
New Genetically Determined Markers of the Functional State of the Cardiovascular System
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
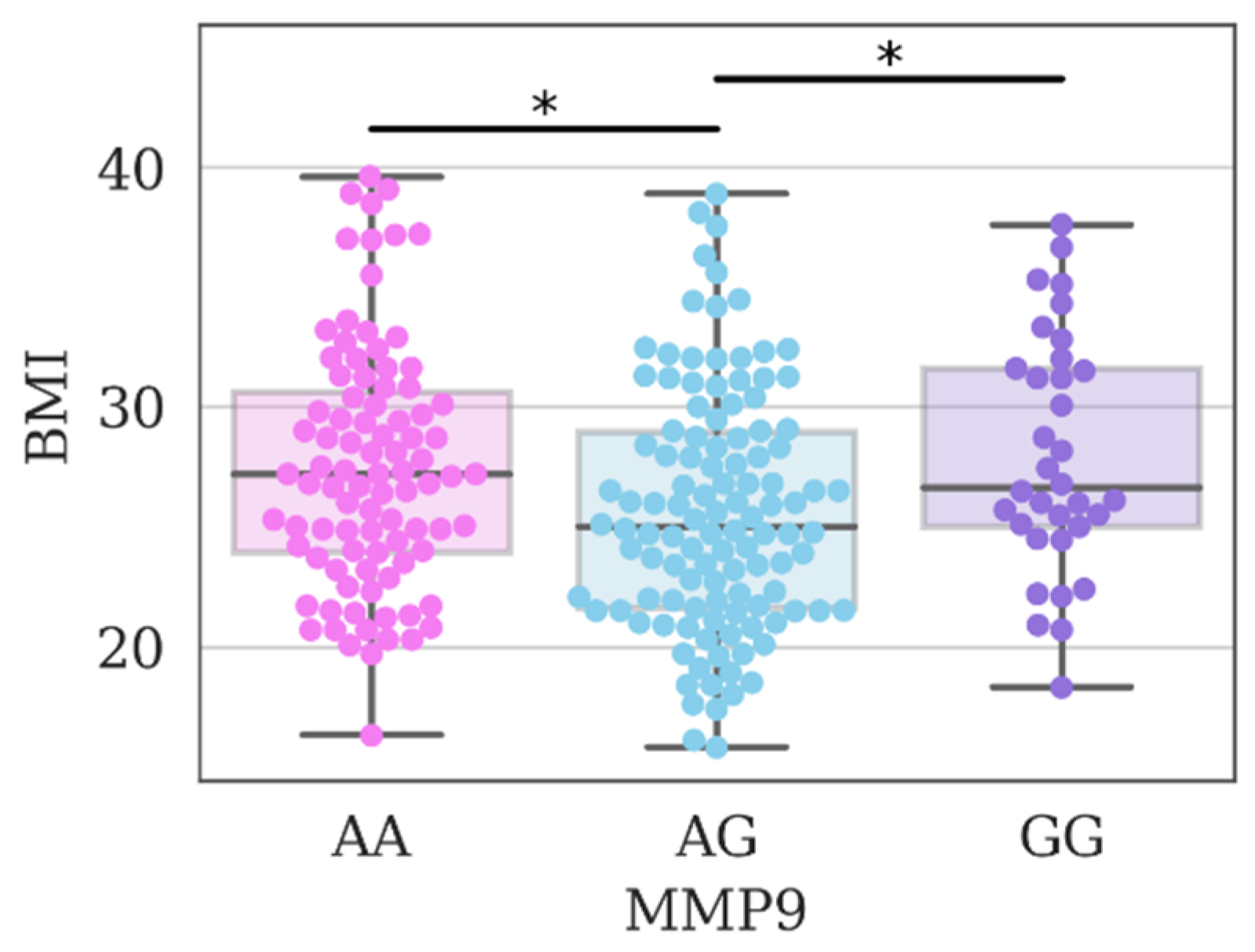
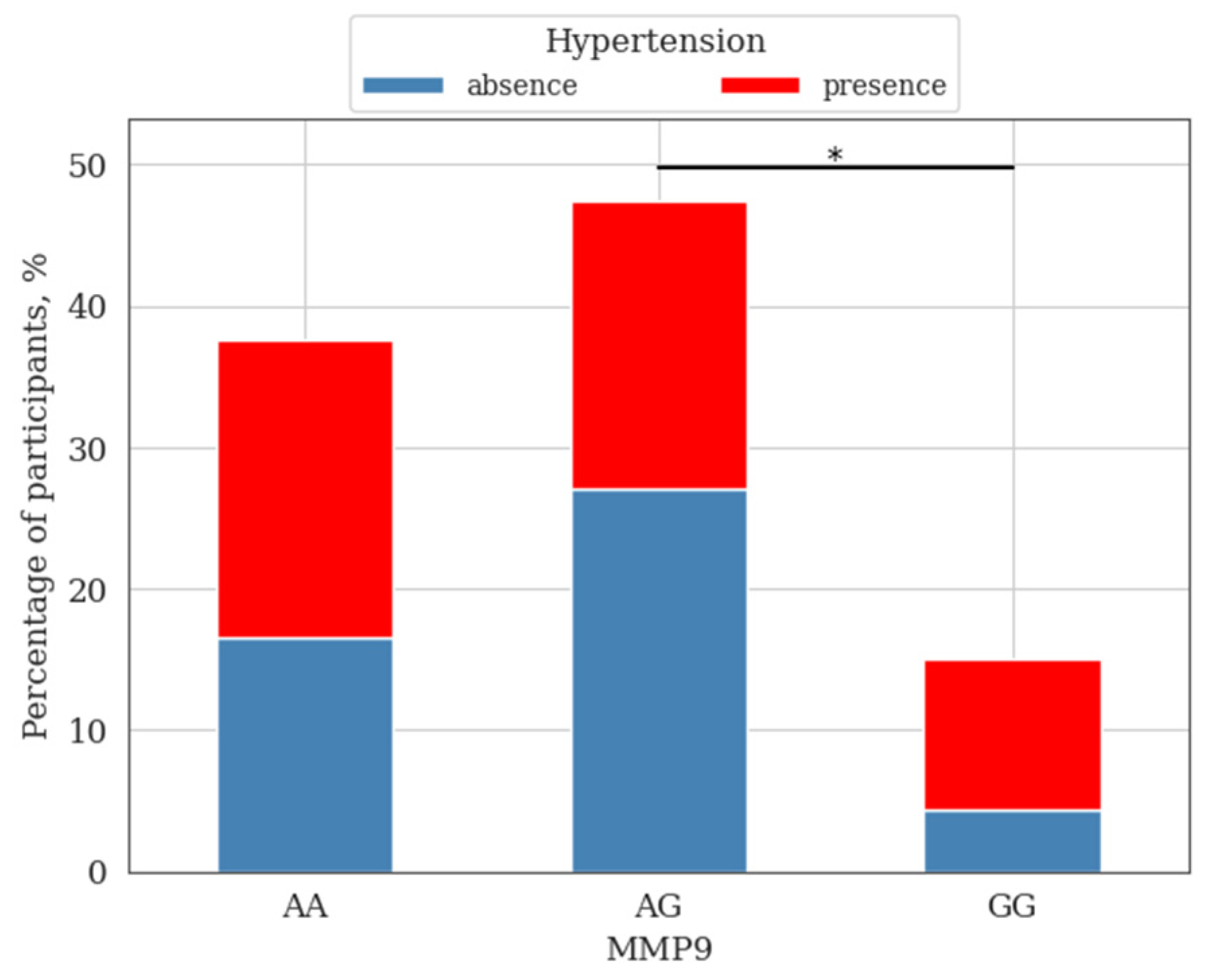
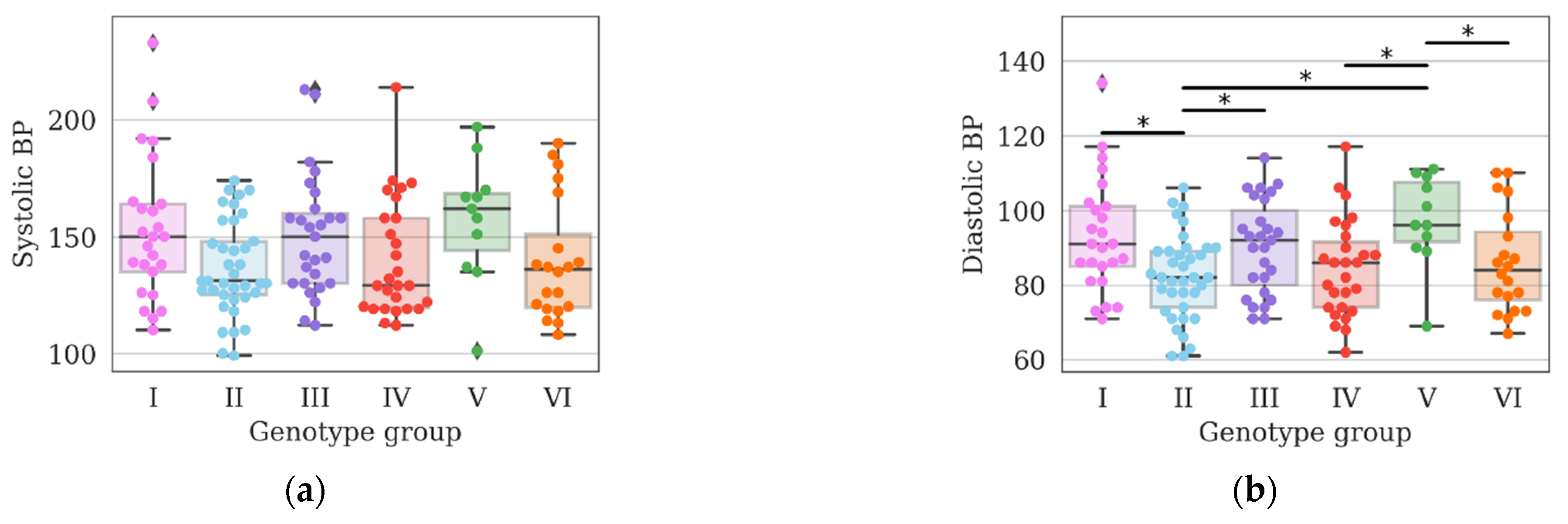
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ho, W.M.; Wu, Y.Y.; Chen, Y.C. Genetic Variants behind Cardiovascular Diseases and Dementia. Genes 2020, 11, 1514. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Ma, B.; Wu, X.; Wang, H.; Zhou, B. Long-Term Effect of Salt Substitute on All-Cause and Cardiovascular Disease Mortality: An Exploratory Follow-Up of a Randomized Controlled Trial. Front. Cardiovasc. Med. 2021, 8, 645902. [Google Scholar] [CrossRef] [PubMed]
- Gkaliagkousi, E.; Lazaridis, A.; Dogan, S.; Fraenkel, E.; Tuna, B.G.; Mozos, I.; Vukicevic, M.; Yalcin, O.; Gopcevic, K. Theories and Molecular Basis of