
Transcriptome Analysis Reveals Altered Expression of Genes Involved in Hypoxia, Inflammation and Immune Regulation in Pdcd10-Depleted Mouse Endothelial Cells

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. RNA Interference
2.3. Western Blotting
2.4. RNA Extraction
2.5. Library Preparation
2.6. Quantitative PCR (qPCR)
2.7. Bioinformatics Workflow
2.8. Confocal Microscopy
2.9. Statistical Analysis
3. Results
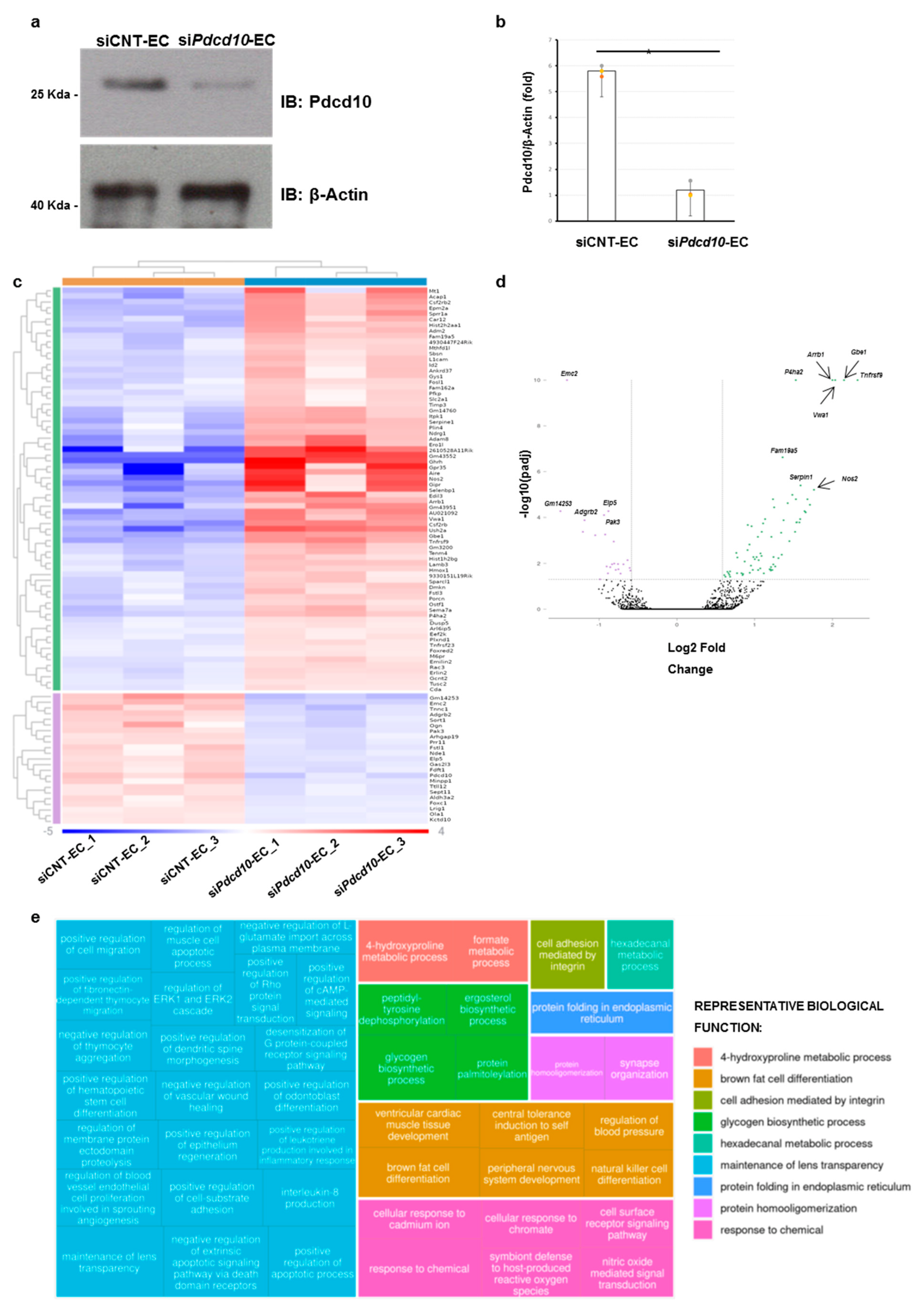
3.1. Pdcd10-Related Transcriptomic Profile
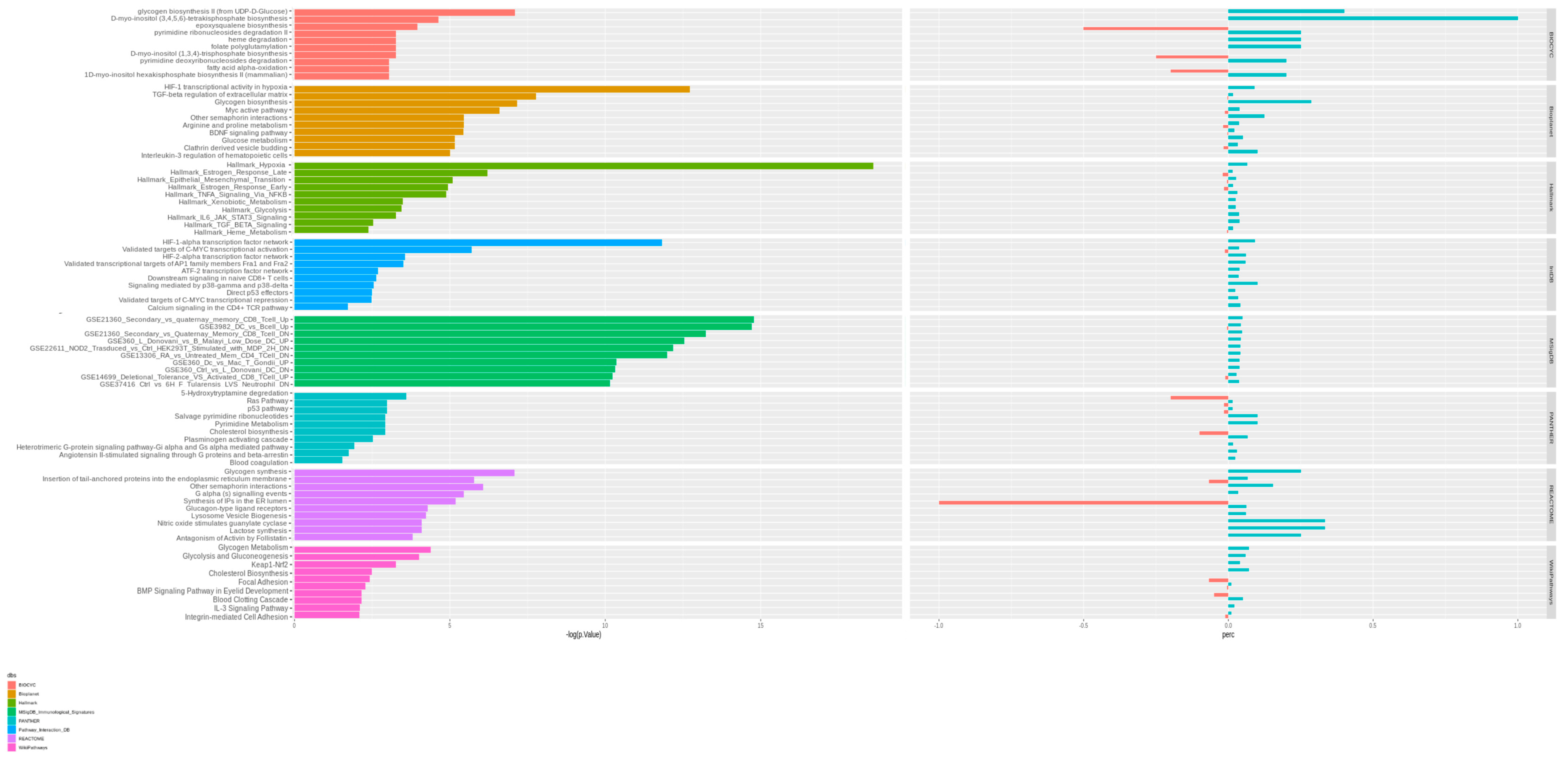
3.2. Pathway Analysis of Differentially Expressed Genes
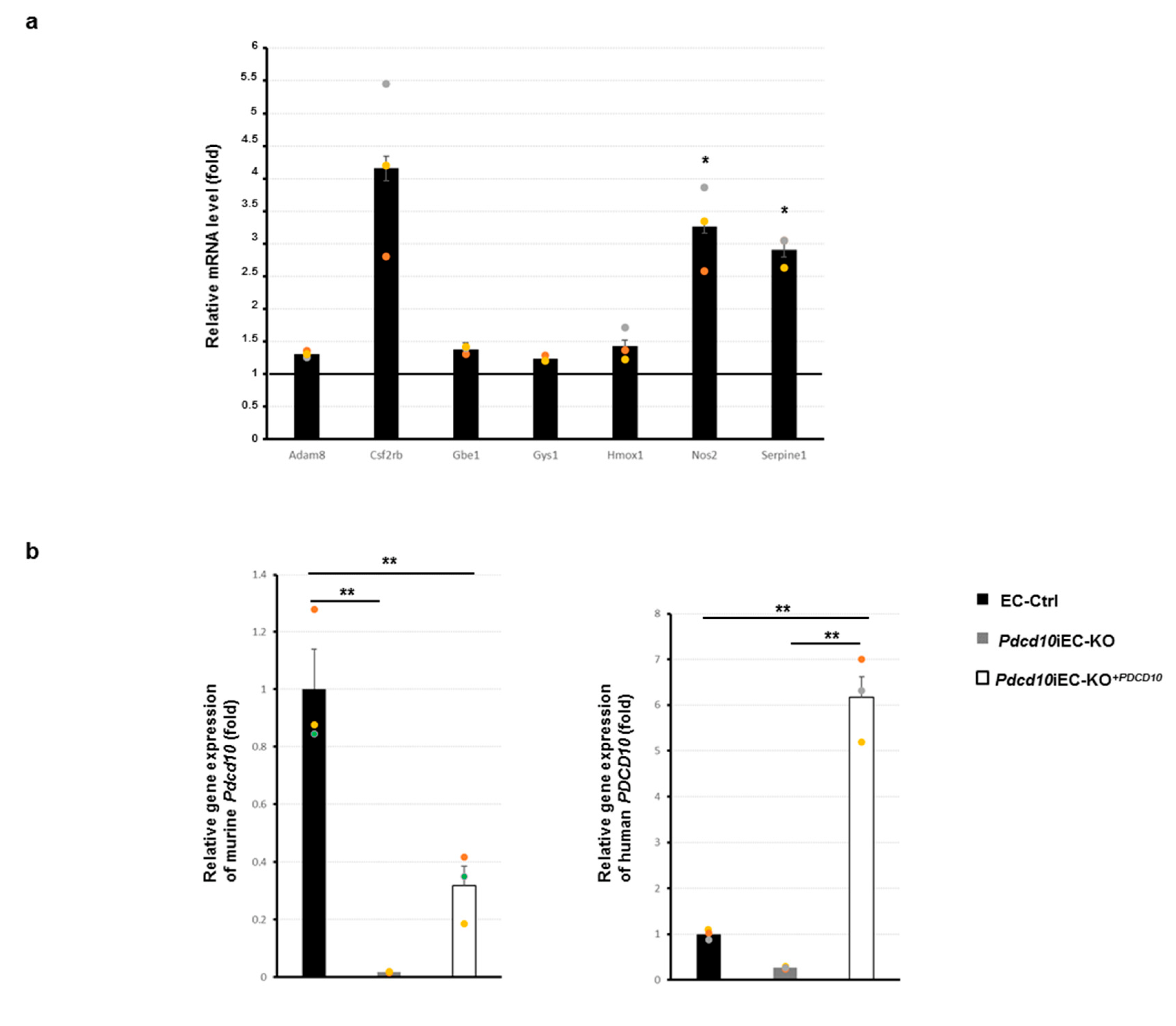
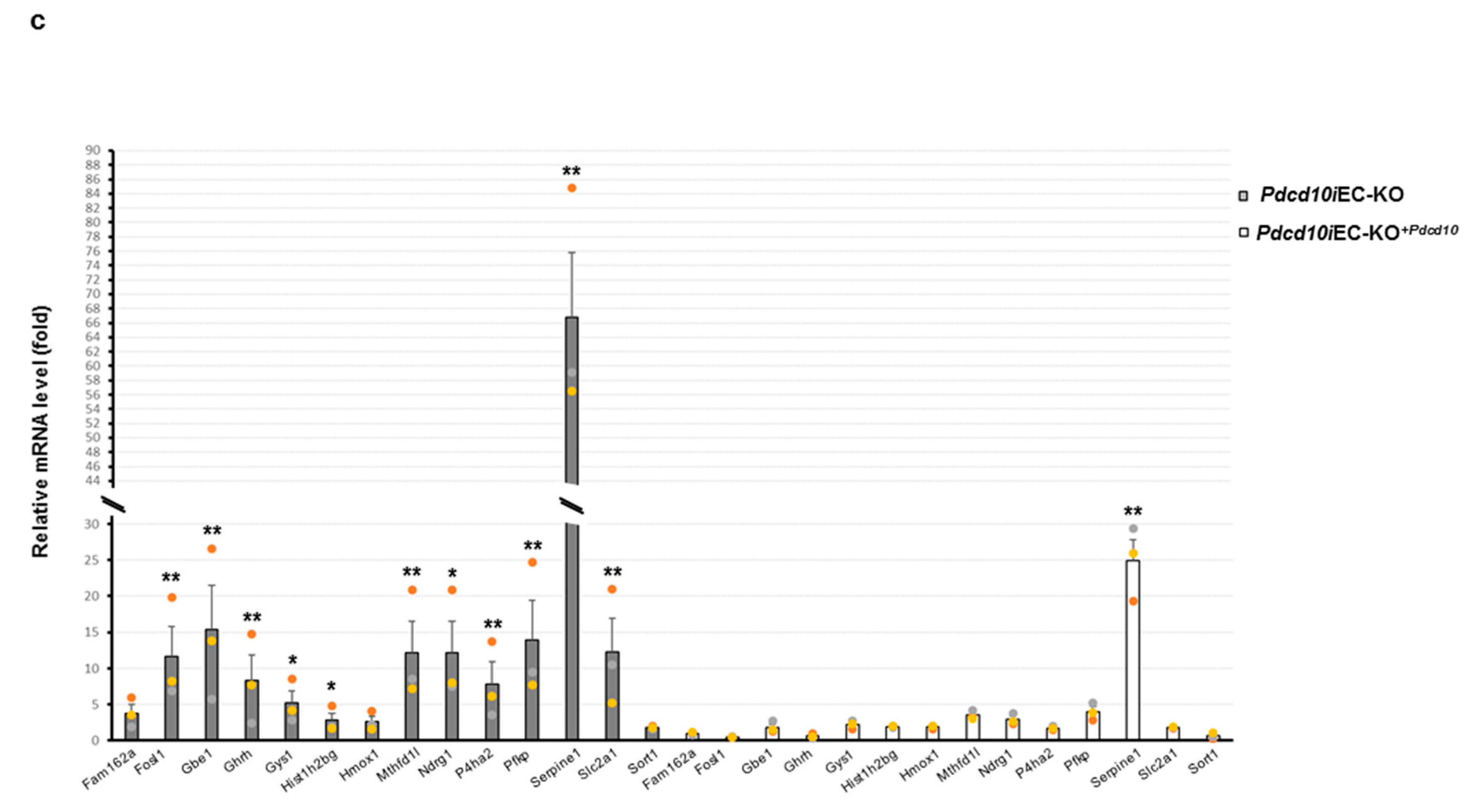
3.3. Validation Study of Differentially Expressed Genes in Mouse Endothelial Cells
4. Discussion
4.1. Hypoxia and HIF-1α Signaling
4.2. NOD2 Signaling
4.3. Immunological Signatures
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Idiculla, P.S.; Gurala, D.; Palanisamy, M.; Vijayakumar, R.; Dhandapani, S.; Nagarajan, E. Cerebral Venous Thrombosis: A Comprehensive Review. Eur. Neurol. 2020, 83, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Akers, A.; Al-Shahi Salman, R.; Awad, A.; Dahlem, K.; Flemming, K.; Hart, B.; Kim, H.; Jusue-Torres, I.; Kondziolka, D.; Lee, C.; et al. Synopsis of Guidelines for the Clinical Management of Cerebral Cavernous Malformations: Consensus Recommen-dations Based on Systematic Literature Review by the Angioma Alliance Scientific Advisory Board Clinical Experts Panel. Neurosurgery 2017, 80, 665–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labauge, P.; Denier, C.; Bergametti, F.; Tournier-Lasserve, E. Genetics of cavernous angiomas. Lancet Neurol. 2007, 6, 237–244. [Google Scholar] [CrossRef]
- Lanfranconi, S.; Scola, E.; Bertani, G.A.; Zarino, B.; Pallini, R.; d’Alessandris, G.; Mazzon, E.; Marino, S.; Carriero, M.R.; Scelzo, E.; et al. Propranolol for familial cerebral cavernous malformation (Treat_CCM): Study protocol for a randomized controlled pilot trial. Trials 2020, 21, 401. [Google Scholar] [CrossRef]
- Shenkar, R.; Shi, C.; Rebeiz, T.; Stockton, R.A.; McDonald, D.A.; Mikati, A.G.; Zhang, L.; Austin, C.; Akers, A.L.; Gallione, C.J.; et al. Exceptional aggressiveness of cerebral cavernous malformation disease associated with PDCD10 mutations. Genet. Med. 2015, 17, 188–196. [Google Scholar] [CrossRef] [Green Version]
- Cuttano, R.; Rudini, N.; Bravi, L.; Corada, M.; Giampietro, C.; Papa, E.; Morini, M.F.; Maddaluno, L.; Baeyens, N.; Adams, R.H.; et al. KLF 4 is a key determinant in the development and progression of cerebral cavernous malformations. EMBO Mol. Med. 2016, 8, 6–24. [Google Scholar] [CrossRef]
- Zhou, Z.; Tang, A.T.; Wong, W.Y.; Bamezai, S.; Goddard, L.M.; Shenkar, R.; Zhou, S.; Yang, J.; Wright, A.C.; Foley, M.; et al. Cerebral cavernous malformations arise from endothelial gain of MEKK3-KLF2/4 signalling. Nature 2016, 532, 122–126. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Zhang, H.; Yu, L.; Gunel, M.; Boggon, T.J.; Chen, H.; Min, W. Stabilization of VEGFR2 Signaling by Cerebral Cavernous Malformation 3 Is Critical for Vascular Development. Sci. Signal. 2010, 3, ra26. [Google Scholar] [CrossRef] [Green Version]
- Voss, K.; Stahl, S.; Hogan, B.M.; Reinders, J.; Schleider, E.; Schulte-Merker, S.; Felbor, U. Functional analyses of human and zebrafish 18-amino acid in-frame deletion pave the way for domain mapping of the cerebral cavernous malformation 3 protein. Hum. Mutat. 2009, 30, 1003–1011. [Google Scholar] [CrossRef]
- Yoruk, B.; Gillers, B.S.; Chi, N.C.; Scott, I.C. Ccm3 functions in a manner distinct from Ccm1 and Ccm2 in a zebrafish model of CCM vascular disease. Dev. Biol. 2012, 362, 121–131. [Google Scholar] [CrossRef] [Green Version]
- Zheng, X.; Xu, C.; Di Lorenzo, A.; Kleaveland, B.; Zou, Z.; Seiler, C.; Chen, M.; Cheng, L.; Xiao, J.; He, J.; et al. CCM3 signaling through sterile 20–like kinases plays an essential role during zebrafish cardiovascular development and cerebral cavernous malformations. J. Clin. Investig. 2010, 120, 2795–2804. [Google Scholar] [CrossRef] [PubMed]
