OprD Repression upon Metal Treatment Requires the RNA Chaperone Hfq in Pseudomonas aeruginosa


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Growth Conditions and Minimum Inhibitory Concentration (MIC) Determination
2.2. DNA Manipulations
2.3. Western Blot Analysis
2.4. RNA Extraction and Reverse Transcription
2.5. Quantitative RT-PCR
2.6. Semi-Quantitative RT-PCR
2.7. Chromosome Immunoprecipitation
3. Results
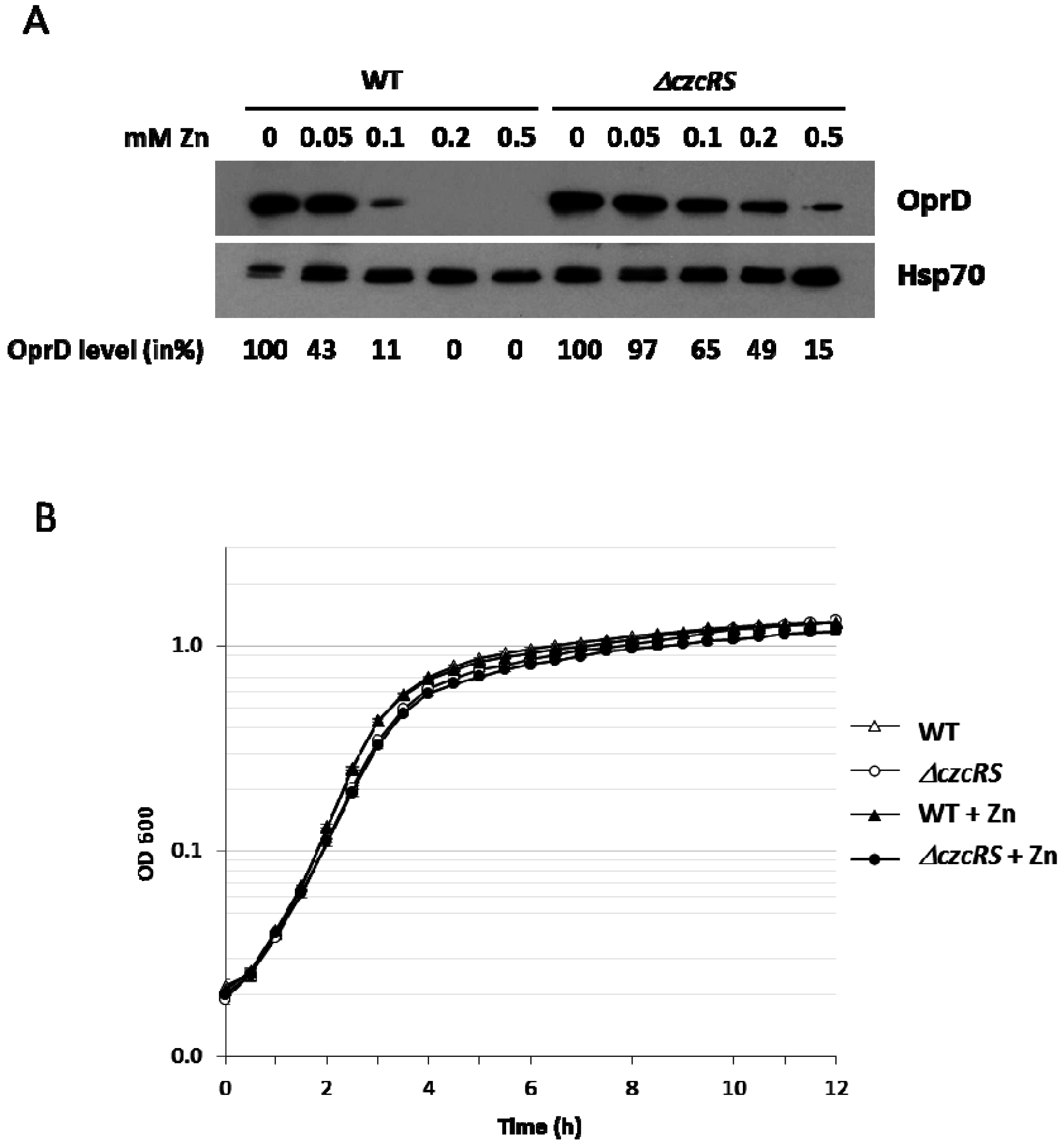
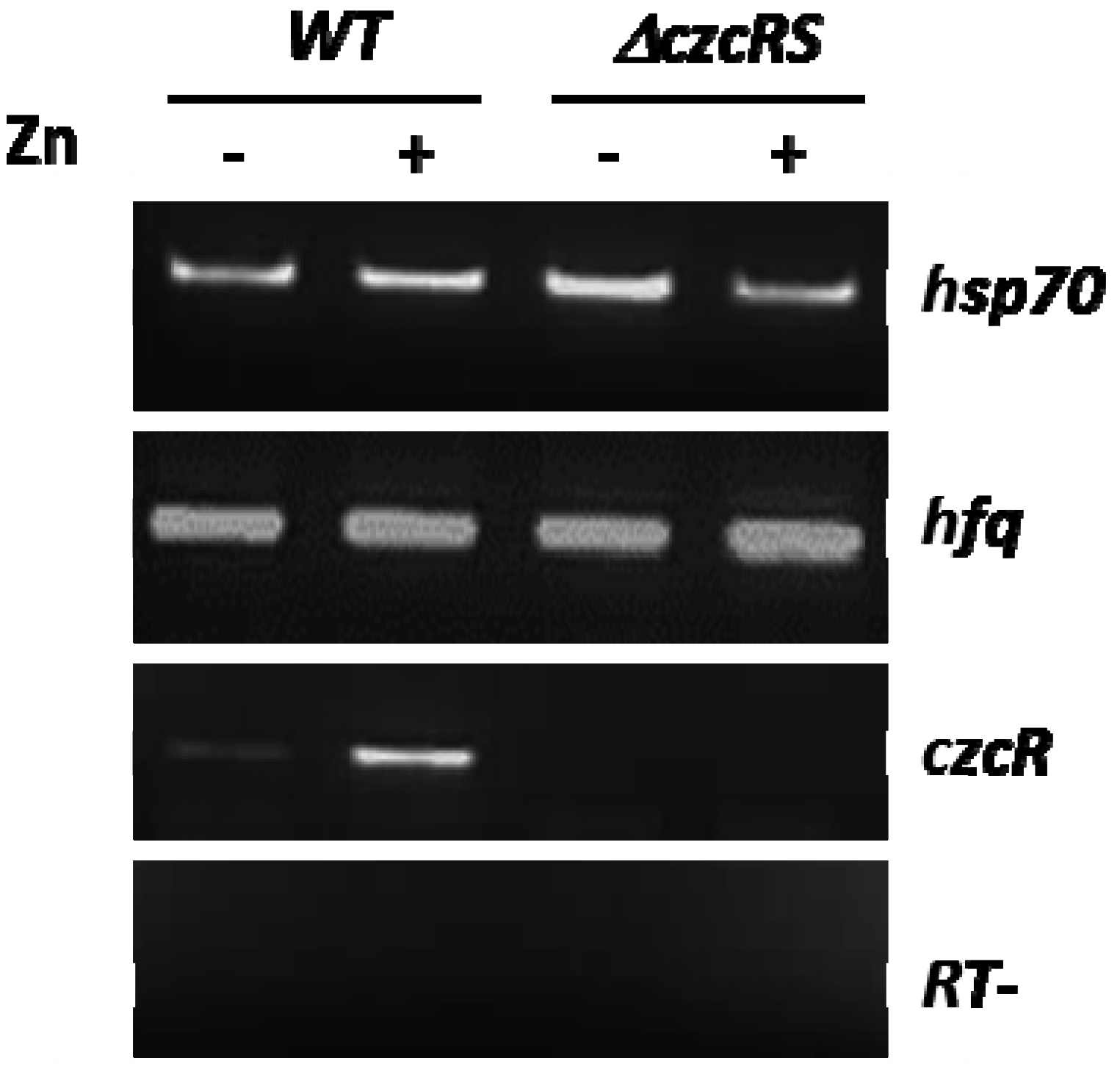
3.1. A CzcR-Independent Mechanism Represses the Production of the OprD Porin in Presence of Zn
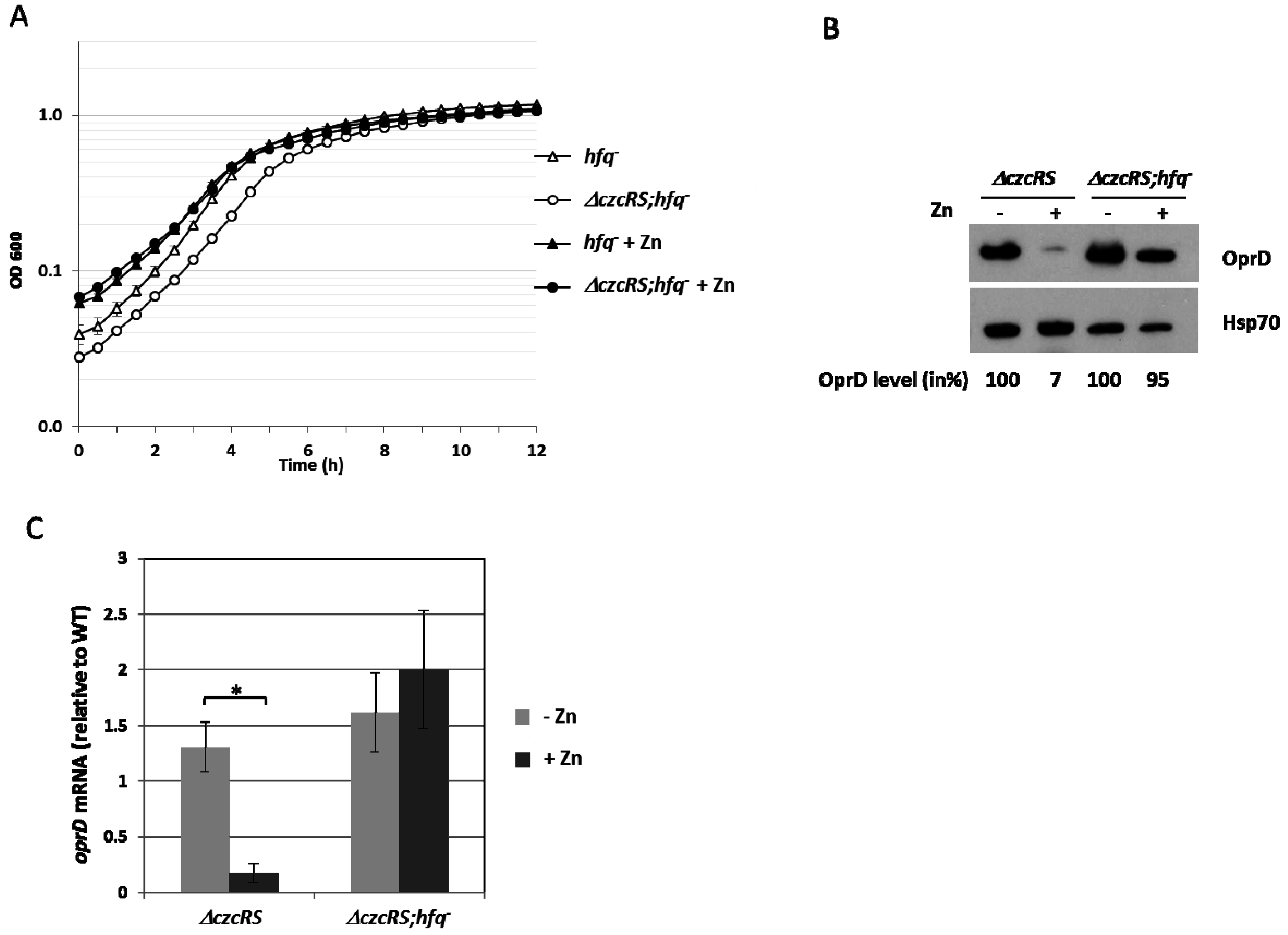
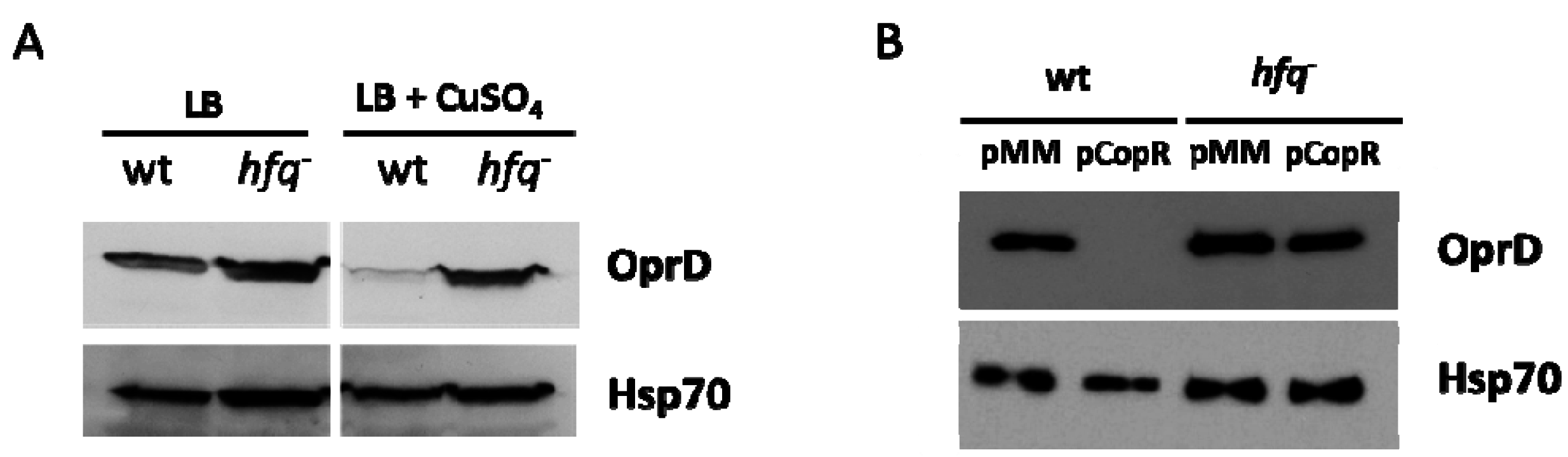
3.2. Hfq is Involved in the Repression of OprD in the Presence of Zn
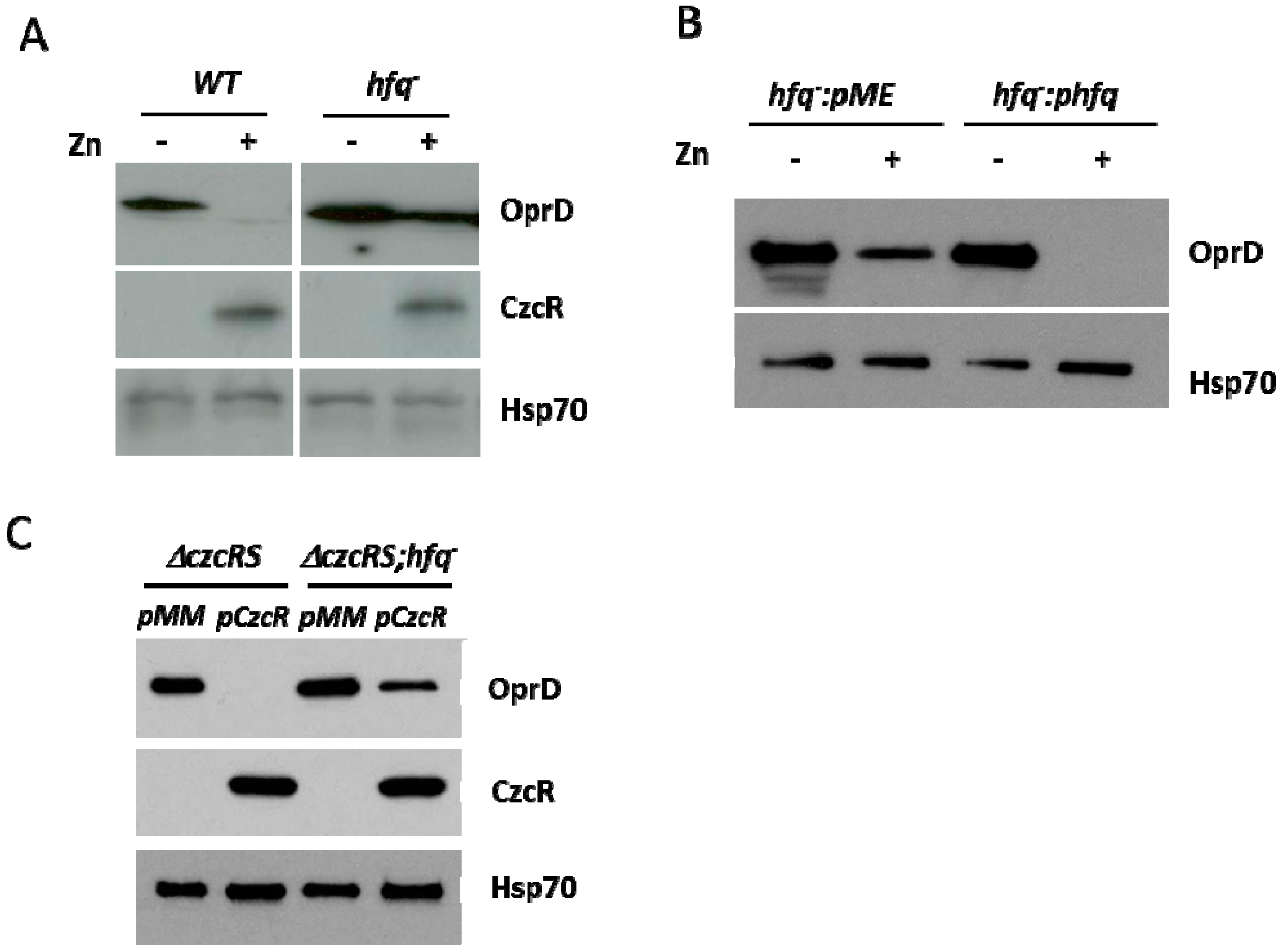
3.3. Hfq is also Involved in the Negative Regulation of OprD Mediated by CzcR
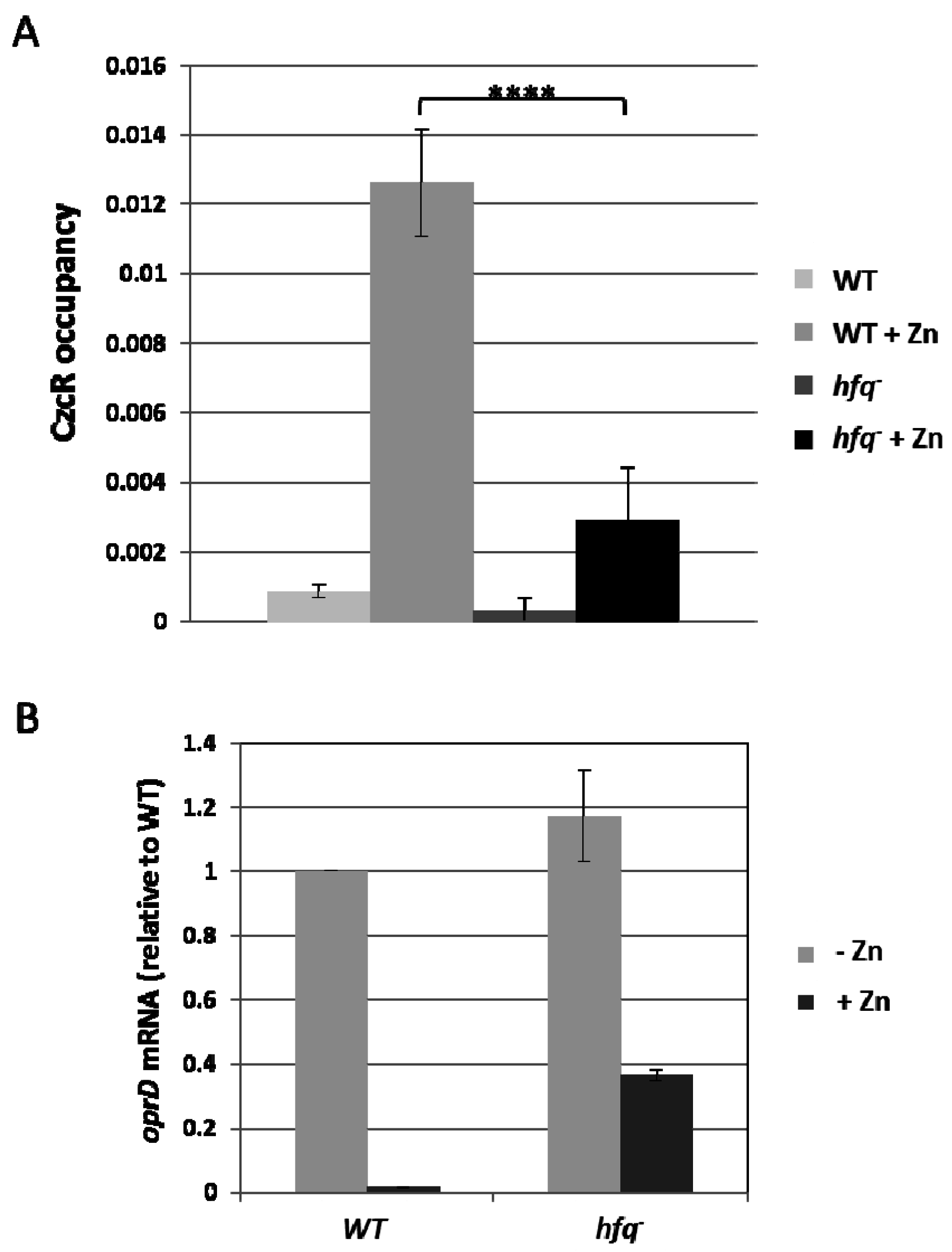
3.4. Hfq is Necessary for Localization of CzcR to the oprD Promoter
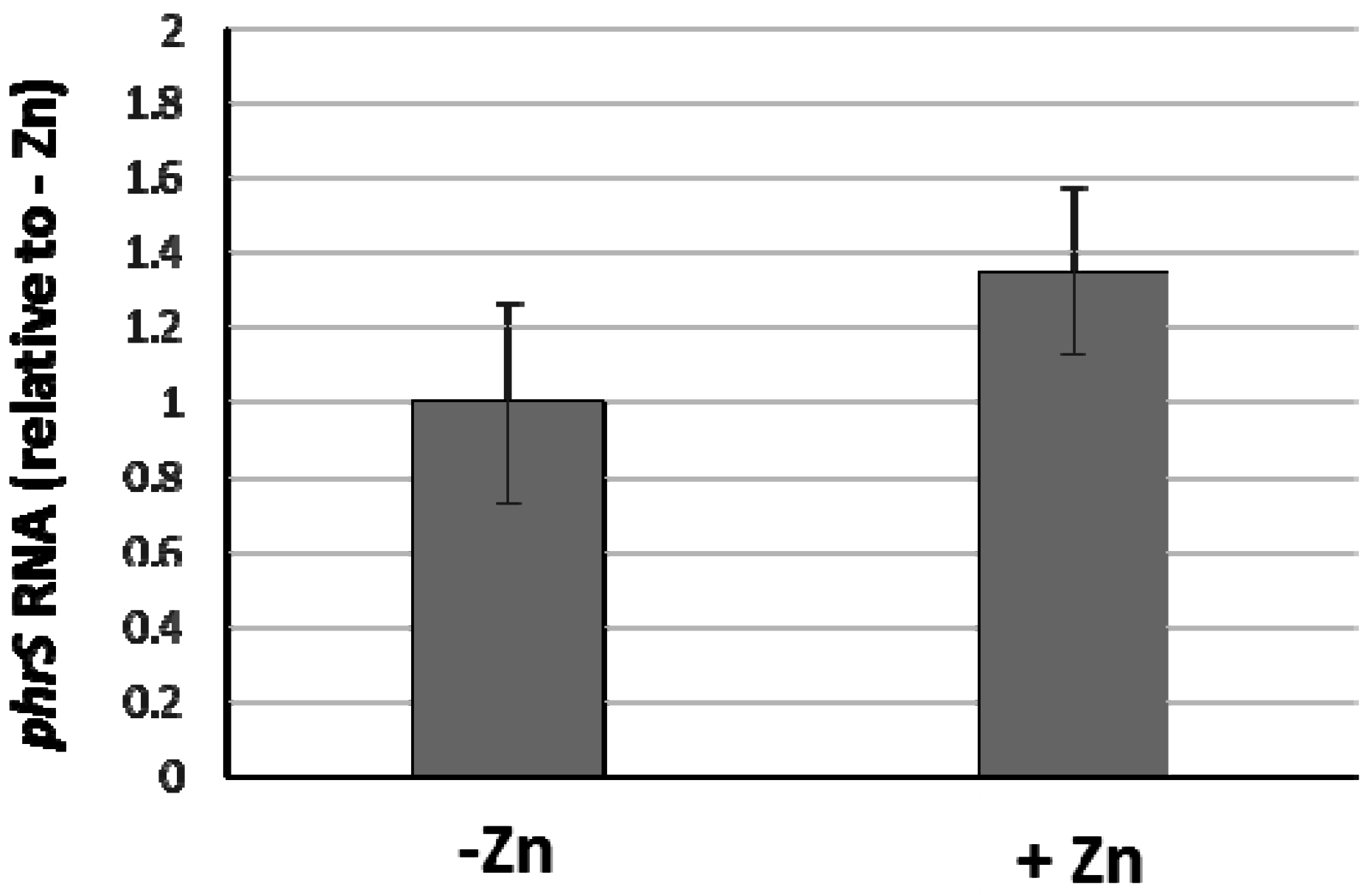
3.5. Is Hfq also Involved in the Repression Mediated by other Transcriptional Regulators?
3.6. Involvement of the Hfq Protein in Carbapenem Resistance
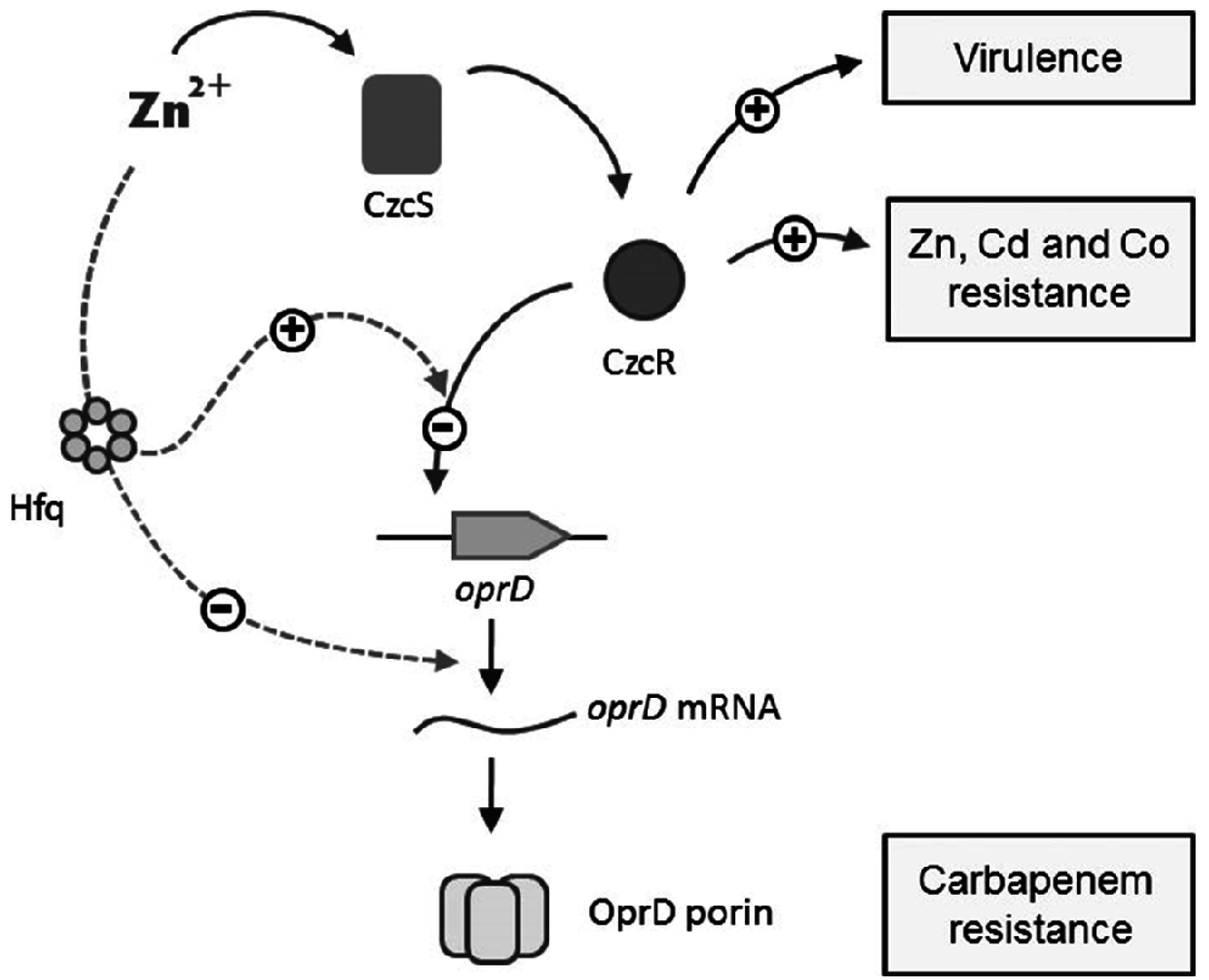
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amplicon | Primer | Sequence (5′ to 3′) | Position | Length |
|---|---|---|---|---|
| Coding region | ||||
| oprD | oprD1 | ATCTACCGCACAAACGATGAAGG | 772 | 156 |
| oprD2 | GCCGAAGCCGATATAATCAAACG | 927 | ||
| oprF | oprF1 | GGTTACTTCCTGACCGACGA | 172 | 664 |
| oprF2 | TCGGTGTTGATGTTGGTGAT | 836 | ||
| hfq | 263 | GTCAAAAGGGCATTCGCTAC | 3 | 176 |
| 331 | AGATCGCGTGCTTGTAAACC | 178 | ||
| hsp70 | 406 | CCACACCGTGATCGTCTATG | 561 | 185 |
| 407 | CGCTTTCCTTCTTGAACTCG | 745 | ||
| czcR | czcR1 | GTCATCACCCGGACGCAGATCAT | 502 | 153 |
| czcR2 | GTAGCCGACGCCGCGAATGGTAT | 654 | ||
