
Seasonal Variations in the Concentration of Particulate Matter in the Air of Cracow Affect the Magnitude of CD4+ T Cell Subsets Cytokine Production in Patients with Inflammatory and Autoimmune Disorders

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Control Groups
2.2. Preparation of the PM from the Air of Cracow
2.3. Cell Isolation
2.4. Cell Viability Assessment
2.5. Cell Culture and Immunostaining for Intracellular Proteins
2.6. Statistics
3. Results
3.1. Characterization of Patients and Healthy Donors
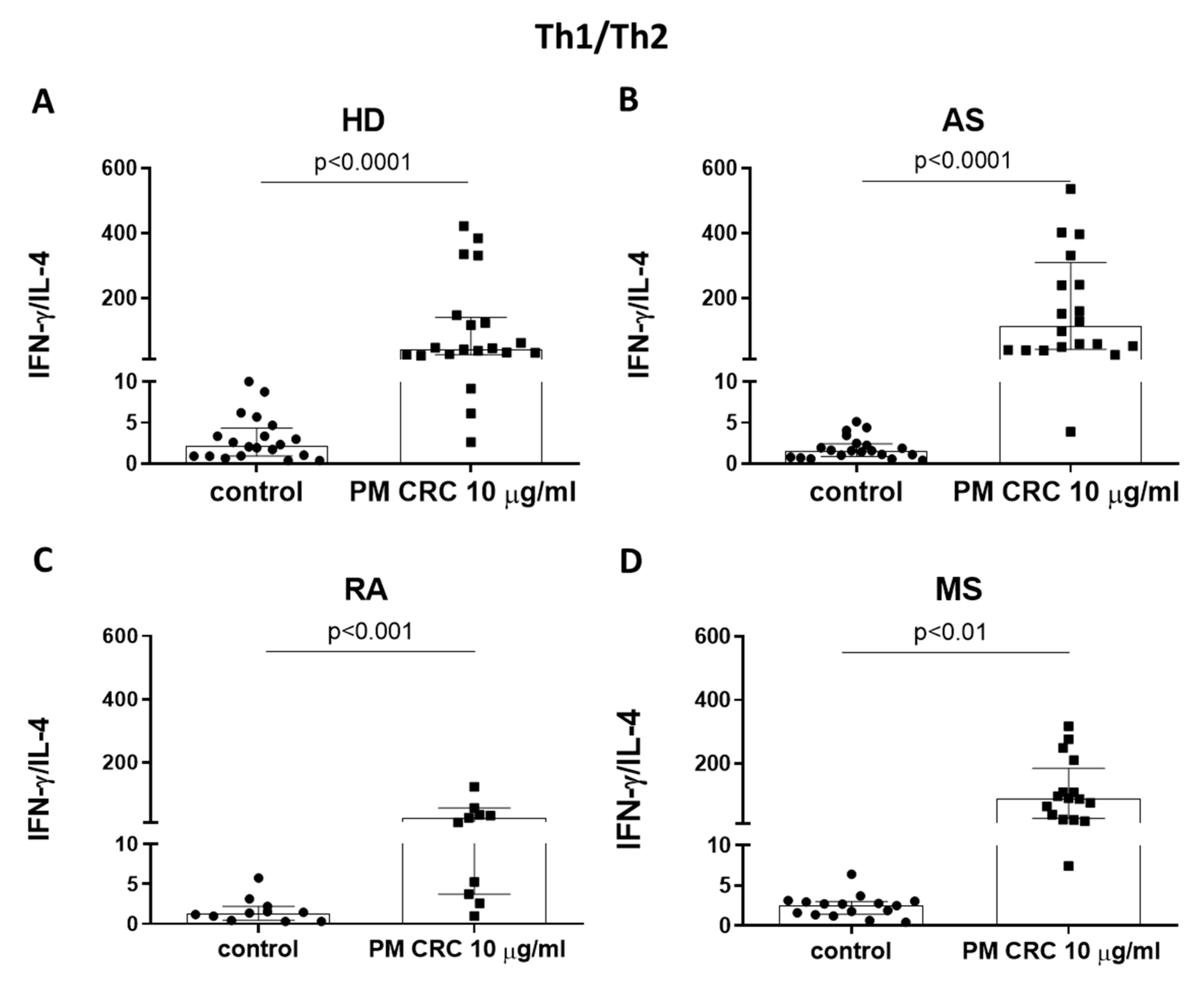
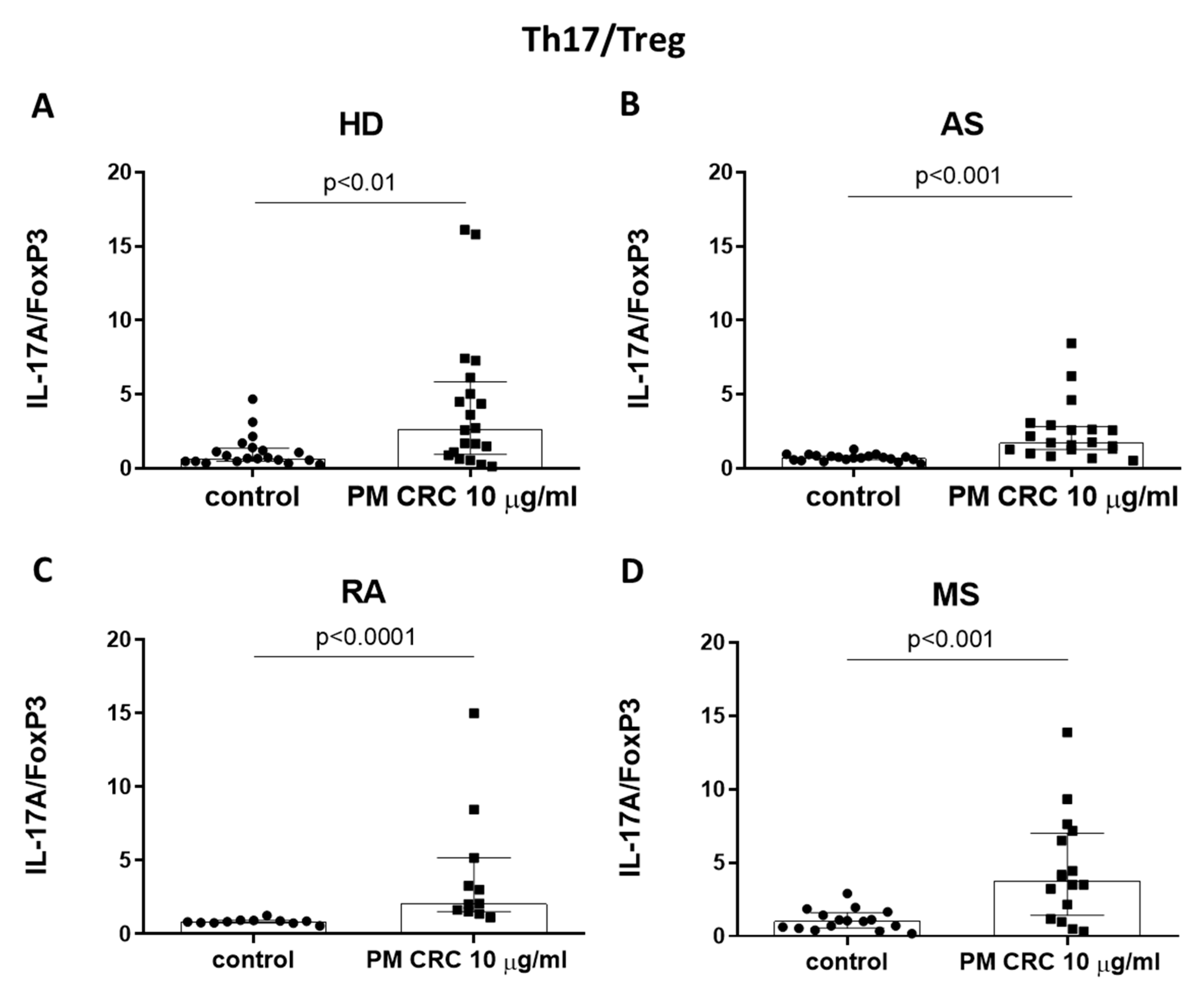
3.2. PM CRC Activate CD4+ T Lymphocytes, Skewing the Balance of Th1/Th2 and Th17/Treg Subsets in Patients and Healthy Donors
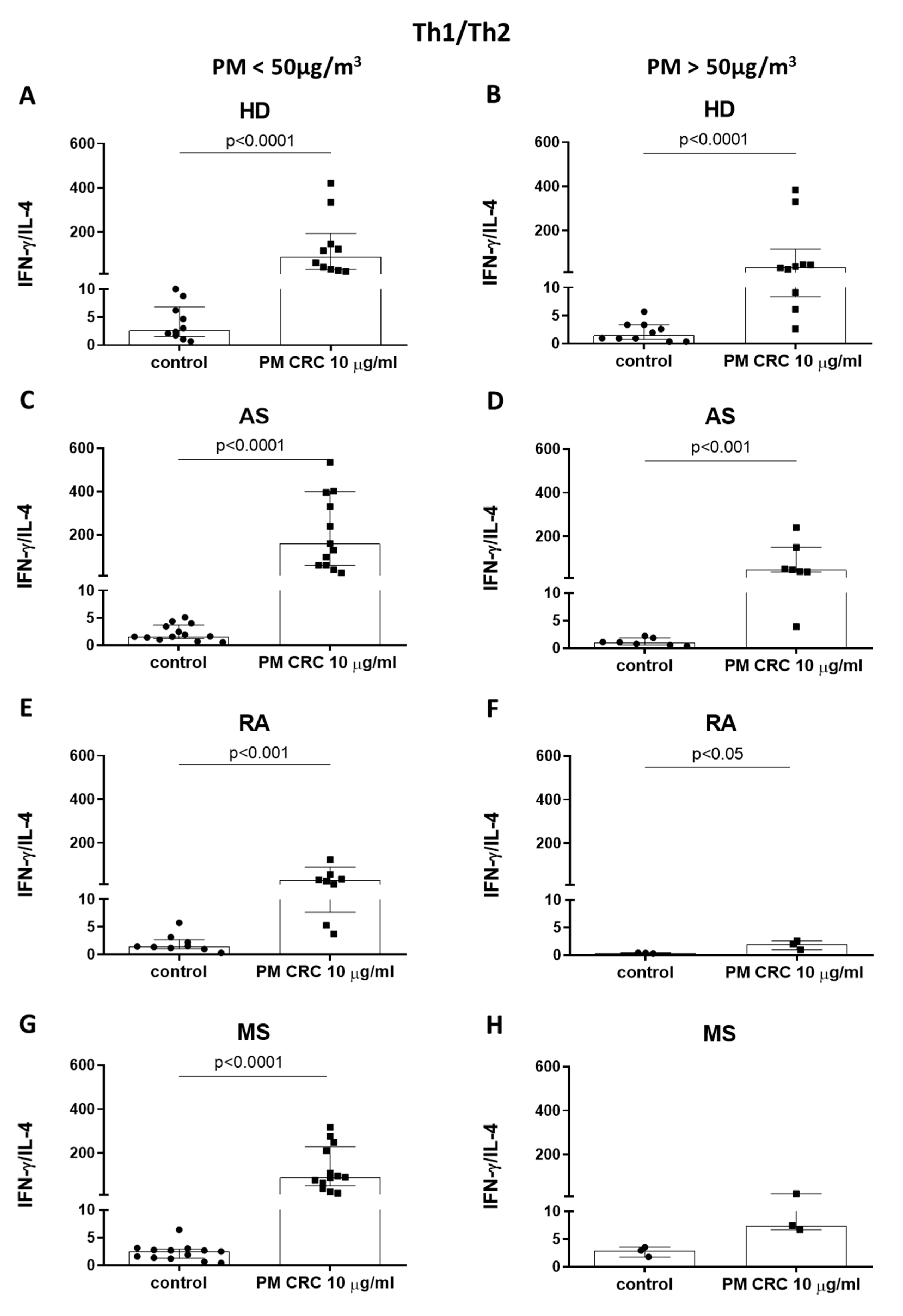
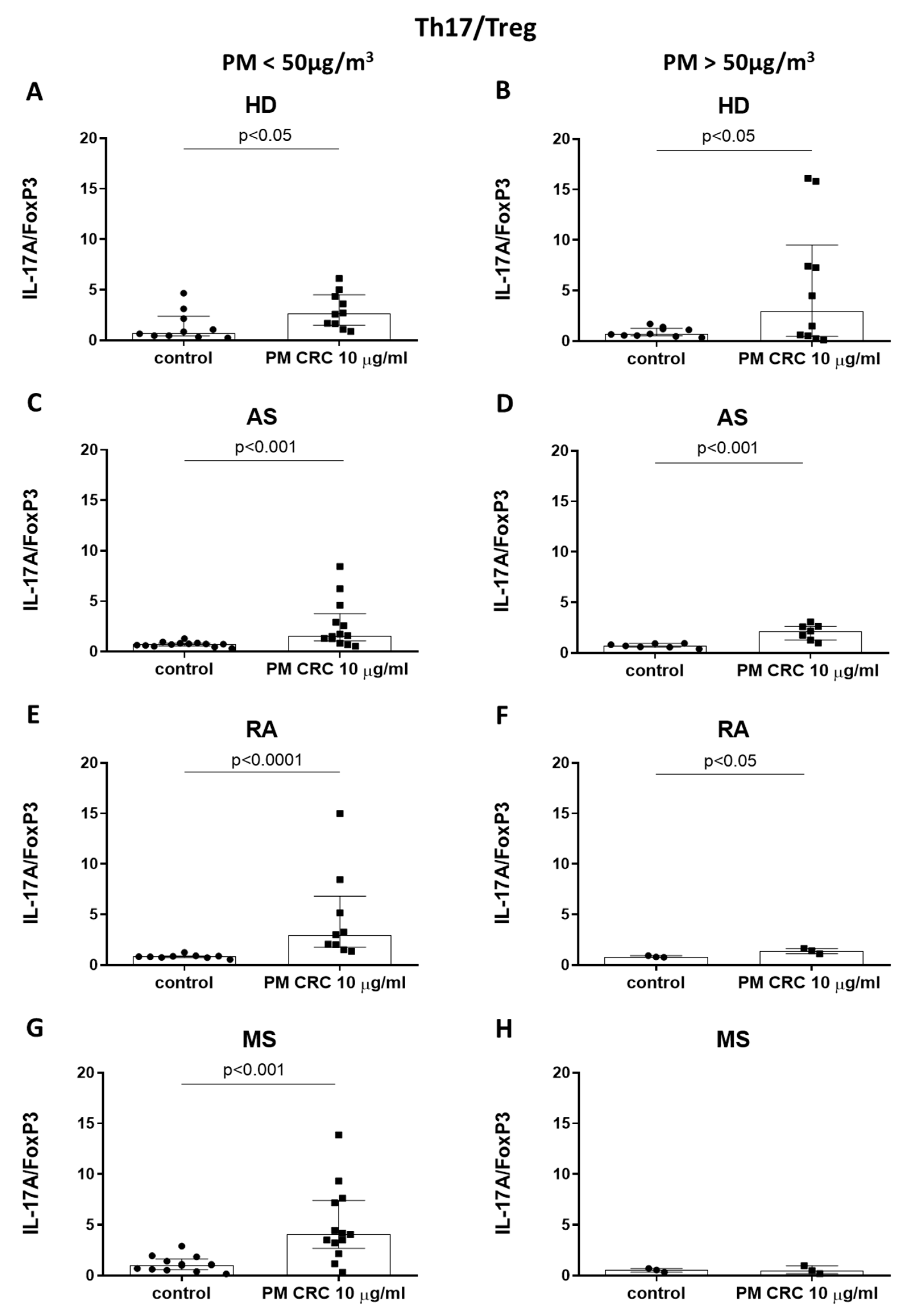
3.3. Polarization of CD4+ T Cells in Patients with Inflammatory or Autoimmune Disorders Depends on the Seasonal Changes in the Concentration of PM in the Air of Cracow
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- The European Parliament; The Council of The European Union. Directive 2008/50/EC of the European Parliament and the Council of 21 May 2008 on ambient air quality and cleaner air for Europe. Off. J. Eur. Union 2008, 152, 1–44. [Google Scholar]
- Polish Minister of the Environment. Regulation of the Minister of the Environment of 24 August 2012 on the levels of certain substances in the air. J. Laws 2012, 1031, 1–9. (In Polish). Available online: https://isap.sejm.gov.pl/isap.nsf/download.xsp/WDU20120001031/O/D20121031.pdf (accessed on 28 February 2022).
