
The Relationships between Dopaminergic, Glutamatergic, and Cognitive Functioning in 22q11.2 Deletion Syndrome: A Cross-Sectional, Multimodal 1H-MRS and 18F-Fallypride PET Study
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Procedure and Instruments
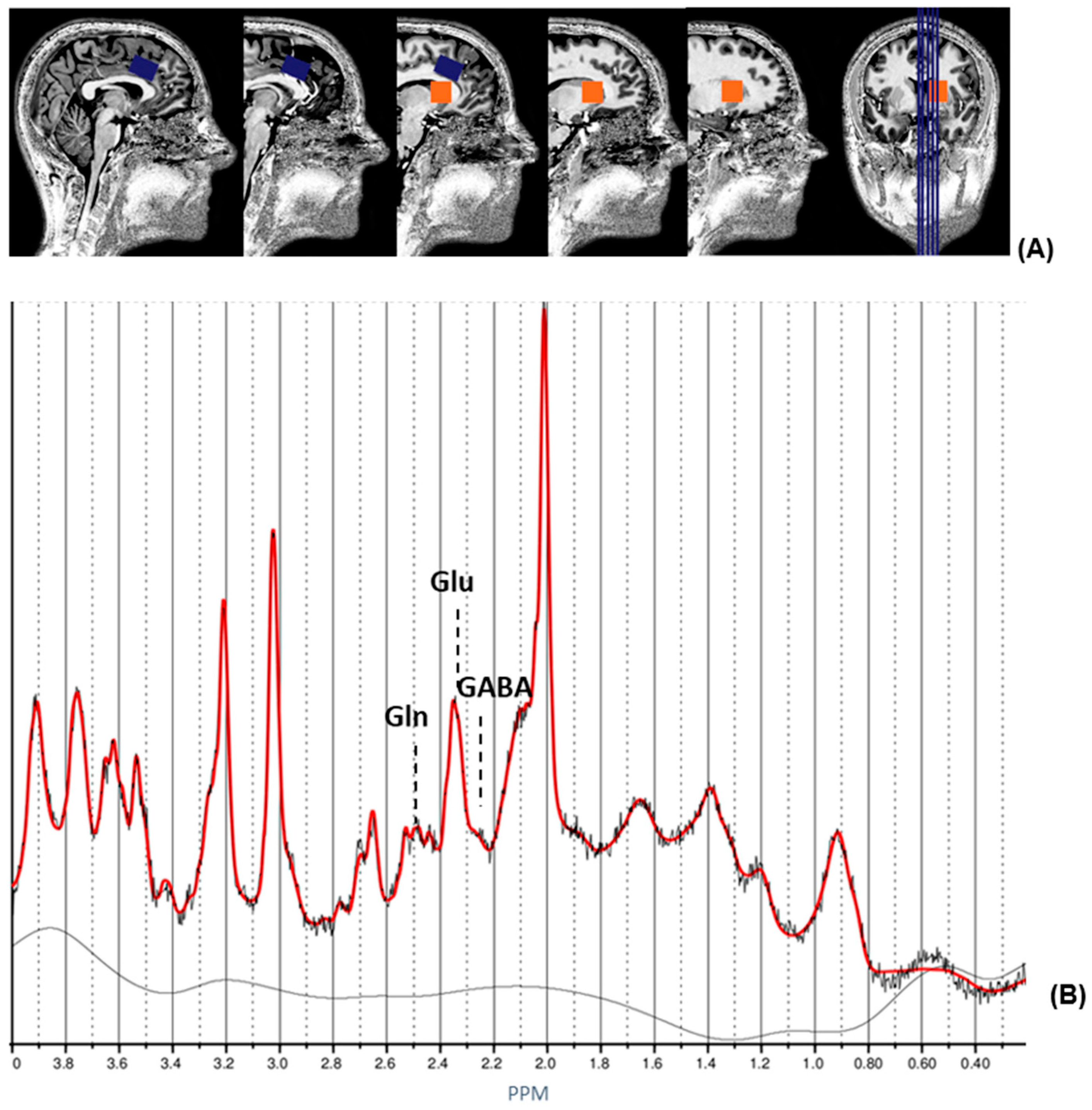
2.3. 1H magnetic Resonance Spectroscopy and Structural MRI
2.4. Positron Emission Tomography
2.5. Statistical Analyses
3. Results
3.1. Demographics
3.2. Association between Dopaminergic and Glutamatergic Functioning in 22q11DS
3.3. Association between Glutamatergic Functioning and ACC Volumes in 22q11DS and Healthy Controls
3.4. Association between Cognitive Functioning and Dopamine D2/3 Receptor Availability in 22q11DS
4. Discussion
4.1. Association between Dopaminergic and Glutamatergic Functioning in 22q11DS
4.2. Association between Glutamatergic Functioning and ACC Volumes in 22q11DS and Healthy Controls
4.3. Association between Cognitive Functioning and Dopamine D2/3 Receptor Availability in 22q11DS
4.4. Strengths and limitations
4.5. Implications and Suggestions for Future Work
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jonas, R.K.; Montojo, C.A.; Bearden, C.E. The 22q11.2 deletion syndrome as a window into complex neuropsychiatric disorders over the lifespan. Biol. Psychiatry 2014, 75, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Guna, A.; Butcher, N.J.; Bassett, A.S. Comparative mapping of the 22q11.2 deletion region and the potential of simple model organisms. J. Neurodev. Disord. 2015, 7, 18. [Google Scholar] [CrossRef] [PubMed]
- Bassett, A.S.; McDonald-McGinn, D.M.; Devriendt, K.; Digilio, M.C.; Goldenberg, P.; Habel, A.; Marino, B.; Oskarsdottir, S.; Philip, N.; Sullivan, K.; et al. Practical guidelines for managing patients with 22q11.2 deletion syndrome. J. Pediatr. 2011, 159, 332–339.e1. [Google Scholar] [CrossRef] [PubMed]
- Schneider, M.; Debbané, M.; Bassett, A.S.; Chow, E.W.; Fung, W.L.A.; Van Den Bree, M.B.; Owen, M.; Murphy, K.C.; Niarchou, M.; Kates, W.R.; et al. Psychiatric disorders from childhood to adulthood in 22q11.2 deletion syndrome: Results from the International Consortium on Brain and Behavior in 22q11.2 Deletion Syndrome. Am. J. Psychiatry 2014, 171, 627–639. [Google Scholar] [CrossRef]
- Perälä, J.; Suvisaari, J.; Saarni, S.I.; Kuoppasalmi, K.; Isometsä, E.; Pirkola, S.; Partonen, T.; Tuulio-Henriksson, A.; Hintikka, J.; Kieseppä, T. Lifetime prevalence of psychotic and bipolar I disorders in a general population. Arch. Gen. Psychiatry 2007, 64, 19–28. [Google Scholar] [CrossRef]
- Vorstman, J.A.; Breetvelt, E.J.; Duijff, S.N.; Eliez, S.; Schneider, M.; Jalbrzikowski, M.; Armando, M.; Vicari, S.; Shashi, V.; Hooper, S.R.; et al. Cognitive decline preceding the onset of psychosis in patients with 22q11.2 deletion syndrome. JAMA Psychiatry 2015, 72, 377–385. [Google Scholar] [CrossRef]