Vascular Aging: A Review of the Literature from VascAgeNet Group on Pathophysiological Mechanisms of Vascular Aging. Int. J. Mol. Sci. 2022, 23, 8672. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, S.M.; Schwartz, H.T.; Horvath, S.; Schadt, E.; Lee, S.I. A Systematic Approach to Multifactorial Cardiovascular Disease. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 2821–2835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernlund, E.; Kissopoulou, A.; Green, H.; Karlsson, J.-E.; Ellegård, R.; Årstrand, H.K.; Jonasson, J.; Gunnarsson, C. Hereditary Hypertrophic Cardiomyopathy in Children and Young Adults—The Value of Reevaluating and Expanding Gene Panel Analyses. Genes. 2020, 11, 1472. [Google Scholar] [CrossRef]
- Kim, J.B. Channelopathies. Korean J. Pediatr. 2014, 57, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Creamer, T.J.; Bramel, E.E.; MacFarlane, E.G. Insights on the Pathogenesis of Aneurysm through the Study of Hereditary Aortopathies. Genes 2021, 12, 183. [Google Scholar] [CrossRef]
- Khera, A.V.; Kathiresan, S. Genetics of coronary artery disease: Discovery, biology and clinical translation. Nat. Rev. Genet. 2017, 18, 331–344. [Google Scholar] [CrossRef]
- Vesa, S.C.; Vlaicu, S.I.; Vacaras, V.; Crisan, S.; Sabin, O.; Pasca, S.; Trifa, A.P.; Rusz-Fogarasi, T.; Sava, M.; Buzoianu, A.D. CYP4F2 and VKORC1 Polymorphisms Amplify the Risk of Carotid Plaque Formation. Genes 2020, 11, 822. [Google Scholar] [CrossRef]
- Kornej, J.; Börschel, C.S.; Benjamin, E.J.; Schnabel, R.B. Epidemiology of Atrial Fibrillation in the 21st Century: Novel Methods and New Insights. Circ. Res. 2020, 127, 4–20. [Google Scholar] [CrossRef]
- Brodehl, A.; Milting, H.; Gerull, B. Special Issue “Cardiovascular Genetics”. Genes 2021, 12, 479. [Google Scholar] [CrossRef]
- Wilde, A.A.M.; Nannenberg, E.; van der Werf, C. Cardiogenetics, 25 years a growing subspecialism. NethHeartJ. 2020, 28 (Suppl S1), 39–43. [Google Scholar] [CrossRef]
- Maskari, R.A.; McEniery, C.M.; Cleary, S.E.; Li, Y.; Siew, K.; Figg, N.L.; Khir, A.W.; Cockcroft, J.R.; Wilkinson, I.B.; O’Shaughnessy, K.M. The matrix proteins aggrecan and fibulin-1 play a key role in determining aortic stiffness. Sci. Rep. 2018, 8, 8550. [Google Scholar] [CrossRef] [Green Version]
- Inozemtseva, A.A.; Usoltseva, E.N. The role of single nucleotide polymorphisms of lipid disorders in predicting the development and assessing the severity of myocardial infarction. Complex Probl. Cardiovasc. Dis. 2015, 1, 51–60. Available online: https://cyberleninka.ru/article/n/rol-odnonukleotidnyh-polimorfizmov-lipidnyh-narusheniy-v-prognozirovanii-razvitiya-i-otsenke-tyazhesti-techeniya-infarkta-miocarda (accessed on 1 April 2020).