- Detter, M.R.; Snellings, D.A.; Marchuk, D.A. Cerebral Cavernous Malformations Develop Through Clonal Expansion of Mu-tant Endothelial. Cells Circ. Res. 2018, 123, 1143–1151. [Google Scholar] [CrossRef] [PubMed]
- Malinverno, M.; Maderna, C.; Abu Taha, A.; Corada, M.; Orsenigo, F.; Valentino, M.; Pisati, F.; Fusco, C.; Graziano, P.; Giannotta, M.; et al. Endothelial cell clonal expansion in the development of cerebral cavernous malformations. Nat. Commun. 2019, 10, 2761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Y.; Chiang, W.C.; Sumpter, R., Jr.; Mishra, P.; Levine, B. Prohibitin 2 Is an Inner Mitochondrial Membrane Mitophagy Receptor. Cell 2017, 168, 224–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, M.; Franco, A.; Fleischer, J.A.; Zhang, L.; Dorn, G.W. Abrogating Mitochondrial Dynamics in Mouse Hearts Accelerates Mitochondrial Senescence. Cell Metab. 2017, 26, 872–883.e5. [Google Scholar] [CrossRef] [Green Version]
- Fusco, C.; Nardella, G.; Augello, B.; Boccafoschi, F.; Palumbo, O.; Fusaro, L.; Notarangelo, A.; Barbano, R.; Parrella, P.; Annicchiarico, G.; et al. Pro-Fibrotic Phenotype in a Patient with Segmental Stiff Skin Syndrome via TGF-beta Signaling Overactivation. Int. J. Mol. Sci. 2020, 21, 5141. [Google Scholar] [CrossRef]
- García-Fernández, E.; Koch, G.; Wagner, R.M.; Fekete, A.; Stengel, S.T.; Schneider, J.; Mielich-Süß, B.; Geibel, S.; Markert, S.; Stigloher, C.; et al. Membrane Microdomain Disassembly Inhibits MRSA Antibiotic Resistance. Cell 2017, 171, 1354–1367. [Google Scholar] [CrossRef]
- Aprile, M.; Cataldi, S.; Ambrosio, M.R.; D’Esposito, V.; Lim, K.; Dietrich, A.; Bluher, M.; Savage, D.B.; Formisano, P.; Ciccodicola, A.; et al. PPARgammaDelta5, a Naturally Occurring Dominant-Negative Splice Isoform, Impairs PPARgamma Function and Adipocyte Differentiation. Cell Rep. 2018, 25, 1577–1592. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Wang, S.; Li, W. RSeQC: Quality control of RNA-seq experiments. Bioinformatics 2012, 28, 2184–2185. [Google Scholar] [CrossRef] [Green Version]
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Near-optimal probabilistic RNA-seq quantification. Nat. Biotechnol. 2016, 34, 525–527. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaimal, V.; Bardes, E.E.; Tabar, S.C.; Jegga, A.G.; Aronow, B.J. ToppCluster: A multiple gene list feature analyzer for com-parative enrichment clustering and network-based dissection of biological systems. Nucleic Acids. Res. 2010, 38, W96–W102. [Google Scholar] [CrossRef] [Green Version]
- Rozen, S.; Skaletsky, H. Primer3 on the WWW for general users and for biologist programmers. Methods Mol. Biol. 2000, 132, 365–386. [Google Scholar] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Dobin, A.; Gingeras, T.R. Mapping RNA-seq Reads with STAR. Curr. Protoc. Bioinform. 2015, 51, 11–14. [Google Scholar] [CrossRef] [Green Version]
- Alexa, A.; Rahnenfuhrer, J.; Lengauer, T. Improved scoring of functional groups from gene expression data by decorrelating GO graph structure. Bioinformatics 2006, 22, 1600–1607. [Google Scholar] [CrossRef] [Green Version]
- Supek, F.; Bošnjak, M.; Škunca, N.; Smuc, T. REVIGO Summarizes and Visualizes Long Lists of Gene Ontology Terms. PLoS ONE 2011, 6, e21800. [Google Scholar] [CrossRef] [Green Version]
- Maddaluno, L.; Rudini, N.; Cuttano, R.; Bravi, L.; Giampietro, C.; Corada, M.; Ferrarini, L.; Orsenigo, F.; Papa, E.; Boulday, G.; et al. EndMT contributes to the onset and progression of cerebral cavernous malformations. Nature 2013, 498, 492–496. [Google Scholar] [CrossRef]
- Mabray, M.C.; Caprihan, A.; Nelson, J.; McCulloch, C.E.; Zafar, A.; Kim, H.; Hart, B.L.; Morrison, L. Effect of Simvastatin on Permeability in Cerebral Cavernous Malformation Type 1 Patients: Results from a Pilot Small Randomized Controlled Clinical Trial. Transl. Stroke Res. 2020, 11, 319–321. [Google Scholar] [CrossRef]
