
Complete Mitochondrial Genome of Malenka flexura (Plecoptera: Nemouridae) and Phylogenetic Analysis


1
College of Plant Protection, Henan Agricultural University, Zhengzhou 450002, China
2
Postdoctoral Research Base, Henan Institute of Science and Technology, Xinxiang 453003, China
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Genes 2022, 13(5), 911; https://0-doi-org.brum.beds.ac.uk/10.3390/genes13050911
Submission received: 6 May 2022
/
Revised: 15 May 2022
/
Accepted: 18 May 2022
/
Published: 19 May 2022
(This article belongs to the Special Issue Phylogeny and Genetic Diversity of Insects)
Abstract
:The genus-level relationships within the subfamily Amphinemurinae have been controversial, although attempts have been made based on morphology and limited molecular data. With the establishment of two new genera, the phylogenetic relationships within Amphinemurinae should be re-examined. In this study, the complete mitochondrial genome (mitogenome) of Malenka flexura of the genus Malenka was firstly sequenced and analyzed. The phylogeny of Amphinemurinae was also reconstructed using 13 proteincoding genes (PCGs) from previously published stoneflies. This mitogenome was 15,744 bp long and encoded the typical 37 genes, as well as a putative control region. The gene arrangement of M. flexura mitogenome is identical with the putative ancestral mitogenome in Drosophila yakuba. Most PCGs used standard ATN as start codons and TAA/TAG as termination codons. All tRNA genes exhibited the typical cloverleaf secondary structure, except for tRNASer(AGN), whose dihydrouridine (DHU) arm was lacking. Some structural elements in the control region were founded, such as tandem repeat regions, stemloop structures, polyN stretch and microsatellite structure, etc. Phylogenetic analyses of sequenced Amphinemurinae mitogenomes unsupported the sister relationship of Amphinemura and Malenka. Finally, the phylogenetic analyses inferred a relationship within Amphinemurinae: Amphinemura + (Malenka + (Protonemura + (Indonemoura + (Sphaeronemoura + Mesonemoura)))).
1. Introduction
The mitochondrial genome (mitogenome) is a complete and relatively independent organelle genome, which contains complete genetic information from molecular sequence to gene structure [1,2]. Insect mitogenome typically constitutes 14–20 kb circular DNA molecules. It encodes 13 protein-coding genes (PCGs), two ribosomal genes (rRNAs), and 22 transfer RNA genes (tRNAs) [1,3]. It also has an A + T-rich region (or control region, CR) that regulates the transcription and replication of the mitogenome [3]. In recent years, the mitogenome has become a major resource for investigating biogeography, species evolution, population genetics structure, and phylogeny of different classification elements [4,5,6,7], because of its small genome size, rare recombination, rapid mutation rate, maternal inheritance, and conserved gene content [1,8].
Stoneflies (Insecta: Plecoptera) are a group of hemimetabolous aquatic insects that are distributed around the world except for Antarctica [9,10]. Currently, over 4000 extant species are described in the order Plecoptera, which are divided into 17 families [9,10,11]. Stoneflies are most commonly associated with clean, cool running water and cool, wet terrestrial environments [10]. The nymphs congregate in riffle areas of streams with an abundance of boulders, gravel, snags, and piled leaves. It is commonly recognized that they could be used as biological indicators of water quality [12].
Nemouridae is one of the largest Plecoptera families, with over 400 species distributed across the nearctic, palearctic, and oriental regions [13]. The nymphs are distinguished by their broad, bristly bodies and divergent wing pads. They can be found in a wide range of streams, but smaller creeks and spring runs are probably the most diverse [9]. Baumann considered Amphinemurinae as a new subfamily of Nemouridae based on the number of lobes on parprocts, and divided this subfamily into five genera, Amphinemura, Indonemoura, Malenka, Mesonemoura and Protonemura [13]. Recently, Sphaeronemoura and Tominemoura were proposed as two new genera in this subfamily [14,15]. Therefore, the Amphinemurinae presently includes seven described genera worldwide.
So far, Baumann’s morphological analyses of the phylogenetic relationships within the Nemouridae are thought to be the most comprehensive [13]. At the subfamily level, the Amphinemurinae is recognized as monophyletic, and the relationship within this subfamily was recovered as (Amphinemura + Malenka) + (Protonemura + (Indonemoura + Mesonemoura)). Early molecular phylogeny of stoneflies has been studied using a single nucleotide sequence [16] and six molecular markers [17]. Both studies used fewer genera of this subfamily, and the results did not support the sister group relationship of Amphinemura and Malenka. Recently, phylogenies for Amphinemurinae fauna were proposed using mitochondrial genomic data, and those results showed that the positions of Mesonemoura, Indonemoura and Protonemura were similar to the traditional morphology-based results [18,19,20]. However, only sequences from five genera of Amphinemurinae species (Amphinemura, Protonemura, Indonemoura, Mesonemoura and Sphaeronemoura) were included in those previous studies.
To better resolve the phylogenetic relationship within this subfamily, more Amphinemurinae mitogenomes, particularly those from Malenka and Tominemoura species, should be obtained. Because the genus Tominemoura has only one described species (Tominemoura trilari), and this species is only found in Sabah, Malaya, the specimen of T. trilari was unable to be obtained. In this study, one complete mitogenome of genera Malenka (Malenka flexura) was sequenced, and its nucleotide compositions, codon usage and RNA structures were analyzed. Finally, the phylogenetic analyses of Amphinemurinae were performed based on the nucleotide sequences of available stonefly mitogenomes. The aim of this research is to improve the understanding of the phylogeny of Amphinemurinae.
2. Materials and Methods
2.1. Sample Collection and DNA Extraction
Wild specimens of M. flexura were collected from Albany in New York, USA, and the voucher specimen for this species (No. VHL-0135) was deposited in the Department of Entomology, Henan Institute of Science and Technology, China. Specimens used in this study were preserved in 100% ethanol and stored at −20 °C. Total genomic DNA was isolated from the thoracic muscle of adults using the DNeasy Blood & Tissue Kit (Qiagen, Hilden, Germany).
2.2. Sequencing and Bioinformatics Analyses
Illumina Hiseq 2500 with 500 cycles of paired-end sequencing (250 bp reads) was performed at Berry Genomics Co., Ltd., Beijing, China. The mitogenome of M. flexura was sequenced and amplified as described in previous studies [18,19,21,22,23]. Illumina sequence reads were assembled into contigs with Geneious 6.1.6 [24]. The tRNA genes were initially identified using the MITOS webservers [25]. The boundaries of PCGs and rRNA genes were identified by comparing with the homologous genes of other published stonefly species. Base composition, codon usage and the relative synonymous codon usage (RSCU) were calculated using MEGA 6.0 [26]. Composition skew analysis was performed using the AT-skew = [A − T]/[A + T] and GC-skew = [G − C]/[G + C] formulas [27]. Stem-loop structures in the control region were predicted by DNAMAN, and the tandem repeat units were identified using the Tandem Repeats Finder server (http://tandem.bu.edu/trf/trf.advanced.submit.html, accessed on 1 May 2022) [28].
