Diatom Taxonomic Composition as a Biological Indicator of the Ecological Health and Status of a River Basin under Agricultural Influence


1
College of Life Science and Technology, Harbin Normal University, Harbin 150025, China
2
Key Laboratory of Biodiversity of Aquatic Organisms, Harbin Normal University, Harbin 150025, China
*
Authors to whom correspondence should be addressed.
Water 2020, 12(7), 2067; https://0-doi-org.brum.beds.ac.uk/10.3390/w12072067
Submission received: 2 June 2020
/
Revised: 14 July 2020
/
Accepted: 18 July 2020
/
Published: 21 July 2020
(This article belongs to the Special Issue Freshwater Biophysical Ecosystem Health)
Abstract
:The Lalin River Basin (LLRB) is a major drainage basin in northeastern China, that has been significantly influenced by agricultural activities. This study focused on exploring diatom taxonomic composition linked to environmental factors at the taxonomic levels of genus and species during ice-covered periods. Nine sampling stations were divided into three groups based on trophic state index (TSI). hierarchical cluster analysis (HCA) and redundancy analysis (RDA) were performed to identify diatom distribution patterns and their relationships to environmental factors. Diatom richness, composition and distribution were analyzed at the levels of genus and species. Our results showed the epipelagic diatom Melosira varians was very abundant at most stations. Benthic diatoms Achnanthidium minutissimum, Encyonema minutum and Gomphonema parvulum were dominant in group-3, which had the highest trophic states. HCA showed the similarity of diatom taxonomic composition spatial distribution patterns between genus and species levels. RDA revealed that the key factors related to genus level distributions are COD, TP and EC, while TP was the key factor in structuring diatom taxonomic composition at the level of species. These results suggest identification of diatoms at genus level can be used as a potential indicator to assess ecological health status of agricultural-influenced rivers during ice cover periods. Further research is necessary to explore the utility of genus level diatom composition as a biological indicator in rivers.
1. Introduction
Rivers are exposed to increasing human pressure all over the world particularly from agricultural activities [1]. During the past decades, however, agricultural practices have significantly increased, causing visible effects on rivers (overuse of pesticides or fertilizer, etc.), but their impacts have been very difficult to assess [2,3]. Ice-covered periods of rivers can play an essential role in nutrient cycling and biodegradation in rivers of the temperate zone [4]. Furthermore, the ice-covered periods can be closely associated with climate reconstruction [5], organic pollution [6] and winter algal blooms [7,8]. In recent years, most studies have focused on developing tools for assessing environmental changes of aquatic ecosystem [9,10]. Diatoms have been widely applied for this purpose and used in bio-monitoring studies in peatlands [11], rivers [12] and lakes [5] as they can tolerate low light levels and low water temperatures, and they can respond rapidly to environmental change [13]. Specifically, the distribution of diatoms in rivers can reflect the combined effects of environmental gradients and reveal the health of river ecosystems [12]. Increasing intensity of agricultural land practices affects rivers, and these impacts would closely limit diatom community assembly [9].
Eutrophication and organic contamination of Chinese agricultural practices, which are undergoing rapid growth due to both economic and population increases, have become a serious threat to river basins [14]. Heilongjiang Province is located in the northeastern of China, which is one of the fastest developing agricultural regions in China. Lalin River Basin (LLRB) is the main grain production district in Heilongjiang Province [15]. More than half of the river area in LLRB is now occupied by grain cultivation [1]. Dense agricultural practices and intensive human activities resulted in high nutrient levels and organic contamination in the LLRB [1,15]. Studies indicate that genus-level identification might increase the accurate prediction of environmental change [16,17]. Diatom-based assessment studies of anthropogenic eutrophic rivers of NE China are still few [18,19], and none of these studies examined conditions during ice cover.
Herein, we hypothesized that the use of genus-level characterization of diatom taxonomic composition has the potential to provide a signal linking river environmental change during ice-cover. In this study, diatom taxonomic composition at both genus- and species-levels and environmental factors were conducted from Lalin River Basin during the ice cover period in 2018. Comprehensive trophic state index (TSI) was used to define the trophic states, hierarchical cluster analysis (HCA) and redundancy analysis (RDA) were performed to determine the diatom distribution pattern and their association with environmental factors at the genus- and species level. The purposes of this study were to (1) determine diatom taxonomic composition in a typical anthropogenic eutrophic river of northeastern China and (2) examine the association of environmental factors and diatom taxonomic composition at genus and species levels during ice cover.
2. Materials and Methods
2.1. Study Area, Sample Collection and Laboratory Analyses
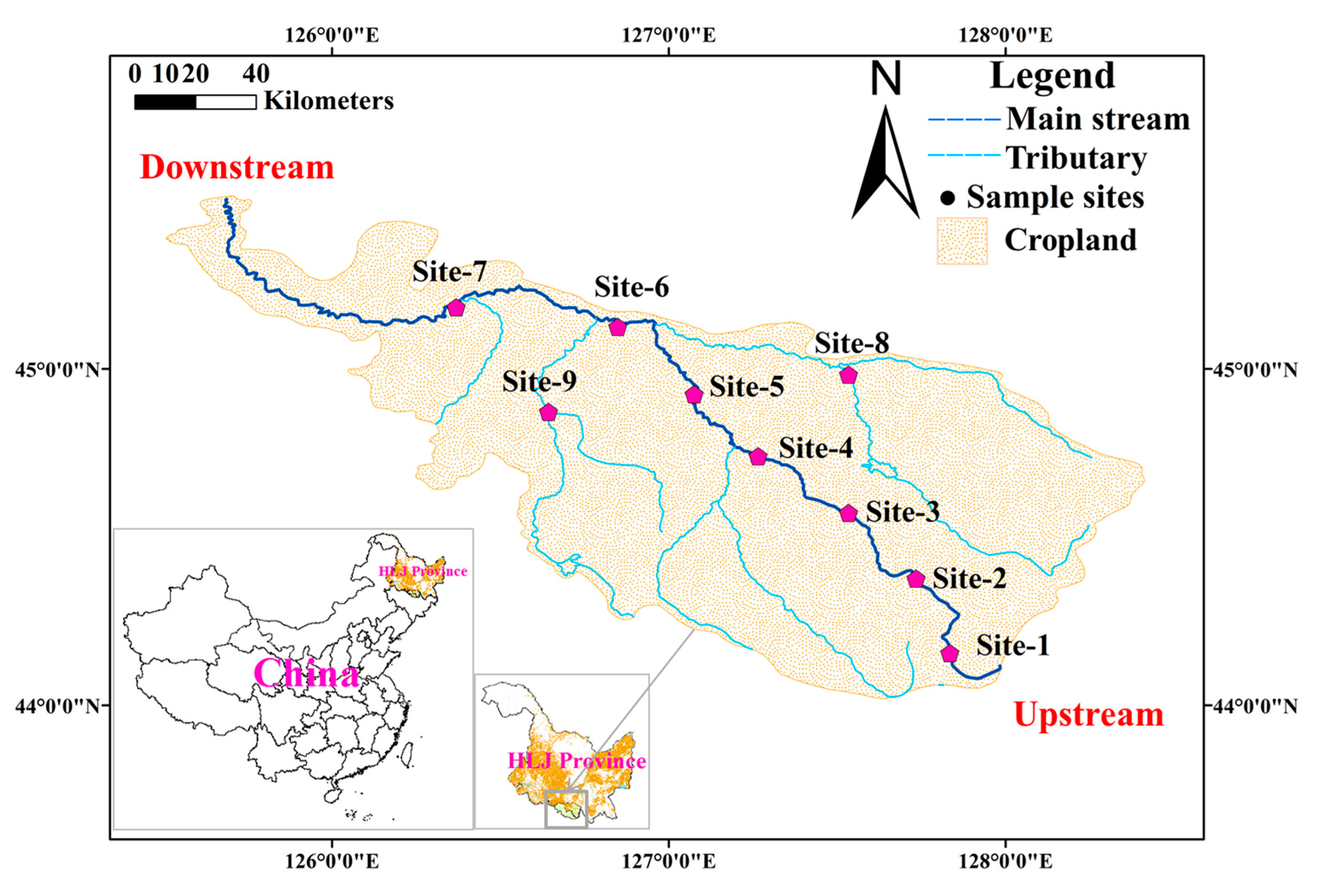
The Lalin River Basin (LLRB) is located in NE China, and it is characterized by long cold winters and warm summers with a mean annual rainfall of ~500–800 mm, maximum temperature in summer of 35.6 °C, and minimum temperature in winter of −45.4 °C [1]. The ice-covered period is more than 100 days per year. This region is famous for its huge grain production and has been referred to as “rice city”. Most of the region of LLRB is eutrophic due to agricultural actives [20]. In this study, a total of 9 stations were established to collect the ice-water samples in February 2018 (Figure 1). These sampling stations spanned from upstream to downstream of LLRB. Remote sensing (RS) showed that the intensity of agricultural practice increased from upstream to downstream. At each sampling station, geographic coordinates were determined using a Garmin Etrex GPS. RS information was downloaded from Resource and Environment data cloud Platform of China (http://www.resdc.cn/) and applied in ArcMap 10.3. The ice-water samples were collected by “ice piton sampler”. A total of 10 L of ice-water samples were collect by plastic bottle preserved with 1% Lugol’s solution and refrigerated under dark conditions until laboratory analysis [21]. A YSI multi-metric probe was used to measure physical factors in the field, such as water temperature (WT), dissolved oxygen (DO), conductivity (EC) and pH. Chemical factors such as total nitrogen (TN), total phosphorus (TP) and chemical oxygen demand (COD) were determined by Chinese national standards for water quality in laboratory within 24 h [22].