- De Brito Beirao, C.A.; Ortiz, G.; De Leeuw, F.; Wiana, M. Air Quality in Europe—2018 Report; European Environment Agency: Copenhagen, Denmark, 2018.
- World Bank Group. In the Spotlight. Air Quality in Poland, What Are the Issues and What Can Be Done? World Bank Group: Washington, DC, USA, 2019. [Google Scholar]
- Khomenko, S.; Cirach, M.; Pereira-Barboza, E.; Mueller, N.; Barrera-Gómez, J.; Rojas-Rueda, D.; de Hoogh, K.; Hoek, G.; Nieuwenhuijsen, M. Premature mortality due to air pollution in European cities: A health impact assessment. Lancet Planet. Health 2021, 5, 121–134. [Google Scholar] [CrossRef]
- Nazar, W.; Plata-Nazar, K. Changes in Air Pollution-Related Behaviour Measured by Google Trends Search Volume Index in Response to Reported Air Quality in Poland. Int. J. Environ. Res. Public Health 2021, 18, 11709. [Google Scholar] [CrossRef] [PubMed]
- Traczyk, P.; Gruszecka-Kosowska, A. The Condition of Air Pollution in Kraków, Poland, in 2005–2020, with Health Risk Assessment. Int. J. Environ. Res. Public Health 2020, 17, 6063. [Google Scholar] [CrossRef] [PubMed]
- Bokwa, A. The climate of the city and air pollution. Aura 2016, 9, 8–13. (In Polish) [Google Scholar]
- Brook, R.D.; Rajagopalan, S.; Pope, C.A.; Brook, J.R.; Bhatnagar, A.; Diez-Roux, A.V.; Holguin, F.; Hong, Y.; Luepker, R.V.; Mittleman, M.A.; et al. American Heart Association Council on Epidemiology and Prevention, Council on the Kidney in Cardiovascular Disease, and Council on Nutrition, Physical Activity, and Metabolism. Particulate matter air pollution and cardiovascular disease: An update to the scientific statement from the American Heart Association. Circulation 2010, 121, 2331–2378. [Google Scholar]
- Al-Kindi, S.G.; Brook, R.D.; Biswal, S.; Rajagopalan, S. Environmental determinants of cardiovascular disease: Lessons learned from air pollution. Nat. Rev. Cardiol. 2020, 17, 656–672. [Google Scholar] [CrossRef]
- Marchini, T.; Zirlik, A.; Wolf, D. Pathogenic Role of Air Pollution Particulate Matter in Cardiometabolic Disease: Evidence from Mice and Humans. Antioxid. Redox Signal. 2020, 33, 263–279. [Google Scholar] [CrossRef]
- Gawda, A.; Majka, G.; Nowak, B.; Marcinkiewicz, J. Air pollution, oxidative stress, and exacerbation of autoimmune diseases. Cent. Eur. J. Immunol. 2017, 42, 305–312. [Google Scholar] [CrossRef]
- Ji, H.; Hershey, G.K.K. Genetic and epigenetic influence on the response to environmental particulate matter. J. Allergy Clin. Immunol. 2013, 129, 33–41. [Google Scholar] [CrossRef] [Green Version]
- Ritz, S.A. Air pollution as a potential contributor to the “epidemic” of autoimmune disease. Med. Hypotheses 2010, 74, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Doria, A.; Zen, M.; Bettio, S.; Gatto, M.; Bassi, N.; Nalotto, L.; Ghirardello, A.; Iaccarino, L.; Punzi, L. Autoinflammation and autoimmunity: Bridging the divide. Autoimmun. Rev. 2012, 12, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Richmond-Bryant, J.; Lu, S.E.; Buckley, B.; Welsh, W.J.; Whitsel, E.A.; Hanna, A.; Yeatts, K.B.; Warren, J.; Herring, A.H.; et al. Cardiovascular outcomes and the physical and chemical properties of metal ions found in particulate matter air pollution: A QICAR study. Environ. Health Perspect. 2013, 121, 558–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, R.O.; Baccarelli, A. Metals and neurotoxicology. J. Nutr. 2007, 137, 2809–2813. [Google Scholar] [CrossRef] [Green Version]
- Sawyer, K.; Mundandhara, S.; Ghio, A.J.; Madden, M.C. The effects of ambient particulate matter on human alveolar macrophage oxidative and inflammatory responses. J. Toxicol. Environ. Health A 2009, 73, 41–57. [Google Scholar] [CrossRef]
- Zelikoff, J.T.; Schermerhorn, K.R.; Fang, K.; Cohen, M.D.; Schlesinger, R.B. A role for associated transition metals in the immunotoxicity of inhaled ambient particulate matter. Environ. Health Perspect. 2002, 110, 871–875. [Google Scholar] [CrossRef] [Green Version]
- Nemmar, A.; Hoet, P.H.M.; Vanquickenborne, B.; Dinsdale, D.; Thomeer, M.; Hoylaerts, M.F.; Vanbilloen, H.; Mortelmans, L.; Nemery, B. Passage of inhaled particles into the blood circulation in humans. Circulation 2002, 105, 411–414. [Google Scholar] [CrossRef] [Green Version]