2.3. Phylogenetic Analyses
A total of 17 nemourid species were used for the phylogenetic analysis, including 14 Amphinemurinae species and three outgroup species from the subfamily Nemourinae (Table 1). In total, 13 PCGs in the 17 species were aligned using the MAFFT algorithm [29] in the TranslatorX online platform [30]. The alignment of individual genes was concatenated together to make the PCG dataset (including 13 PCGs) after removing ambiguously aligned positions.
Phylogenetic trees were reconstructed based on the PCG dataset under the maximum likelihood (ML) and Bayesian inference (BI) methods. According to the Akaike information criterion (AIC), the best-fit model GTR + I+G for the dataset was determined using ModelFinder [31]. ML analysis was performed using IQ-TREE Web Server [31] with 10,000 bootstrap replicates. BI analysis was carried out with MrBayes 3.2.6 [32] under the following conditions: 10 million generations with sampling every 1000 generations, four independent Markov chains, and a burn-in of 25% trees.
3. Results and Discussion
3.1. Mitogenome Organization and Base Composition
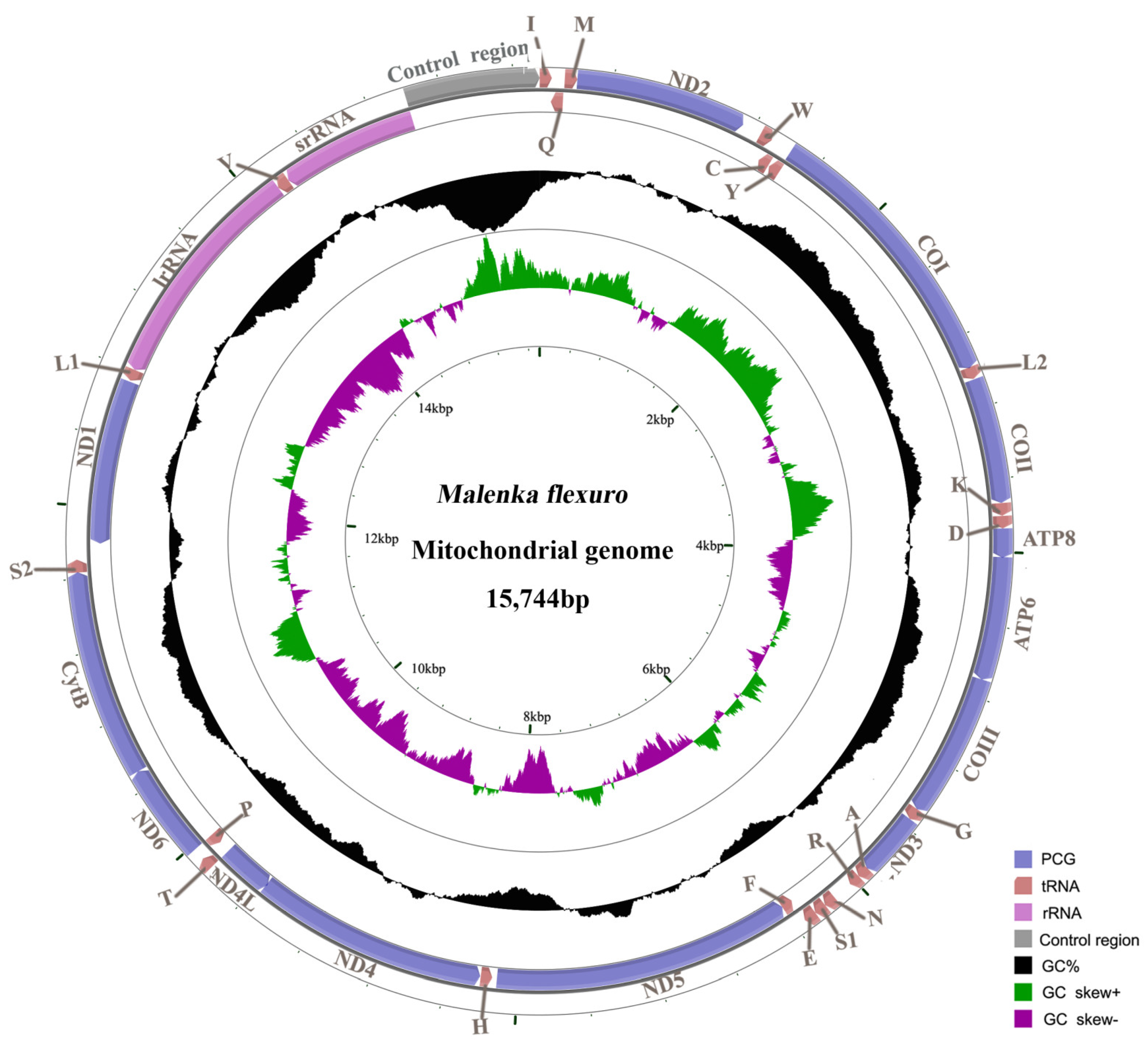
The complete mitogenome of M. flexura is 15,744 bp in length (GenBank accession number ON411527; Figure 1), which is consistent with other sequenced Amphinemurinae species [18]. It is a double-stranded circular molecule, including 13 PCGs, 22 tRNA genes, 2 rRNA genes and a large non-coding region (control region) (Table 2 and Figure 1). The gene order of the M. flexura mitogenome is identical to other sequenced stoneflies and the model insect, Drosophila yakuba [33]. There are 51 overlapping nucleotides distributed in 13 gene junctions; the tRNATrp/tRNACys and tRNATyr/COI gene junctions possess the longest overlap (8 bp). The ATP8/ATP6 and ND4/ND4L gene junctions overlap seven nucleotides (ATGNTAA), and are often found across the Metazoa [34,35]. Except for the large non-coding region, there are 225 nucleotides dispersed in 11 intergenic spacers, ranging in size from 1 to 111 bp. (Table 2).
The overall nucleotide composition of the M. flexura mitogenome is 32.3% A, 18.8% G, 36.3% T and 12.5% C, respectively (Table 3). The A + T content of the whole mitogenome, PCGs, tRNAs, rRNAs and the control region is 68.6%, 66.6%, 70.9%, 71.9% and 85.2% (Table 3). Therefore, the nucleotide composition of the M. flexura mitogenome is biased toward A and T nucleotides, and the control region is usually considered the most A + T rich in stonefly mitogenomes [18,19,20,21,22,23].