- Erichsen, H.C.; Chanock, S.J. SNPs in cancer research and treatment. Br. J. Cancer 2004, 90, 747–751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belotte, J.; Fletcher, N.M.; Saed, M.G.; Abusamaan, M.S.; Dyson, G.; Diamond, M.P.; Saed, G.M. A Single Nucleotide Polymorphism in Catalase Is Strongly Associated with Ovarian Cancer Survival. PLoS ONE 2015, 10, e0135739. [Google Scholar] [CrossRef] [Green Version]
- Palomino-Morales, R.; Gonzalez-Juanatey, C.; Vazquez-Rodriguez, T.R.; Rodriguez, L.; Miranda-Filloy, J.A.; Fernandez-Gutierrez, B.; Llorca, J.; Martin, J.; Gonzalez-Gay, M.A. A1298C polymorphism in the MTHFR gene predisposes to cardiovascular risk in rheumatoid arthritis. Arthritis Res. Ther. 2010, 12, R71. [Google Scholar] [CrossRef] [Green Version]
- Fawzy, M.S.; Toraih, E.A.; Aly, N.M.; Fakhr-Eldeen, A.; Badran, D.I.; Hussein, M.H. Atherosclerotic and thrombotic genetic and environmental determinants in Egyptian coronary artery disease patients: A pilot study. BMC Cardiovasc. Disord. 2017, 17, 26. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Zhang, X.; Sang, L.; Li, X.T.; Sun, H.Y.; Yang, J.; Yuan, Y. The interaction effects between TLR4 and MMP9 gene polymorphisms contribute to aortic aneurysm risk in a Chinese Han population. BMC Cardiovasc Disord. 2019, 19, 72. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.J.; Stanfill, A.; Pourmotabbed, T. The role of matrix metalloproteinase polymorphisms in ischemic stroke. Int. J. Mol. Sci. 2016, 17, 1323. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Shen, J.X.; Wu, H.T.; Li, X.L.; Wen, X.F.; Du, C.W.; Zhang, G.J. Collagen 1A1 (COL1A1) promotes metastasis of breast cancer and is a potential therapeutic target. Discov Med. 2018, 25, 211–223. [Google Scholar] [PubMed]
- Vafaie, F.; Yin, H.; O’Neil, C.; Nong, Z.; Watson, A.; Arpino, J.M.; Chu, M.W.; Wayne Holdsworth, D.; Gros, R.; Pickering, J.G. Collagenase-resistant collagen promotes mouse aging and vascular cell senescence. Aging Cell 2014, 13, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Ikramov, O.A. Results of the analysis of the association of rs1107946 polymorphism of the col1a1 gene with the risk of myopia development. Re-Health J. 2020, 4, 29–33. [Google Scholar]
- Miyamoto-Mikami, E.; Kumagai, H.; Tanisawa, K.; Taga, Y.; Hirata, K.; Kikuchi, N.; Kamiya, N.; Kawakami, R.; Midorikawa, T.; Kawamura, T.; et al. Female Athletes Genetically Susceptible to Fatigue Fracture Are Resistant to Muscle Injury: Potential Role of COL1A1 Variant. Med. Sci. Sport. Exerc. 2021, 53, 1855–1864. [Google Scholar] [CrossRef] [PubMed]
- Ibragimov, Z.Z.; Boboev, K.T.; Alimov, T.R. Association of polymorphism rs1107946 gene COL1A1 with osteoporosis. J. Biomed. Pract. 2018, 1, 26–32. [Google Scholar] [CrossRef]
- Aboyans, V.; Criqui, M.H.; Abraham, P.; Allison, M.A.; Creager, M.A.; Diehm, C.; Fowkes, F.G.; Hiatt, W.R.; Jönsson, B.; Lacroix, P.; et al. Measurement and interpretation of the ankle-brachial index: A scientific statement from the American Heart Association. Circulation 2012, 126, 2890–2909. [Google Scholar] [CrossRef] [Green Version]
- Tomiyama, H.; Matsumoto, C.; Shiina, K.; Yamashina, A. Brachial-Ankle PWV: Current Status and Future Directions as a Useful Marker in the Management of Cardiovascular Disease and/or Cardiovascular Risk Factors. J. AtherosclerThromb. 2016, 23, 128–146. [Google Scholar] [CrossRef] [Green Version]