Before qualitative and quantitative analysis, diatom samples were transferred into a beaker and heated to 80 °C with 30% H2O2 [13]. All digestion and centrifugation were according to Chen et al. [11]. Diatom counts from each station were expressed as relative abundances to eliminate the effects of rare species [23]. At least 600 diatom frustules (>2 µm) were counted under at 1000× magnification using Zeiss imager A2 microscope. Dominant species were defined as those with a relative abundance >5% in at least one station. Diatom taxa identification was mainly based on Lange-Bertalot et al. [24], Lange-Bertalot et al. [25], and Wojtal [26].
2.2. Multivariate Statistical Analysis
In total, 10 physicochemical factors were considered for multivariate statistical analysis, including WT, DO, EC, pH, ORP, Tur., Chl-a, TN, TP and COD. All 10 factors and diatom data were normalized by [log10(x+1)] transformation. Comprehensive trophic state index (TSI) was used to describe the trophic status [22]. The equations for TSI are as follows:
TSI(TN) = 10(5.453 + 1.694 ln TN)
TSI(TP) = 10(9.436 + 1.642 ln TP)
TSI(Chl-a) = 10(2.5 + 1.086 ln Chl-a)
TSI(COD) = 10(1.2 + 1.566 ln COD)
TSIM(∑) = [TSI (TN) + TSI (TP) + TSI (COD) + TSI (Chl-a)]/4
Evaluation standard:0 < TSIM < 30 oligotrophic, 30 < TSIM < 50 mesotrophic, TSIM > 50 eutrophic, 50 < TSIM < 60 light eutrophic, 60 < TSIM < 70 middle eutrophic, TSIM > 70 high eutrophic.
Hierarchical Cluster analysis (HCA) was performed for both species- and genus-level data to examine diatom distribution patterns along the trophic gradient. HCA, Ward method based on Euclidian distances was conducted by HemI software. Ordinate model was conducted by CANOCO 4.5 on environmental factors and diatom data. Linear model or unimodal model were determined by Detrended Correspondence Analysis (DCA) [27]. When gradient lengths were less than 3, linear model RDA was fit for ordinate; conversely, unimodal model CCA was more suited for ordinate while gradient lengths were more than 4. Moreover, CCA and RDA were compatible while the gradient lengths were within 3-4. In this study, dominant diatom taxa and 10 environmental factors were selected to carry out model (16 species and 11 genera, respectively). Monte Carlo permutation test (499 permutations) was necessary to determine the significant of relation between environmental factors and diatom taxonomic composition (cut-off point of p = 0.05). Independent-samples t test (t-test) was performed to identify the different of environmental factors, which was conducted by SPSS 22.0.
3. Results
3.1. Environmental Gradient of LLRB during Ice Cover
Sampling stations in this study were alkaline (mean pH 8.46) and had high nutrient concentrations (mean TN, TP and COD were 4.41, 0.60 and 33.44, respectively) (see Table 1). TSI indicated that trophic states of these 9 sampling stations ranged from mesotrophic to highly eutrophic (Table 2). The trophic traits conformed to the prior studies and RS information (Figure 1) [1,20]. According the TSI, the 9 sampling stations were divided into three groups manually. Group-1 included stations 1-2, which were mesotrophic states; group-2 included stations 3-5 and stations 8-9, which were middle eutrophic states, and group-3 included stations 6-7, which were highly eutrophic states. T-test showed that most physical factors had no significant difference beside EC (p < 0.05). Meanwhile, in chemical factors, TP and COD had significant differences (p < 0.05) among the three groups (Table 2).
3.2. Diatom Taxonomic Composition at Genus And Species Levels
In this study, a total of 50 diatom taxa were identified, which belong to 25 genera. Most taxa were pennate diatoms, and higher richness was mainly observed in Navicula (7 taxa), Gomphonema (6 taxa), Fragilaria (4 taxa), Surirella (4 taxa), Nitzschia (3 taxa) and Cymbella (3 taxa). The dominant species (relative abundance of more than 5% in at least one sample) included Achnanthidium minutissimum, Cocconeis placentula, Cymbella tumida, Encyonema minutum, Encyonema silesiacum, Fragilaria vaucheriae var. capitellata, Gomphonema acuminatum, Gomphonema parvulum, Hannaea arcus, Melosira varians, Navicula viridula var. rostellata, Navicula viridula, Surirella angusta, Surirella minuta, Surirella visurgis and Synedra ulna. These 16 dominated species contributed 79.33–90.33% of relative abundance in all sampling stations.