- Evers, L.; Van Amelsvoort, T.; Candel, M.; Boer, H.; Engelen, J.M.; Curfs, L. Psychopathology in adults with 22q11 deletion syndrome and moderate and severe intellectual disability. J. Intellect. Disabil. Res. 2014, 58, 915–925. [Google Scholar] [CrossRef]
- Yavich, L.; Forsberg, M.M.; Karayiorgou, M.; Gogos, J.A.; Männistö, P.T. Site-specific role of catechol-O-methyltransferase in dopamine overflow within prefrontal cortex and dorsal striatum. J. Neurosci. 2007, 27, 10196–10209. [Google Scholar] [CrossRef]
- Rogdaki, M.; Devroye, C.; Ciampoli, M.; Veronese, M.; Ashok, A.H.; McCutcheon, R.A.; Jauhar, S.; Bonoldi, I.; Gudbrandsen, M.; Daly, E.; et al. Striatal dopaminergic alterations in individuals with copy number variants at the 22q11.2 genetic locus and their implications for psychosis risk: A [18F]-DOPA PET study. Mol. Psychiatry 2021, 1–12. [Google Scholar] [CrossRef]
- Van Duin, E.D.; Ceccarini, J.; Booij, J.; Kasanova, Z.; Vingerhoets, C.; van Huijstee, J.; Heinzel, A.; Mohammadkhani-Shali, S.; Winz, O.; Mottaghy, F.; et al. Lower [18F] fallypride binding to dopamine D2/3 receptors in frontal brain areas in adults with 22q11.2 deletion syndrome: A positron emission tomography study. Psychol. Med. 2020, 50, 799–807. [Google Scholar] [CrossRef]
- Cohen, S.M.; Nadler, J.V. Proline-induced potentiation of glutamate transmission. Brain Res. 1997, 761, 271–282. [Google Scholar] [CrossRef]
- Henzi, V.; Reichling, D.B.; Helm, S.W.; MacDermott, A.B. L-proline activates glutamate and glycine receptors in cultured rat dorsal horn neurons. Mol. Pharmacol. 1992, 41, 793–801. [Google Scholar] [PubMed]
- Goodman, B.; Rutberg, J.; Lin, W.; Pulver, A.; Thomas, G.; Geraghty, M. Hyperprolinaemia in patients with deletion (22)(q11. 2) syndrome. J. Inherit. Metab. Dis. 2000, 23, 847–848. [Google Scholar] [CrossRef]
- Evers, L.J.; van Amelsvoort, T.A.; Bakker, J.A.; de Koning, M.; Drukker, M.; Curfs, L.M. Glutamatergic markers, age, intellectual functioning and psychosis in 22q11 deletion syndrome. Psychopharmacology 2015, 232, 3319–3325. [Google Scholar] [CrossRef] [PubMed]
- Raux, G.; Bumsel, E.; Hecketsweiler, B.; Van Amelsvoort, T.; Zinkstok, J.; Manouvrier-Hanu, S.; Fantini, C.; Brévière, G.-M.M.; Di Rosa, G.; Pustorino, G.; et al. Involvement of hyperprolinemia in cognitive and psychiatric features of the 22q11 deletion syndrome. Hum. Mol. Genet. 2007, 16, 83–91. [Google Scholar] [CrossRef]
- Cohen, S.M.; Nadler, J.V. Proline-induced inhibition of glutamate release in hippocampal area CA1. Brain Res. 1997, 769, 333–339. [Google Scholar] [CrossRef]
- Lau, A.; Tymianski, M. Glutamate receptors, neurotoxicity and neurodegeneration. Pflügers Arch.-Eur. J. Physiol. 2010, 460, 525–542. [Google Scholar] [CrossRef]