| phrS | 449 | CAACTGGAGGCCATCAACAT | 21 | 160 |
| 450 | CCTTGCGTGCTCTGTGTATC | 180 | ||
| Promoter | ||||
| poprD | poprD R | CGCAGATGCGACATGCGTCA | −178 | 101 |
| poprD L | GGCGCTCCACTTCATCACTT | −77 | ||
| Primers used for DNA cloning | ||||
| Hfq:6His | 609 | GCCGAATTCGCGAAGCTGGAGCCATCTAC | −200 | 449 |
| 667 | GCCGGATCCTCAATGATGATGATGAT GATGAGCGTTGCCCGGCTCGG | 249 | ||
| Hfq insertion verification | 314 | TGCCGTATACTGTCGCTCAG | PA4945 | 1390/3534 * |
| 315 | ATTGAACAGGGTCGATTTGC | PA4944 | ||


References
- Lyczak, J.B.; Cannon, C.L.; Pier, G.B. Establishment of Pseudomonas Aeruginosa infection: Lessons from a versatile opportunist. Microbes Infect. 2000, 2, 1051–1060. [Google Scholar] [CrossRef]
- Poole, K. Pseudomonas Aeruginosa: Resistance to the max. Front. Microbiol. 2011. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.; Mohammad, M.M.; Patel, D.R.; Movileanu, L.; van den Berg, B. Structural insight into oprd substrate specificity. Nat. Struct. Mol. Biol. 2007, 14, 1108–1109. [Google Scholar] [CrossRef] [PubMed]
- Ochs, M.M.; Lu, C.D.; Hancock, R.E.; Abdelal, A.T. Amino acid-mediated induction of the basic amino acid-specific outer membrane porin oprd from Pseudomonas Aeruginosa. J. Bacteriol. 1999, 181, 5426–5432. [Google Scholar] [PubMed]
- Caille, O.; Rossier, C.; Perron, K. A copper-activated two-component system interacts with zinc and imipenem resistance in Pseudomonas Aeruginosa. J. Bacteriol. 2007, 189, 4561–4568. [Google Scholar] [CrossRef] [PubMed]
- Ochs, M.M.; McCusker, M.P.; Bains, M.; Hancock, R.E. Negative regulation of the Pseudomonas Aeruginosa outer membrane porin oprd selective for imipenem and basic amino acids. Antimicrob. Agents Chemother. 1999, 43, 1085–1090. [Google Scholar] [PubMed]