- Kar, S.; Bali, K.K.; Baisantry, A.; Geffers, R.; Samii, A.; Bertalanffy, H. Genome-Wide Sequencing Reveals MicroRNAs Downregulated in Cerebral Cavernous Malformations. J. Mol. Neurosci. 2017, 61, 178–188. [Google Scholar] [CrossRef]
- Subhash, S.; Kalmbach, N.; Wegner, F.; Petri, S.; Glomb, T.; Dittrich-Breiholz, O.; Huang, C.; Bali, K.K.; Kunz, W.S.; Samii, A.; et al. Transcriptome-wide Profiling of Cerebral Cavernous Malformations Patients Reveal Important Long noncoding RNA molecular signatures. Sci. Rep. 2019, 9, 18203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koskimäki, J.; Polster, S.P.; Li, Y.; Romanos, S.; Srinath, A.; Zhang, D.; Carrión-Penagos, J.; Lightle, R.; Moore, T.; Lyne, S.B.; et al. Common transcriptome, plasma molecules, and imaging signatures in the aging brain and a Mendelian neurovascular disease, cerebral cavernous malformation. GeroScience 2020, 42, 1351–1363. [Google Scholar] [CrossRef] [PubMed]
- Scimone, C.; Donato, L.; Alibrandi, S.; Esposito, T.; Alafaci, C.; D’Angelo, R.; Sidoti, A. Transcriptome analysis provides new molecular signatures in sporadic Cerebral Cavernous Malformation endothelial cells. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165956. [Google Scholar] [CrossRef] [PubMed]
- Abou-Fadel, J.; Zhang, J. Systems Wide Analysis of CCM Signaling Complex Alterations in CCM-Deficient Models Using Omics Approaches. Methods Mol. Biol. 2020, 2152, 325–344. [Google Scholar] [CrossRef]
- Koskimaki, J.; Zhang, D.; Li, Y.; Saadat, L.; Moore, T.; Lightle, R.; Polster, S.P.; Carrion-Penagos, J.; Lyne, S.B.; Zeineddine, H.A.; et al. Transcriptome clarifies mechanisms of lesion genesis versus progression in models of Ccm3 cerebral cavernous malfor-mations. Acta Neuropathol. Commun. 2019, 7, 132. [Google Scholar] [CrossRef]
- Koskimäki, J.; Girard, R.; Li, Y.; Saadat, L.; Zeineddine, H.A.; Lightle, R.; Moore, T.; Lyne, S.; Avner, K.; Shenkar, R.; et al. Comprehensive transcriptome analysis of cerebral cavernous malformation across multiple species and genotypes. JCI Insight 2019, 4, e126167. [Google Scholar] [CrossRef] [Green Version]
- Orsenigo, F.; Conze, L.L.; Jauhiainen, S.; Corada, M.; Lazzaroni, F.; Malinverno, M.; Sundell, V.; Cunha, S.I.; Brännström, J.; Globisch, M.A.; et al. Mapping endothelial-cell diversity in cerebral cavernous malformations at single-cell resolution. eLife 2020, 9, e61413. [Google Scholar] [CrossRef]
- Ma, X.; Zhao, H.; Shan, J.; Long, F.; Chen, Y.; Chen, Y.; Zhang, Y.; Han, X.; Ma, D. PDCD10 Interacts with Ste20-related Kinase MST4 to Promote Cell Growth and Transformation via Modulation of the ERK Pathway. Mol. Biol. Cell 2007, 18, 1965–1978. [Google Scholar] [CrossRef] [Green Version]
- Bravi, L.; Malinverno, M.; Pisati, F.; Rudini, N.; Cuttano, R.; Pallini, R.; Martini, M.; Larocca, L.M.; Locatelli, M.; Levi, V.; et al. Endothelial Cells Lining Sporadic Cerebral Cavernous Malformation Cavernomas Undergo Endothelial-to-Mesenchymal Transition. Stroke 2016, 47, 886–890. [Google Scholar] [CrossRef]
- Lim, C.S.; Kiriakidis, S.; Paleolog, E.M.; Davies, A.H. Increased activation of the hypoxia-inducible factor pathway in varicose veins. J. Vasc. Surg. 2012, 55, 1427–1439. [Google Scholar] [CrossRef] [Green Version]
- Mikati, A.G.; Khanna, O.; Zhang, L.; Girard, R.; Shenkar, R.; Guo, X.; Shah, A.; Larsson, H.B.W.; Tan, H.; Li, L.; et al. Vascular Permeability in Cerebral Cavernous Malformations. J. Cereb. Blood Flow Metab. 2015, 35, 1632–1639. [Google Scholar] [CrossRef] [PubMed]