- Deutsch, S.I.; Rosse, R.B.; Schwartz, B.L.; Mastropaolo, J. A revised excitotoxic hypothesis of schizophrenia: Therapeutic implications. Clin. Neuropharmacol. 2001, 24, 43–49. [Google Scholar] [CrossRef]
- Sun, D.; Ching, C.R.K.; Lin, A.; Forsyth, J.K.; Kushan, L.; Vajdi, A.; Jalbrzikowski, M.; Hansen, L.; Villalon-Reina, J.E.; Qu, X.; et al. Large-scale mapping of cortical alterations in 22q11.2 deletion syndrome: Convergence with idiopathic psychosis and effects of deletion size. Mol. Psychiatry 2020, 25, 1822–1834. [Google Scholar] [CrossRef]
- Rogdaki, M.; Hathway, P.; Gudbrandsen, M.; McCutcheon, R.A.; Jauhar, S.; Daly, E.; Howes, O. Glutamatergic function in a genetic high-risk group for psychosis: A proton magnetic resonance spectroscopy study in individuals with 22q11.2 deletion. Eur. Neuropsychopharmacol. 2019, 29, 1333–1342. [Google Scholar] [CrossRef]
- Vingerhoets, C.; Tse, D.H.; van Oudenaren, M.; Hernaus, D.; van Duin, E.; Zinkstok, J.; Ramaekers, J.G.; Jansen, J.F.; McAlonan, G.; van Amelsvoort, T. Glutamatergic and GABAergic reactivity and cognition in 22q11.2 deletion syndrome and healthy volunteers: A randomized double-blind 7-Tesla pharmacological MRS study. J. Psychopharmacol. 2020, 34, 856–863. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Alves, F.; Boot, E.; Schmitz, N.; Nederveen, A.; Vorstman, J.; Lavini, C.; Pouwels, P.; de Haan, L.; Linszen, D.; van Amelsvoort, T. Proton magnetic resonance spectroscopy in 22q11 deletion syndrome. PLoS ONE 2011, 6, e21685. [Google Scholar] [CrossRef]
- Brugger, S.P.; Angelescu, I.; Abi-Dargham, A.; Mizrahi, R.; Shahrezaei, V.; Howes, O.D. Heterogeneity of striatal dopamine function in schizophrenia: Meta-analysis of variance. Biol. Psychiatry 2020, 87, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Howes, O.D.; Montgomery, A.J.; Asselin, M.C.; Murray, R.M.; Valli, I.; Tabraham, P.; Bramon-Bosch, E.; Valmaggia, L.; Johns, L.; Broome, M.; et al. Elevated striatal dopamine function linked to prodromal signs of schizophrenia. Arch. Gen. Psychiatry 2009, 66, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Huttunen, J.; Heinimaa, M.; Svirskis, T.; Nyman, M.; Kajander, J.; Forsback, S.; Solin, O.; Ilonen, T.; Korkeila, J.; Ristkari, T.; et al. Striatal dopamine synthesis in first-degree relatives of patients with schizophrenia. Biol. Psychiatry 2008, 63, 114–117. [Google Scholar] [CrossRef]
- McCutcheon, R.; Beck, K.; Jauhar, S.; Howes, O.D. Defining the locus of dopaminergic dysfunction in schizophrenia: A meta-analysis and test of the mesolimbic hypothesis. Schizophr. Bull. 2018, 44, 1301–1311. [Google Scholar] [CrossRef] [PubMed]