The nucleotide composition of metazoan mitogenomes usually has a clear strand bias [36,37], which can be measured as AT- and GC-skews [27]. In this study, the M. flexura mitogenome shows a negative AT-skew and a positive GC-skew (Table 3), revealing a bias in the use of T and G nucleotides. For the J-strand, most insect mitogenomes show a positive AT-skew and negative GC-skew [38], while results of this study show a negative AT-skew of PCGs and a positive GC-skew of tRNA genes. Like the M. flexura mitogenome, the strand bias of some other stonefly mitogenomes is also inconsistent with that of most other insects (positive AT skew and negative GC skew for the J-strand) [18,21,39,40]. The balance between mutational and selection pressures during replication and transcription may cause nucleotide compositional asymmetries, which might serve as a possible signal for replication orientation and gene direction [27,38].
3.2. Protein-Coding Genes and Codon Usage
Most PCGs of M. flexura use ATN as the start codon, such as ATT (4 PCGs), and ATG (7 PCGs). However, two exceptions, ND5 and ND1 genes, initiate with GTG and TTG as a start codon, respectively (Table 2). The use of these two nonstandard start codons is also found in other Amphinemurinae species [18]. In some species, TTG is also employed to shorten intergenic spacer and avoid gene overlap [41,42]. Eight PCGs (ND2, COI, ATP8, ATP6, COIII, ND4, ND4L and ND6) terminate with the stop codon TAA, three PCGs (ND3, CytB and ND1) terminate with TAG, and two PCGs (COII and ND5) end with incomplete stop codon T (Table 2). The use of an incomplete stop codon (T) is common in stoneflies [18,20,21,22,23,39,40,43] and animal mitogenomes [3], and can form a complete TAA terminal signal by post-transcriptional polyadenylation [44,45].
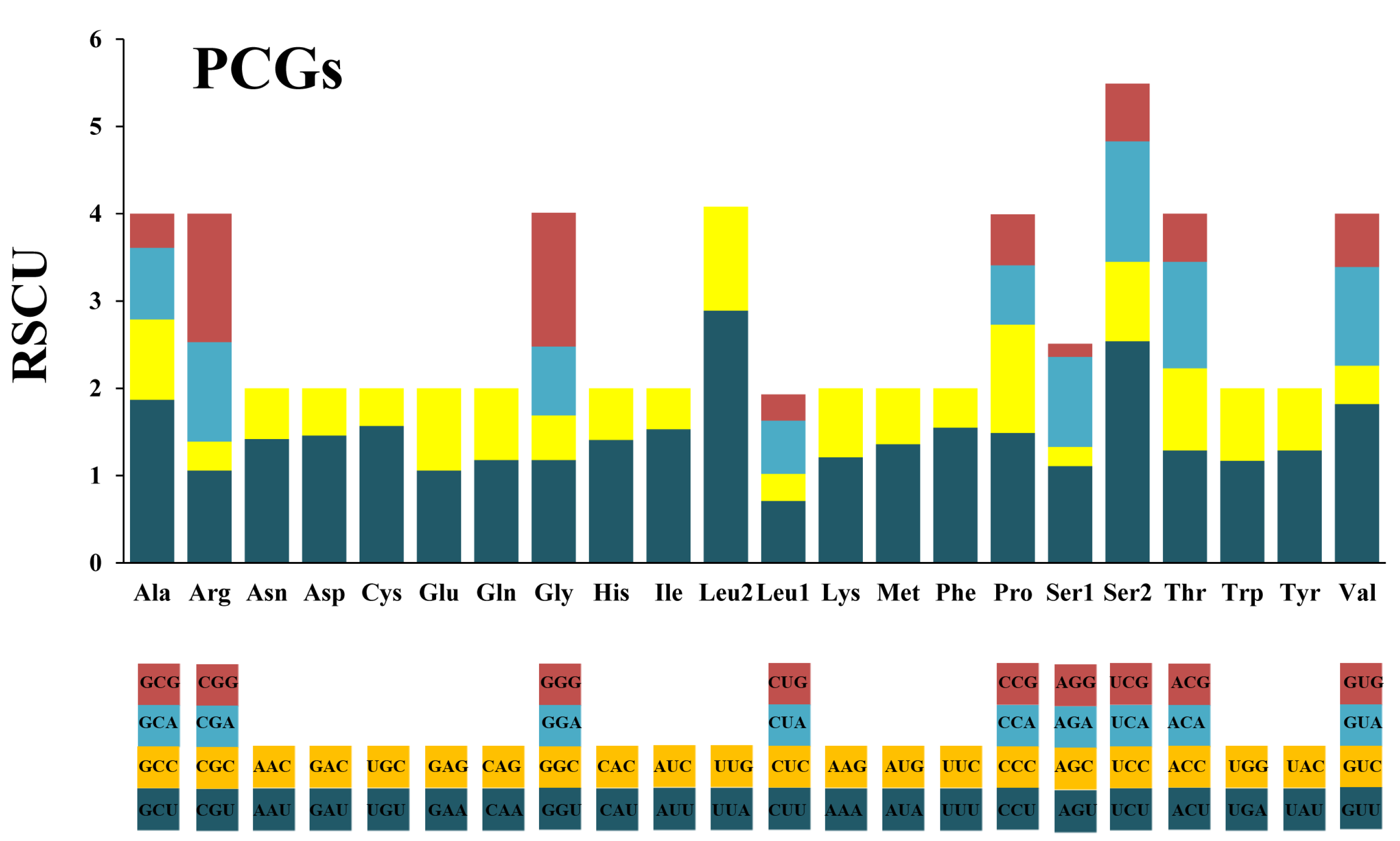
The influence of a strong biased codon usage is reflected by the relative synonymous codon usage (RSCU) [46]. The result shows that both two-fold and four-fold degenerate codons are preferable over codons ending with A or U (Figure 2). Another five prevalent AT-rich codons (TTA, ATT, TTT, ATA and AAT) also contribute to the compositional biases for AT (Table 4).
3.3. Transfer and Ribosomal RNA Genes
The lengths of tRNAs are ranged from 63 bp to 71 bp (Table 2). All tRNA genes exhibit the typical cloverleaf secondary structure, except for tRNASer(AGN), whose dihydrouridine (DHU) arm is lacking (Figure S1). According to the secondary structure of M. flexura tRNA genes, there are 42 unmatched base pairs in these tRNAs. Thirty-four of these are weak G-U pairs, which are in acceptor arms (9 bp), DHU arms (10 bp), anticodon arms (12 bp), and TΨC arms (3 bp). The remaining are U-U (1 bp), A-G (2 bp), U-C (3 bp) and A-C (2 bp) mismatches (Figure S1).
