
Biodegradation of Microtoxic Phenylpropanoids (Phenylpropanoic Acid and Ibuprofen) by Bacteria and the Relevance for Their Removal from Wastewater Treatment Plants
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Enrichment, Isolation and Growth
2.3. Characterization of the Isolates
2.4. CoA Ligase Activity of the IpfF Protein with Ibuprofen-like Substrates
2.5. Resting Cell Assays
2.6. Horizontal Gene Transfer
2.7. Analytical Techniques
3. Results and Discussion
3.1. Characterization of Ibuprofen Mineralizing Consortia
3.2. Horizontal Transfer of ipf Genes
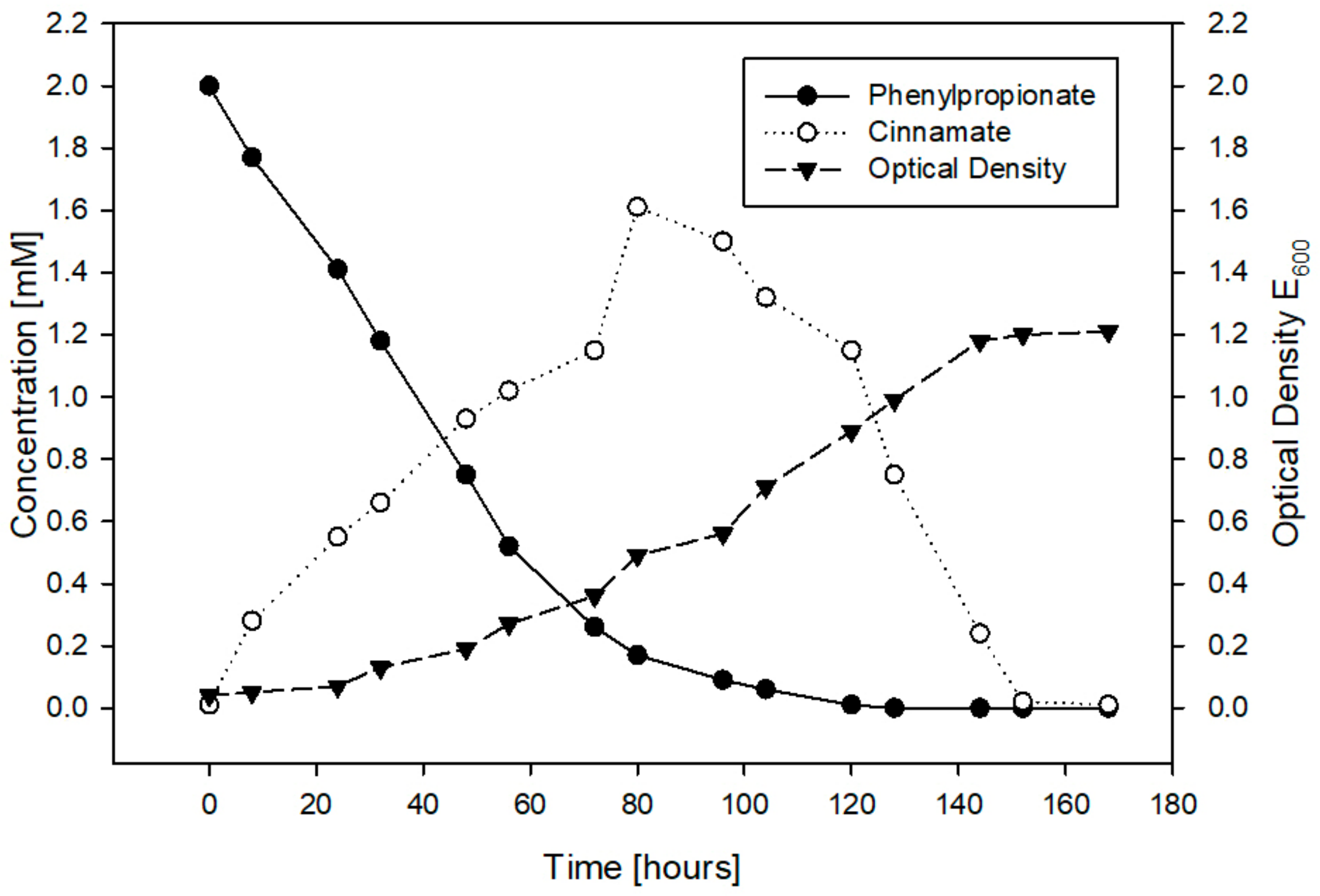
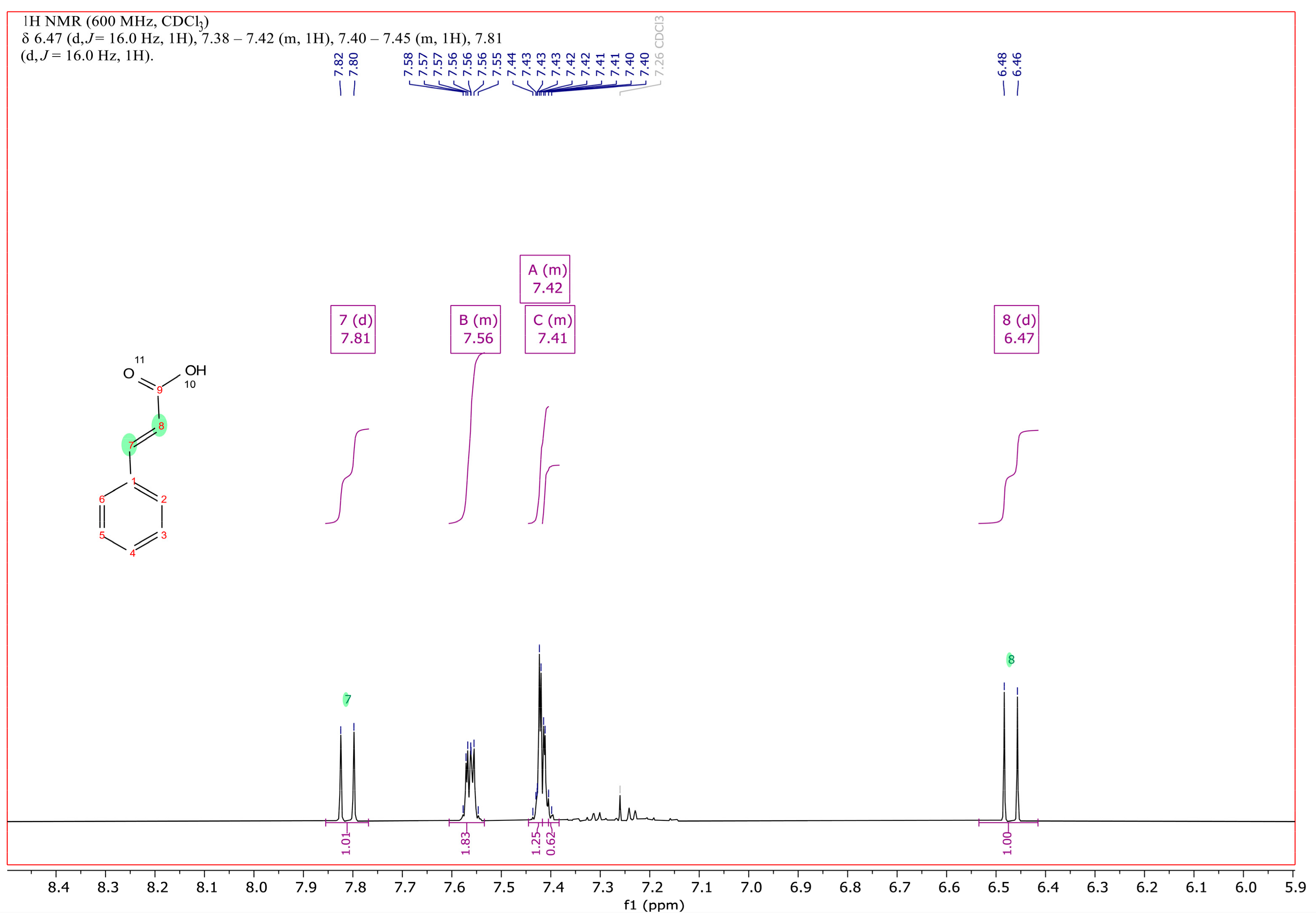
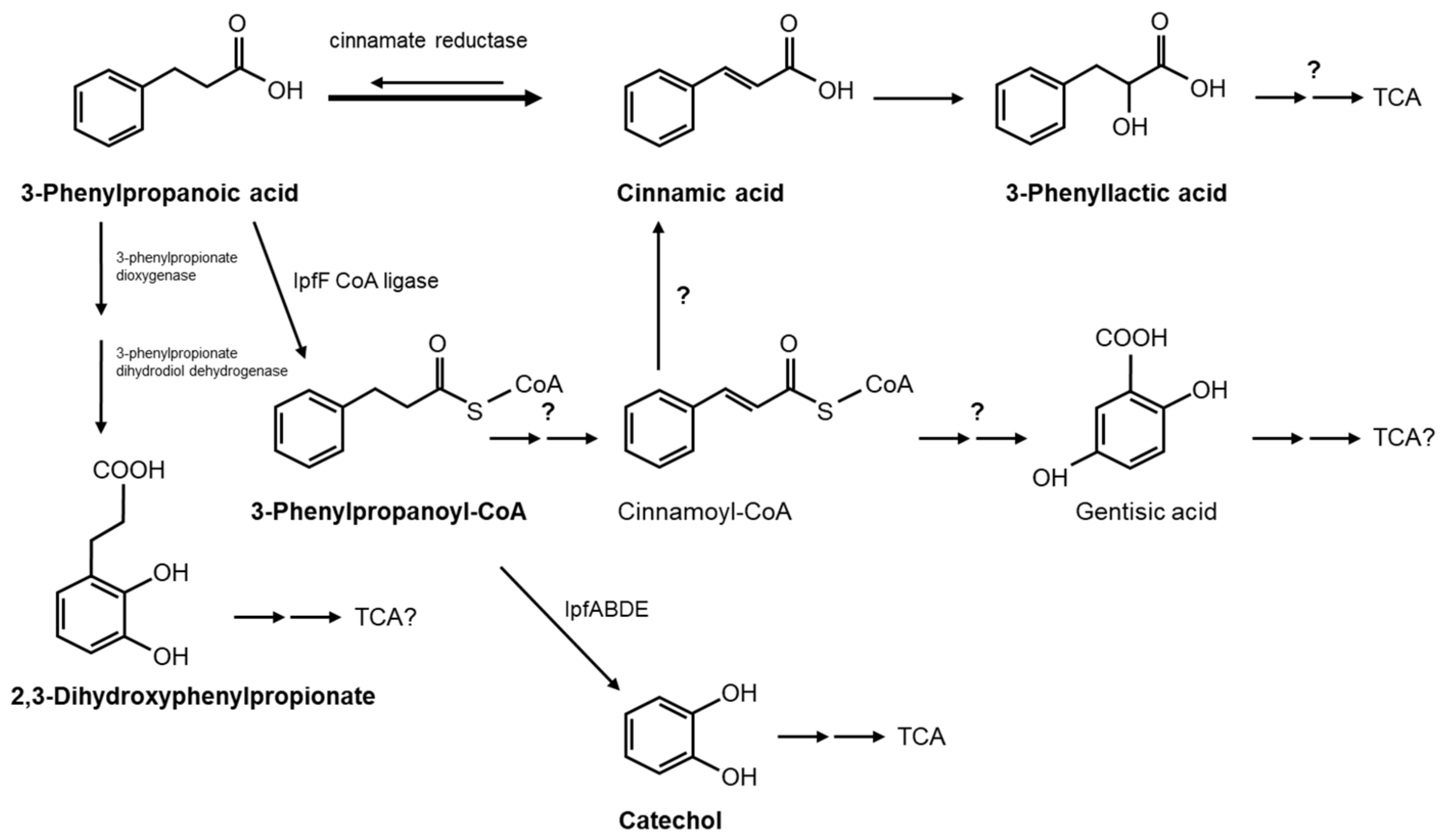
3.3. Catabolism of 3-Phenylpropanoic Acid (3PPA)
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bouissou-Schurtz, C.; Houeto, P.; Guerbet, M.; Bachelot, M.; Casellas, C.; Mauclaire, A.-C.; Panetier, P.; Delval, C.; Masset, D. Ecological risk assessment of the presence of pharmaceutical residues in a French national water survey. Regul. Toxicol. Pharmacol. 2014, 69, 296–303. [Google Scholar] [CrossRef]
- Chopra, S.; Kumar, D. Ibuprofen as an emerging organic contaminant in environment, distribution and remediation. Heliyon 2020, 6, e04087. [Google Scholar] [CrossRef]
- Aguilar-Romero, I.; de la Torre-Zúñiga, J.; Quesada, J.M.; Haïdur, A.; O’Connell, G.; McAmmond, B.M.; van Hamme, J.D.; Romero, E.; Wittich R-Mvan Dillewijn, P. Effluent decontamination by the ibuprofen-mineralizing strain, Sphingopyxis granuli RW412: Metabolic processes. Environ. Pollut. 2021, 274, 116536. [Google Scholar] [CrossRef]
- Murdoch, R.W.; Hay, A.G. Formation of catechols via removal of acid side chains from ibuprofen and related aromatic acids. Appl. Environ. Microbiol. 2005, 71, 6121–6125. [Google Scholar] [CrossRef]