- Howes, O.; McCutcheon, R.; Stone, J. Glutamate and dopamine in schizophrenia: An update for the 21st century. J. Psychopharmacol. 2015, 29, 97–115. [Google Scholar] [CrossRef]
- Kokkinou, M.; Ashok, A.H.; Howes, O.D. The effects of ketamine on dopaminergic function: Meta-analysis and review of the implications for neuropsychiatric disorders. Mol. Psychiatry 2018, 23, 59–69. [Google Scholar] [CrossRef]
- Breier, A.; Adler, C.M.; Weisenfeld, N.; Su, T.P.; Elman, I.; Picken, L.; Malhotra, A.K.; Pickar, D. Effects of NMDA antagonism on striatal dopamine release in healthy subjects: Application of a novel PET approach. Synapse 1998, 29, 142–147. [Google Scholar] [CrossRef]
- Vollenweider, F.X.; Vontobel, P.; Øye, I.; Hell, D.; Leenders, K.L. Effects of (S)-ketamine on striatal dopamine: A [11C] raclopride PET study of a model psychosis in humans. J. Psychiatr. Res. 2000, 34, 35–43. [Google Scholar] [CrossRef]
- Jauhar, S.; McCutcheon, R.; Borgan, F.; Veronese, M.; Nour, M.; Pepper, F.; Rogdaki, M.; Stone, J.; Egerton, A.; Turkheimer, F.; et al. The relationship between cortical glutamate and striatal dopamine in first-episode psychosis: A cross-sectional multimodal PET and magnetic resonance spectroscopy imaging study. Lancet Psychiatry 2018, 5, 816–823. [Google Scholar] [CrossRef]
- Van Duin, E.D.; Kasanova, Z.; Hernaus, D.; Ceccarini, J.; Heinzel, A.; Mottaghy, F.; Mohammadkhani-Shali, S.; Winz, O.; Frank, M.; Beck, M.C.; et al. Striatal dopamine release and impaired reinforcement learning in adults with 22q11.2 deletion syndrome. Eur. Neuropsychopharmacol. 2018, 28, 732–742. [Google Scholar] [CrossRef] [PubMed]
- Sheehan, D. MINI-Mini International neuropsychiatric interview-english version 5.0. 0-DSM-IV. J Clin Psychiatry 1998, 59, 34–57. [Google Scholar]
- Levaux, M.N.; Potvin, S.; Sepehry, A.A.; Sablier, J.; Mendrek, A.; Stip, E. Computerized assessment of cognition in schizophrenia: Promises and pitfalls of CANTAB. Eur. Psychiatry 2007, 22, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Velthorst, E.; Levine, S.Z.; Henquet, C.; De Haan, L.; Van Os, J.; Myin-Germeys, I.; Reichenberg, A. To cut a short test even shorter: Reliability and validity of a brief assessment of intellectual ability in schizophrenia—A control-case family study. Cogn. Neuropsychiatry 2013, 18, 574–593. [Google Scholar] [CrossRef]
- Frahm, J.; Merboldt, K.-D.; Hänicke, W. Localized proton spectroscopy using stimulated echoes. J. Magn. Reson. 1987, 72, 502–508. [Google Scholar] [CrossRef]
- Provencher, S.W. Automatic quantitation of localized in vivo 1H spectra with LCModel. NMR Biomed. 2001, 14, 260–264. [Google Scholar] [CrossRef]
- Smith, S.; Levante, T.; Meier, B.H.; Ernst, R.R. Computer simulations in magnetic resonance. An object-oriented programming approach. J. Magn. Reson. Ser. A 1994, 106, 75–105. [Google Scholar] [CrossRef]