- Theivanthiran, B.; Batra, S.; Balamayooran, G.; Cai, S.; Kobayashi, K.; Flavell, R.A.; Jeyaseelan, S. NOD2 Signaling Contributes to Host Defense in the Lungs against Escherichia coli Infection. Infect. Immun. 2012, 80, 2558–2569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tukhvatulin, A.I.; Dzharullaeva, A.S.; Tukhvatulina, N.M.; Shcheblyakov, D.V.; Shmarov, M.M.; Dolzhikova, I.V.; Stan-hope-Baker, P.; Naroditsky, B.S.; Gudkov, A.V.; Logunov, D.Y.; et al. Powerful Complex Immunoadjuvant Based on Synergistic Effect of Combined TLR4 and NOD2 Activation Significantly Enhances Magnitude of Humoral and Cellular Adaptive Immune Responses. PLoS ONE 2016, 11, e0155650. [Google Scholar] [CrossRef] [PubMed]
- Ramanan, D.; Tang, M.S.; Bowcutt, R.; Loke, P.; Cadwell, K. Bacterial Sensor Nod2 Prevents Inflammation of the Small Intestine by Restricting the Expansion of the Commensal Bacteroides vulgatus. Immunity 2014, 41, 311–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cryan, J.F.; O’Riordan, K.J.; Sandhu, K.; Peterson, V.; Dinan, T.G. The gut microbiome in neurological disorders. Lancet Neurol. 2020, 19, 179–194. [Google Scholar] [CrossRef]
- Tang, A.T.; Choi, J.P.; Kotzin, J.J.; Yang, Y.; Hong, C.; Hobson, N.; Girard, R.; Zeineddine, H.A.; Lightle, R.; Moore, T.; et al. Endothelial TLR4 and the microbiome drive cerebral cavernous malformations. Nature 2017, 545, 305–310. [Google Scholar] [CrossRef]
- Tang, A.T.; Sullivan, K.R.; Hong, C.C.; Goddard, L.M.; Mahadevan, A.; Ren, A.; Pardo, H.; Peiper, A.; Griffin, E.; Tanes, C.; et al. Distinct cellular roles for PDCD10 define a gut-brain axis in cerebral cavernous malformation. Sci. Transl. Med. 2019, 11, eaaw3521. [Google Scholar] [CrossRef]
- Kabesch, M.; Depner, M.; Dahmen, I.; Weiland, S.K.; Vogelberg, C.; Niggemann, B.; Lau, S.; Illig, T.; Klopp, N.; Wahn, U.; et al. Original article: Polymorphisms in eosinophil pathway genes, asthma and atopy. Allergy 2007, 62, 423–428. [Google Scholar] [CrossRef]
- Oh, Y.M.; Park, H.B.; Shin, J.H.; Lee, J.E.; Park, H.Y.; Kho, D.H.; Lee, J.S.; Choi, H.; Okuda, T.; Kokame, K.; et al. Ndrg1 is a T-cell clonal anergy factor negatively regulated by CD28 costimulation and interleukin-2. Nat. Commun. 2015, 6, 8698. [Google Scholar] [CrossRef] [Green Version]
- Siracusa, M.C.; Saenz, S.A.; Hill, D.A.; Kim, B.S.; Headley, M.B.; Doering, T.A.; Wherry, E.J.; Jessup, H.K.; Siegel, L.A.; Kam-bayashi, T.; et al. TSLP promotes interleukin-3-independent basophil haematopoiesis and type 2 inflammation. Nature 2011, 477, 229–233. [Google Scholar] [CrossRef]
- Morioka, S.; Perry, J.S.A.; Raymond, M.H.; Medina, C.B.; Zhu, Y.; Zhao, L.; Serbulea, V.; Onengut-Gumuscu, S.; Leitinger, N.; Kucenas, S.; et al. Efferocytosis induces a novel SLC program to promote glucose uptake and lactate release. Nature 2018, 563, 714–718. [Google Scholar] [CrossRef] [PubMed]
- Kaech, S.M.; Cui, W. Transcriptional control of effector and memory CD8+ T cell differentiation. Nat. Rev. Immunol. 2012, 12, 749–761. [Google Scholar] [CrossRef] [PubMed]
- Korn, T.; Kallies, A. T cell responses in the central nervous system. Nat. Rev. Immunol. 2017, 17, 179–194. [Google Scholar] [CrossRef] [PubMed]
- Manglani, M.; McGavern, D.B. New advances in CNS immunity against viral infection. Curr. Opin. Virol. 2018, 28, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Shenkar, R.; Kinloch, A.; Henderson, S.G.; Shaaya, M.; Chong, A.S.; Clark, M.R.; Awad, I.A. Immune complex for-mation and in situ B-cell clonal expansion in human cerebral cavernous malformations. J. Neuroimmunol. 2014, 15, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Wan, M.; Ma, L.; Liu, X.; He, J. Protective effects of ADAM8 against cisplatin-mediated apoptosis in non-small-cell lung cancer. Cell Biol. Int. 2013, 37, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Wang, J.; Xiao, Y.; Wang, C.; Qiu, Q.; Lao, M.; Yu, Y.; Li, Z.; Zhang, H.; Ye, Y.; et al. Glycogen Metabolism and Rheumatoid Arthritis: The Role of Glycogen Synthase 1 in Regulation of Synovial Inflammation via Blocking AMP-Activated Protein Kinase Activation. Front. Immunol. 2018, 9, 1714. [Google Scholar] [CrossRef]