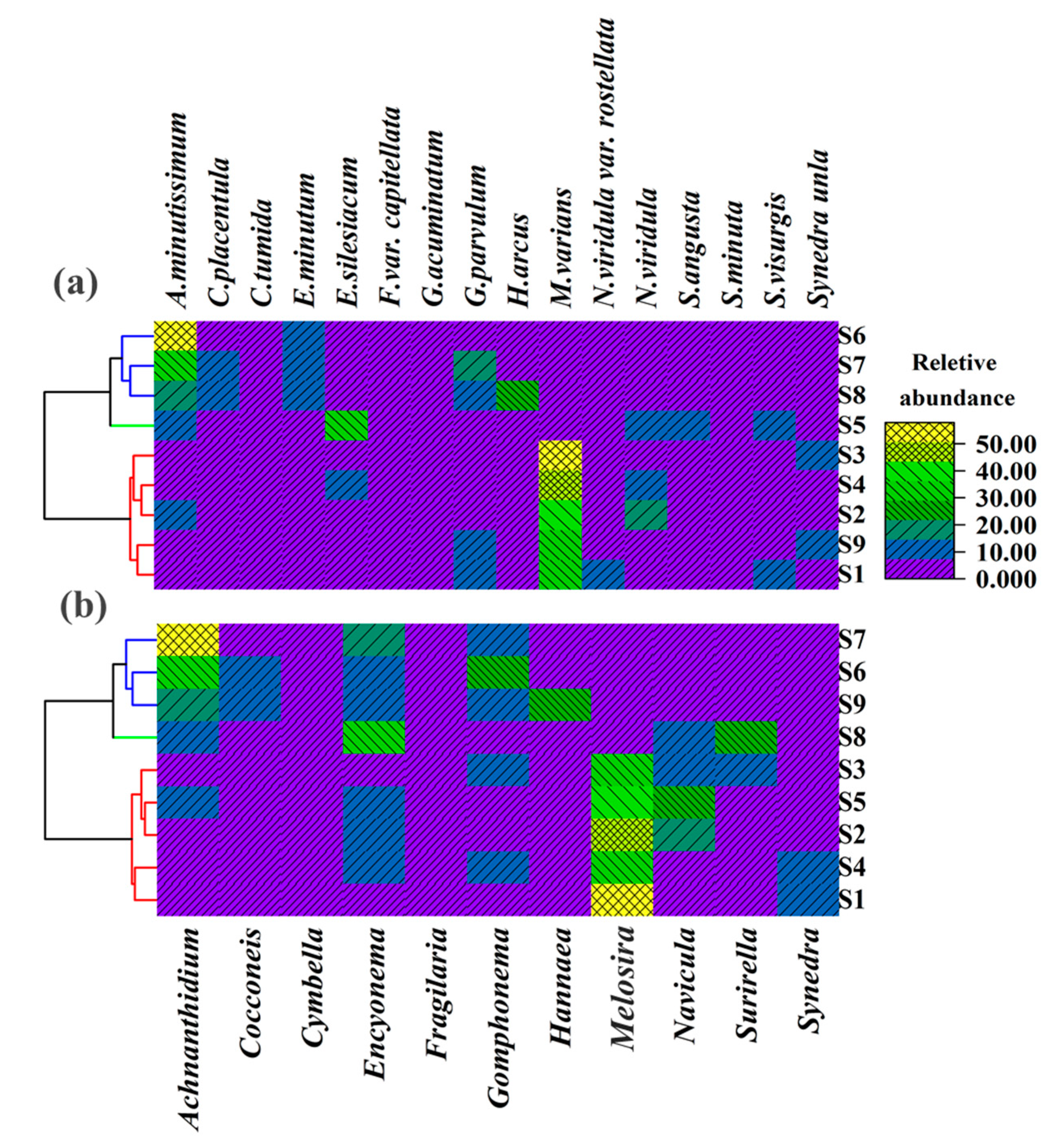
Hierarchical clustering (HCA) (Ward, Euclidean Distance) based on species level diatom taxonomic composition indicated all 9 sampling stations can be divided into three categories: mesotrophic to middle eutrophic (staiton-1, station-2, station-3, station-4 and station-9), middle eutrophic (station-5) and middle eutrophic to high eutrophic (station-6, station-7 and station-8). At genus-level HCA showed a comparable pattern with species-level, especially station-6 and station-7, which had the highest TSI and clustered into the same category (Figure 2a,b).
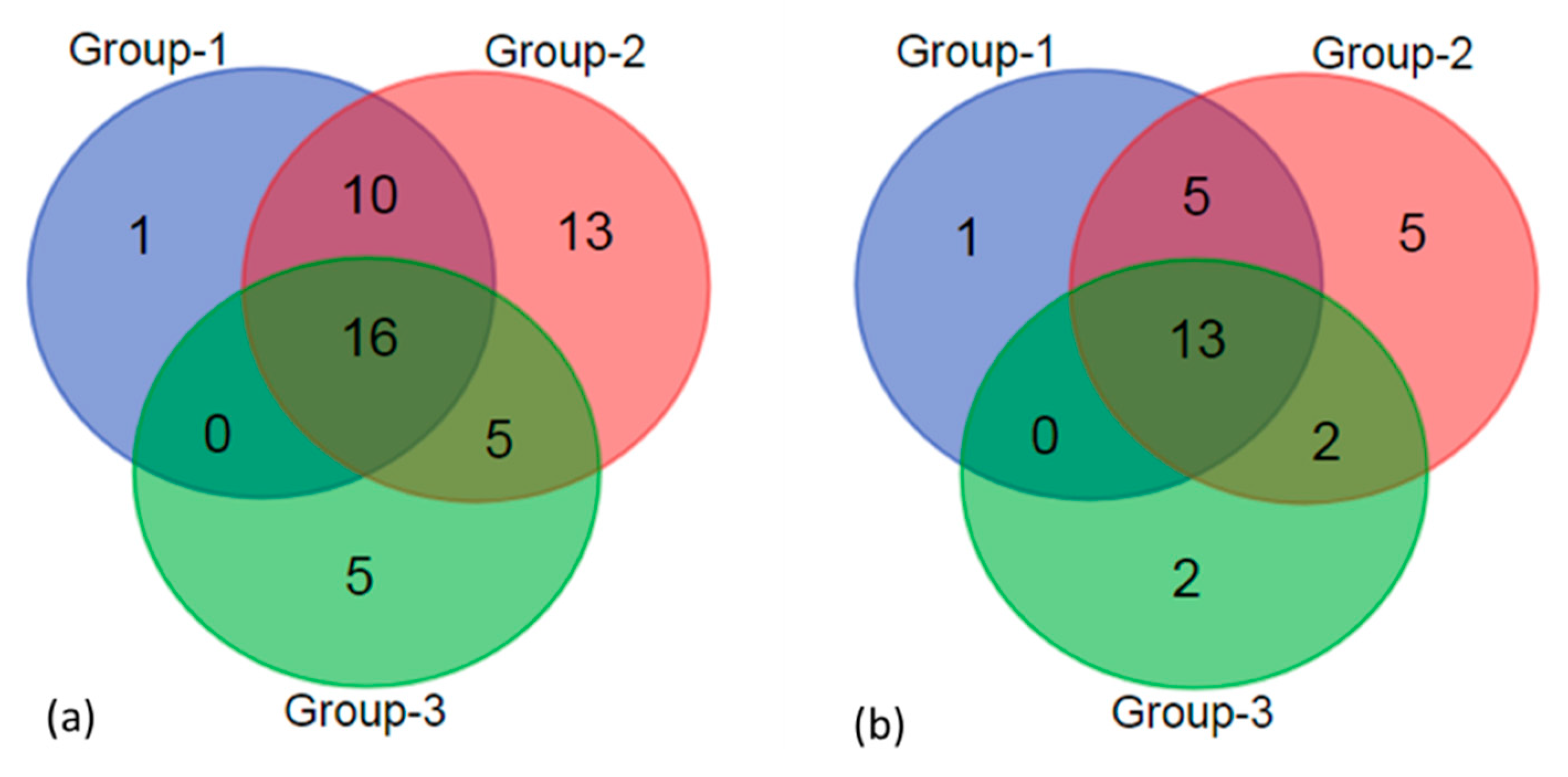
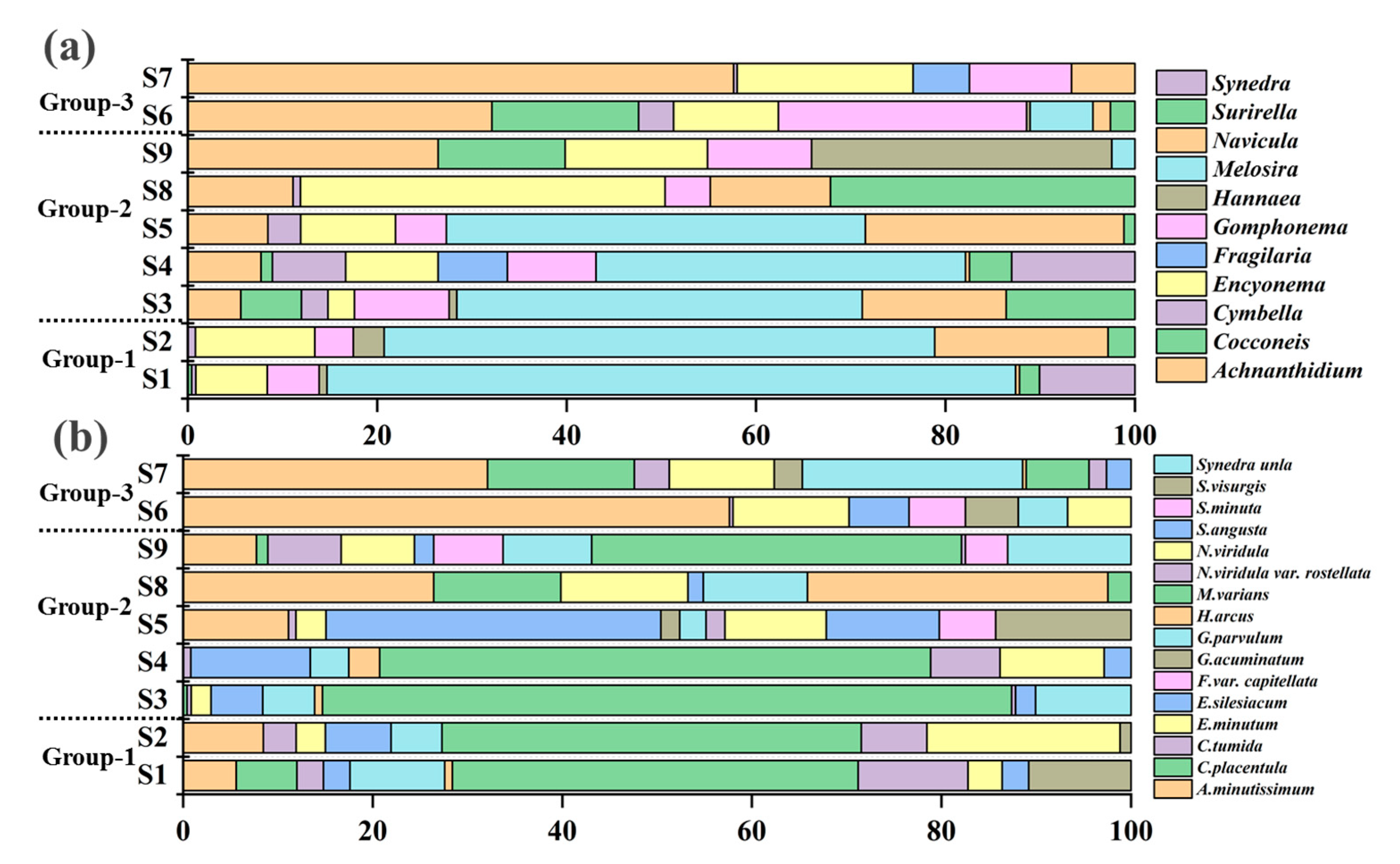
Species level data yielded diatom richness for three groups as 27, 44 and 26 taxa, respectively. The sixteen common species occurred in all three groups (Figure 3a). Along the trophic gradient, the common species show a decreasing trend, which is 10 (group-1 and group-2), 5 (group-2 and group-3) and 0 (group-1 and group-3) (Figure 3a). In group-1, Melosira varians was the pre-dominant species with 37% relative abundance, which was much higher than that in the other two groups. In addition, Navicula viridula, Gomphonema parvulum and Achnanthidium minutissimum were found to be relatively important in group-1 (Figure 4a). Melosira varians was pre-dominant in group-2. Achnanthidium minutissimum and Encyonema silesiacum were co-dominant in group-2, which were found to have higher relative abundance than group-1. Achnanthidium minutissimum was pre-dominated in group-3, which contributed more than 40% relative abundance (Figure 4a).
In the genus-level analysis, the diatom richness of three groups (group-1 to group-3) was 19, 25 and 18 genera, respectively. Thirteen common genera occurred in all three groups (Figure 3b). Similar to the species-level data, the common species exhibited a decreasing trend, which is 5 (group-1 and group-2), 2 (group-2 and group-3) and 0 (group-1 and group-3) (Figure 3b). We found that the dominant diatoms in the genus level analysis had a pattern similar to the species-level data. The