- Quadrelli, S.; Mountford, C.; Ramadan, S. Hitchhiker’s guide to voxel segmentation for partial volume correction of in vivo magnetic resonance spectroscopy. Magn. Reson. Insights 2016, 9, MRI-S32903. [Google Scholar] [CrossRef]
- Marques, J.P.; Kober, T.; Krueger, G.; van der Zwaag, W.; Van de Moortele, P.F.; Gruetter, R. MP2RAGE, a self bias-field corrected sequence for improved segmentation and T1-mapping at high field. Neuroimage 2010, 49, 1271–1281. [Google Scholar] [CrossRef]
- Fischl, B. FreeSurfer. Neuroimage 2012, 62, 774–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serrarens, C.; Otter, M.; Campforts, B.; Stumpel, C.T.; Jansma, H.; van Amelsvoort, T.A.; Vingerhoets, C. Altered subcortical and cortical brain morphology in adult women with 47,XXX: A 7-Tesla magnetic resonance imaging study. J. Neurodev. Disord. 2022, 14, 14. [Google Scholar] [CrossRef] [PubMed]
- Kasanova, Z.; Ceccarini, J.; Frank, M.J.; van Amelsvoort, T.; Booij, J.; Heinzel, A.; Mottaghy, F.; Myin-Germeys, I. Striatal dopaminergic modulation of reinforcement learning predicts reward—Oriented behavior in daily life. Biol. Psychol. 2017, 127, 1–9. [Google Scholar] [CrossRef]
- Lammertsma, A.A.; Hume, S.P. Simplified reference tissue model for PET receptor studies. Neuroimage 1996, 4, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Alpert, N.M.; Badgaiyan, R.D.; Livni, E.; Fischman, A.J. A novel method for noninvasive detection of neuromodulatory changes in specific neurotransmitter systems. Neuroimage 2003, 19, 1049–1060. [Google Scholar] [CrossRef]
- Stone, J.M.; Howes, O.D.; Egerton, A.; Kambeitz, J.; Allen, P.; Lythgoe, D.J.; O’Gorman, R.L.; McLean, M.A.; Barker, G.J.; McGuire, P. Altered relationship between hippocampal glutamate levels and striatal dopamine function in subjects at ultra high risk of psychosis. Biol. Psychiatry 2010, 68, 599–602. [Google Scholar] [CrossRef]
- Kegeles, L.S.; Abi-Dargham, A.; Frankle, W.G.; Gil, R.; Cooper, T.B.; Slifstein, M.; Hwang, D.R.; Huang, Y.; Haber, S.N.; Laruelle, M. Increased synaptic dopamine function in associative regions of the striatum in schizophrenia. Arch. Gen. Psychiatry 2010, 67, 231–239. [Google Scholar] [CrossRef]
- Mintun, M.A.; Raichle, M.E.; Kilbourn, M.R.; Wooten, G.F.; Welch, M.J. A quantitative model for the in vivo assessment of drug binding sites with positron emission tomography. Ann. Neurol. Off. J. Am. Neurol. Assoc. Child Neurol. Soc. 1984, 15, 217–227. [Google Scholar] [CrossRef]
- Haijma, S.V.; Van Haren, N.; Cahn, W.; Koolschijn, P.C.M.; Hulshoff Pol, H.E.; Kahn, R.S. Brain volumes in schizophrenia: A meta-analysis in over 18,000 subjects. Schizophr. Bull. 2013, 39, 1129–1138. [Google Scholar] [CrossRef]
