center; Damrauer, S.M.; Ritchie, M.D.; Rader, D.J.; Bućan, M. Polygenic Risk of Psychiatric Disorders Exhibits Cross-trait Associations in Electronic Health Record Data from European Ancestry Individuals. Biol. Psychiatry 2021, 89, 236–245. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Lucas, A.M.; Zhang, X.; Chaudhary, K.; Cho, J.H.; Nadkarni, G.; Dobbyn, A.; Chittoor, G.; Josyula, N.S.; Katz, N.; et al. Exome-wide evaluation of rare coding variants using electronic health records identifies new gene-phenotype associations. Nat. Med. 2021, 27, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, H.; Takeuchi, H.; Kikuchi, Y.; Ono, C.; Yu, Z.; Iizuka, K.; Takano, Y.; Kakuto, Y.; Funakoshi, S.; Ono, T.; et al. Ethnicity-Dependent Effects of Schizophrenia Risk Variants of the OLIG2 Gene on OLIG2 Transcription and White Matter Integrity. Schizophr. Bull. 2020, 46, 1619–1628. [Google Scholar] [CrossRef]
- Ohi, K.; Shimada, T.; Yasuyama, T.; Uehara, T.; Kawasaki, Y. Variability of 128 schizophrenia-associated gene variants across distinct ethnic populations. Transl. Psychiatry 2017, 7, e988. [Google Scholar] [CrossRef]
- Alizadeh, N.; Nosrat, N.; Jahani, Z.; Ahmadiani, A.; Asadi, S.; Shams, J. Association of HTR1A gene polymorphisms with obsessive-compulsive disorder and its treatment response: The influence of sex and clinical characteristics. Int. J. Neurosci. 2019, 129, 264–272. [Google Scholar] [CrossRef]
- Shin, J.G.; Kim, J.H.; Park, C.S.; Kim, B.J.; Kim, J.W.; Choi, I.G.; Hwang, J.; Shin, H.D.; Woo, S.I. Gender-Specific Associations between CHGB Genetic Variants and Schizophrenia in a Korean Population. Yonsei Med. J. 2017, 58, 619–625. [Google Scholar] [CrossRef]
- Lee, P.H.; Feng, Y.A.; Smoller, J.W. Pleiotropy and Cross-Disorder Genetics Among Psychiatric Disorders. Biol. Psychiatry 2021, 89, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Bandelow, B.; Baldwin, D.; Abelli, M.; Bolea-Alamanac, B.; Bourin, M.; Chamberlain, S.R.; Cinosi, E.; Davies, S.; Domschke, K.; Fineberg, N.; et al. Biological markers for anxiety disorders, OCD and PTSD: A consensus statement. Part II: Neurochemistry, neurophysiology and neurocognition. World J. Biol. Psychiatry 2017, 18, 162–214. [Google Scholar] [CrossRef] [PubMed]
- Hawn, S.E.; Sheerin, C.M.; Lind, M.J.; Hicks, T.A.; Marraccini, M.E.; Bountress, K.; Bacanu, S.A.; Nugent, N.R.; Amstadter, A.B. GxE effects of FKBP5 and traumatic life events on PTSD: A meta-analysis. J. Affect. Disord. 2019, 243, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.Y.; Wu, Y.L.; Cheng, C.H.; Liu, X.X.; Zhou, L. Association of Brain-Derived Neurotrophic Factor rs6265 G > A polymorphism and Post-traumatic Stress Disorder susceptibility: A systematic review and meta-analysis. Brain Behav. 2021, 11, e02118. [Google Scholar] [CrossRef]
- Koenen, K.C. Nature-nurture interplay: Genetically informative designs contribute to understanding the effects of trauma and interpersonal violence. J. Interpers. Violence 2005, 20, 507–512. [Google Scholar] [CrossRef]
- McKinney, K.A.; Lange, M.M. Familial fugue—A case report. Can. J. Psychiatry 1983, 28, 654–656. [Google Scholar] [CrossRef]