- Andreuzzi, E.; Capuano, A.; Pellicani, R.; Poletto, E.; Doliana, R.; Maiero, S.; Fornasarig, M.; Magris, R.; Colombatti, A.; Cannizzaro, R.; et al. Loss of Multimerin-2 and EMILIN-2 Expression in Gastric Cancer Associate with Altered Angiogenesis. Int. J. Mol. Sci. 2018, 19, 3983. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol | GeneID | Description | Ensembl_gene_id | Log Fold Change | p-Value | p-Adj |
|---|---|---|---|---|---|---|
| Tnfrsf9 | 21942 | tumor necrosis factor receptor superfamily_ member 9 | ENSMUSG00000028965 | 2.323 | 2.1 × 10−28 | 6.9 × 10−24 |
| Gbe1 | 74185 | glucan (1_4-alpha-)_ branching enzyme 1 | ENSMUSG00000022707 | 2.149 | 6.6 × 10−27 | 1.0 × 10−22 |
| Arrb1 | 109689 | arrestin_ beta 1 | ENSMUSG00000018909 | 2.000 | 4.6 × 10−21 | 5.1 × 10−17 |
| P4ha2 | 18452 | procollagen-proline_ 2-oxoglutarate 4-dioxygenase (proline 4-hydroxylase)_ alpha II polypeptide | ENSMUSG00000018906 | 1.529 | 1.8 × 10−17 | 1.5 × 10−13 |
| Vwa1 | 246228 | von Willebrand factor A domain containing 1 | ENSMUSG00000042116 | 2.035 | 3.4 × 10−16 | 2.2 × 10−12 |
| Emc2 | 66736 | ER membrane protein complex subunit 2 | ENSMUSG00000022337 | −1.413 | 4.9 × 10−16 | 2.6 × 10−12 |
| Fam19a5 | 106014 | family with sequence similarity 19_ member A5 | ENSMUSG00000054863 | 1.360 | 5.0 × 10−11 | 2.3 × 10−7 |
| Epm2a | 13853 | epilepsy_ progressive myoclonic epilepsy_ type 2 gene alpha | ENSMUSG00000055493 | 1.590 | 9.7 × 10−10 | 4.0 × 10−6 |
| Csf2rb | 12983 | colony stimulating factor 2 receptor_ beta_ low-affinity (granulocyte-macrophage) | ENSMUSG00000071713 | 1.763 | 1.7 × 10−9 | 6.2 × 10−6 |
| Hist2h2aa1 | 15267 | histone cluster 2_ H2aa1 | ENSMUSG00000064220 | 1.484 | 3.1 × 10−9 | 1.0 × 10−5 |
| Lamb3 | 16780 | laminin_ beta 3 | ENSMUSG00000026639 | 1.236 | 5.8 × 10−9 | 1.6 × 10−5 |
| Serpine1 | 18787 | serine (or cysteine) peptidase inhibitor_ clade E_ member 1 | ENSMUSG00000037411 | 1.530 | 5.4 × 10−9 | 1.6 × 10−5 |
| Adam8 | 11501 | a disintegrin and metallopeptidase domain 8 | ENSMUSG00000025473 | 1.679 | 6.3 × 10−9 | 1.6 × 10−5 |
| Ndrg1 | 17988 | N-myc downstream regulated gene 1 | ENSMUSG00000005125 | 1.425 | 1.0 × 10−8 | 2.3 × 10−5 |
| Gipr | 381853 | gastric inhibitory polypeptide receptor | ENSMUSG00000030406 | 1.711 | 1.2 × 10−8 | 2.8 × 10−5 |
| Tenm4 | 23966 | teneurin transmembrane protein 4 | ENSMUSG00000048078 | 1.251 | 1.7 × 10−8 | 3.6 × 10−5 |
| Elp5 | 54351 | elongator acetyltransferase complex subunit 5 | ENSMUSG00000018565 | −0.881 | 3.0 × 10−8 | 5.3 × 10−5 |
| Ghrh | 14601 | growth hormone releasing hormone | ENSMUSG00000027643 | 1.642 | 2.8 × 10−8 | 5.3 × 10−5 |
| Gm14253 | 628707 | programmed cell death 10 pseudogene | ENSMUSG00000082321 | −1.498 | 3.0 × 10−8 | 5.3 × 10−5 |
| Nos2 | 18126 | nitric oxide synthase 2_ inducible | ENSMUSG00000020826 | 1.655 | 3.8 × 10−8 | 5.9 × 10−5 |
| Selenbp1 | 20341 | selenium binding protein 1 | ENSMUSG00000068874 | 1.655 | 3.7 × 10−8 | 5.9 × 10−5 |
| Pak3 | 18481 | p21 protein (Cdc42/Rac)-activated kinase 3 | ENSMUSG00000031284 | −0.937 | 5.3 × 10−8 | 7.9 × 10−5 |
| Fstl3 | 83554 | follistatin-like 3 | ENSMUSG00000020325 | 1.134 | 8.3 × 10−8 | 1.1 × 10−4 |
| Gm43552 | NULL | predicted gene 43552 | ENSMUSG00000105835 | 1.585 | 8.4 × 10−8 | 1.1 × 10−4 |
| Pdcd10 | 56426 | programmed cell death 10 | ENSMUSG00000027835 | −1.188 | 1.0 × 10−7 | 1.3 × 10−4 |
| Itpk1 | 217837 | inositol 1_3_4-triphosphate 5/6 kinase | ENSMUSG00000057963 | 1.372 | 9.9 × 10−8 | 1.3 × 10−4 |
| Ero1l | 50527 | ERO1-like (S. cerevisiae) | ENSMUSG00000021831 | 1.543 | 1.3 × 10−7 | 1.5 × 10−4 |
| Edil3 | 13612 | EGF-like repeats and discoidin I-like domains 3 | ENSMUSG00000034488 | 1.537 | 1.4 × 10−7 | 1.6 × 10−4 |
| AU021092 | 239691 | expressed sequence AU021092 | ENSMUSG00000051669 | 1.576 | 1.4 × 10−7 | 1.6 × 10−4 |
| Hmox1 | 15368 | heme oxygenase 1 | ENSMUSG00000005413 | 1.109 | 1.6 × 10−7 | 1.7 × 10−4 |
| Erlin2 | 244373 | ER lipid raft associated 2 | ENSMUSG00000031483 | 0.912 | 1.9 × 10−7 | 2.0 × 10−4 |