The large rRNA subunit gene (lrRNA) is 1339 bp long, with an A + T content of 73.6%, whereas the small rRNA subunit gene (srRNA) is 790 bp long, with an A + T content of 68.9%. The lrRNA and srRNA genes present in the M. flexura mitogenome are located between tRNALeu(CUN) and tRNAVal, and between tRNAVal and the control region, respectively. Results of the rRNAs secondary structures show that lrRNA and srRNA have five (I–II, IV–VI, with domain III absent) and three (I–III) structural domains, respectively (Figures S2 and S3). Both lrRNA and srRNA have characteristics that are similar to those found in most published plecopteran species [18,20,21,22,23,39].
3.4. The Control Region
The control region contains essential elements involved in the initiation of replication and transcription of the mitogenome [47]. The control region of M. flexura is 735 bp in length and is located between srRNA and tRNAIle (Figure 1 and Table 2). It contains the highest A + T content (85.2%) in the entire mitogenome.
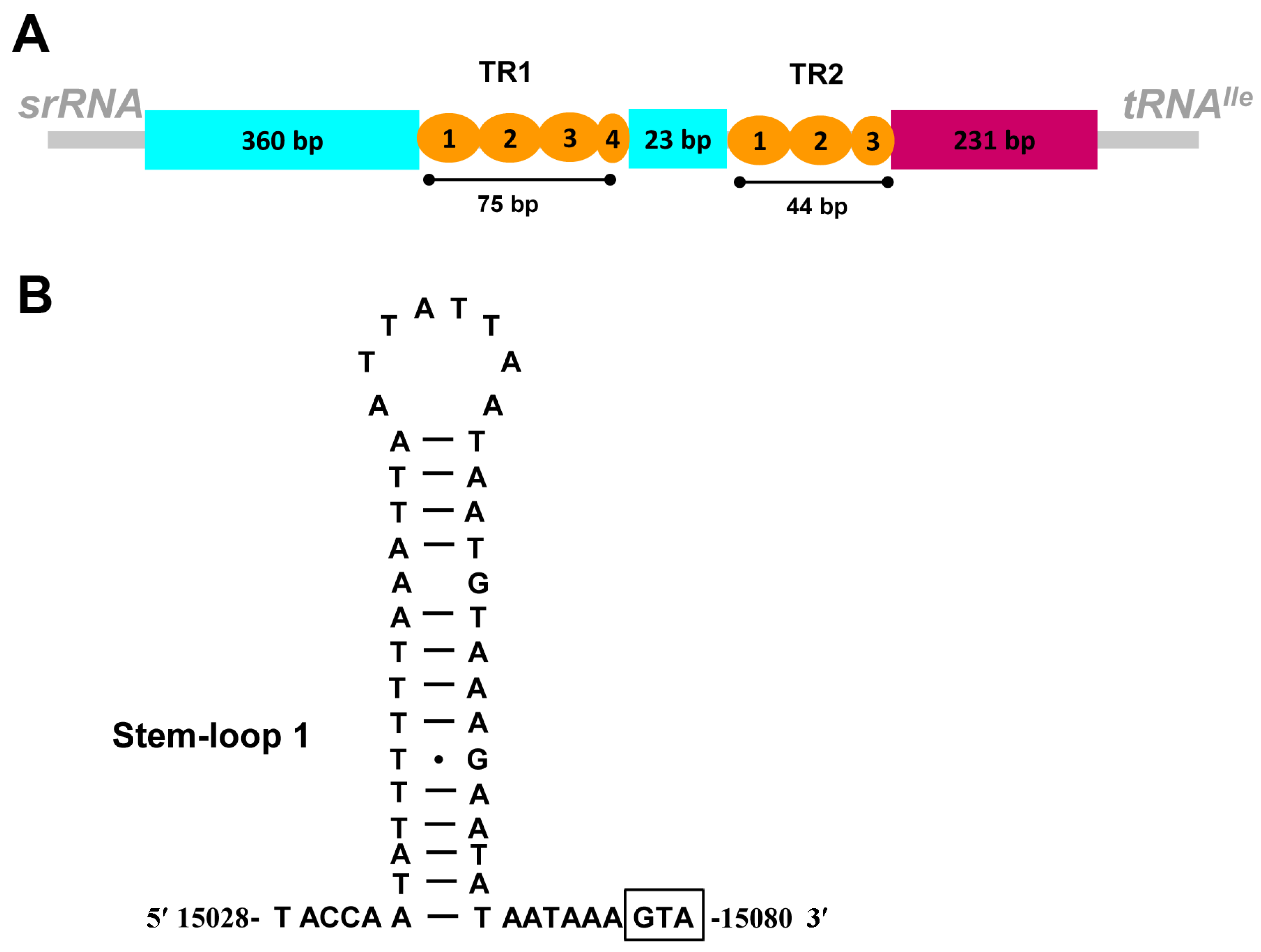
The control region of M. flexura can be divided into five parts: (1) a 360 bp leading sequence adjacent to srRNA composed of a stem-loop structure; (2) a 75 bp tandemly repeated sequence block consisting of three complete and one incomplete tandem repeat units; (3) a 23 bp region; (4) a 44 bp region including two complete and one incomplete tandem repeats units; (5) a 231 bp region at the end of the control region (Figure 3A).
One stem-loop (SL1, position: 15,028 bp–15,080 bp) structure is predicted in the control region (Figure 3B). The proposed SL structure with a 3′ flanking “G(A)nT” motif is not detected in SL1, but it is modified as “GTA”. The stem-loop structure in the control region is identified in many insects and it is thought to be the site of the initiation of secondary strand synthesis in Drosophila [48]. In addition, one microsatellite sequence (position: 15,376 bp–15,391 bp), (AT)8, is detected in the control region. Similar to other stoneflies mitogenomes [18,40,49], several poly-N stretch (≥7 bp) were also found near to the tRNAIle, such as poly-T (9 bp, position: 15,668 bp–15,676 bp) and poly-A (9 bp, position: 15,574 bp–15,582 bp). Poly-T stretch is considered to be essential for the initiation of replication in insects [50].
3.5. Phylogenetic Relationships
In the present study, concatenated nucleotide sequences of 13 PCGs from 17 nemourid mitogenomes were used to reconstruct phylogenetic relationships by the BI and ML methods. Two methods generated the same tree topologies (Figure 4).