- Harkness, D. Testing Cernak’s hypothesis: Is dissociation the mediating variable that links substance abuse in the family of origin with offspring codependency? J. Psychoactive Drugs 2001, 33, 75–82. [Google Scholar] [CrossRef]
- Domozych, W.; Dragan, W.L. Genetic and environmental basis of the relationship between dissociative experiences and Cloninger’s temperament and character dimensions—A pilot study. Pol. Psychol. Bull. 2016, 47, 412–420. [Google Scholar] [CrossRef]
- Grabe, H.-J.; Spitzer, C.; Freyberger, H.J. Relationship of dissociation to temperament and character in men and women. Am. J. Psychiatry 1999, 156, 1811–1813. [Google Scholar]
- Waller, N.G.; Ross, C.A. The prevalence and biometric structure of pathological dissociation in the general population: Taxometric and behavior genetic findings. J. Abnorm. Psychol. 1997, 106, 499–510. [Google Scholar] [CrossRef]
- Bracha, H.S. Human brain evolution and the “Neuroevolutionary Time-depth Principle”: Implications for the Reclassification of fear-circuitry-related traits in DSM-V and for studying resilience to warzone-related posttraumatic stress disorder. Prog. Neuropsychopharmacol. Biol. Psychiatry 2006, 30, 827–853. [Google Scholar] [CrossRef] [PubMed]
- Şar, V. The many faces of dissociation: Opportunities for innovative research in psychiatry. Clin. Psychopharmacol. Neurosci. 2014, 12, 171–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasmusson, A.M.; Pineles, S.L. Neurotransmitter, Peptide, and Steroid Hormone Abnormalities in PTSD: Biological Endophenotypes Relevant to Treatment. Curr. Psychiatry Rep. 2018, 20, 52. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Mateu, F.; Escámez, T.; Koenen, K.C.; Alonso, J.; Sánchez-Meca, J. Meta-analyses of the 5-HTTLPR polymorphisms and post-traumatic stress disorder. PLoS ONE 2013, 8, e66227. [Google Scholar] [CrossRef]
- Suliman, S.; Hemmings, S.M.; Seedat, S. Brain-Derived Neurotrophic Factor (BDNF) protein levels in anxiety disorders: Systematic review and meta-regression analysis. Front. Integr. Neurosci. 2013, 7, 55. [Google Scholar] [CrossRef] [Green Version]
- Mendonça, M.S.; Mangiavacchi, P.M.; Rios, Á. Regulatory functions of FKBP5 intronic regions associated with psychiatric disorders. J. Psychiatr. Res. 2021, 143, 1–8. [Google Scholar] [CrossRef]
- Gulsun, M.; Doruk, A.; Uzun, O.; Turkbay, T.; Ozsahin, A. Effect of dissociative experiences on drug treatment of panic disorder. Clin. Drug Investig. 2007, 27, 583–590. [Google Scholar] [CrossRef]
- Bob, P.; Susta, M.; Gregusova, A.; Jasova, D.; Raboch, J.; Mishara, A. Traumatic stress, dissociation, and limbic irritability in patients with unipolar depression being treated with SSRIs. Psychol. Rep. 2010, 107, 685–696. [Google Scholar] [CrossRef]
- Simeon, D.; Hollander, E.; Stein, D.J.; DeCaria, C.; Cohen, L.J.; Saoud, J.B.; Islam, N.; Hwang, M. Induction of depersonalization by the serotonin agonist meta-chlorophenylpiperazine. Psychiatry Res. 1995, 58, 161–164. [Google Scholar] [CrossRef]
- Lacy, T.; Mathis, M. Dissociative symptoms from combined treatment with sertraline and trazodone. J. Neuropsychiatr. Clin. Neurosci. 2003, 15, 241–242. [Google Scholar] [CrossRef]