| Emilin2 | 246707 | elastin microfibril interfacer 2 | ENSMUSG00000024053 | 0.960 | 2.3 × 10−7 | 2.3 × 10−4 |
| Aire | 11634 | autoimmune regulator (autoimmune polyendocrinopathy candidiasis ectodermal dystrophy) | ENSMUSG00000000731 | 1.520 | 4.4 × 10−7 | 4.1 × 10−4 |
| Id2 | 15902 | inhibitor of DNA binding 2 | ENSMUSG00000020644 | 1.091 | 4.2 × 10−7 | 4.1 × 10−4 |
| Tnnc1 | 21924 | troponin C_ cardiac/slow skeletal | ENSMUSG00000091898 | −1.209 | 4.3 × 10−7 | 4.1 × 10−4 |
| Adm2 | 223780 | adrenomedullin 2 | ENSMUSG00000054136 | 1.341 | 4.9 × 10−7 | 4.5 × 10−4 |
| Gas2l3 | 237436 | growth arrest-specific 2 like 3 | ENSMUSG00000074802 | −0.923 | 6.1 × 10−7 | 5.4 × 10−4 |
| Adgrb2 | 230775 | adhesion G protein-coupled receptor B2 | ENSMUSG00000028782 | −1.050 | 6.8 × 10−7 | 5.8 × 10−4 |
| Sema7a | 20361 | sema domain_ immunoglobulin domain (Ig)_ and GPI membrane anchor_ (semaphorin) 7A | ENSMUSG00000038264 | 1.215 | 8.4 × 10−7 | 6.9 × 10−4 |
| Sbsn | 282619 | suprabasin | ENSMUSG00000046056 | 1.041 | 8.4 × 10−7 | 6.9 × 10−4 |
| Ush2a | 22283 | Usher syndrome 2A (autosomal recessive_ mild) | ENSMUSG00000026609 | 1.447 | 1.3 × 10−6 | 1.0 × 10−3 |
| Prr11 | 270906 | proline rich 11 | ENSMUSG00000020493 | −0.812 | 1.4 × 10−6 | 1.1 × 10−3 |
| Ostf1 | 20409 | osteoclast stimulating factor 1 | ENSMUSG00000024725 | 0.933 | 1.9 × 10−6 | 1.5 × 10−3 |
| Timp3 | 21859 | tissue inhibitor of metalloproteinase 3 | ENSMUSG00000020044 | 0.909 | 3.5 × 10−6 | 2.6 × 10−3 |
| Tnfrsf23 | 79201 | tumor necrosis factor receptor superfamily_ member 23 | ENSMUSG00000037613 | 0.764 | 4.7 × 10−6 | 3.4 × 10−3 |
| Gys1 | 14936 | glycogen synthase 1_ muscle | ENSMUSG00000003865 | 1.075 | 5.1 × 10−6 | 3.6 × 10−3 |
| Sprr1a | 20753 | small proline-rich protein 1A | ENSMUSG00000050359 | 1.348 | 5.9 × 10−6 | 4.1 × 10−3 |
| Dmkn | 73712 | dermokine | ENSMUSG00000060962 | 1.088 | 6.8 × 10−6 | 4.7 × 10−3 |
| Plin4 | 57435 | perilipin 4 | ENSMUSG00000002831 | 1.242 | 7.8 × 10−6 | 5.2 × 10−3 |
| Sparcl1 | 13602 | SPARC-like 1 | ENSMUSG00000029309 | 1.025 | 8.1 × 10−6 | 5.2 × 10−3 |
| Mthfd1l | 270685 | methylenetetrahydrofolate dehydrogenase (NADP+ dependent) 1-like | ENSMUSG00000040675 | 0.984 | 8.2 × 10−6 | 5.2 × 10−3 |
| Gcnt2 | 14538 | glucosaminyl (N-acetyl) transferase 2_ I-branching enzyme | ENSMUSG00000021360 | 0.778 | 8.7 × 10−6 | 5.5 × 10−3 |
| Acap1 | 216859 | ArfGAP with coiled-coil_ ankyrin repeat and PH domains 1 | ENSMUSG00000001588 | 1.306 | 9.5 × 10−6 | 5.9 × 10−3 |
| Hist1h2bg | 319181 | histone cluster 1_ H2bg | ENSMUSG00000058385 | 1.002 | 1.0 × 10−5 | 6.5 × 10−3 |
| Kctd10 | 330171 | potassium channel tetramerisation domain containing 10 | ENSMUSG00000001098 | −0.635 | 1.2 × 10−5 | 7.2 × 10−3 |
| Porcn | 53627 | porcupine homolog (Drosophila) | ENSMUSG00000031169 | 1.051 | 1.2 × 10−5 | 7.2 × 10−3 |
| Fam162a | 70186 | family with sequence similarity 162_ member A | ENSMUSG00000003955 | 0.872 | 1.4 × 10−5 | 8.2 × 10−3 |
| Arhgap19 | 71085 | Rho GTPase activating protein 19 | ENSMUSG00000025154 | −0.806 | 1.8 × 10−5 | 1.0 × 10−2 |
| Sept11 | 52398 | septin 11 | ENSMUSG00000058013 | −0.757 | 1.9 × 10−5 | 1.0 × 10−2 |
| Fdft1 | 14137 | farnesyl diphosphate farnesyl transferase 1 | ENSMUSG00000021273 | −0.834 | 1.9 × 10−5 | 1.0 × 10−2 |
| Foxc1 | 17300 | forkhead box C1 | ENSMUSG00000050295 | −0.687 | 2.0 × 10−5 | 1.1 × 10−2 |
| Car12 | 76459 | carbonic anyhydrase 12 | ENSMUSG00000032373 | 1.240 | 2.0 × 10−5 | 1.1 × 10−2 |
| Fosl1 | 14283 | fos-like antigen 1 | ENSMUSG00000024912 | 0.952 | 2.4 × 10−5 | 1.2 × 10−2 |
| Fstl1 | 14314 | follistatin-like 1 | ENSMUSG00000022816 | −0.903 | 2.6 × 10−5 | 1.3 × 10−2 |
| Mt1 | 17748 | metallothionein 1 | ENSMUSG00000031765 | 1.264 | 2.6 × 10−5 | 1.3 × 10−2 |
| 2610528A11Rik | 70045 | RIKEN cDNA 2610528A11 gene | ENSMUSG00000096001 | 1.210 | 2.9 × 10−5 | 1.4 × 10−2 |