- Perron, K.; Caille, O.; Rossier, C.; Van Delden, C.; Dumas, J.L.; Kohler, T. Czcr-czcs, a two-component system involved in heavy metal and carbapenem resistance in Pseudomonas Aeruginosa. J. Biol. Chem. 2004, 279, 8761–8768. [Google Scholar] [CrossRef] [PubMed]
- Baker-Austin, C.; Wright, M.S.; Stepanauskas, R.; McArthur, J.V. Co-selection of antibiotic and metal resistance. Trends Microbiol. 2006, 14, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Dieppois, G.; Ducret, V.; Caille, O.; Perron, K. The transcriptional regulator czcr modulates antibiotic resistance and quorum sensing in Pseudomonas Aeruginosa. PLoS ONE 2012, 7, e38148. [Google Scholar] [CrossRef] [PubMed]
- Marguerettaz, M.; Dieppois, G.; Que, Y.-A.; Ducret, V.; Zuchuat, S.; Perron, K. Sputum containing zinc enhances carbapenem resistance, biofilm formation and virulence of Pseudomonas Aeruginosa. Microb. Pathog. 2014, 77, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Rodrigue, A.; Quentin, Y.; Lazdunski, A.; Mejean, V.; Foglino, M. Two-component systems in Pseudomonas Aeruginosa: Why so many? Trends Microbiol. 2000, 8, 498–504. [Google Scholar] [CrossRef]
- Chen, S.; Zhang, A.; Blyn, L.B.; Storz, G. MicC, a second small-RNA regulator of Omp protein expression in escherichia coli. J. Bacteriol. 2004, 186, 6689–6697. [Google Scholar] [CrossRef] [PubMed]
- Song, T.; Sabharwal, D.; Wai, S.N. VrrA mediates Hfq-dependent regulation of OmpT synthesis in Vibrio cholerae. J. Mol. Biol. 2010, 400, 682–688. [Google Scholar] [CrossRef] [PubMed]
- Frohlich, K.S.; Papenfort, K.; Berger, A.A.; Vogel, J. A conserved RpoS-dependent small RNA controls the synthesis of major porin OmpD. Nucleic. Acids Res. 2012, 40, 3623–3640. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, V.; Sittka, A.; Tomer, R.; Tedin, K.; Brinkmann, V.; Vogel, J. A small non-coding RNA of the invasion gene island (SPI-1) represses outer membrane protein synthesis from the Salmonella core genome. Mol. Microbiol. 