- Kawai, N.; Honda, M.; Nakamura, S.; Samatra, P.; Sukardika, K.; Nakatani, Y.; Shimojo, N.; Oohashi, T. Catecholamines and opioid peptides increase in plasma in humans during possession trances. Neuroreport 2001, 12, 3419–3423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boulet, C.; Lopez-Castroman, J.; Mouchabac, S.; Olié, E.; Courtet, P.; Thouvenot, E.; Abbar, M.; Conejero, I. Stress response in dissociation and conversion disorders: A systematic review. Neurosci. Biobehav. Rev. 2022, 132, 957–967. [Google Scholar] [CrossRef] [PubMed]
- Procopio, D.O.; Saba, L.M.; Walter, H.; Lesch, O.; Skala, K.; Schlaff, G.; Vanderlinden, L.; Clapp, P.; Hoffman, P.L.; Tabakoff, B. Genetic markers of comorbid depression and alcoholism in women. Alcohol. Clin. Exp. Res. 2013, 37, 896–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Cao, H.; Kural, K.C.; Fang, Q.; Zhang, F. Integrative analysis of shared genetic pathogenesis by autism spectrum disorder and obsessive-compulsive disorder. Biosci. Rep. 2019, 39, BSR20191942. [Google Scholar] [CrossRef] [Green Version]
- de Mooij-van Malsen, A.J.; van Lith, H.A.; Oppelaar, H.; Hendriks, J.; de Wit, M.; Kostrzewa, E.; Breen, G.; Collier, D.A.; Olivier, B.; Kas, M.J. Interspecies trait genetics reveals association of Adcy8 with mouse avoidance behavior and a human mood disorder. Biol. Psychiatry 2009, 66, 1123–1130. [Google Scholar] [CrossRef]
- Prontera, P.; Napolioni, V.; Ottaviani, V.; Rogaia, D.; Fusco, C.; Augello, B.; Serino, D.; Parisi, V.; Bernardini, L.; Merla, G.; et al. DPP6 gene disruption in a family with Gilles de la Tourette syndrome. Neurogenetics 2014, 15, 237–242. [Google Scholar] [CrossRef]
- Lin, L.; Murphy, J.G.; Karlsson, R.M.; Petralia, R.S.; Gutzmann, J.J.; Abebe, D.; Wang, Y.X.; Cameron, H.A.; Hoffman, D.A. DPP6 Loss Impacts Hippocampal Synaptic Development and Induces Behavioral Impairments in Recognition, Learning and Memory. Front. Cell. Neurosci. 2018, 12, 84. [Google Scholar] [CrossRef] [Green Version]
- Hébert, M.; Langevin, R.; Charest, F. Disorganized attachment and emotion dysregulation as mediators of the association between sexual abuse and dissociation in preschoolers. J. Affect. Disord. 2020, 267, 220–228. [Google Scholar] [CrossRef]
- Lewis, J.; Binion, G.; Rogers, M.; Zalewski, M. The Associations of Maternal Emotion Dysregulation and Early Child Dissociative Behaviors. J. Trauma Dissociation 2020, 21, 203–216. [Google Scholar] [CrossRef]
- Golanska, E.; Sieruta, M.; Gresner, S.M.; Hulas-Bigoszewska, K.; Corder, E.H.; Styczynska, M.; Peplonska, B.; Barcikowska, M.; Liberski, P.P. Analysis of APBB2 gene polymorphisms in sporadic Alzheimer’s disease. Neurosci. Lett. 2008, 447, 164–166. [Google Scholar] [CrossRef]
- Günak, M.M.; Billings, J.; Carratu, E.; Marchant, N.L.; Favarato, G.; Orgeta, V. Post-traumatic stress disorder as a risk factor for dementia: Systematic review and meta-analysis. Br. J. Psychiatry 2020, 217, 600–608. [Google Scholar] [CrossRef] [PubMed]