| Minpp1 | 17330 | multiple inositol polyphosphate histidine phosphatase 1 | ENSMUSG00000024896 | −0.857 | 2.9 × 10−5 | 1.4 × 10−2 |
| Lrig1 | 16206 | leucine-rich repeats and immunoglobulin-like domains 1 | ENSMUSG00000030029 | −0.607 | 3.2 × 10−5 | 1.5 × 10−2 |
| L1cam | 16728 | L1 cell adhesion molecule | ENSMUSG00000031391 | 1.072 | 3.4 × 10−5 | 1.6 × 10−2 |
| Ttll12 | 223723 | tubulin tyrosine ligase-like family_ member 12 | ENSMUSG00000016757 | −0.752 | 3.9 × 10−5 | 1.8 × 10−2 |
| Gpr35 | 64095 | G protein-coupled receptor 35 | ENSMUSG00000026271 | 1.193 | 3.8 × 10−5 | 1.8 × 10−2 |
| Gm14760 | 654474 | glyceraldehyde-3-phosphate dehydrogenase pseudogene | ENSMUSG00000081221 | 1.204 | 3.9 × 10−5 | 1.8 × 10−2 |
| Nde1 | 67203 | nuclear distribution gene E homolog 1 (A nidulans) | ENSMUSG00000022678 | −0.786 | 4.3 × 10−5 | 1.9 × 10−2 |
| 4930447F24Rik | 76873 | RIKEN cDNA 4930447F24 gene | ENSMUSG00000102224 | 0.976 | 4.2 × 10−5 | 1.9 × 10−2 |
| Csf2rb2 | 12984 | colony stimulating factor 2 receptor_ beta 2_ low-affinity (granulocyte-macrophage) | ENSMUSG00000071714 | 1.227 | 4.4 × 10−5 | 1.9 × 10−2 |
| Ankrd37 | 654824 | ankyrin repeat domain 37 | ENSMUSG00000050914 | 1.045 | 4.5 × 10−5 | 1.9 × 10−2 |
| Ola1 | 67059 | Obg-like ATPase 1 | ENSMUSG00000027108 | −0.598 | 4.8 × 10−5 | 2.0 × 10−2 |
| Gm43951 | NULL | predicted gene_ 43951 | ENSMUSG00000107877 | 1.208 | 4.8 × 10−5 | 2.0 × 10−2 |
| Plxnd1 | 67784 | plexin D1 | ENSMUSG00000030123 | 0.683 | 5.3 × 10−5 | 2.2 × 10−2 |
| Foxred2 | 239554 | FAD-dependent oxidoreductase domain containing 2 | ENSMUSG00000016552 | 0.650 | 5.6 × 10−5 | 2.3 × 10−2 |
| 9330151L19Rik | 414085 | RIKEN cDNA 9330151L19 gene | ENSMUSG00000097061 | 0.937 | 6.3 × 10−5 | 2.5 × 10−2 |
| Rac3 | 170758 | RAS-related C3 botulinum substrate 3 | ENSMUSG00000018012 | 0.831 | 6.7 × 10−5 | 2.6 × 10−2 |
| Dusp5 | 240672 | dual specificity phosphatase 5 | ENSMUSG00000034765 | 0.773 | 6.7 × 10−5 | 2.6 × 10−2 |
| Arl6ip5 | 65106 | ADP-ribosylation factor-like 6 interacting protein 5 | ENSMUSG00000035199 | 0.665 | 6.9 × 10−5 | 2.7 × 10−2 |
| Sort1 | 20661 | sortilin 1 | ENSMUSG00000068747 | −0.887 | 7.1 × 10−5 | 2.7 × 10−2 |
| Gm3200 | 100041204 | glyceraldehyde-3-phosphate dehydrogenase pseudogene | ENSMUSG00000097388 | 1.118 | 7.1 × 10−5 | 2.7 × 10−2 |
| Aldh3a2 | 11671 | aldehyde dehydrogenase family 3_ subfamily A2 | ENSMUSG00000010025 | −0.725 | 7.3 × 10−5 | 2.7 × 10−2 |
| Slc2a1 | 20525 | solute carrier family 2 (facilitated glucose transporter)_ member 1 | ENSMUSG00000028645 | 0.851 | 7.7 × 10−5 | 2.8 × 10−2 |
| Tusc2 | 80385 | tumor suppressor candidate 2 | ENSMUSG00000010054 | 0.675 | 8.5 × 10−5 | 3.1 × 10−2 |
| M6pr | 17113 | mannose-6-phosphate receptor_ cation dependent | ENSMUSG00000007458 | 0.610 | 8.9 × 10−5 | 3.2 × 10−2 |
| Pfkp | 56421 | phosphofructokinase_ platelet | ENSMUSG00000021196 | 0.844 | 8.9 × 10−5 | 3.2 × 10−2 |
| Cda | 72269 | cytidine deaminase | ENSMUSG00000028755 | 0.684 | 1.0 × 10−4 | 3.8 × 10−2 |
| Eef2k | 13631 | eukaryotic elongation factor-2 kinase | ENSMUSG00000035064 | 0.624 | 1.0 × 10−4 | 3.8 × 10−2 |
| Ogn | 18295 | osteoglycin | ENSMUSG00000021390 | −0.989 | 1.4 × 10−4 | 4.9 × 10−2 |
| Term Name | p-Value | FDR-Adjusted p-Value | N. of Genes that Are Also in This Filter or Cluster | N. of Upregulated Genes | N. of Downregulated Genes |
|---|---|---|---|---|---|
| HYPOXIA | 8.2 × 10−9 | 3.0 × 10−7 | 13 | 13 | 0 |
| HIF-1-α transcription factor network | 7.4 × 10−6 | 0.00020 | 6 | 6 | 0 |
| GSE21360_SECONDARY_VS_QUATERNARY_MEMORY_CD8_TCELL_UP | 3.8 × 10−7 | 0.00071 | 9 | 9 | 0 |
| GSE3982_DC_VS_BCELL_UP | 4.1 × 10−7 | 0.00071 | 9 | 8 | 1 |
| GSE21360_SECONDARY_VS_QUATERNARY_MEMORY_CD8_TCELL_DN | 1.8 × 10−6 | 0.00208 | 8 | 8 | 0 |
| GSE22611_NOD2_TRANSDUCED_VS_CTRL_HEK293T_STIMULATED_WITH_MDP_2H_DN | 5.1 × 10−6 | 0.00352 | 8 | 8 | 0 |
| GSE37416_CTRL_VS_6H_F_TULARENSIS_LVS_NEUTROPHIL_DN | 3.9 × 10−5 | 0.01260 | 7 | 7 | 0 |