The monophyly of each genus is generally well supported (bootstrap probabilities (BSPs) ≥ 74; Bayesian posterior probabilities (BPPs) ≥ 0.99). In morphology, Indonemoura, Mesonemoura, Protonemura and Sphaeronemoura are similar to each other in having a sclerotized projection (tigellus) on the median lobe of the paraproct [13]. Although Shimizu and Sivec regarded the genus Sphaeronemoura not to be closely related to Mesonemoura [14], result of this study supports Sphaeronemoura as a sister group of Mesonemoura. This result is similar to that of Cao et al. (2019) [18]. In addition, the position of Protonemura and Indonemoura is consistent with the traditionally proposed relationships [13] and previous studies [18,19,20]. Moreover, the traditional morphology-based classification among Amphinemurinae was well supported [13].
Malenka is the sister genus of Amphinemura and is restricted to western North America. They usually have a distinct median notch with other genera of Amphinemurinae [13]. However, phylogenetic analyses in this study do not support the sister group relationship of Amphinemura and Malenka. This result is similar to that of Thomas et al. (2000) [16] and Terry (2003) [17], but differs from the morphological studies of Baumann (1975) [13]. More comprehensive sampling especially for those stoneflies from the Malenka is expected to better resolve the mitochondrial phylogeny of Amphinemurinae. Finally, the best-supported phylogenetic relationship found in this study is as follows: Amphinemura + (Malenka + (Protonemura + (Indonemoura + (Sphaeronemoura + Mesonemoura)))).
4. Conclusions
Currently, the position of five genera in Amphinemurinae has been resolved based on morphology. However, the results of early molecular studies differ from morphological results. With the establishment of two new genera (Sphaeronemoura and Tominemoura), the phylogenetic relationships within Amphinemurinae should be re-examined. In this study, one complete mitogenome from genus Malenka, in the subfamily Amphinemurinae, was presented. Its mitogenome organizations and phylogenetic relationships with the other species from Amphinemurinae were analyzed. The M. flexura mitogenome resulted in a DNA molecule with genomic features typical for insect mitogenomes, such as conserved gene order, gene content, nucleotide composition, codon usage of PCGs and RNA secondary structures. In addition, some structural elements were also found in the control region, such as tandem repeats regions, poly-N stretch, stem-loop structures, etc. The phylogenetic analyses indicated that within Amphinemurinae, Amphinemura and Malenka were not supported as a sister-group relationship. The relationship between Amphinemura and Malenka would be improved if a more comprehensive taxon sampling was used. Finally, phylogenetic analyses inferred a relationship within Amphinemurinae: Amphinemura + (Malenka + (Protonemura + (Indonemoura + (Sphaeronemoura + Mesonemoura)))).
Supplementary Materials
The following supporting information can be downloaded at: https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/genes13050911/s1, Figure S1: Predicted secondary structure of 22 tRNAs in M. flexura mitogenome; Figure S2: Predicted secondary structure of the lrRNA gene in M. flexura mitogenome; Figure S3: Predicted secondary structure of the srRNA gene in M. flexura mitogenome.
Author Contributions
Conceptualization, supervision and funding acquisition, Y.W. and F.Y.; investigation, J.C., X.G., C.G. and X.W.; writing—review and editing, J.C., X.G., Y.W. and F.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Program for Science & Technology Innovation Talents in Universities of Henan (No. 21HASTIT042), the Key Scientific Research Project of Henan Province (No. 21A210009, 22A210004), and the innovation and entrepreneurship training program for college students in Henan Province, China (No. S202110467005).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are deposited in GenBank with accession number ON411527. The data are available from the corresponding authors upon reasonable request.
Acknowledgments
We thank Boris C. Kondratieff (Colorado State University, USA) for sharing the samples.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Boore, J.L. Animal mitochondrial genomes. Nucleic Acids Res. 1999, 27, 1767–1780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, C.; Buckley, T.R.; Frati, F.; Stewart, J.B.; Beckenbach, A.T. Incorporating molecular evolution into phylogenetic analysis, and a new compilation of conserved polymerase chain reaction primers for animal mitochondrial DNA. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 545–579. [Google Scholar] [CrossRef] [Green Version]
- Wolstenholme, D. Genetic novelties in mitochondrial genomes of multicellular animals. Curr. Opin. Genet. Dev. 1992, 2, 918–925. [Google Scholar] [CrossRef]
- Cameron, S.L. Insect mitochondrial genomics: Implications for evolution and phylogeny. Annu. Rev. Èntomol. 2014, 59, 95–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Leavengood, J.M., Jr.; Chapman, E.G.; Burkhardt, D.; Song, F.; Jiang, P.; Liu, J.; Zhou, X.G.; Cai, W.Z. Mitochondrial phylogenomics of Hemiptera reveals adaptive innovations driving the diversification of true bugs. Proc. Roy. Soc. B–Biol. Sci. 2017, 284, 1862. [Google Scholar] [CrossRef] [PubMed]
- Song, F.; Li, H.; Jiang, P.; Zhou, X.; Liu, J.; Sun, C.; Vogler, A.P.; Cai, W. Capturing the phylogeny of holometabola with mitochondrial genome data and bayesian site-Heterogeneous mixture models. Genome Biol. Evol. 2016, 8, 1411–1426. [Google Scholar] [CrossRef]
- Song, F.; Li, H.; Liu, G.-H.; Wang, W.; James, P.; Colwell, D.D.; Tran, A.; Gong, S.; Cai, W.; Shao, R. Mitochondrial genome fragmentation unites the parasitic lice of Eutherian Mammals. Syst. Biol. 2019, 68, 430–440. [Google Scholar] [CrossRef] [Green Version]
- Simon, C.; Frati, F.; Beckenbach, A.; Crespi, B.; Liu, H.; Flook, P. Evolution, weighting, and phylogenetic utility of mitochondrial gene-sequences and a compilation of conserved polymerase chain–reaction primers. Ann. Entomol. Soc. Am. 1994, 87, 651–701. [Google Scholar] [CrossRef]
- Plecoptera Species File Online. Available online: http://Plecoptera.SpeciesFile.org (accessed on 10 April 2022).