2007, 66, 1174–1191. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, V.; Papenfort, K.; Lucchini, S.; Hinton, J.C.; Vogel, J. Coding sequence targeting by MicC RNA reveals bacterial mrna silencing downstream of translational initiation. Nat. Struct. Mol. Biol. 2009, 16, 840–846. [Google Scholar] [CrossRef] [PubMed]
- Vogel, J.; Luisi, B.F. Hfq and its constellation of RNA. Nat. Rev. Microbiol. 2011, 9, 578–589. [Google Scholar] [CrossRef] [PubMed]
- Repoila, F.; Darfeuille, F. Small regulatory non-coding RNAs in bacteria: Physiology and mechanistic aspects. Biol. Cell 2009, 101, 117–131. [Google Scholar] [CrossRef] [PubMed]
- Mura, C.; Randolph, P.S.; Patterson, J.; Cozen, A.E. Archaeal and eukaryotic homologs of Hfq: A structural and evolutionary perspective on Sm function. RNA Biol. 2013, 10, 636–651. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, M.A.; Pearson, R.F.; Moller, T.; Valentin-Hansen, P.; Brennan, R.G. Structures of the pleiotropic translational regulator Hfq and an Hfq-RNA complex: A bacterial Sm-like protein. EMBO J. 2002, 21, 3546–3556. [Google Scholar] [CrossRef] [PubMed]
- Someya, T.; Baba, S.; Fujimoto, M.; Kawai, G.; Kumasaka, T.; Nakamura, K. Crystal structure of hfq from bacillus subtilis in complex with selex-derived RNA aptamer: Insight into RNA-binding properties of bacterial hfq. Nucleic Acids Res. 2012, 40, 1856–1867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sauter, C.; Basquin, J.; Suck, D. Sm-like proteins in eubacteria: The crystal structure of the Hfq protein from Escherichia coli. Nucleic Acids Res. 2003, 31, 4091–4098. [Google Scholar] [CrossRef] [PubMed]
- Moskaleva, O.; Melnik, B.; Gabdulkhakov, A.; Garber, M.; Nikonov, S.; Stolboushkina, E.; Nikulin, A. The structures of mutant forms of Hfq from Pseudomonas Aeruginosa reveal the importance of the conserved his57 for the protein hexamer organization. Acta Crystallogr. Sect. F 2010, 66, 760–764. [Google Scholar] [CrossRef] [PubMed]