- Murdoch, R.W.; Hay, A.G. Genetic and chemical characterization of ibuprofen degradation by Sphingomonas Ibu-2. Microbiology 2013, 159, 621–632. [Google Scholar] [CrossRef]
- Aulestia, M.; Flores, A.; Mangas, E.L.; Pérz-Pulido, A.J.; Santero, E.; Camacho, E.M. Isolation and genomic characterization of the ibuprofen-degrading bacterium Sphingomonas strain MPO218. Environ. Microbiol. 2021, 23, 267–280. [Google Scholar] [CrossRef]
- Aulestia, M.; Flores, A.; Acosta-Jurado, S.; Santero, E.; Camacho, E.M. Genetic characterization of the ibuprofen-degradative pathway of Rhizorhabdus wittichii MPO218. Appl. Environ. Microbiol. 2022, 88, e0038822. [Google Scholar] [CrossRef]
- Balciunas, E.M.; Kappelmeyer, U.; Harms, H.; Heipieper, H.J. Increasing ibuprofen degradation in constructed wetlands by bioaugmentation with gravel containing biofilms of an ibuprofen-degrading Sphingobium yanoikuyae. Eng. Life Sci. 2020, 20, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Murdoch, R.W.; Hay, A.G. The biotransformation of ibuprofen to trihydroxyibuprofen in activated sludge and by Variovorax Ibu-1. Biodegradation 2015, 26, 105–113. [Google Scholar] [CrossRef]
- Salgado, R.; Brito, D.; Noronha, J.P.; Almeida, B.; Bronze, M.R.; Oehmen, A.; Carvalho, G.; Barreto-Crespo, M.T. Metabolite identification of ibuprofen biodegradation by Patulibacter medicamentivorans under aerobic conditions. Environ. Technol. 2020, 41, 450–465. [Google Scholar] [CrossRef]
- Chopra, S.; Kumar, D. Characteristics and growth kinetics of biomass of Citrobacter freundii strains PYI-2 and Citrobacter portucalensis strain YPI-+2 during the biodegradation of ibuprofen. Int. Microbiol. 2022, 25, 615–628. [Google Scholar] [CrossRef]