- Zwick, P. Phylogenetic system and zoogeography of the Plecoptera. Annu. Rev. Èntomol. 2000, 45, 709–746. [Google Scholar] [CrossRef]
- South, E.J.; Skinner, R.K.; DeWalt, R.E.; A Davis, M.; Johnson, K.P.; A Teslenko, V.; Lee, J.J.; Malison, R.L.; Hwang, J.M.; Bae, Y.J.; et al. A new family of stoneflies (Insecta: Plecoptera), Kathroperlidae, fam. n., with a phylogenomic analysis of the Paraperlinae (Plecoptera: Chloroperlidae). Insect Syst. Divers. 2021, 5, 1. [Google Scholar] [CrossRef]
- Karr, J.R. Defining and measuring river health. Freshw. Biol. 1999, 41, 221–234. [Google Scholar] [CrossRef] [Green Version]
- Baumann, R.W. Revision of the stonefly family Nemouridae (Plecoptera): A study of the world fauna at the generic level. Smithson. Contrib. Zoöl. 1975, 211, 1–74. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, T.; Sivec, I. Sphaeronemoura, a new genus of the Amphinemurinae (Nemouridae, Plecoptera) from asia. In Trends in Research in Ephemeroptera and Plecoptera; Dominguez, E., Ed.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2001; pp. 393–399. [Google Scholar]
- Sivec, I.; Stark, B.P. Tominemoura, a new stonefly genus from Sabah, East Malaysia (Plecoptera: Nemouridae). Illiesia 2009, 5, 199–203. [Google Scholar]
- Thomas, M.A.; Walsh, K.A.; Wolf, M.R.; McPheron, B.A.; Marden, J.H. Molecular phylogenetic analysis of evolutionary trends in stonefly wing structure and locomotor behavior. Proc. Natl. Acad. Sci. USA 2000, 97, 13178–13183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terry, M.D. Phylogeny of the polyneopterous insects with emphasis on Plecoptera: Molecular and morpological evidence. Ph.D. Thesis, Brigham Young University, Provo, UT, USA, 2003; pp. 1–118. [Google Scholar]
- Cao, J.-J.; Wang, Y.; Li, W.-H. Comparative mitogenomic analysis of species in the subfamily Amphinemurinae (Plecoptera: Nemouridae) reveal conserved mitochondrial genome organization. Int. J. Biol. Macromol. 2019, 138, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Wang, Y.; Guo, X.; Wang, G.; Li, W.; Murányi, D. Two complete mitochondrial genomes from Leuctridae (Plecoptera: Nemouroidea): Implications for the phylogenetic relationships among stoneflies. J. Insect Sci. 2021, 21, 16. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Cao, J.; Murányi, D.; Chen, X.; Yan, F. The complete mitochondrial genome of Amphinemura bulla Shimizu, 1997 (Plecoptera: Nemouridae) from Japan. Mitochondrial DNA Part B 2021, 6, 846–847. [Google Scholar] [CrossRef]
- Wang, Y.; Cao, J.; Li, W. The complete mitochondrial genome of the styloperlid stonefly species Styloperla spinicercia Wu (Insecta: Plecoptera) with family-level phylogenetic analyses of the Pteronarcyoidea. Zootaxa 2017, 4243, 125–138. [Google Scholar] [CrossRef]
- Wang, Y.; Cao, J.; Murányi, D.; Li, W. Comparison of two complete mitochondrial genomes from Perlodidae (Plecoptera: Perloidea) and the family-level phylogenetic implications of Perloidea. Gene 2018, 675, 254–264. [Google Scholar] [CrossRef]
- Wang, Y.; Cao, J.J.; Li, N.; Ma, G.-Y.; Li, W.-H. The first mitochondrial genome from Scopuridae (Insecta: Plecoptera) reveals structural features and phylogenetic implications. Int. J. Biol. Macromol. 2019, 122, 893–902. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Bernt, M.; Donath, A.; Jühling, F.; Externbrink, F.; Florentz, C.; Fritzsch, G.; Pütz, J.; Middendorf, M.; Stadler, P.F. MITOS: Improved de novo metazoan mitochondrial genome annotation. Mol. Phylogenetics Evol. 2013, 69, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perna, N.T.; Kocher, T.D. Patterns of nucleotide composition at fourfold degenerate sites of animal mitochondrial genomes. J. Mol. Evol. 1995, 41, 353–358. [Google Scholar] [CrossRef]
- Benson, G. Tandem repeats finder: A program to analyze DNA sequences. Nucleic Acids Res. 1999, 27, 573–580. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Abascal, F.; Zardoya, R.; Telford, M.J. TranslatorX: Multiple alignment of nucleotide sequences guided by amino acid translations. Nucleic Acids Res. 2010, 38, W7–W13. [Google Scholar] [CrossRef] [Green Version]
- Trifinopoulos, J.; Nguyen, L.T.; von Haeseler, A.; Minh, B.Q. W-IQTREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Clary, D.O.; Wolstenholme, D.R. The ribosomal RNA genes of Drosophila mitochondrial DNA. Nucleic Acids Res. 1985, 13, 4029–4045. [Google Scholar] [CrossRef] [Green Version]
- Stewart, J.B.; Beckenbach, A.T. Insect mitochondrial genomics: The complete mitogenome sequence of the meadow spittlebug Philaenus spumarius (Hemiptera: Auchenorrhyncha: Cercopoidae). Genome 2005, 48, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Carapelli, A.; Vannini, L.; Nardi, F.; Boore, J.L.; Beani, L.; Dallai, R.; Frati, F. The mitochondrial genome of the entomophagous endoparasite Xenos vesparum (Insecta: Strepsiptera). Gene 2006, 376, 248–259. [Google Scholar] [CrossRef] [Green Version]
- Hassanin, A.; Léger, N.; Deutsch, J. Evidence for multiple reversals of asymmetric mutational constraints during the evolution of the mitochondrial genome of Metazoa, and consequences for phylogenetic inferences. Syst. Biol. 2005, 54, 277–298. [Google Scholar] [CrossRef] [PubMed]
- Hassanin, A. Phylogeny of Arthropoda inferred from mitochondrial sequences: Strategies for limiting the misleading effects of multiple changes in pattern and rates of substitution. Mol. Phylogenetics Evol. 2006, 38, 100–116. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.J.; Shi, M.; Chen, X.X.; Sharkey, M.J.; van Achterberg, C.; Ye, G.Y.; He, J.H. New views on strand asymmetry in insect mitochondrial genomes. PLoS ONE 2010, 5, e12708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.T.; Du, Y.Z. First Mitochondrial Genome from Nemouridae (Plecoptera) Reveals novel features of the elongated control region and phylogenetic implications. Int. J. Mol. Sci. 2017, 18, 996. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.Y.; Ji, X.Y.; Yu, W.W.; Du, Y.Z. Complete mitochondrial genome of the stonefly Cryptoperla stilifera Sivec (Plecop-tera: Peltoperlidae) and the phylogeny of Polyneopteran insects. Gene 2014, 537, 177–183. [Google Scholar] [CrossRef]
- Bae, J.S.; Kim, I.; Sohn, H.D.; Jin, B.R. The mitochondrial genome of the firefly, Pyrocoelia rufa: Complete DNA sequence, genome organization, and phylogenetic analysis with other insects. Mol. Phylogenetics Evol. 2004, 32, 978–985. [Google Scholar] [CrossRef]
- Sheffield, N.C.; Song, H.; Cameron, S.L.; Whiting, M.F. A Comparative analysis of mitochondrial genomes in Coleoptera (Arthropoda: Insecta) and genome descriptions of six new beetles. Mol. Biol. Evol. 2008, 25, 2499–2509. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Ding, S.; Yang, D. The complete mitochondrial genome of a stonefly species, Kamimuria chungnanshana Wu, 1948 (Plecoptera: Perlidae). Mitochondrial DNA Part A 2016, 27, 3810–3811. [Google Scholar] [CrossRef] [PubMed]
- Ojala, D.; Montoya, J.; Attardi, G. tRNA punctuation model of RNA processing in human mitochondria. Nature 1981, 290, 470–474. [Google Scholar] [CrossRef] [PubMed]
- Boore, J.L. Complete mitochondrial genome sequence of Urechis caupo, a representative of the phylum Echiura. BMC Genom. 2004, 5, e67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharp, P.M.; Li, W.H. An evolutionary perspective on synonymous codon usage in unicellular organisms. J. Mol. Evol. 1986, 24, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Taanman, J.W. The mitochondrial genome: Structure, transcription, translation and replication. Biochim. Biophys. Acta BBA-Bioenergies 1999, 1410, 103–123. [Google Scholar] [CrossRef] [Green Version]
- Clary, D.O.; Wolstenholme, D.R. Drosophila mitochondrial DNA: Conserved sequences in the A+T–rich region and sup-porting evidence for a secondary structure model of the small ribosomal RNA. J. Mol. Evol. 1987, 25, 116–125. [Google Scholar] [CrossRef]
- Chen, Z.T.; Du, Y.Z. Comparison of the complete mitochondrial genome of the stonefly Sweltsa longistyla (Plecoptera: Chloroperlidae) with mitogenomes of three other stoneflies. Gene 2015, 558, 82–87. [Google Scholar] [CrossRef]
- Zhang, D.X.; Hewitt, G.M. Insect mitochondrial control region: A review of its structure, evolution and usefulness in evolutionary studies. Biochem. Syst. Ecol. 1997, 25, 99–120. [Google Scholar] [CrossRef]
Figure 1.