- Nikulin, A.; Stolboushkina, E.; Perederina, A.; Vassilieva, I.; Blaesi, U.; Moll, I.; Kachalova, G.; Yokoyama, S.; Vassylyev, D.; Garber, M.; et al. Structure of Pseudomonas Aeruginosa Hfq protein. Acta Crystallogr. Sect. D 2005, 61, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Sonnleitner, E.; Schuster, M.; Sorger-Domenigg, T.; Greenberg, E.P.; Blasi, U. Hfq-dependent alterations of the transcriptome profile and effects on quorum sensing in Pseudomonas Aeruginosa. Mol. Microbiol. 2006, 59, 1542–1558. [Google Scholar] [CrossRef] [PubMed]
- Sonnleitner, E.; Hagens, S.; Rosenau, F.; Wilhelm, S.; Habel, A.; Jager, K.E.; Blasi, U. Reduced virulence of a hfq mutant of Pseudomonas Aeruginosa o1. Microb. Pathog. 2003, 35, 217–228. [Google Scholar] [CrossRef]
- Lenz, D.H.; Mok, K.C.; Lilley, B.N.; Kulkarni, R.V.; Wingreen, N.S.; Bassler, B.L. The small RNA chaperone hfq and multiple small rnas control quorum sensing in vibrio harveyi and vibrio cholerae. Cell 2004, 118, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Matuschek, E.; Brown, D.F.; Kahlmeter, G. Development of the eucast disk diffusion antimicrobial susceptibility testing method and its implementation in routine microbiology laboratories. Clin. Microbiol. Infect. 2014, 20, O255–O266. [Google Scholar] [CrossRef] [PubMed]
- Furste, J.P.; Pansegrau, W.; Frank, R.; Blocker, H.; Scholz, P.; Bagdasarian, M.; Lanka, E. Molecular cloning of the plasmid RP4 primase region in a multi-host-range tacp expression vector. Gene 1986, 48, 119–131. [Google Scholar] [CrossRef]
- Rist, M.; Kertesz, M.A. Construction of improved plasmid vectors for promoter characterization in Pseudomonas Aeruginosa and other gram-negative bacteria. FEMS Microbiol. Lett. 1998, 169, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2001. [Google Scholar]