- Marsman, A.; Mandl, R.C.; Klomp, D.W.; Bohlken, M.M.; Boer, V.O.; Andreychenko, A.; Cahn, W.; Kahn, R.S.; Luijten, P.R.; Pol, H.E.H. GABA and glutamate in schizophrenia: A 7 T 1H-MRS study. NeuroImage Clin. 2014, 6, 398–407. [Google Scholar] [CrossRef]
- Stone, J.M.; Day, F.; Tsagaraki, H.; Valli, I.; McLean, M.A.; Lythgoe, D.J.; O’Gorman, R.L.; Barker, G.J.; McGuire, P.K.; OASIS. Glutamate dysfunction in people with prodromal symptoms of psychosis: Relationship to gray matter volume. Biol. Psychiatry 2009, 66, 533–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraguljac, N.V.; White, D.M.; Reid, M.A.; Lahti, A.C. Increased hippocampal glutamate and volumetric deficits in unmedicated patients with schizophrenia. JAMA Psychiatry 2013, 70, 1294–1302. [Google Scholar] [CrossRef] [PubMed]
- Plitman, E.; Patel, R.; Chung, J.K.; Pipitone, J.; Chavez, S.; Reyes-Madrigal, F.; Gómez-Cruz, G.; León-Ortiz, P.; Chakravarty, M.M.; de la Fuente-Sandoval, C.; et al. Glutamatergic metabolites, volume and cortical thickness in antipsychotic-naive patients with first-episode psychosis: Implications for excitotoxicity. Neuropsychopharmacology 2016, 41, 2606–2613. [Google Scholar] [CrossRef] [PubMed]
- Rea, L.M.; Parker, R.A. Designing and Conducting Survey Research: A Comprehensive Guide; John Wiley & Sons: Hoboken, NJ, USA, 2014. [Google Scholar]
- Bäckman, L.; Ginovart, N.; Dixon, R.A.; Wahlin, T.B.R.; Wahlin, Å.; Halldin, C.; Farde, L. Age-related cognitive deficits mediated by changes in the striatal dopamine system. Am. J. Psychiatry 2000, 157, 635–637. [Google Scholar] [CrossRef] [PubMed]
- Cervenka, S.; Bäckman, L.; Cselényi, Z.; Halldin, C.; Farde, L. Associations between dopamine D2-receptor binding and cognitive performance indicate functional compartmentalization of the human striatum. Neuroimage 2008, 40, 1287–1295. [Google Scholar] [CrossRef]
- Chen, P.S.; Yang, Y.K.; Lee, Y.S.; Yeh, T.L.; Lee, I.H.; Chiu, N.T.; Chu, C.L. Correlation between different memory systems and striatal dopamine D2/D3 receptor density: A single photon emission computed tomography study. Psychol. Med. 2005, 35, 197–204. [Google Scholar] [CrossRef]
- Reeves, S.J.; Grasby, P.M.; Howard, R.J.; Bantick, R.A.; Asselin, M.C.; Mehta, M.A. A positron emission tomography (PET) investigation of the role of striatal dopamine (D2) receptor availability in spatial cognition. Neuroimage 2005, 28, 216–226. [Google Scholar] [CrossRef]
- Volkow, N.D.; Gur, R.C.; Wang, G.J.; Fowler, J.S.; Moberg, P.J.; Ding, Y.S.; Hitzemann, R.; Smith, G.; Logan, J. Association between decline in brain dopamine activity with age and cognitive and motor impairment in healthy individuals. Am. J. Psychiatry 1998, 155, 344–349. [Google Scholar]
- Goldman-Rakic, P.S.; Muly, E.C., III; Williams, G.V. D₁ receptors in prefrontal cells and circuits. Brain Res. Rev. 2000, 31, 295–301. [Google Scholar] [CrossRef]