Map of the mitogenome of M. flexura. tRNA genes are labeled using abbreviations. The content of Guanine and Cytosine nucleotides (GC content) is plotted as the deviation from the average GC content of the entire sequence. GC skew is plotted as the deviation from the average GC skew of the entire sequence.
Figure 1.
Map of the mitogenome of M. flexura. tRNA genes are labeled using abbreviations. The content of Guanine and Cytosine nucleotides (GC content) is plotted as the deviation from the average GC content of the entire sequence. GC skew is plotted as the deviation from the average GC skew of the entire sequence.

Figure 2.
The relative synonymous codon usage (RSCU) in the mitogenome of M. flexura. PCGs represent protein-coding genes.
Figure 2.
The relative synonymous codon usage (RSCU) in the mitogenome of M. flexura. PCGs represent protein-coding genes.

Figure 3.
Control region of the M. flexura mitogenome. (A) Structure elements found in the control region of M. flexura. (B) Putative stem-loop structures found in the control region of M. flexura.
Figure 3.
Control region of the M. flexura mitogenome. (A) Structure elements found in the control region of M. flexura. (B) Putative stem-loop structures found in the control region of M. flexura.

Figure 4.
Mitochondrial phylogenetic relationships among 17 stoneflies. Bayesian inference and aximum likelihood analysis inferred from PCGs supported the same topological structure. Values at nodes are Bayesian posterior probabilities (BPPs) and ML bootstrap probabilities (BSPs). The tree was rooted with three outgroups.
Figure 4.
Mitochondrial phylogenetic relationships among 17 stoneflies. Bayesian inference and aximum likelihood analysis inferred from PCGs supported the same topological structure. Values at nodes are Bayesian posterior probabilities (BPPs) and ML bootstrap probabilities (BSPs). The tree was rooted with three outgroups.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
General information of nemourid species used in this study.
| Subfamily | Species | Number (bp) | Accession Number |
|---|---|---|---|
| Amphinemurinae | Amphinemura longispina | 15,709 | MH085446 |
| Amphinemura yao | 15,876 | MH085447 | |
| Indonemoura auriformis | 15,718 | MN419915 | |
| Indonemoura jacobsoni | 15,642 | MH085448 | |
| Indonemoura nohirae | 15,738 | MH085449 | |
| Malenka flexura | 15,744 | ON411527 | |
| Mesonemoura metafiligera | 15,739 | MH085450 | |
| Mesonemoura tritaenia | 15,778 | MH085451 | |
| Protonemura datongensis | 15,756 | MT276842 | |
| Protonemura kohnoae | 15,707 | MH085452 | |
| Protonemura orbiculata | 15,758 | MH085453 | |
| Sphaeronemoura acutispina | 15,016 | MH085455 * | |
| Sphaeronemoura elephas | 15,846 | MN944385 | |
| Sphaeronemoura grandicauda | 15,661 | MH085454 * | |
| Nemourinae (Outgroup) | Nemoura meniscata | 15,895 | MN944386 |
| Nemoura nankinensis | 16,602 | KY940360 | |
| Nemoura papilla | 15,774 | MK290826 |
* Incomplete mitogenome sequence.
Table 2.
Organization of the M. flexura mitochondrial genome.