- Levine, M.E.; Lu, A.T.; Quach, A.; Chen, B.H.; Assimes, T.L.; Bandinelli, S.; Hou, L.; Baccarelli, A.A.; Stewart, J.D.; Li, Y.; et al. An epigenetic biomarker of aging for lifespan and healthspan. Aging 2018, 18, 573–591. [Google Scholar] [CrossRef] [Green Version]
- Wilcox, R. Introduction to Robust Estimation and Hypothesis Testing, 3rd ed.; Academic Press: New York, NY, USA, 2012. [Google Scholar]
- Agresti, A. Categorical Data Analysis, 2nd ed.; Wiley: New York, NY, USA, 2002. [Google Scholar]
- Heidari, M.M.; Khatami, M.; Hadadzadeh, M.; Kazemi, M.; Mahamed, S.; Malekzadeh, P.; Mirjalili, M. Polymorphisms in NOS3, MTHFR, APOB and TNF-α Genes and Risk of Coronary Atherosclerotic Lesions in Iranian Patients. Res. Cardiovasc. Med. 2015, 5, e29134. [Google Scholar] [CrossRef] [Green Version]
- Gilowska, I.; Kasper, Ł.; Bogacz, K.; Szczegielniak, J.; Szymasek, T.; Kasper, M.; Majorczyk, E. Impact of Matrix Metalloproteinase 9 on COPD Development in Polish Patients: Genetic Polymorphism, Protein Level, and Their Relationship with Lung Function. Biomed. Res Int. 2018, 6417415. [Google Scholar] [CrossRef]
- Feng, X.; Yu, F.; Zhou, X.; Liu, Z.; Liao, D.; Huang, Q.; Li, X.; Jin, X.; Xia, J. MMP9 rs17576 Is Simultaneously Correlated with Symptomatic Intracranial Atherosclerotic Stenosis and White Matter Hyperintensities in Chinese Population. Cerebrovasc Dis. 2021, 50, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Balatsky, A.V.; Samohodskaya, L.M.; Boytsov, S.A.; Tkachuk, V.A. Association of molecular genetic factors with signs of instability of atherosclerotic lesions. Russ. J. Cardiol. 2018, 8, 32–38. [Google Scholar] [CrossRef]
- Fan, D.; Zheng, C.; Wu, W.; Chen, Y.; Chen, D.; Hu, X.; Shen, C.; Chen, M.; Li, R.; Chen, Y. MMP9 SNP and MMP SNP-SNP interactions increase the risk for ischemic stroke in the Han Hakka population. BrainBehav 2022, 12, e2473. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene Product | Polymorphism |
|---|---|---|
| MMP9 | matrix metalloproteinase-9 | rs17576 Gln279Arg |
MMP12 COL1A1 MTHFR ApoB | matrix metalloproteinase-12 pro- α1 chains of type 1 collagen methylenetetrahydrofolate reductase apolipoprotein B | rs652438 Asn357Ser rs1107946 C1997A rs1801131 A1298C rs5742904 R3500Q Arg3500Gln |
| Study Groups | |||
|---|---|---|---|
| Polymorphism | Long-Livers | Descendants of Long-Livers | Control Group |
| rs17576 of the MMP9 gene |  |  |  |
| rs652438 of the MMP12 gene |  |  |  |
| rs1107946 of the COL1A1 gene |  |  |  |
| rs1801131 of the MTHFR gene |  |  |  |
| rs5742904 of the ApoB gene |  |  |  |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kondakova, E.V.; Ilina, V.M.; Ermakova, L.M.; Krivonosov, M.I.; Kuchin, K.V.; Vedunova, M.V. New Genetically Determined Markers of the Functional State of the Cardiovascular System. Genes 2023, 14, 185. https://0-doi-org.brum.beds.ac.uk/10.3390/genes14010185
Kondakova EV, Ilina VM, Ermakova LM, Krivonosov MI, Kuchin KV, Vedunova MV. New Genetically Determined Markers of the Functional State of the Cardiovascular System. Genes. 2023; 14(1):185. https://0-doi-org.brum.beds.ac.uk/10.3390/genes14010185
Chicago/Turabian StyleKondakova, Elena V., Valeria M. Ilina, Lyubov M. Ermakova, Mikhail I. Krivonosov, Kirill V. Kuchin, and Maria V. Vedunova. 2023. "New Genetically Determined Markers of the Functional State of the Cardiovascular System" Genes 14, no. 1: 185. https://0-doi-org.brum.beds.ac.uk/10.3390/genes14010185