dominance of common genera changed significantly, with Melosira being replaced by Achnanthidium and Encyonema (Figure 4b). The change in dominant taxa in both the genus-level and species-level data that was also seen with increasing nutrients suggested that the diatom taxonomic composition had a vital link to environmental changes during the ice-covered period.
3.3. Diatom Taxonomic Composition Linked to Environmental Factors
A total of 16 dominant species were included in the ordination analysis. DCA showed the length gradients were 1.45 and 1.64 in species-level and genus-level data, respectively. Thus, the linear model RDA was selected to explore the correlation between diatom diversity patterns and environmental factors.
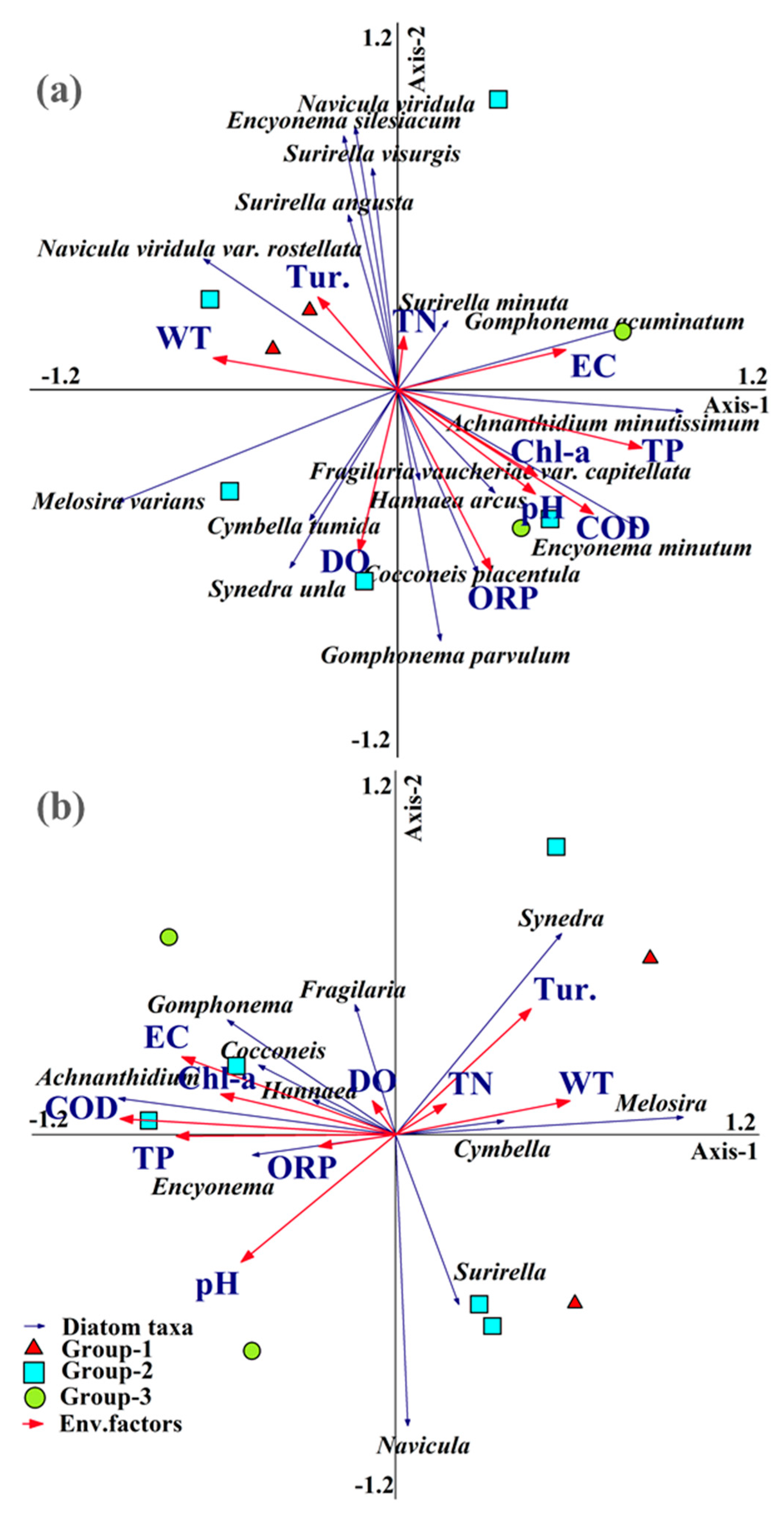
At the species level, the results of RDA showed that the first and second ordination axes explain 29.3% and 11.9% of the total variance, respectively. Monte Carlo permutation test indicated that TP (p < 0.002) was the key factor that facilitates dynamic diatom community ecological distribution during ice cover. The first axis is significantly positively correlated with TP (r2 = 0.67, p < 0.002) (Table 3). The second axis is negatively correlated with DO and ORP (r2 from −0.47 to −0.48, p > 0.05). Therefore, the first axis represents the effects of eutrophication on diatom diversity pattern, while axis 2 represents the impacts of physical factors. At the genus level, the results of RDA showed that the first and second ordination axes explain 40.3% and 20.9% of the total variance, respectively. COD, TP and EC were key factors related to diatom genus-level community assembly. The first axis is significantly negatively correlated with the COD, TP and EC (r2 = from −0.67 to −0.79, p < 0.002 or p < 0.05). The second axis is negatively correlated with pH and Tur. (r2 from −0.47 to −0.48, p > 0.05). As with the species level analysis, the eutrophic conditions affected the diatom composition strongly, while physical factors did not significantly correlate with diatom community structure as reflected in the genus-level data.