- Cox, S.M.; Frank, M.J.; Larcher, K.; Fellows, L.K.; Clark, C.A.; Leyton, M.; Dagher, A. Striatal D1 and D2 signaling differentially predict learning from positive and negative outcomes. Neuroimage 2015, 109, 95–101. [Google Scholar] [CrossRef]
- Damsa, C.; Bumb, A.; Bianchi-Demicheli, F.; Vidailhet, P.; Sterck, R.; Andreoli, A.; Beyenburg, S. “Dopamine-dependent” side effects of selective serotonin reuptake inhibitors: A clinical review. J. Clin. Psychiatry 2004, 65, 4690. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| PET 22q11DS (N = 10) Mean (SD) | MRI 22q11DS (N = 17) Mean (SD) | MRI HC (N = 20) Mean (SD) | Statistic | p-Value | |
|---|---|---|---|---|---|
| Sex (F/M) | 5/5 | 11/6 | 12/8 | 0.09 | 0.77 1 |
| Age, years | 37.07 (11.12) | 34.17 (11.41) | 30.70 (8.20) | 145.00 | 0.46 2 |
| FSIQ | 82.60 (12.23) | 76.65 (12.32) | 120.21 (16.23) 3 | 4.50 | <0.0012 |
| Smoking in the previous year (yes/no) | 0/9 3 | 2/13 3 | 2/16 3 | NA | 1.00 4 |
| Current SSRI use (yes/no) | 1/9 | 2/15 | 1/19 | NA | 0.58 4 |
| Time between MRI and PET scan, days | 180.10 (349.52) | NA | NA | NA | NA |
| ACC Glutamate | ACC Glutamine | ACC Glx | Striatum Glutamate | Striatum Glutamine | Striatum Glx | |
|---|---|---|---|---|---|---|
| BPND 18F-fallypride ACC | r = 0.15 p = 0.68 | r = 0.01 p = 0.99 | r = 0.07 p = 0.86 | r = 0.47 p = 0.17 | r = 0.18 p = 0.64 | r = 0.56 p = 0.09 |
| BPND 18F-fallypride CNC (mean) | r = −0.33 p = 0.35 | r = −0.27 p = 0.45 | r = −0.46 p = 0.19 | r = 0.21 p = 0.56 | r = 0.27 p = 0.49 | r = 0.17 p = 0.65 |
| BPND 18F-fallypride putamen (mean) | r = −0.52 p = 0.13 | r = −0.31 p = 0.39 | r = −0.46 p = 0.19 | r = −0.10 p = 0.78 | r = 0.10 p = 0.80 | r = −0.07 p = 0.86 |
| BPND 18F-fallypride VST (mean) | r = −0.31 p = 0.39 | r = −0.20 p = 0.58 | r = −0.30 p = 0.41 | r = 0.30 p = 0.41 | r = −0.33 p = 0.38 | r = 0.19 p = 0.60 |
| Left Rostral ACC Volume | Right Rostral ACC Volume | Left Caudal ACC Volume | Right Caudal ACC Volume | ||
|---|---|---|---|---|---|
| 22q11DS | ACC glutamate | r = 0.34 p = 0.19 | r = 0.05 p = 0.85 | r = 0.36 p = 0.18 | r = −0.37 p = 0.16 |
| ACC glutamine | r = 0.30 p = 0.25 | r = −0.01 p = 0.98 | r = 0.03 p = 0.92 | r = −0.12 p = 0.67 | |
| ACC Glx | r = 0.01 p = 0.98 | r = −0.30 p = 0.26 | r = 0.14 p = 0.59 | r = −0.43 p = 0.09 | |
| Striatum glutamate | r = −0.45 p = 0.08 | r = −0.05 p = 0.85 | r = 0.09 p = 0.74 | r = 0.01 p = 0.96 | |
| Striatum glutamine | r = −0.05 p = 0.85 | r = 0.20 p = 0.48 | r = 0.12 p = 0.67 | r = −0.21 p = 0.44 | |
| Striatum Glx | r = 0.02 p = 0.94 | r = 0.12 p = 0.66 | r = 0.31 p = 0.25 | r = −0.09 p = 0.73 | |
| HC | ACC glutamate | r = 0.22 p = 0.36 | r = 0.49 p = 0.03 | r = −0.14 p = 0.54 | r = 0.12 p = 0.61 |