- Gałuszka, A.; Stec, M.; Węglarczyk, K.; Kluczewska, A.; Siedlar, M.; Baran, J. Transition metal containing particulate matter promotes Th1 and Th17 inflammatory response by monocyte activation in organic and inorganic compounds dependent manner. Int. J. Environ. Res. Public Health 2020, 17, 1227. [Google Scholar] [CrossRef] [Green Version]
- Crane, I.J.; Forrester, J.V. Th1 and Th2 lymphocytes in autoimmune disease. Crit. Rev. Immunol. 2005, 25, 75–102. [Google Scholar] [CrossRef]
- Lee, F.E.; Georas, S.N.; Beck, L.A. IL-17: Important for host defense, autoimmunity, and allergy? J. Investig. Dermatol. 2010, 130, 2540–2542. [Google Scholar]
- Araujo, J.A.; Barajas, B.; Kleinman, M.; Wang, X.; Bennett, B.J.; Gong, K.W.; Navab, M.; Harkema, J.; Sioutas, C.; Lusis, A.J.; et al. Ambient particulate pollutants in the ultrafine range promote early atherosclerosis and systemic oxidative stress. Circ. Res. 2008, 102, 589–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oikonen, M.; Laaksonen, M.; Laippala, P.; Oksaranta, O.; Lilius, E.M.; Lindgren, S.; Rantio-Lehtimäki, A.; Anttinen, A.; Koski, K.; Erälinna, J.P. Ambient air quality and occurrence of multiple sclerosis relapse. Neuroepidemiology 2003, 22, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Hou, T.; Liao, J.; Zhang, C.; Sun, C.; Li, X.; Wang, G. Elevated expression of miR-146, miR-139 and miR-340 involved in regulating Th1/Th2 balance with acute exposure of fine particulate matter in mice. Int. Immunopharmacol. 2018, 54, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Dong, T.; Jiang, X.; Wang, C.; Zhang, Y.; Li, Y.; Zheng, G.; Li, X.; Bai, J.; Li, H. Chronic and low-level particulate matter exposure can sustainably mediate lung damage and alter CD4 T cells during acute lung injury. Mol. Immunol. 2019, 112, 51–58. [Google Scholar] [CrossRef]
- Polman, C.H.; Reingold, S.C.; Banwell, B.; Clanet, M.; Cohen, J.A.; Filippi, M.; Fujihara, K.; Havrdova, E.; Hutchinson, M.; Kappos, L.; et al. Diagnostic criteria for multiple sclerosis: 2010 revisions to the McDonald criteria. Ann. Neurol. 2011, 69, 292–302. [Google Scholar] [CrossRef] [Green Version]
- Adams, H.P., Jr.; Bendixen, B.H.; Kappelle, L.J.; Biller, J.; Love, B.B.; Gordon, D.L.; Marsh, E.E., III. Classification of subtype of acute ischemic stroke. Definitions for use in a multicenter clinical trial. TOAST. Trial of Org 10172 in Acute Stroke Treatment. Stroke 1993, 24, 35–41. [Google Scholar] [CrossRef] [Green Version]
- Mikrut, M.; Mazuryk, O.; Macyk, W.; van Eldik, R.; Stochel, G. Generation and photogeneration of hydroxyl radicals and singlet oxygen by particulate matter and its inorganic components. J. Environ. Chem. Eng. 2021, 9, 106478. [Google Scholar] [CrossRef]
- Baran, J.; Kowalczyk, D.; Ożóg, M.; Zembala, M. Three-color flow cytometry detection of intracellular cytokines in peripheral blood mononuclear cells: Comparative analysis of phorbol myristate acetate-ionomycin and phytohemagglutinin stimulation. Clin. Diagn. Lab. Immunol. 2001, 8, 303–313. [Google Scholar] [CrossRef] [Green Version]
- Wnuk, M.; Maluchnik, M.; Perwieniec, J.; Podwojcic, K.; Szelag, M.; Walkiewicz, D.; Zakrzewski, M.; Kulakowska, A.; Brola, W.; Rejdak, K.; et al. Multiple sclerosis incidence and prevalence in Poland: Data from administrative health claims. Mult. Scler. Relat. Disord. 2021, 55, 103162. [Google Scholar] [CrossRef]
- Iltchev, P.; Śliwczyński, A.; Czeleko, T.; Sierocka, A.; Tłustochowicz, M.; Tłustochowicz, W.; Timler, D.; Brzozowska, M.; Szatko, F.; Marczak, M. Epidemiology of Rheumatoid Arthritis (RA) in rural and urban areas of Poland—2008–2012. Ann. Agric. Environ. Med. 2016, 23, 350–356. [Google Scholar] [CrossRef] [Green Version]
- Kruger, P.C.; Guzik, T.J.; Eikelboom, J.W. How can the results of the COMPASS trial benefit patients with coronary or peripheral artery disease in Poland? Kardiol. Pol. 2019, 77, 661–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kvien, T.K.; Uhlig, T.; Ødegård, S.; Heiberg, M.S. Epidemiological aspects of rheumatoid arthritis: The sex ratio. Ann. N. Y. Acad. Sci. 2006, 1069, 212–222. [Google Scholar] [CrossRef] [PubMed]