4. Discussion
In this study, we determined the diatom taxonomic composition of a typical agriculture-impacted, eutrophied river in NE China during ice cover. In addition, we explored the relationships between diatom richness, relative abundance and dominant taxa distribution with environmental factors at species-level and genus-level. Based on trophic state indices, all 9 sampling stations exhibit an obvious nutrient gradient during the study period (group-1 to group-3). Our results verified our hypothesis that genus-level diatom taxonomic composition was a potential signal for indicating river environmental changes during ice cover.
Compared with blue-green algae, diatoms have a great ability to tolerate extreme conditions such as low water temperature, low light, extreme pH and nutrient deficiencies [11,21,28]. Less light can penetrate opaque snow and ice during most of the ice-covered period in aquatic ecosystems [29]. During winter of LLRB, the ice cover period is long and low water temperatures prevail throughout the whole winter [1]. Compared with the warm seasons in LLRB, the diatom biodiversity is very low, 130 taxa vs. 50 taxa (unpublished data). Similar results were also reported in temperate aquatic systems in North America and Europe [10,30]. Thus, we suggest the low richness of LLRB was attributed to limited light penetration and low WT during ice cover. Ice cover was considered to have a direct impact on diatom community assembly by limiting the ability for movement [10]. Moreover, snow and ice cover obstructed the refraction of light in water body [5]. Centric diatoms were widely used as an indicator of changing lake thermal structure and capable of tolerating low light levels; they dominated in an ice-covered lake [29]. Katz et al. [8] reported that epipelagic Melosira had a potential ability to cause the under-ice blooms in winter. In this study, the dominant species (or genera) Melosira varians is generally considered to be tolerant of low light and ice cover [8]. Nutrient addition, it was shown, can increase the benthic diatom abundance in template rivers [12]. In this study, benthic diatom Achnanthidium, Encyonema and Gomphonema were co-dominants in highest nutrient state stations (group-3). Encyonema silesiacum and Gomphonema parvulum are considered as relevant eutrophic indicators that commonly occur in warm seasons of eutrophic lakes [31] and rivers [9,32]. The dominance of Encyonema silesiacum and Gomphonema parvulum in highest trophic stations of LLRB suggested that these two species had a potential ability to indicate the ecological states in agricultural eutrophic rivers during ice-covered periods. It is interesting to note that Achnanthidium minutissimum replaces Melosira varians as the dominant taxon in group-3, which had the highest nutrient conditions. Achnanthidium minutissimum was reported to have a wide range of tolerance for temperature, acidity, conductivity and higher primary productivity [12] and was capable of dominating in the fast flow of lotic environments during ice melt [33]. In our study, group-3 stations are located in downstream of LLRB, which present highest EC, Chl-a, TN and TP concentrations (Table 1). The habitat of group-3 provided an appropriate condition for Achnanthidium minutissimum population. Although some research has shown that Achnanthidium minutissimum was sensitive to high P_PO4 concentration in warm seasons (0.3 mg/L) [9], it occurred with high TP concentrations in several stations during ice cover. Moreover, the mechanism of Achnanthidium minutissimum being adept at high TP habitats still need further exploration.
Understanding how diatom community assembly is linked to environmental factors is a vital key to improving agricultural eutrophic rivers ecosystem management [2]. In Lalin River Basin, the main factors emanating from agricultural activities that impacted the aquatic organisms were organic and persistent organic pollutants [1,20]. Our results indicated that environmental factors were closely associated with diatom taxonomic composition at genus and species levels. In RDA, the station scores the weight sums of the species ecological information [34]. Therefore, the position of one sampling station (station here) in the ordination diagram represents the weights of all species in the site to the two axes. RDA showed three group regions were separately clustered together along eutrophic gradient, although there are some crossing points among them. Stations from group-3 are located in the axis 1, which are included in the arrows of TP. These results represent the higher eutrophic in this group. Stations of the group-2 are scattered in the diagram, mostly on opposite sides of TP, suggesting a middle eutrophication in this area. Group-1 grouped on the positive side of axis 1, indicating the relative lower trophic level in the area. Based on RDA, we found that spatial distribution of eutrophication limited the diatom community assembly, on the contrary, the physical factors had no significant correlation with diatom taxonomic composition expected from EC during ice-covered periods. At the species level, the diatom community strongly associated with the TP gradient (Axis-1) included Achnanthidium minutissimum and Gomphonema acuminatum. The results suggest that TP enhances the capability of Achnanthidium minutissimum and Gomphonema acuminatum to adapt in bad conditions and we infer these species have a potential ability to indicate eutrophication during ice-covered period. The effects of TP on the ecological distribution of diatoms have been revealed in freshwater [12], marine [35], and estuaries ecosystems [13]. General N:P ratio was the key issue to dynamic phytoplankton diversity pattern in aquatic ecosystem [35]. TP was linked to diatom nutrient utilization; when N:P ratio > 16:9, the phytoplankton ecological distribution was mainly determined by phosphorus [12]. In this study N:P ratio was 6.84, in which TP limited diatom community assembly. The three groups station distribution in RDA based on genus-level showed a comparable pattern with species level (Figure 5b). Furthermore, RDA showed the genus-level data were more sensitive to COD and EC than the species composition (Figure 5b). In genus level, stations in the Group-3 are located in the negative direction of axis 1, which are included in the arrows of EC, COD and TP. EC is an index of the total ionic concentration in rivers and had an effect on the diatoms community assembly, with individual diatoms showing affinity for particular ions under effect of herbicide contaminate rivers [36].The effects of EC on the distribution of diatom taxonomic composition have been reported by recent studies in rivers [37], lakes [38] and wetlands [39] during warm seasons. Pestryakova et al. recorded that diatom composition changed significantly along conductivity gradients in cold lake [40]. Agricultural activity had resulted in polycyclic aromatic hydrocarbon pollution and high ionic concentration in LLRB [1,20]. The measured EC values ranged from 34.7 to 270.4 μS cm−1, with the median value of 164.3 μS cm−1. EC was significantly different between Group-1 and Group-3. Our results suggested that the genus diatom taxonomic composition had a similar response along the EC gradient during ice cover. COD is an indicator of change in organic source of water column derived from agricultural waster sewage and herbicide abuse [41] and was reported to influence the benthic diatom taxonomic composition in a heavily polluted stream at similar latitude river in warm season. However, the mechanisms that link COD to diatom in ice-covered rivers are rarely reported. In our study, RDA indicated that COD was another covariate factor to dynamic diatom taxonomic composition distribution at genus level, and positively correlated with TP and EC. Chen et al. [1] noted that the organic pollution of LLRB was mainly in the form of agricultural sewage, and this is the reason why pollution-tolerant benthic taxa Encyonema and Gomphonema were co-dominants here. Most physical factors such as pH, WT and DO did not have any significant effect on diatom taxonomic composition during our research study. The effects on phytoplankton community assembly of physical factors are mostly dependent on the water flow regime during ice cover [33]. Notably, we found the nutrient stated had a change in some stations (station-8, see in Table 2). Rivers can recover the pollutant load within certain limits [42]. The TSI change at station 8 suggested that limitation of river self-purification ability to diatom diversity cannot be ignored in agricultural influenced rivers during ice cover [43]. Thus, the chemical factors, self-purification ability and velocity monitor in further river regulations during ice cover is necessary.