- Bob, P.; Raboch, J.; Maes, M.; Susta, M.; Pavlat, J.; Jasova, D.; Vevera, J.; Uhrova, J.; Benakova, H.; Zima, T. Depression, traumatic stress and interleukin-6. J. Affect. Disord. 2010, 120, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Hori, H.; Kim, Y. Inflammation and post-traumatic stress disorder. Psychiatry Clin. Neurosci. 2019, 73, 143–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michopoulos, V.; Beurel, E.; Gould, F.; Dhabhar, F.S.; Schultebraucks, K.; Galatzer-Levy, I.; Rothbaum, B.O.; Ressler, K.J.; Nemeroff, C.B. Association of Prospective Risk for Chronic PTSD Symptoms with Low TNFα and IFNγ Concentrations in the Immediate Aftermath of Trauma Exposure. Am. J. Psychiatry 2020, 177, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Bertule, M.; Sebre, S.B.; Kolesovs, A. Childhood abuse experiences, depression and dissociation symptoms in relation to suicide attempts and suicidal ideation. J. Trauma Dissociation 2021, 22, 598–614. [Google Scholar] [CrossRef]
- Caspi, A.; Hariri, A.R.; Holmes, A.; Uher, R.; Moffitt, T.E. Genetic sensitivity to the environment: The case of the serotonin transporter gene and its implications for studying complex diseases and traits. Am. J. Psychiatry 2010, 167, 509–527. [Google Scholar] [CrossRef]
- Cattaneo, A.; Cattane, N.; Begni, V.; Pariante, C.M.; Riva, M.A. The human BDNF gene: Peripheral gene expression and protein levels as biomarkers for psychiatric disorders. Transl. Psychiatry 2016, 6, e958. [Google Scholar] [CrossRef]
- Rafiq, S.; Campodonico, C.; Varese, F. The relationship between childhood adversities and dissociation in severe mental illness: A meta-analytic review. Acta Psychiatr. Scand. 2018, 138, 509–525. [Google Scholar] [CrossRef]
- Zerubavel, N.; Messman-Moore, T.L.; DiLillo, D.; Gratz, K.L. Childhood Sexual Abuse and Fear of Abandonment Moderate the Relation of Intimate Partner Violence to Severity of Dissociation. J. Trauma Dissociation 2018, 19, 9–24. [Google Scholar] [CrossRef]
- Garcia-Esteve, L.; Torres-Gimenez, A.; Canto, M.; Roca-Lecumberri, A.; Roda, E.; Velasco, E.R.; Echevarría, T.; Andero, R.; Subirà, S. Prevalence and risk factors for acute stress disorder in female victims of sexual assault. Psychiatry Res. 2021, 306, 114240. [Google Scholar] [CrossRef]
- Stone, J.; LaFrance Jr, C.; Brown, R.; Spiegel, D.; Levenson, J.L.; Sharpe, M. Conversion disorder: Current problems and potential solutions for DSM-5. J. Psychosom. Res. 2011, 71, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Agargun, M.Y.; Ozer, O.A.; Kara, H.; Sekeroglu, R.; Selvi, Y.; Eryonucu, B. Serum lipid levels in patients with dissociative disorder. Am. J. Psychiatry 2004, 161, 2121–2123. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.K.; Ratanatharathorn, A.; Maihofer, A.X.; Naviaux, R.K.; Aiello, A.E.; Amstadter, A.B.; Ashley-Koch, A.E.; Baker, D.G.; Beckham, J.C.; Boks, M.P.; et al. Epigenome-wide meta-analysis of PTSD across 10 military and civilian cohorts identifies methylation changes in AHRR. Nat. Commun. 2020, 11, 5965. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
PRISMA flow diagram for the current systematic review.


{kind=link}
Table 1.
Summary of studies included in the current review.