- Sellner, J.; Kraus, J.; Awad, A.; Milo, R.; Hemmer, B.; Stüve, O. The increasing incidence and prevalence of female multiple sclerosis—A critical analysis of potential environmental factors. Autoimmun. Rev. 2011, 10, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Fairweather, D. Sex differences in inflammation during atherosclerosis. Clin. Med. Insights Cardiol. 2015, 8, 49–59. [Google Scholar] [CrossRef] [Green Version]
- Druet, P.; Sheela, R.; Pelletier, L. Th1 and Th2 cells in autoimmunity. Clin. Exp. Immunol. 1995, 101, 9–12. [Google Scholar] [CrossRef]
- Pryjma, J.; Baran, J.; Ernst, M.; Woloszyn, M.; Flad, H.D. Altered antigen-presenting capacity of human monocytes after phagocytosis of bacteria. Infect. Immun. 1994, 62, 1961–1967. [Google Scholar] [CrossRef] [Green Version]
- Samek, L.; Styszko, K.; Stegowski, Z.; Zimnoch, M.; Skiba, A.; Turek-Fijak, A.; Gorczyca, Z.; Furman, P.; Kasper-Giebl, A.; Rozanski, K. Comparison of PM10 Sources at Traffic and Urban Background Sites Based on Elemental, Chemical and Isotopic Composition: Case Study from Krakow, Southern Poland. Atmosphere 2021, 12, 1364. [Google Scholar] [CrossRef]
- Measurement data bank of the Chief Inspectorate of Environmental Protection. (In Polish). Available online: https://powietrze.gios.gov.pl/pjp/archives (accessed on 15 February 2021).
- Cohen, A.J.; Brauer, M.; Burnett, R.; Anderson, H.R.; Frostad, J.; Estep, K.; Balakrishnan, K.; Brunekreef, B.; Dandona, L.; Dandona, R.; et al. Estimates and 25-year trends of the global burden of disease attributable to ambient air pollution: An analysis of data from the Global Burden of Diseases Study 2015. Lancet 2017, 389, 1907–1918. [Google Scholar] [CrossRef] [Green Version]
- Ma, Q.Y.; Huang, D.Y.; Zhang, H.J.; Wang, S.; Chen, X.F. Exposure to particulate matter 2.5 (PM2.5) induced macrophage-dependent inflammation, characterized by increased Th1/Th17 cytokine secretion and cytotoxicity. Int. Immunopharmacol. 2017, 50, 139–145. [Google Scholar] [CrossRef]
- Angelici, L.; Piola, M.; Cavalleri, T.; Randi, G.; Cortini, F.; Bergamaschi, R.; Baccarelli, A.A.; Bertazzi, P.A.; Pesatori, A.C.; Bollati, V. Effects of particulate matter exposure on multiple sclerosis hospital admission in Lombardy region, Italy. Environ. Res. 2016, 145, 68–73. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Xu, Z.; Dan, Y.L.; Cheng, J.; Zhao, C.N.; Mao, Y.M.; Xiang, K.; Hu, Y.Q.; He, Y.S.; Pan, H.F. Association between traffic-related air pollution and hospital readmissions for rheumatoid arthritis in Hefei, China: A time-series study. Environ. Pollut. 2021, 268, 115628. [Google Scholar] [CrossRef] [PubMed]
- Hart, J.E.; Källberg, H.; Laden, F.; Bellander, T.; Costenbader, K.H.; Holmqvist, M.; Klareskog, L.; Alfredsson, L.; Karlson, E.W. Ambient air pollution exposures and risk of rheumatoid arthritis: Results from the Swedish EIRA case-control study. Ann. Rheum. Dis. 2013, 72, 888–894. [Google Scholar] [CrossRef] [PubMed]
- Hennig, F.; Geisel, M.H.; Kälsch, H.; Lucht, S.; Mahabadi, A.A.; Moebus, S.; Erbel, R.; Lehmann, N.; Jöckel, K.H.; Scherag, A.; et al. Air Pollution and Progression of Atherosclerosis in Different Vessel Beds-Results from a Prospective Cohort Study in the Ruhr Area, Germany. Environ. Health Perspect. 2020, 128, 107003. [Google Scholar] [CrossRef] [PubMed]
- Hasslöf, H.; Molnár, P.; Andersson, E.M.; Spanne, M.; Gustafsson, S.; Stroh, E.; Engström, G.; Stockfelt, L. Long-term exposure to air pollution and atherosclerosis in the carotid arteries in the Malmö diet and cancer cohort. Environ. Res. 2020, 191, 110095. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, J.D.; Adar, S.D.; Barr, R.G.; Budoff, M.; Burke, G.L.; Curl, C.L.; Daviglus, M.L.; Diez Roux, A.V.; Gassett, A.J.; Jacobs, D.R., Jr.; et al. Association between air pollution and coronary artery calcification within six metropolitan areas in the USA (the Multi-Ethnic Study of Atherosclerosis and Air Pollution): A longitudinal cohort study. Lancet 2016, 388, 696–704. [Google Scholar] [CrossRef] [Green Version]
- Cortese, A.; Lova, L.; Comoli, P.; Volpe, E.; Villa, S.; Mallucci, G.; La Salvia, S.; Romani, A.; Franciotta, D.; Bollati, V.; et al. Air pollution as a contributor to the inflammatory activity of multiple sclerosis. J. Neuroinflamm. 2020, 17, 334. [Google Scholar] [CrossRef]