- Marchlewicz, A.; Guzik, U.; Smulek, W.; Wojcieszynska, D. Exploring the degradation of ibuprofen by Bacillus thuringiensis B1 (2015b): The new pathway and factors affecting degradation. Molecules 2017, 22, 1676. [Google Scholar] [CrossRef]
- Chen, Y.; Rosazza, J.P. Microbial transformation of ibuprofen by a Nocardia species. Appl. Environ. Microbiol. 1994, 60, 1292–1296. [Google Scholar] [CrossRef]
- Sharma, K.; Kaushik, G.; Thotakura, N.; Raza, K.; Sharma, N.; Nimesh, S. Fate of ibuprofen under optimized batch biodegradation experiments using Micrococcus yunnanensis isolated from pharmaceutical sludge. Int. J. Environ. Sci. Technol. 2019, 16, 8315–8328. [Google Scholar] [CrossRef]
- Lu, Z.; Sun, W.; Li, C.; Ao, X.; Yang, C.; Li, S. Bioremoval of non-steroidal anti-inflammatory drugs by Pseudoxanthomonas sp. DIN-3 isolated from biological activated carbon process. Water Res. 2019, 161, 459–472. [Google Scholar] [CrossRef]
- Ferrando-Climent, L.; Collado, N.; Buttiglieri, G.; Gros, M.; Rodriguez-Roda, I.; Rodriguez-Mozaz, S.; Barcelò, D. Comprehensive study of ibuprofen and its metabolites in activated sludge batch experiments and aquatic environment. Sci. Total Environ. 2012, 438, 404–413. [Google Scholar] [CrossRef]
- Girardi, C.; Nowak, K.M.; Carranza-Diaz, O.; Lewkow, B.; Miltner, A.; Gehre, M.; Schäffer, A.; Kästner, M. Microbial degradation of the pharmaceutical ibuprofen and the herbicide 2,4-D in water and soil—Use of limits of data obtained from aqueous systems for predicting their fate in soil. Sci. Total Environ. 2013, 444, 32–42. [Google Scholar] [CrossRef]
- Nowak, K.M.; Girardi, C.; Miltner, A.; Gehre, M.; Schäffer, A.; Kästner, M. Contribution of microorganisms to non-extractable residue formation during biodegradation of ibuprofen in soil. Sci. Total Environ. 2013, 445–446, 377–384. [Google Scholar] [CrossRef]