Diatom taxonomic composition has been linked to environmental change in rivers and lakes during cold or ice-covered periods [29,30]. Based on TSI, we found a significant nutrient gradient, where downstream stations (group-3) were higher in organic materials than middle and upstream stations (group-2 and group-1). T-test indicted EC, TP and COD, which are the key dynamic factors linking diatom taxonomic composition to environmental change in this study, were significantly different (p < 0.05 or p < 0.01). The Venn diagram showed that the common taxa number along the environmental gradient had the same trend in both the species-level and genus-level data. The hierarchical cluster analyses grouped together the sampling stations with similar nutrient states at species-level and genus level. The results suggest that species level diatom taxonomic composition was coupled to environmental gradients. The results of the HCA based on genus level data showed station-6 and station-7, where highest nutrient states were divided into the same category. The hierarchical cluster further proved that genus level diatom taxonomic composition had a potential ability to indicate environmental change. Previous studies argued that diatom at genus level shows only a weak link to environmental reconstruction [44]. Some studies indicated assessment of the integrity at genus level led to the loss of any signal of sensitivity of diatom diversity [45]. Exploring the key factors for dynamic community assembly is the vital component of a mechanism for assessment of aquatic ecological function and health. In our study, we found that RDA showed that genus level had a higher total variance (61.2%) than species level (41.2%) in diatom taxonomic composition linked to environmental factors. RDA revealed that genus-level attributes for environmental conditions in ice-covered rivers appear to be relatively more precise compared to species-level attributes. This finding was consistent with a previous study in peatland of central China [16]. Furthermore, the genus-level composition was more sensitive to COD and EC changes than the species composition. Growns [17] notes that lower richness of species within each relevant genus excludes the chances of differing responses of several species with the same genus to environmental factors. In general, the diatom richness of rivers in ice-covered periods is lower [30]. Hence, the similarity of the diatom genus and species composition linked to river environments can be ascribed to the fact that most of the genera have few species in ice cover. During ice melt, some diatoms valves were broken, resulting in loss of necessary information and increasing the monitoring time in river regulation and environmental research [33]. Both of RDA and HCA showed that species level and genus level diatom taxonomic compositions corresponded to environmental gradients of LLRB. This finding is consistent with our hypotheses. Under ice-covered condition, genus level identification is timesaving and a relevant tool for exploring the diatom taxonomic composition linked to environmental change. However, to further utilize diatom genus compositions to assess ecological health in rivers during the ice-covered period, the hydrological information gathered while ice melts and freezes is necessary.
5. Conclusions
The present study demonstrated that the taxonomic composition of diatoms at the species and genus level allows to detect changes in river ecosystems during ice-covered periods caused by environmental factors as a result of agricultural activities. The lower richness of diatom (50 taxa at the species level and 11 at the genus level) were found in ice cover. Based on multivariate statistical analysis, we found that the diatom taxonomic composition was able to distinguish the habitat trophic characteristic within the LLRB, showing a dominant diatom taxa changed at all 9 stations known to be affected by agricultural contamination and other agricultural activities. The results indicated that the spatial variations of mainly chemical factors were closely related to diatom taxonomic composition at genus and species levels. Since the use of information about taxa at a genus level is not developed yet and requires substantial knowledge of the ecology of species, we inferred that the genus level diatom composition is a potential signal for indicating river environmental changes during ice-covered periods. Overall, further exploring the effectiveness of genus level diatom composition as indicator of environmental change in rivers is necessary.
Author Contributions
X.L. formulated a concept of the paper, wrote its original draft, performed software, Y.L. review, editing the manuscript, investigation, improved the original text, figures and funding, Y.F. performed the project administration. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Natural Science Foundation of China (31870187; 31970213), Scientific Research Foundation of Harbin Normal University (XKB201912).
Acknowledgments
We are grateful to the distinguished reviewers for their helpful comments and suggestions. We thank J.P. Kociolek for his linguistic assistance during the preparation of this manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Chen, R.H.; Teng, Y.G.; Chen, H.Y.; Hu, B.; Yue, W.F. Groundwater pollution and risk assessment based on source apportionment in a typical cold agricultural region in Northeastern China. Sci. Total Environ. 2019, 696, 133972. [Google Scholar] [CrossRef] [PubMed]
- Hicks, M.B.; Taylor, J.M. Diatom Assemblage Changes in Agricultural Alluvial Plain Streams and Application for Nutrient Management. J. Environ. Qual. 2019, 48, 83–92. [Google Scholar] [CrossRef] [Green Version]
- Bellinger, B.J.; Angradi, T.R.; Bolgrien, D.W.; Jicha, T.M.; Hill, B.H.; Reavie, E.D. Longitudinal variation and response to anthropogenic stress in diatom assemblages of the Lower Mississippi River, USA. River Syst. 2013, 21, 29–54. [Google Scholar] [CrossRef]
- Kamari, M.; Alho, P.; Veijalainen, N.; Aaltonen, J.; Huokuna, M.; Lotsari, E. River ice cover influence on sediment transportation at present and under projected hydroclimatic conditions. Hydrol. Process. 2015, 29, 4738–4755. [Google Scholar] [CrossRef]
- Keatley, B.E.; Douglas, M.S.V.; Smol, J.P. Prolonged ice cover dampens diatom community responses to recent climatic change in High Arctic lakes. Arct. Antarct. Alp. Res. 2008, 40, 364–372. [Google Scholar] [CrossRef] [Green Version]
- Walter, P. Environmental pollution Sea ice cover linked to toxic mercury. Chem. Ind. 2011, 3, 7-7. [Google Scholar]
- Binding, C.E.; Greenberg, T.A.; Bukata, R.P.; Smith, D.E.; Twiss, M.R. The MERIS MCI and its potential for satellite detection of winter diatom blooms on partially ice-covered Lake Erie. J. Plankton Res. 2012, 34, 569–573. [Google Scholar] [CrossRef] [Green Version]
- Katz, S.L.; Izmest’eva, L.R.; Hampton, S.E.; Ozersky, T.; Shchapov, K.; Moore, M.V.; Shimaraeva, S.V.; Silow, E.A. The “Melosira” years of Lake Baikal: Winter environmental conditions at ice onset predict under-ice algal blooms in spring. Limnol. Oceanogr. 2015, 60, 1950–1964. [Google Scholar] [CrossRef]
- Pardo, I.; Delgado, C.; Abrain, R.; Gomez-Rodriguez, C.; Garcia-Rosello, E.; Garcia, L.; Reynoldson, T.B. A predictive diatom-based model to assess the ecological status of streams and rivers of Northern Spain. Ecol. Indic. 2018, 90, 519–528. [Google Scholar] [CrossRef]
- Frenette, J.J.; Thibeault, P.; Lapierre, J.F.; Hamilton, P.B. Presence of algae in freshwater ice cover of fluvial LAC Saint-Pierre (St. Lawrence river, Canada). J. Phycol. 2008, 44, 284–291. [Google Scholar] [CrossRef]
- Chen, X.; McGowan, S.; Bu, Z.J.; Yang, X.D.; Cao, Y.M.; Bai, X.; Zeng, L.H.; Liang, J.; Qiao, Q.L. Diatom-based water-table reconstruction in Sphagnum peatlands of northeastern China. Water Res. 2020, 174, 115648. [Google Scholar] [CrossRef] [PubMed]