- Tedgui, A.; Mallat, Z. Cytokines in atherosclerosis: Pathogenic and regulatory pathways. Physiol. Rev. 2006, 86, 515–581. [Google Scholar] [CrossRef] [Green Version]
- Taleb, S.; Tedgui, A.; Mallat, Z. IL-17 and Th17 cells in atherosclerosis: Subtle and contextual roles. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 258–264. [Google Scholar] [CrossRef] [Green Version]
- Noronha, A.; Toscas, A.; Jensen, M.A. Interferon beta decreases T cell activation and interferon gamma production in multiple sclerosis. J. Neuroimmunol. 1993, 46, 145–153. [Google Scholar] [CrossRef]
- Panitch, H.S.; Hirsch, R.L.; Haley, A.S.; Johnson, K.P. Exacerbations of multiple sclerosis in patients treated with gamma interferon. Lancet 1987, 1, 893–895. [Google Scholar] [CrossRef]
- Steinman, L. A brief history of T(H)17, the first major revision in the T(H)1/T(H)2 hypothesis of T cell-mediated tissue damage. Nat. Med. 2007, 13, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Luchtman, D.W.; Ellwardt, E.; Larochelle, C.; Zipp, F. IL-17 and related cytokines involved in the pathology and immunotherapy of multiple sclerosis: Current and future developments. Cytokine Growth Factor Rev. 2014, 25, 403–413. [Google Scholar] [CrossRef]
- Rostami, A.; Ciric, B. Role of Th17 cells in the pathogenesis of CNS inflammatory demyelination. J. Neurol. Sci. 2013, 333, 76–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Firestein, G.S. Evolving concepts of rheumatoid arthritis. Nature 2003, 423, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Davis, L.S.; Cush, J.J.; Schulze-Koops, H.; Lipsky, P.E. Rheumatoid synovial CD4+ T cells exhibit a reduced capacity to differentiate into IL-4-producing T-helper-2 effector cells. Arthritis Res. 2001, 3, 54–64. [Google Scholar] [CrossRef] [Green Version]
- van der Graaff, W.L.; Prins, A.P.; Niers, T.M.; Dijkmans, B.A.; van Lier, R.A. Quantitation of interferon gamma- and interleukin-4-producing T cells in synovial fluid and peripheral blood of arthritis patients. Rheumatology 1999, 38, 214–220. [Google Scholar] [CrossRef] [Green Version]
- Kanik, K.S.; Hagiwara, E.; Yarboro, C.H.; Schumacher, H.R.; Wilder, R.L.; Klinman, D.M. Distinct patterns of cytokine secretion characterize new onset synovitis versus chronic rheumatoid arthritis. J. Rheumatol. 1998, 25, 16–22. [Google Scholar]
- Mikrut, M.; Macyk, W.; van Eldik, R.; Stochel, G. Physicochemical Analysis of Water Extracts of Particulate Matter from Polluted Air in the Area of Kraków, Poland. Atmosphere 2021, 12, 565. [Google Scholar] [CrossRef]
- Jeanjean, M.; Bind, M.A.; Roux, J.; Ongagna, J.C.; de Sèze, J.; Bard, D.; Leray, E. Ozone, NO2 and PM10 are associated with the occurrence of multiple sclerosis relapses. Evidence from seasonal multi-pollutant analyses. Environ. Res. 2018, 163, 43–52. [Google Scholar] [CrossRef] [Green Version]
- Roux, J.; Bard, D.; Le Pabic, E.; Segala, C.; Reis, J.; Ongagna, J.C.; de Sèze, J.; Leray, E. Air pollution by particulate matter PM10 may trigger multiple sclerosis relapses. Environ. Res. 2017, 156, 404–410. [Google Scholar] [CrossRef]
- Deiuliis, J.A.; Kampfrath, T.; Zhong, J.; Oghumu, S.; Maiseyeu, A.; Chen, L.C.; Sun, Q.; Satoskar, A.R.; Rajagopalan, S. Pulmonary T cell activation in response to chronic particulate air pollution. Am. J. Physiol. Lung Cell. Mol. Physiol. 2012, 302, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Ulmer, A.J.; Flad, H.; Rietschel, T.; Mattern, T. Induction of proliferation and cytokine production in human T lymphocytes by lipopolysaccharide (LPS). Toxicology 2000, 152, 37–45. [Google Scholar] [CrossRef]
- Ifrim, D.C.; Quintin, J.; Joosten, L.A.; Jacobs, C.; Jansen, T.; Jacobs, L.; Gow, N.A.; Williams, D.L.; van der Meer, J.W.; Netea, M.G. Trained immunity or tolerance: Opposing functional programs induced in human monocytes after engagement of various pattern recognition receptors. Clin. Vaccine Immunol. 2014, 21, 534–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biswas, S.K.; Lopez-Collazo, E. Endotoxin tolerance: New mechanisms, molecules and clinical significance. Trends Immunol. 2009, 30, 475–487. [Google Scholar] [CrossRef] [PubMed]