- Neelam Khatkar, A.; Sharma, K.K. Phenylpropanoids and its derivatives: Biological activities and its role in food, pharmaceutical and cosmetic industries. Crit. Rev. Food Sci. Nutr. 2020, 60, 2655–2675. [Google Scholar] [CrossRef] [PubMed]
- Korneev, S.M. Hydrocinnamic acids: Application and strategy of synthesis. Synthesis 2013, 45, 1000–1015. [Google Scholar] [CrossRef]
- Narayana, K.J.P.; Prabhakar, P.; Vijayalahshmi, M.; Venkateswarlu, Y.; Krishna, P.S.J. Biological activity of phenylpropionic acid isolated from a terrestrial Streptomycetes. Pol. J. Microbiol. 2007, 56, 191–197. [Google Scholar]
- Griel, C.; Hecht CStreuber, S.; Ganss, Y.; Kremp, H. Determination of phenylacetic acid for early detection of of process faults in biogas plants. Chem. Ing. Technol. 2010, 82, 2223–2229. [Google Scholar] [CrossRef]
- Schaider, L.A.; Rodgers, K.M.; Rudel, R.A. Review of organic wastewater compound concentrations and removal in onsite wastewater treatment systems. Environ. Sci. Technol. 2017, 51, 7304–7317. [Google Scholar] [CrossRef]
- Wagner, A.O.; Prem, E.M.; Markt, R.; Kaufmann, R.; Illmer, P. Formation of phenylacetic acid and phenylpropionic acid under different overload conditions during mesophilic and thermophillic anaerobic digestion. Biotechnol. Biofuels 2019, 12, 26. [Google Scholar] [CrossRef]
- Sander, P.; Wittich, R.M.; Fortnagel, P.; Wilkes, H.; Francke, W. Degradation of 1,2,4-trichloro- and 1,2,4,5-tetrachlorobenzene by Pseudomonas strains. Appl. Environ. Microbiol. 1991, 57, 1430–1440. [Google Scholar] [CrossRef]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Bartels, I.; Knackmuss, H.J.; Reineke, W. Suicide inactivation of catechol 2,3-dioxygenase from Pseudomonas putida mt-2 by 3-halocatechols. Appl. Environ. Microbiol. 1984, 47, 500–505. [Google Scholar] [CrossRef]