- Becker, M.E.; Becker, T.J.; Bellucci, C.J. Diatom tolerance metrics to identify total phosphorus as candidate cause of aquatic life impairment in Connecticut, USA freshwater streams. Ecol. Indic. 2018, 93, 638–646. [Google Scholar] [CrossRef]
- Kim, H.K.; Cho, I.H.; Hwang, E.A.; Kim, Y.J.; Kim, B.H. Benthic Diatom Communities in Korean Estuaries: Species Appearances in Relation to Environmental Variables. Int. J. Environ. Res. Public Health 2019, 16, 2681. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Cao, J.X.; Pei, G.F.; Liu, G.X. Using benthic diatom assemblages to assess human impacts on streams across a rural to urban gradient. Environ. Sci. Pollut. Res. 2015, 22, 18093–18106. [Google Scholar] [CrossRef]
- Liu, Y.; Shen, J.M.; Chen, Z.L.; Ren, N.Q.; Li, Y.F. Distribution of polycyclic aromatic hydrocarbons in surface water and sediment near a drinking water reservoir in Northeastern China. Environ. Sci. Pollut. Res. 2013, 20, 2535–2545. [Google Scholar] [CrossRef]
- Chen, X.; Bu, Z.J.; Stevenson, M.A.; Cao, Y.M.; Zeng, L.H.; Qin, B. Variations in diatom communities at genus and species levels in peatlands (central China) linked to microhabitats and environmental factors. Sci. Total Environ. 2016, 568, 137–146. [Google Scholar] [CrossRef]
- Growns, I. Is genus or species identification of periphytic diatoms required to determine the impacts of river regulation? J. Appl. Phycol. 1999, 11, 273–283. [Google Scholar] [CrossRef]
- Xiao, W.; Huang, Y.; Mi, W.; Wu, H.; Bi, Y. Variation of Diatoms and Silicon in a Tributary of the Three Gorges Reservoir: Evidence of Interaction. Water 2019, 11, 1369. [Google Scholar] [CrossRef] [Green Version]
- Xue, H.; Zheng, B.; Meng, F.; Wang, Y.; Zhang, L.; Cheng, P. Assessment of Aquatic Ecosystem Health of the Wutong River Based on Benthic Diatoms. Water 2019, 11, 727. [Google Scholar] [CrossRef] [Green Version]
- Yin, W.J.; Teng, Y.G.; Zhai, Y.Z.; Hu, L.T.; Zhao, X.B.; Zhang, M.L. Suitability for developing riverside groundwater sources along Songhua River, Northeast China. Hum. Ecol. Risk Assess. 2018, 24, 2088–2100. [Google Scholar] [CrossRef]
- Kim, H.K.; Kwon, Y.S.; Kim, Y.J.; Kim, B.H. Distribution of Epilithic Diatoms in Estuaries of the Korean Peninsula in Relation to Environmental Variables. Water 2015, 7, 6702–6718. [Google Scholar] [CrossRef] [Green Version]
- Wang, M. Evaluate method and classification standard on lake eutrophication. Environ. Monit. China 2002, 18, 47–49. [Google Scholar]
- Tan, X.; Ma, P.M.; Bunn, S.E.; Zhang, Q.F. Development of a benthic diatom index of biotic integrity (BD-IBI) for ecosystem health assessment of human dominant subtropical rivers, China. J. Environ. Manag. 2015, 151, 286–294. [Google Scholar] [CrossRef]
- Lange-Bertalot, H.; Cavacini, P.; Tagliaventi, N.; Alfinito, S. Diatoms of Sardinia. Rare and 76 New Species in Rock Pools and Other Ephemeral Waters; Iconographia Diatomologica 12; A.R.G. Gantner Verlag, KG: Ruggell, Germany, 2003; pp. 1–438. [Google Scholar]
- Lange-Bertalot, H.; Hofmann, G.; Werum, M.; Cantonati, M. Freshwater Benthic Diatoms of Central Europe:Over 800 Common Species Used in Ecological Assessment; English edition with updated taxonomy and addedspecies; Koeltz Botanical Books: Schmitten, Oberreifenberg, Germany, 2017; pp. 1–942. [Google Scholar]
- Wojtal, A.Z. Species Composition and Distribution of Diatom Assemblages in Spring Waters from Various Geological Formations in Southern Poland; Bibliotheca Diatomologica 59; J. Cramer: Stuttgart, Germany, 2013; pp. 1–436. [Google Scholar]
- Lai, J. Canoco 5: A new version of an ecological multivariate data ordination program. Biodivers. Sci. 2013, 21, 765–768. [Google Scholar]
- Gang, H.X.; Li, R.H.; Zhao, Y.M.; Liu, G.F.; Chen, S.; Jiang, J. Loss of GLK1 transcription factor function reveals new insights in chlorophyll biosynthesis and chloroplast development. J. Exp. Bot. 2019, 70, 3125–3138. [Google Scholar] [CrossRef]
- Twiss, M.R.; McKay, R.M.L.; Bourbonniere, R.A.; Bullerjahn, G.S.; Carrick, H.J.; Smith, R.E.H.; Winter, J.G.; D’Souza, N.A.; Furey, P.C.; Lashaway, A.R.; et al. Diatoms abound in ice-covered Lake Erie: An investigation of offshore winter limnology in Lake Erie over the period 2007 to 2010. J. Great Lakes Res. 2012, 38, 18–30. [Google Scholar] [CrossRef]
- Lotter, A.F.; Bigler, C. Do diatoms in the Swiss Alps reflect the length of ice-cover? Aquat. Sci. 2000, 62, 125–141. [Google Scholar] [CrossRef] [Green Version]
- Pei, G.F.; Liu, G.X.; Hu, Z.Y. Variation of benthic diatoms in a shallow, eutrophic Chinese Lake, Lake Donghu. J. Freshw. Ecol. 2006, 21, 277–284. [Google Scholar] [CrossRef]
- Soares, M.C.S.; Huszar, V.L.M.; Roland, F. Phytoplankton dynamics in two tropical rivers with different degrees of human impact (southeast Brazil). River Res. Appl. 2007, 23, 698–714. [Google Scholar] [CrossRef]
- Bona, F.; La Morgia, V.; Falasco, E. Predicting river diatom removal after shear stress induced by ice melting. River Res. Appl. 2012, 28, 1289–1298. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.X.; Wang, Z.H.; Guo, X.; Gu, Y.G.; Liang, W.B.; Liu, L. Impacts of metal contamination and eutrophication on dinoflagellate cyst assemblages along the Guangdong coast of southern China. Mar. Pollut. Bull. 2017, 120, 239–249. [Google Scholar] [CrossRef]
- Agawin, N.S.R.; Duarte, C.M.; Agusti, S.; Vaque, D. Effect of N: P ratios on response of Mediterranean picophytoplankton to experimental nutrient inputs. Aquat. Microb. Ecol. 2004, 34, 57–67. [Google Scholar] [CrossRef] [Green Version]
- Wood, R.J.; Mitrovic, S.M.; Lim, R.P.; Warne, M.S.; Dunlop, J.; Kefford, B. Benthic diatoms as indicators of herbicide toxicity in rivers—A new SPEcies At Risk (SPEAR(herbicides)) index. Ecol. Indic. 2019, 99, 203–213. [Google Scholar] [CrossRef]
- Tan, X.; Zhang, Q.F.; Burford, M.A.; Sheldon, F.; Bunn, S.E. Benthic Diatom Based Indices for Water Quality Assessment in Two Subtropical Streams. Front. Microbiol. 2017, 8, 601. [Google Scholar] [CrossRef] [Green Version]
- Maslennikova, A.V. Development and application of an electrical conductivity transfer function, using diatoms from lakes in the Urals, Russia. J. Paleolimnol. 2020, 63, 129–146. [Google Scholar] [CrossRef]
- Tibby, J.; Gell, P.A.; Fluin, J.; Sluiter, I.R.K. Diatom-salinity relationships in wetlands: Assessing the influence of salinity variability on the development of inference models. Hydrobiologia 2007, 591, 207–218. [Google Scholar] [CrossRef]
- Pestryakova, L.A.; Herzschuh, U.; Gorodnichev, R.; Wetterich, S. The sensitivity of diatom taxa from Yakutian lakes (north-eastern Siberia) to electrical conductivity and other environmental variables. Polar Res. 2018, 37, 1485625. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.; Lee, H.; Kim, Y.K.; Sohn, K.; Lee, K. Hydrogen peroxide interference in chemical oxygen demand during ozone based advanced oxidation of anaerobically digested livestock wastewater. Int. J. Environ. Sci. Technol. 2011, 8, 381–388. [Google Scholar] [CrossRef] [Green Version]
- Zubaidah, T.; Karnaningroem, N.; Slamet, A. The Self-Purification Ability in the Rivers of Banjarmasin, Indonesia. J. Ecol. Eng. 2019, 20, 177–182. [Google Scholar] [CrossRef]
- Xiao, C.G.; Chen, J.; Chen, D.; Chen, R.D. Effects of river sinuosity on the self-purification capacity of the Shiwuli River, China. Water Sci. Technol. Water Supply 2019, 19, 1152–1159. [Google Scholar] [CrossRef]
- Wiese, R.; Renaudie, J.; Lazarus, D.B. Testing the accuracy of genus-level data to predict species diversity in Cenozoic marine diatoms. Geology 2016, 44, 1051–1054. [Google Scholar] [CrossRef]
- Lazarus, D.; Barron, J.; Renaudie, J.; Diver, P.; Turke, A. Cenozoic Planktonic Marine Diatom Diversity and Correlation to Climate Change. PLoS ONE 2014, 9, e84857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
The location of sampling stations in Lalin River Basin.