- Choi, K.H.; Kumar, A.; Schweizer, H.P. A 10-min method for preparation of highly electrocompetent Pseudomonas Aeruginosa cells: Application for DNA fragment transfer between chromosomes and plasmid transformation. J. Microbiol. Methods 2006, 64, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST©) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002. [Google Scholar] [CrossRef]
- Perron, K.; Comte, R.; van Delden, C. Dksa represses ribosomal gene transcription in Pseudomonas Aeruginosa by interacting with RNA polymerase on ribosomal promoters. Mol. Microbiol. 2005, 56, 1087–1102. [Google Scholar] [CrossRef] [PubMed]
- Vogel, J.; Papenfort, K. Small non-coding RNAs and the bacterial outer membrane. Curr. Opin. Microbiol. 2006, 9, 605–611. [Google Scholar] [CrossRef] [PubMed]
- Gales, A.C.; Jones, R.N.; Sader, H.S. Global assessment of the antimicrobial activity of polymyxin b against 54 731 clinical isolates of gram-negative bacilli: Report from the sentry antimicrobial surveillance programme (2001–2004). Clin. Microbiol. Infect. 2006, 12, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Lister, P.D.; Wolter, D.J.; Hanson, N.D. Antibacterial-resistant Pseudomonas Aeruginosa: Clinical impact and complex regulation of chromosomally encoded resistance mechanisms. Clin. Microbiol. Rev. 2009, 22, 582–610. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Luo, Y.F.; Williams, B.J.; Blackwell, T.S.; Xie, C.M. Structure and function of oprd protein in Pseudomonas Aeruginosa: From antibiotic resistance to novel therapies. Int. J. Med. Microbiol. 2012, 302, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Kohler, T.; Epp, S.F.; Curty, L.K.; Pechere, J.C. Characterization of mext, the regulator of the mexe-mexf-oprn multidrug efflux system of Pseudomonas Aeruginosa. J. Bacteriol. 1999, 181, 6300–6305. [Google Scholar] [PubMed]