| ACC glutamine | r = 0.09 p = 0.71 | r = −0.15 p = 0.54 | r = 0.51 p = 0.03 | r = −0.11 p = 0.65 | |
| ACC Glx | r = 0.10 p = 0.67 | r = 0.25 p = 0.29 | r = 0.25 p = 0.30 | r = −0.53 p = 0.02 | |
| Striatum glutamate | r = -0.01 p = 0.97 | r = 0.40 p = 0.08 | r = −0.18 p = 0.44 | r = 0.31 p = 0.18 | |
| Striatum glutamine | r = −0.33 p = 0.23 | r = -0.31 p = 0.26 | r = −0.07 p = 0.80 | r = −0.37 p = 0.18 | |
| Striatum Glx | r = −0.16 p = 0.49 | r = 0.14 p = 0.54 | r = −0.39 p = 0.09 | r = 0.15 p = 0.53 |
| BPND 18F-Fallypride ACC | BPND 18F-Fallypride CNC (Mean) | BPND 18F-Fallypride Putamen (Mean) | BPND 18F-Fallypride VST (Mean) | |
|---|---|---|---|---|
| Visual memory | r = −0.72 p = 0.02 | r = 0.21 p = 0.56 | r = 0.36 p = 0.31 | r = −0.21 p = 0.56 |
| Verbal memory | r = −0.62 p > 0.05 | r = 0.09 p = 0.80 | r = -0.08 p = 0.83 | r = -0.56 p = 0.09 |
| Working memory | r = −0.63 p > 0.05 | r = −0.03 p = 0.93 | r = 0.24 p = 0.50 | r = −0.26 p = 0.47 |
| Attention 2 | r = −0.55 p = 0.13 | r = 0.20 p = 0.61 | r = 0.07 p = 0.87 | r = −0.33 p = 0.38 |
| Processing speed | r = -0.03 p = 0.93 | r = 0.15 p = 0.68 | r = −0.13 p = 0.73 | r = −0.02 p = 0.96 |
| Executive functioning | r = −0.74 p = 0.01 | r = −0.29 p = 0.42 | r = −0.08 p = 0.83 | r = -0.24 p = 0.51 |
| Social cognition | r = −0.60 p = 0.07 | r = 0.09 p = 0.80 | r = 0.12 p = 0.74 | r = −0.46 p = 0.18 |
| Composite score | r = −0.78 p = 0.01 | r = 0.06 p = 0.88 | r = 0.07 p = 0.86 | r = -0.43 p = 0.21 |
| FSIQ | r = −0.45 p = 0.19 | r = 0.26 p = 0.47 | r = 0.34 p = 0.34 | r = 0.27 p = 0.46 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Hooijdonk, C.F.M.; Tse, D.H.Y.; Roosenschoon, J.; Ceccarini, J.; Booij, J.; van Amelsvoort, T.A.M.J.; Vingerhoets, C. The Relationships between Dopaminergic, Glutamatergic, and Cognitive Functioning in 22q11.2 Deletion Syndrome: A Cross-Sectional, Multimodal 1H-MRS and 18F-Fallypride PET Study. Genes 2022, 13, 1672. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13091672
van Hooijdonk CFM, Tse DHY, Roosenschoon J, Ceccarini J, Booij J, van Amelsvoort TAMJ, Vingerhoets C. The Relationships between Dopaminergic, Glutamatergic, and Cognitive Functioning in 22q11.2 Deletion Syndrome: A Cross-Sectional, Multimodal 1H-MRS and 18F-Fallypride PET Study. Genes. 2022; 13(9):1672. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13091672
Chicago/Turabian Stylevan Hooijdonk, Carmen F. M., Desmond H. Y. Tse, Julia Roosenschoon, Jenny Ceccarini, Jan Booij, Therese A. M. J. van Amelsvoort, and Claudia Vingerhoets. 2022. "The Relationships between Dopaminergic, Glutamatergic, and Cognitive Functioning in 22q11.2 Deletion Syndrome: A Cross-Sectional, Multimodal 1H-MRS and 18F-Fallypride PET Study" Genes 13, no. 9: 1672. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13091672