| GSE360_CTRL_VS_L_DONOVANI_DC_DN | 3.3 × 10−5 | 0.01260 | 7 | 7 | 0 |
| GSE15930_NAIVE_VS_72H_IN_VITRO_STIM_IL12_CD8_TCELL_DN | 6.4 × 10−5 | 0.01271 | 7 | 5 | 2 |
| Glycogen biosynthesis II (from UDP-D-Glucose) | 0.00083 | 0.01995 | 2 | 2 | 0 |
| ESTROGEN_RESPONSE_LATE | 0.00201 | 0.03626 | 7 | 3 | 4 |
| GSE25677_MPL_VS_R848_STIM_BCELL_UP | 0.00031 | 0.04286 | 6 | 6 | 0 |
| GSE36078_UNTREATED_VS_AD5_INF_MOUSE_LUNG_DC_UP | 0.00043 | 0.04286 | 6 | 5 | 1 |
| GSE43955_1H_VS_42H_ACT_CD4_TCELL_WITH_TGFB_IL6_DN | 0.00044 | 0.04286 | 6 | 6 | 0 |
| GSE17721_CpG_VS_GARDIQUIMOD_1H_BMDC_UP | 0.00042 | 0.04286 | 6 | 4 | 2 |
| GSE3982_MAC_VS_BCELL_UP | 0.00039 | 0.04286 | 6 | 6 | 0 |
| GSE17301_CTRL_VS_48H_ACD3_ACD28_IFNA2_STIM_CD8_TCELL_UP | 0.00040 | 0.04286 | 6 | 4 | 2 |
| EPITHELIAL_MESENCHYMAL_TRANSITION | 0.00620 | 0.05306 | 6 | 5 | 1 |
| ESTROGEN_RESPONSE_EARLY | 0.00722 | 0.05306 | 6 | 3 | 3 |
| TNFA_SIGNALING_VIA_NFKB | 0.00758 | 0.05306 | 6 | 6 | 0 |
| Pathway | Gene | Description | Aliases |
|---|---|---|---|
| Hypoxia | Gbe1 | glucan (1,4-alpha), branching enzyme 1 | 2310045H19RIK, 2810426P10RIK, D16ERTD536E |
| Serpine1 | serine (or cysteine) peptidase inhibitor, clade E, member 1 | PAI-1, PAI1, PLANH1 | |
| Car12 | carbonic anhydrase 12 | 2310047E01RIK, AI314958, CA-XII, CA12 | |
| Fam162a | family with sequence similarity 162, member A | 2310056P07RIK, HGTD-P | |
| Gcnt2 | glucosaminyl (N-acetyl) transferase 2, I-branching enzyme | 5330430K10RIK, IGNT, IGNTA, IGNTB, IGNTC | |
| Selenbp1 | selenium binding protein 1 | LP56, LPSB, SBP56 | |
| P4ha2 | procollagen-proline, 2-oxoglutarate 4-dioxygenase (proline 4-hydroxylase), alphaII polypeptide | AA407196, C76437, P4HL | |
| Hmox1 | heme oxygenase 1 | D8WSU38E, HO-1, HO1, HEMOX, HMOX, HSP32 | |
| Slc2a1 | solute carrier family 2 (facilitated glucose transporter), member 1 | GLUT-1, GLUT1 | |
| Ero1l | ERO1-like (S. cerevisiae) | ERO1-L | |
| Ndrg1 | N-myc downstream-regulated gene 1 | CAP43, CMT4D, DRG1, HMSNL, NMSL, NDR1, NDRL, PROXY1, RTP, TDD5 | |
| Gys1 | glycogen synthase 1, muscle | GYS3, MGS | |
| Pfkp | phosphofructokinase, platelet | 1200015H23RIK, 9330125N24RIK, ATP-PFK, PFK-C, PFK-P | |
| HIF-1-alpha | Nos2 | nitric oxide synthase 2, inducible | MAC-NOS, NOS-II, NOS-2, NOS2A, I-NOS, INOS |
| Hmox1 | heme oxygenase 1 | D8WSU38E, HO-1, HO1, HEMOX, HMOX, HSP32 | |
| Serpine1 | serine (or cysteine) peptidase inhibitor, clade E, member 1 | PAI-1, PAI1, PLANH1 | |
| Id2 | inhibitor of DNA binding 2 | AI255428, C78922, IDB2, BHLHB26 | |
| Ndrg1 | N-myc downstream-regulated gene 1 | CAP43, CMT4D, DRG1, HMSNL, NMSL, NDR1, NDRL, PROXY1, RTP, TDD5 | |
| Slc2a1 | solute carrier family 2 (facilitated glucose transporter), member 1 | GLUT-1, GLUT1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fusco, C.; Nardella, G.; Di Filippo, L.; Dejana, E.; Cacchiarelli, D.; Petracca, A.; Micale, L.; Malinverno, M.; Castori, M. Transcriptome Analysis Reveals Altered Expression of Genes Involved in Hypoxia, Inflammation and Immune Regulation in Pdcd10-Depleted Mouse Endothelial Cells. Genes 2022, 13, 961. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13060961
Fusco C, Nardella G, Di Filippo L, Dejana E, Cacchiarelli D, Petracca A, Micale L, Malinverno M, Castori M. Transcriptome Analysis Reveals Altered Expression of Genes Involved in Hypoxia, Inflammation and Immune Regulation in Pdcd10-Depleted Mouse Endothelial Cells. Genes. 2022; 13(6):961. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13060961
Chicago/Turabian StyleFusco, Carmela, Grazia Nardella, Lucio Di Filippo, Elisabetta Dejana, Davide Cacchiarelli, Antonio Petracca, Lucia Micale, Matteo Malinverno, and Marco Castori. 2022. "Transcriptome Analysis Reveals Altered Expression of Genes Involved in Hypoxia, Inflammation and Immune Regulation in Pdcd10-Depleted Mouse Endothelial Cells" Genes 13, no. 6: 961. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13060961