- Sonnleitner, E.; Sorger-Domenigg, T.; Madej, M.J.; Findeiss, S.; Hackermuller, J.; Huttenhofer, A.; Stadler, P.F.; Blasi, U.; Moll, I. Detection of small RNAs in Pseudomonas Aeruginosa by rnomics and structure-based bioinformatic tools. Microbiology 2008, 154, 3175–3187. [Google Scholar] [CrossRef] [PubMed]
- Ellis, M.J.; Trussler, R.S.; Haniford, D.B. Hfq binds directly to the ribosome-binding site of IS10 transposase mrna to inhibit translation. Mol. Microbiol. 2015, 96, 633–650. [Google Scholar] [CrossRef] [PubMed]
- Sobrero, P.; Valverde, C. Evidences of autoregulation of hfq expression in sinorhizobium meliloti strain 2011. Arch. Microbiol. 2011, 193, 629–639. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Huang, X.; Li, S.; Huang, J.; Wei, X.; Li, Y.; Xu, Y. The RNA chaperone hfq regulates antibiotic biosynthesis in the rhizobacterium Pseudomonas Aeruginosa M18. J. Bacteriol. 2012, 194, 2443–2457. [Google Scholar] [CrossRef] [PubMed]
- Stock, A.M.; Robinson, V.L.; Goudreau, P.N. Two-component signal transduction. Annu. Rev. Biochem. 2000, 69, 183–215. [Google Scholar] [CrossRef] [PubMed]
- Sobrero, P.; Valverde, C. The bacterial protein Hfq: Much more than a mere RNA-binding factor. Crit. Rev. Microbiol. 2012, 38, 276–299. [Google Scholar] [CrossRef] [PubMed]
- Ohniwa, R.L.; Muchaku, H.; Saito, S.; Wada, C.; Morikawa, K. Atomic force microscopy analysis of the role of major DNA-binding proteins in organization of the nucleoid in Escherichia Coli. PLoS ONE 2013, 8, e72954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Mowafi, S.A.; Alumasa, J.N.; Ades, S.E.; Keiler, K.C. Cell-based assay to identify inhibitors of the hfq-sRNA regulatory pathway. Antimicrob. Agents Chemother. 2014, 58, 5500–5509. [Google Scholar] [CrossRef] [PubMed]