| Authors | Study Population and Sample Size | Study Design | Phenotype | Polymorphisms Studied, if Applicable | Study Quality-Overall | Study Quality–Bias | Study Results |
|---|---|---|---|---|---|---|---|
| Lochner et al., 2004 [40] | Patients with obsessive–compulsive disorder or trichotillomania (n = 114); Caucasian ethnicity | Association | Dissociative symptoms, total | DRD4 48-bp VNTR, DAT 40-bp VNTR, 5HTTLPR, HTR1B G861C, HTR2C T102C, TPH1 Val81Met, COMT rs4680, MAOA C1460T polymorphisms | Moderate | Poor | No association between dissociation and any of the studied polymorphisms. |
| Koenen et al., 2005 [41] | Children with acute injuries (n = 46) | Association | Dissociative symptoms, total | FKBP5 rs3800373, rs1360870 SNPs | Poor | Poor | FKBP5 rs3800373 C allele and rs1360870 T allele significantly associated with dissociation during and after injury |
| Lochner et al., 2007 [42] | Patients with obsessive–compulsive disorder; Caucasian ethnicity (n = 83) | Association, G×E (childhood maltreatment) | Dissociative symptoms, total | 5-HTTLPR polymorphism | Moderate | Good | Interaction between 5-HTT s (particularly s/s genotype) and childhood trauma associated with dissociation |
| Savitz et al., 2008 [43] | Patients with bipolar disorder and their relatives (n = 178) | Association, G×E (childhood trauma) | Dissociative symptoms, total | COMT rs4680, DRD4 48-bp VNTR, BDNF Val66Met, 5-HTTLPR, DAT 3’-VNTR | Moderate | Poor | BDNF Met associated with lower self-reported dissociation; interaction between COMT Val and childhood trauma associated with dissociation |
| Tadic et al., 2009 [44] | Patients with borderline personality disorder (n = 161) and healthy controls (n = 156); Caucasian ethnicity | Association with control group | Dissociative symptoms, total | SCN9A rs16851799, rs7607967, rs4371369, rs4597545, rs4387806, rs6754031, rs12620053, rs13017637, rs12994338, rs4447616 SNPs and haplotype | Good | Good | Possible association between SCN9A rs7607967 (G) and dissociation; possible interaction between rs4371369 (G) and rs4387806 (C), sex, and dissociation |
| McLean et al., 2011 [45] | Motor vehicle accident victims; Caucasian ethnicity (n = 89) | Association | Dissociative symptoms, total | COMT rs4633, rs4680, rs4818 and rs6269 haplotype | Moderate | Good | Association between “pain-vulnerable” COMT haplotype (A_C_C_G) and dissociative symptoms following trauma. |
| Pieper et al., 2011 [46] | Adult twin pairs (n = 184) | Association, G×E (traumatic stress) | Dissociative symptoms, total | 5-HTTLPR genotype | Moderate | Good | 5-HTTLPR s/s genotype associated with dissociation in general; s/s genotype associated with pathological dissociation in those with a history of trauma |
| Dackis et al., 2012 [47] | High-risk, low-income women with (n = 170) and without (n = 66) childhood maltreatment | G×E (childhood maltreatment) with control group | Dissociative symptoms, total | FKBP5 rs3800373, rs9296158, rs1360870, rs9470080 haplotype | Good | Good | Interaction between FKBP5 CATT haplotype and childhood trauma associated with dissociation |
| Schmahl et al., 2013 [48] | Women hospitalized for borderline personality disorder (n = 31) | Gene expression | Dissociative symptoms, total | 29 genes selected based on prior associations with depression | Moderate | Good | IL6 gene expression positively correlated with dissociation; IL1B, MAPK1, MAPK3, MAPK8, GNAI2, ARRB1, ARRB2, CREB1 expression negatively correlated with dissociation |
| Wolf et al., 2014 [49] | Adults with a history of trauma exposure (n = 484); Caucasian ethnicity | Genome-wide association | Depersonalization / derealization symptoms | Not applicable | Good | Good | No genome-wide significant associations; ten suggestive associations with depersonalization / derealization; highest peaks at ADCY8 rs263232 and DPP6 rs71534169; no replication of earlier associations with 5-HTTLPR, COMT or FKBP5 |
| Reiner et al., 2016 [50] | Pre-menopausal women with depression (n = 43) and healthy controls (n = 41); Caucasian ethnicity | Association, G×E (unresolved attachment) with control group | Depersonalization / derealization symptoms | OXTR rs53576 (A/G) SNP | Moderate | Good | Trend towards higher depersonalization / derealization symptoms in women with the OXTR rs53576 GG genotype; interaction between OXTR GG genotype and unresolved attachment associated with dissociation. |