- Yabuuchi, E.; Yamamoto, H.; Terabubo, S.; Okamura, N.; Naka, T.; Fujiwara, N.; Kobayashi, K.; Kosako, Y.; Hiraishi, A. Proposal of Sphingomonas wittichii sp. nov. for strain RW1T, known as a dibenzo-p-dioxin metabolizer. Int. J. Syst. Evol. Microbiol. 2001, 51, 281–292. [Google Scholar] [CrossRef]
- Nelson, K.E.; Weinel, C.; Paulsen, I.T.; Dodson, R.J.; Hilbert, H.; Martins dos Santos, V.A.; Fouts, D.E.; Gill, S.R.; Pop, M.; Holmes, M.; et al. Complete genome sequence and comparative analysis of the metabolically versatile Pseudomonas putida KT2440. Environ. Microbiol. 2002, 4, 799–808. [Google Scholar] [CrossRef]
- Choi, K.H.; Kumar, A.; Schweizer, H.P. A 10-min method for preparation of highly electrocompetent Pseudomonas aeruginosa cells: Application for DNA fragment transfer between chromosomes and plasmid transformation. J. Microbiol. Methods 2006, 64, 391–397. [Google Scholar] [CrossRef]
- Żur, J.; Piński, A.; Marchlewicz, A.; Hupert-Kocurek, K.; Wojcieszyńska, D.; Guzik, U. Organic micropollutants paracetamol and ibuprofen—Toxicity, biodegradation, and genetic background of their utilization by bacteria. Environ. Sci. Pollut. Res. 2018, 25, 21498–21524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juhas, M.; van der Meer, J.R.; Gaillard, M.; Harding, R.M.; Hood, D.W.; Crook, D.W. Genomic islands: Tools of bacterial horizontal gene transfer and evolution. FEMS Microbiol. Rev. 2009, 33, 376–393. [Google Scholar] [CrossRef] [PubMed]
- Stolz, A. degradative plasmids from sphingomonads. FEMS Microbiol. Lett. 2013, 350, 9–19. [Google Scholar] [CrossRef]
- Burlingame, R.; Chapman, P.J. Catabolism of phenylpropionic acid and its 3-hydroxy derivative by Escherichia coli. J. Bacteriol. 1983, 155, 113–121. [Google Scholar] [CrossRef]