Figure 2.
Hierarchical clustering based on diatom taxonomic composition: (a) based on species-level identification, and (b) based on in genus-level identification.
Figure 2.
Hierarchical clustering based on diatom taxonomic composition: (a) based on species-level identification, and (b) based on in genus-level identification.

Figure 3.
Venn diagram of the richness of diatom at species-level (a) and genus-level (b).

Figure 4.
The composition of diatom in LLRB at genus-level (a) and species-level (b) during ice-covered period.
Figure 4.
The composition of diatom in LLRB at genus-level (a) and species-level (b) during ice-covered period.

Figure 5.
Redundancy analysis (RDA) ordinations of diatom species level (a), genus level (b) and sampling station with 9 environmental factors.
Figure 5.
Redundancy analysis (RDA) ordinations of diatom species level (a), genus level (b) and sampling station with 9 environmental factors.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Environmental factors in Lalin River Basin (LLRB) during ice-covered period in 2018.
| Stations | WT (°C) | DO (mg/L) | EC (μS/cm) | pH | ORP (mV) | Tur. (NTU) | Chl-a (μg/L) | TN (mg/L) | TP (mg/L) | COD (mg/L) |
|---|---|---|---|---|---|---|---|---|---|---|
| S1 | 1.44 | 1.55 | 54.47 | 8.57 | 405 | 27.37 | 0.58 | 3.60 | 0.06 | 5.67 |
| S2 | 0.16 | 2.20 | 61.73 | 8.23 | 404 | 26.77 | 0.37 | 3.63 | 0.05 | 7.33 |
| S3 | 3.54 | 1.70 | 164.5 | 8.38 | 394 | 26.53 | 1.71 | 5.00 | 0.22 | 10.67 |
| S4 | 1.44 | 1.72 | 168.2 | 7.66 | 409 | 27.1 | 1.17 | 4.93 | 0.29 | 17.00 |
| S5 | 0.97 | 0.46 | 194.6 | 8.17 | 406 | 27.57 | 1.22 | 3.37 | 0.44 | 16.67 |
| S6 | 0.06 | 1.94 | 263.5 | 9.27 | 410 | 23.87 | 2.52 | 5.03 | 1.67 | 72.00 |
| S7 | 0.49 | 1.54 | 267.63 | 8.42 | 415 | 27.93 | 6.51 | 8.03 | 1.77 | 83.33 |
| S8 | 0.49 | 2.09 | 270.47 | 8.44 | 436 | 26.57 | 0.83 | 2.07 | 0.58 | 20.67 |
| S9 | 0.03 | 1.43 | 34.07 | 9.00 | 449 | 24.67 | 1.09 | 1.30 | 0.34 | 67.67 |
| T-test | ||||||||||
| Group-1 × Group-2 | p > 0.05 | p > 0.05 | p > 0.05 | p > 0.05 | p > 0.05 | p > 0.05 | p > 0.05 | p > 0.05 | p < 0.05 | p > 0.05 |
| Group-1 × Group-3 | p > 0.05 | p > 0.05 | p < 0.05 | p > 0.05 | p > 0.05 | p > 0.05 | p > 0.05 | p > 0.05 | p < 0.05 | p < 0.05 |
| Group-2 × Group-3 | p > 0.05 | p > 0.05 | p > 0.05 | p > 0.05 | p > 0.05 | p > 0.05 | p > 0.05 | p > 0.05 | p < 0.05 | p < 0.05 |
Table 2.
Comprehensive trophic index (TSI) of LLRB during ice-covered period in 2018.
| TSI (Chl-a) | TSI (TN) | TSI (TP) | TSI (COD) | TSI (∑) | Trophic Status | |
|---|---|---|---|---|---|---|
| S1 | 19 | 76 | 49 | 47 | 48 | mesotrophic |
| S2 | 14 | 76 | 46 | 54 | 48 | mesotrophic |
| S3 | 31 | 82 | 70 | 64 | 62 | middle eutrophic |
| S4 | 27 | 82 | 74 | 76 | 65 | middle eutrophic |
| S5 | 27 | 75 | 81 | 76 | 65 | middle eutrophic |
| S6 | 35 | 82 | 103 | 115 | 84 | high eutrophic |
| S7 | 45 | 90 | 104 | 119 | 89 | high eutrophic |
| S8 | 23 | 67 | 85 | 82 | 64 | middle eutrophic |
| S9 | 26 | 59 | 77 | 113 | 69 | middle eutrophic |
Table 3.
Summary statistics of Redundancy analysis at species level and genus level.
| Environmental Factors | r2 | p | ||||
|---|---|---|---|---|---|---|
| Species-Level | Genus-Level | Species-Level | Genus-Level | |||
| Axis-1 | Axis-2 | Axis-1 | Axis-2 | |||
| WT | −0.601 | 0.104 | 0.575 | 0.111 | 0.260 | 0.168 |
| DO | −0.127 | −0.472 | −0.075 | 0.112 | 0.422 | 0.912 |
| EC | 0.549 | 0.130 | −0.674 | −0.037 | 0.278 | 0.038 |
| pH | 0.449 | −0.340 | −0.509 | −0.419 | 0.232 | 0.204 |
| ORP | 0.306 | −0.482 | −0.253 | 0.257 | 0.298 | 0.816 |
| Tur. | −0.260 | 0.304 | 0.448 | 0.416 | 0.362 | 0.150 |
| Chl-a | 0.455 | −0.281 | −0.576 | 0.133 | 0.540 | 0.220 |
| TN | 0.020 | 0.175 | 0.167 | 0.102 | 0.954 | 0.832 |
| TP | 0.674 | −0.190 | −0.724 | −0.004 | 0.004 | 0.030 |
| COD | 0.638 | −0.406 | −0.793 | 0.053 | 0.076 | 0.002 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lu, X.; Liu, Y.; Fan, Y. Diatom Taxonomic Composition as a Biological Indicator of the Ecological Health and Status of a River Basin under Agricultural Influence. Water 2020, 12, 2067. https://0-doi-org.brum.beds.ac.uk/10.3390/w12072067
AMA Style
Lu X, Liu Y, Fan Y. Diatom Taxonomic Composition as a Biological Indicator of the Ecological Health and Status of a River Basin under Agricultural Influence. Water. 2020; 12(7):2067. https://0-doi-org.brum.beds.ac.uk/10.3390/w12072067
Chicago/Turabian StyleLu, Xinxin, Yan Liu, and Yawen Fan. 2020. "Diatom Taxonomic Composition as a Biological Indicator of the Ecological Health and Status of a River Basin under Agricultural Influence" Water 12, no. 7: 2067. https://0-doi-org.brum.beds.ac.uk/10.3390/w12072067
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.