- Dalpke, A.H.; Lehner, M.D.; Hartung, T.; Heeg, K. Differential effects of CpG-DNA in Toll-like receptor-2/-4/-9 tolerance and cross-tolerance. Immunology 2005, 116, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Shoenfelt, J.; Mitkus, R.J.; Zeisler, R.; Spatz, R.O.; Powell, J.; Fenton, M.J.; Squibb, K.A.; Medvedev, A.E. Involvement of TLR2 and TLR4 in inflammatory immune responses induced by fine and coarse ambient air particulate matter. J. Leukoc. Biol. 2009, 86, 303–312. [Google Scholar] [CrossRef] [Green Version]
- Aalapati, S.; Ganapathy, S.; Manapuram, S.; Anumolu, G.; Prakya, B.M. Toxicity and bioaccumulation of inhaled cerium oxide nanoparticles in CD1 mice. Nanotoxicology 2014, 8, 786–798. [Google Scholar]
- Steenhof, M.; Gosens, I.; Strak, M.; Godri, K.J.; Hoek, G.; Cassee, F.R.; Mudway, I.S.; Kelly, F.J.; Harrison, R.M.; Lebret, E.; et al. In vitro toxicity of particulate matter (PM) collected at different sites in the Netherlands is associated with PM composition, size fraction and oxidative potential–the RAPTES project. Part. Fibre Toxicol. 2011, 8, 26. [Google Scholar] [CrossRef] [Green Version]
- Tousoulis, D.; Oikonomou, E.; Economou, E.K.; Crea, F.; Kaski, J.C. Inflammatory cytokines in atherosclerosis: Current therapeutic approaches. Eur. Heart J. 2016, 37, 1723–1732. [Google Scholar] [CrossRef] [Green Version]
- Sharief, M.K.; Hentges, R. Association between tumor necrosis factor-alpha and disease progression in patients with multiple sclerosis. N. Engl. J. Med. 1991, 325, 467–472. [Google Scholar] [CrossRef]
- Lubberts, E. IL-17/Th17 targeting: On the road to prevent chronic destructive arthritis? Cytokine 2008, 41, 84–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feldmann, M.; Brennan, F.M.; Maini, R.N. Role of cytokines in rheumatoid arthritis. Annu. Rev. Immunol. 1996, 14, 397–440. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Noh, J.; Noh, Y.; Oh, S.S.; Koh, S.B.; Kim, C. Gender Difference in the Effects of Outdoor Air Pollution on Cognitive Function among Elderly in Korea. Front. Public Health 2019, 7, 375. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Frequency of Disease in the Population [%] | Number | Female to Male Ratio [%] | Age [Mean Years ± SD] | ||||
|---|---|---|---|---|---|---|---|---|
| Overall | Female | Male | Overall | Female | Male | |||
| MS | 0.13 * | 16 | 10 | 6 | 1.670 | 30.88 ± 7.36 | 31.00 ± 7.97 | 30.5 ± 7.78 |
| RA | 0.44 * | 11 | 7 | 4 | 1.750 | 43.55 ± 10.51 | 40.50 ± 9.75 | 45.29 ± 11.27 |
| AS | 3.00 ** | 20 | 7 | 13 | 0.538 | 69.90 ± 11.02 | 75.14 ± 10.30 | 67.08 ± 10.70 |
| HD | 20 | 7 | 13 | 0.538 | 38.90 ± 11.17 | 39.57 ± 11.87 | 38.54 ± 11.25 | |
| Season | |||
|---|---|---|---|
| Summer | Winter | ||
| PM10 | PM2.5 | PM10 | PM2.5 |
| 17.53 ± 9.27 | 9.98 ± 5.46 | 76.43 ± 36.77 | 50.46 ± 27.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gałuszka-Bulaga, A.; Węglarczyk, K.; Latacz, P.; Jodłowska-Cicio, K.; Korkosz, M.; Pera, J.; Słowik, A.; Siedlar, M.; Baran, J. Seasonal Variations in the Concentration of Particulate Matter in the Air of Cracow Affect the Magnitude of CD4+ T Cell Subsets Cytokine Production in Patients with Inflammatory and Autoimmune Disorders. Atmosphere 2022, 13, 529. https://0-doi-org.brum.beds.ac.uk/10.3390/atmos13040529
Gałuszka-Bulaga A, Węglarczyk K, Latacz P, Jodłowska-Cicio K, Korkosz M, Pera J, Słowik A, Siedlar M, Baran J. Seasonal Variations in the Concentration of Particulate Matter in the Air of Cracow Affect the Magnitude of CD4+ T Cell Subsets Cytokine Production in Patients with Inflammatory and Autoimmune Disorders. Atmosphere. 2022; 13(4):529. https://0-doi-org.brum.beds.ac.uk/10.3390/atmos13040529
Chicago/Turabian StyleGałuszka-Bulaga, Adrianna, Kazimierz Węglarczyk, Paweł Latacz, Katarzyna Jodłowska-Cicio, Mariusz Korkosz, Joanna Pera, Agnieszka Słowik, Maciej Siedlar, and Jarek Baran. 2022. "Seasonal Variations in the Concentration of Particulate Matter in the Air of Cracow Affect the Magnitude of CD4+ T Cell Subsets Cytokine Production in Patients with Inflammatory and Autoimmune Disorders" Atmosphere 13, no. 4: 529. https://0-doi-org.brum.beds.ac.uk/10.3390/atmos13040529