| McCoy et al., 2017 [51] | Patients from academic medical centers participating in biobanking programs (n = 10845), Northern European ethnicity | Genome-wide association | Anxiety, phobic and dissociative disorders | Not applicable | Good | Poor | Possible association between the group “anxiety, phobic and dissociative disorders” and locus on chromosome 4 containing the APBB2 gene. |
| Yaylaci et al., 2017 [52] | Low-income adolescents with (n = 279) and without (n = 171) childhood maltreatment | G×E interaction (childhood maltreatment) with control group | Dissociative symptoms, total | FKBP5 rs3800373, rs9296158, rs1360870, rs9470080 haplotype | Good | Good | Possible protective effect of FKBP5 CATT haplotype on dissociation in those with an early-onset and longer duration of maltreatment |
| Yu et al., 2017 [53] | Patients with depression (n = 203) and healthy controls (n = 196); Mexican-American ethnicity | Association with control group | Depersonalization/derealization symptoms | 19 SNPs identified in a prior study: rs41310573, rs201935337, rs140395831, rs56293203, rs78562453, rs115054458, rs143696449, rs748441912, rs62001028, rs150952348, rs782472239, rs112610420, rs142029931, rs201483250, rs200897153, rs3744550, rs115668237, rs56344012 rs200520741 | Moderate | Good | Evidence of a latent depressive subtype associated with 19 SNPs, associated with lower depersonalization / derealization scores |
| Honma et al., 2018 [54] | Normal individuals (n = 76), Japanese ethnicity | Association | Dissociative symptoms, total | COMT rs4680 genotype | Moderate | Good | COMT rs4680 Val/Val genotype associated with total dissociative symptoms and depersonalization/derealization symptoms but not dissociative amnesia symptoms |
| Kember et al., 2021 [55] | Patients from an academic medical center participating in a biobanking program (n = 10182); European ethnicity | Polygenic risk score | Anxiety, phobic and dissociative disorders | PRS for six common psychiatric disorders (schizophrenia, bipolar disorder, depression, attention deficit-hyperactivity disorder and anorexia nervosa) | Good | Good | PRS for depression and bipolar disorder both significantly associated with “anxiety, phobic and dissociative disorders” |
| Park et al., 2021 [56] | Patients from an academic medical center participating in a biobanking program (n = 10,845) | Exome-wide association | Anxiety, phobic and dissociative disorders | Not applicable | Good | Good | No association identified for the group “anxiety, phobic and dissociative disorders” |
Abbreviations: 5-HTT, serotonin transporter gene; 5-HTTLPR, serotonin transporter gene promoter region polymorphism; ADCY8, adenylyl cyclase type 8 gene; APBB2, amyloid beta-4 precursor protein-binding family B member 2 gene; ARRB1, arrestin beta-1 gene; ARRB2, arrestin beta-2 gene; BDNF, brain-derived neurotrophic factor gene; COMT, catechol O-methyltransferase gene; CREB1, cyclic AMP response element binding protein 1 gene; DAT, dopamine transporter gene; DPP6, dipeptidyl peptidase 6 gene; DRD4, dopamine type 4 receptor gene; FKBP5, FK506 binding protein 5 gene; GNAI2, G-protein subunit alpha I2 gene; HTR1B, serotonin type 1B receptor gene; HTR2C, serotonin type 2C receptor gene; IL1B, interleukin-1 beta gene; IL6, interleukin-6 gene; MAOA, monoamine oxidase A gene; MAPK1, mitogen-activated protein kinase 1 gene; MAPK3, mitogen-activated protein kinase 3 gene; MAPK8, mitogen-activated protein kinase 8 gene; OXTR, oxytocin receptor gene; PRS, polygenic risk score; SCN9A, sodium voltage-gated channel alpha subunit 9; SNP, single nucleotide polymorphism; TPH1, tryptophan hydroxylase 1 gene; VNTR, variable number of tandem repeats.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Rajkumar, R.P. The Molecular Genetics of Dissociative Symptomatology: A Transdiagnostic Literature Review. Genes 2022, 13, 843. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13050843
AMA Style
Rajkumar RP. The Molecular Genetics of Dissociative Symptomatology: A Transdiagnostic Literature Review. Genes. 2022; 13(5):843. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13050843
Chicago/Turabian StyleRajkumar, Ravi Philip. 2022. "The Molecular Genetics of Dissociative Symptomatology: A Transdiagnostic Literature Review" Genes 13, no. 5: 843. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13050843
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.