- Díaz, E.; Ferrández, A.; García, J.L. Characterization of the hca cluster encoding the dioxygenolytic pathway for initial catabolism of 3-phenylpropionic acid in Escherichia coli K-12. J. Bacteriol. 1998, 180, 2915–2923. [Google Scholar] [CrossRef] [PubMed]
- Teufel, R.; Friedrich, T.; Fuchs, G. An oxygenase that forms and deoxygenates toxic epoxide. Nature 2012, 483, 359–362. [Google Scholar] [CrossRef]
- Moreno-Ruiz, E.; Hernáez, M.J.; Martínez-Pérez, O.; Santero, E. Identification and functional characterization of Sphingomonas macrogolitabida strain TFA genes involved in the first two steps of the tetralin catabolic pathway. J. Bacteriol. 2003, 185, 2026–2030. [Google Scholar] [CrossRef]
- Pinyakong, O.; Habe, H.; Omori, T. The unique aromatic catabolic genes in sphingomonads degrading polycyclic aromatic hydrocarbons (PAHs). J. Gen. Appl. Microbiol. 2003, 49, 1–19. [Google Scholar] [CrossRef]
- Lal, R.; Dogra, C.; Malhotra, S.; Sharma, P.; Pal, R. Diversity, distribution and divergence of lin genes in hexachlorocyclohexane- degrading sphingomonads. Trends Biotechnol. 2006, 24, 121–130. [Google Scholar] [CrossRef]
- Mordaka, P.M.; Hall, S.J.; Minton, N.; Stephens, G. Recombinant expression and characterization of the oxygen-sensitive 2-enoate reductase from Clostridium sporogenes. Microbiology 2018, 164, 122–132. [Google Scholar] [CrossRef]
- Fu, W.; Oriel, P. Degradation of 3-phenylpropionic acid by Haloferax sp. D1227. Extremophiles 1999, 3, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, S.; Sova, M.; Ergün, S. Antimicrobial activity of trans-cinnamic acid and commonly used antibiotics against important fish pathogens and nonpathogenic isolates. J. Appl. Microbiol. 2018, 125, 1714–1727. [Google Scholar] [CrossRef] [PubMed]
- Mu, W.; Yu, S.; Zhu, L.; Zhang, T.; Jiang, B. Recent research on 3-phenyllactic acid, a broad-spectrum antimicrobial compound. Appl. Microbiol. Biotechnol. 2012, 95, 1155–1163. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Substrate | Product | Estimated Concentration CoA Product (mM) 1 |