| Strain or Plasmid | Relevant Characteristic(s) | Reference or Source |
|---|---|---|
| Pseudomonas aeruginosa | ||
| Wild type (WT) | PAO1 wild type | WT, laboratory collection |
| ΔczcRS | PAO1 ΔczcRS | [5] |
| hfq− | PAO1 hfq::aadA, Smr, Spr | [26] |
| ΔczcRS, hfq− | PAO1 ΔczcRS, hfq::aadA, Smr, Spr | This study |
| Escherichia coli | ||
| Top10 | F-mcrA Δ(mrr-hsdRMS-mcrBC) Φ80lacZΔM15 Δ lacX74 recA1 araD139 Δ(araleu)7697 galU galK rpsL (StrR) endA1 nupG | Invitrogen |
| Plasmids | ||
| pMMB66EH | Broad host range expressing vector, Cbr | [29] |
| pczcRWT pcopRWT | pMMB66EH derivative containing czcR gene, Cbr pMMB66EH derivative containing copR gene, Cbr | [7] [5] |
| pME4510 pME-Hfq | Multicopy broad host range vector, Gmr pME4510 derivative containing hfq gene under its own promoter, Gmr | [30] This study |
| Strain | Medium Supplemented with: | ||
|---|---|---|---|
| No Metal | Zn | Cu | |
| WT | 1 | 16 | >32 |
| ΔczcRS | 1 | 3 | >32 |
| hfq− | 0.38 | 2 | 4 |
| ΔczcRS;hfq− | 0.25 | 0.38 | 2 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ducret, V.; Gonzalez, M.R.; Scrignari, T.; Perron, K. OprD Repression upon Metal Treatment Requires the RNA Chaperone Hfq in Pseudomonas aeruginosa. Genes 2016, 7, 82. https://0-doi-org.brum.beds.ac.uk/10.3390/genes7100082
Ducret V, Gonzalez MR, Scrignari T, Perron K. OprD Repression upon Metal Treatment Requires the RNA Chaperone Hfq in Pseudomonas aeruginosa. Genes. 2016; 7(10):82. https://0-doi-org.brum.beds.ac.uk/10.3390/genes7100082
Chicago/Turabian StyleDucret, Verena, Manuel R. Gonzalez, Tiziana Scrignari, and Karl Perron. 2016. "OprD Repression upon Metal Treatment Requires the RNA Chaperone Hfq in Pseudomonas aeruginosa" Genes 7, no. 10: 82. https://0-doi-org.brum.beds.ac.uk/10.3390/genes7100082