| Gene | Direction | Coordinates (bp) | Size (bp) | Anticodon or Start/Stop Codons | IGN (bp) |
|---|---|---|---|---|---|
| tRNAIle | J | 1–66 | 66 | 30–32 GAT | 0 |
| tRNAGln | N | 64–132 | 69 | 100–102 TTG | −3 |
| tRNAMet | J | 137–204 | 68 | 167–169 CAT | 4 |
| ND2 | J | 205–1239 | 1035 | ATG/TAA | 0 |
| tRNATrp | J | 1247–1315 | 69 | 1277–1279 TCA | 7 |
| tRNACys | N | 1308–1370 | 63 | 1339–1341 GCA | −8 |
| tRNATyr | N | 1377–1442 | 66 | 1409–1411 GTA | 6 |
| COI | J | 1435–2979 | 1545 | ATT/TAA | −8 |
| tRNALeu(UUR) | J | 2975–3041 | 67 | 3017–3019 TAA | −5 |
| COII | J | 3045–3732 | 688 | ATG/T- | 3 |
| tRNALys | J | 3733–3803 | 71 | 3763–3765 CTT | 0 |
| tRNAAsp | J | 3803–3870 | 68 | 3832–3834 GTC | −1 |
| ATP8 | J | 3871–4029 | 159 | ATT/TAA | 0 |
| ATP6 | J | 4023–4700 | 678 | ATG/TAA | −7 |
| COIII | J | 4700–5488 | 789 | ATG/TAA | −1 |
| tRNAGly | J | 5488–5553 | 66 | 5517–5519 TCC | −1 |
| ND3 | J | 5554–5907 | 354 | ATT/TAG | 3 |
| tRNAAla | J | 5906–5969 | 64 | 5935–5937 TGC | −2 |
| tRNAArg | J | 5970–6032 | 63 | 5999–6001 TCG | 0 |
| tRNAAsn | J | 6144–6209 | 66 | 6174–6172 GTT | 111 |
| tRNASer(AGN) | J | 6209–6277 | 69 | 6235–6237GCT | −1 |
| tRNAGlu | J | 6277–6345 | 69 | 6307–6309 TTC | −1 |
| tRNAPhe | N | 6344–6408 | 65 | 6376–6378 GAA | −2 |
| ND5 | N | 6409–8143 | 1735 | GTG/T- | 0 |
| tRNAHis | N | 8144–8209 | 66 | 8177–8179 GTG | 0 |
| ND4 | N | 8213–9553 | 1341 | ATG/TAA | 3 |
| ND4L | N | 9547–9843 | 297 | ATG/TAA | −7 |
| tRNAThr | J | 9846–9911 | 66 | 9877–9879 TGT | 2 |
| tRNAPro | N | 9911–9975 | 65 | 9943–9945 TGG | −1 |
| ND6 | J | 9977–10,501 | 525 | ATT/TAA | 0 |
| CytB | J | 10,501–11,637 | 1137 | ATG/TAG | −1 |
| tRNASer(UCN) | J | 11,636–11,705 | 70 | 11,667–11,669 TGA | −2 |
| ND1 | N | 11,792–12,742 | 951 | TTG/TAG | 86 |
| tRNALeu(CUN) | N | 12,744–12,809 | 66 | 12,778–12,780TAG | 1 |
| lrRNA | N | 12,810–14,148 | 1339 | 0 | |
| tRNAVal | N | 14,149–14,219 | 71 | 14,184–14,1186 TAC | 0 |
| srRNA | N | 14,220–15,009 | 790 | 0 | |
| CR | 15,010–15,744 | 735 | 0 |
CR—control region; IGN—intergenic nucleotides; J—majority strand; N—minority strand.
Table 3.
The nucleotide composition of the M. flexura mitogenome.
| Feature | Proportion of Nucleotides (%) | AT Skew | GC Skew | ||||
|---|---|---|---|---|---|---|---|
| T | C | A | G | A + T | |||
| Whole mitogenome | 36.3 | 12.5 | 32.3 | 18.8 | 68.6 | −0.059 | 0.200 |
| Protein-coding genes | 39.3 | 17.1 | 27.3 | 16.3 | 66.6 | −0.180 | −0.022 |
| Protein-coding genes J-strand | 35.6 | 20.7 | 29.1 | 14.7 | 64.7 | −0.101 | −0.171 |
| Protein-coding genes N-strand | 45.1 | 11.4 | 24.4 | 19.1 | 69.6 | −0.297 | 0.2552 |
| tRNA genes | 35.5 | 12.2 | 35.4 | 16.9 | 70.9 | −0.001 | 0.160 |
| tRNA genes J-strand | 34.9 | 13.9 | 36.0 | 15.2 | 70.9 | 0.015 | 0.044 |
| tRNA genes N-strand | 36.5 | 9.2 | 34.5 | 19.9 | 71.0 | −0.029 | 0.368 |
| rRNA genes | 39.0 | 10.1 | 32.9 | 18.1 | 71.9 | −0.084 | 0.285 |
| lrRNA | 40.4 | 9.0 | 33.2 | 17.4 | 73.6 | −0.097 | 0.320 |
| srRNA | 36.5 | 12.0 | 32.3 | 19.2 | 68.9 | −0.061 | 0.231 |
| Control region | 43.6 | 8.1 | 41.6 | 6.7 | 85.2 | −0.023 | −0.100 |
Table 4.
Codon number in the M. flexura mitochondrial PCGs.
| Codon | Count | Codon | Count | Codon | Count | Codon | Count |
|---|---|---|---|---|---|---|---|
| UUU(F) | 270 | UCU(S) | 95 | UAU(Y) | 109 | UGU(C) | 34 |
| UUC(F) | 51 | UCC(S) | 31 | UAC(Y) | 48 | UGC(C) | 8 |
| UUA(L) | 364 | UCA(S) | 74 | UAA(*) | 0 | UGA(W) | 87 |
| UUG(L) | 46 | UCG(S) | 12 | UAG(*) | 0 | UGG(W) | 19 |
| CUU(L) | 102 | CCU(P) | 63 | CAU(H) | 58 | CGU(R) | 14 |
| CUC(L) | 31 | CCC(P) | 38 | CAC(H) | 25 | CGC(R) | 7 |
| CUA(L) | 71 | CCA(P) | 42 | CAA(Q) | 64 | CGA(R) | 32 |
| CUG(L) | 12 | CCG(P) | 9 | CAG(Q) | 117 | CGG(R) | 7 |
| AUU(I) | 271 | ACU(T) | 87 | AAU(N) | 127 | AGU(S) | 45 |
| AUC(I) | 41 | ACC(T) | 27 | AAC(N) | 23 | AGC(S) | 17 |
| AUA(I) | 153 | ACA(T) | 75 | AAA(K) | 52 | AGA(S) | 63 |
| AUG(M) | 38 | ACG(T) | 12 | AAG(K) | 20 | AGG(S) | 3 |
| GUU(V) | 105 | GCU(A) | 92 | GAU(D) | 52 | GGU(G) | 53 |
| GUC(V) | 28 | GCC(A) | 38 | GAC(D) | 18 | GGC(G) | 34 |
| GUA(V) | 77 | GCA(A) | 54 | GAA(E) | 55 | GGA(G) | 83 |
| GUG(V) | 23 | GCG(A) | 24 | GAG(E) | 24 | GGG(G) | 79 |
* represent the stop codons.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Cao, J.; Guo, X.; Guo, C.; Wang, X.; Wang, Y.; Yan, F. Complete Mitochondrial Genome of Malenka flexura (Plecoptera: Nemouridae) and Phylogenetic Analysis. Genes 2022, 13, 911. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13050911
AMA Style
Cao J, Guo X, Guo C, Wang X, Wang Y, Yan F. Complete Mitochondrial Genome of Malenka flexura (Plecoptera: Nemouridae) and Phylogenetic Analysis. Genes. 2022; 13(5):911. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13050911
Chicago/Turabian StyleCao, Jinjun, Xuan Guo, Caiyue Guo, Xuan Wang, Ying Wang, and Fengming Yan. 2022. "Complete Mitochondrial Genome of Malenka flexura (Plecoptera: Nemouridae) and Phylogenetic Analysis" Genes 13, no. 5: 911. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13050911
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.