|---|---|---|
| Ibuprofen | ibuprofenyl-CoA | 0.44 |
| Diclofenac | diclofenacyl-CoA | 0.04 |
| Ibufenac | ibufenacyl-CoA | 0.41 |
| Naproxen | naproxenyl-CoA | 0.12 |
| Ketoprofen | ketoprofenyl-CoA | 0.00 |
| Benzoic acid | benzoyl-CoA | 0.00 |
| Phenylacetic acid | kphenylacetyl-CoA | 0.35 |
| 2-Phenylpropanoic acid | 2-phenylpropanoyl-CoA | 0.27 * |
| 3-Phenylpropanoic acid (3PPA) | 3-phenylpropanoyl-CoA | 0.28 * |
| m-Tolylacetic acid | m-tolylacetyl-CoA | 0.42 * |
| p-Tolylacetic acid | p-tolylacetyl-CoA | 0.35 * |
| α,4-Dimethylphenylacetic acid | α,4-dimethylphenylacetyl-CoA | 0.42 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wittich, R.-M.; Haïdour, A.; Aguilar-Romero, I.; de la Torre-Zúñiga, J.; van Dillewijn, P. Biodegradation of Microtoxic Phenylpropanoids (Phenylpropanoic Acid and Ibuprofen) by Bacteria and the Relevance for Their Removal from Wastewater Treatment Plants. Genes 2023, 14, 442. https://0-doi-org.brum.beds.ac.uk/10.3390/genes14020442
Wittich R-M, Haïdour A, Aguilar-Romero I, de la Torre-Zúñiga J, van Dillewijn P. Biodegradation of Microtoxic Phenylpropanoids (Phenylpropanoic Acid and Ibuprofen) by Bacteria and the Relevance for Their Removal from Wastewater Treatment Plants. Genes. 2023; 14(2):442. https://0-doi-org.brum.beds.ac.uk/10.3390/genes14020442
Chicago/Turabian StyleWittich, Regina-Michaela, Ali Haïdour, Inés Aguilar-Romero, Jesús de la Torre-Zúñiga, and Pieter van Dillewijn. 2023. "Biodegradation of Microtoxic Phenylpropanoids (Phenylpropanoic Acid and Ibuprofen) by Bacteria and the Relevance for Their Removal from Wastewater Treatment Plants" Genes 14, no. 2: 442. https://0-doi-org.brum.beds.ac.uk/10.3390/genes14020442