
Amphibian Metacommunity Responses to Agricultural Intensification in a Mediterranean Landscape


1
Área de Ecología, Departamento de Biodiversidad y Gestión Ambiental, Facultad de Ciencias Biológicas, Callejón Campus Vegazana s/n, Universidad de León (ULE), 24071 León, Spain
2
Departamento de Biodiversidad y Biología Evolutiva, Museo Nacional de Ciencias Naturales (MNCN-CSIC), c/ José Gutiérrez Abascal, 28006 Madrid, Spain
3
Departamento de Biología Animal (Zoología), Universidad de Salamanca, 37007 Salamanca, Spain
*
Author to whom correspondence should be addressed.
Land 2021, 10(9), 924; https://0-doi-org.brum.beds.ac.uk/10.3390/land10090924
Submission received: 31 July 2021
/
Revised: 28 August 2021
/
Accepted: 29 August 2021
/
Published: 2 September 2021
(This article belongs to the Special Issue Wildlife Protection and Habitat Management: Practice and Perspectives)
Abstract
:Agricultural intensification has been associated with biodiversity declines, habitat fragmentation and loss in a number of organisms. Given the prevalence of this process, there is a need for studies clarifying the effects of changes in agricultural practices on local biological communities; for instance, the transformation of traditional rainfed agriculture into intensively irrigated agriculture. We focused on pond-breeding amphibians as model organisms to assess the ecological effects of agricultural intensification because they are sensitive to changes in habitat quality at both local and landscape scales. We applied a metacommunity approach to characterize amphibian communities breeding in a network of ponds embedded in a terrestrial habitat matrix that was partly converted from rainfed crops to intensive irrigated agriculture in the 1990s. Specifically, we compared alpha and beta diversity, species occupancy and abundance, and metacommunity structure between irrigated and rainfed areas. We found strong differences in patterns of species occurrence, community structure and pairwise beta diversity between agricultural management groups, with a marked community structure in rainfed ponds associated with local features and the presence of some rare species that were nearly absent in the irrigated area, which was characterized by a random community structure. Natural vegetation cover at the landscape scale, significantly lower on the irrigated area, was an important predictor of species occurrences. Our results suggest that maintaining both local and landscape heterogeneity is key to preserving diverse amphibian communities in Mediterranean agricultural landscapes.
1. Introduction
Biodiversity loss due to anthropogenic activities is a global concern, which calls for practices favoring coexistence between human needs and the viability of natural communities. In the current sixth mass extinction [1,2], we need to understand how human activities affect biodiversity and find ways to mitigate their negative effects [3]. One of the main threats to biodiversity is the increasing replacement of natural habitats with human settlements and farmland [4]. The amount of land cover devoted to agricultural practices has increased for centuries in a continuous trend, and after the “green revolution” of the 20th century, higher crop yields were achieved with the use of heavy machinery, agrochemicals, irrigation and a shift to extensive monocultures [4,5,6]. This intensified agriculture has been associated with biodiversity declines and deleterious effects for farmland animal and plant communities [7,8,9,10].
Traditional extensive agriculture, characterized by rainfed (supplied primarily with rainfall) heterogeneous crops in smaller plot sizes, is usually associated with landscape heterogeneity. The terrestrial habitat matrix includes patches of natural vegetation separating different crops and can potentially host more species than homogeneous habitats [11,12]. The transformation of traditional extensive agriculture to intensive (irrigated) agriculture implies the replacement of patches of natural habitats by crops and an increasing isolation of the remaining patches. The effect of this process of landscape homogenization on farmland biodiversity is an important research topic with applied implications [13,14,15].
Water bodies, such as ponds, represent key ecosystems in agricultural landscapes [16], hosting sets of biological communities linked by the dispersal of multiple, potentially interacting species (i.e., metacommunities [17]). Metacommunity ecology is a powerful framework to assess the impact of human activities on entire biotic assemblages, explicitly addressing the role of factors acting at different spatial scales, like environmental conditions, predation, competition, or dispersal [17]. Most metacommunity studies have focused on discrete water bodies that are considered local populations, especially in temperate and cold regions [18,19]. Mediterranean ponds in agricultural landscapes have been comparatively less studied [20] (but see [21,22]). Mediterranean ponds typically differ from the comparatively stable environments of temperate ponds in their marked seasonality, with strong and largely unpredictable changes in hydroperiod across seasons and years [23,24]. From a metacommunity perspective, this unpredictability can result in stronger environmental sorting (sensu [17]) in Mediterranean communities, with increasing probability of local extinction events following extended dry periods.
Since water is a major limiting factor in Mediterranean agro-ecosystems, irrigation is often used to achieve higher crop yields, and is in fact one of the main drivers of agricultural intensification in the region [25]. Negative effects of the conversion of rainfed to irrigated agriculture on biodiversity have been reported in terrestrial organisms like farmland and steppe birds [26,27] and insects [28], but few studies have focused on aquatic or semi-aquatic organisms, such as amphibians. Amphibians are key components of Mediterranean biotas, with many regional endemics (N = 68) [29], but threats like habitat destruction, alteration and fragmentation are causing population declines and extinctions [30,31]. However, artificial habitats can be used successfully by some species [32,33]. Most community studies of Mediterranean pond-breeding amphibians have been carried out in natural rather than in agricultural areas and have focused mainly on the influence of local wetland features on individual species [34,35]. Negative effects of irrigation, mediated by the increased use of fertilizers and pesticides alongside strong changes in landscape transformation, have been described in biotic communities associated with Mediterranean ponds [36]. However, higher humidity as a consequence of irrigation, and the construction of water channels could also have a positive effect, creating humid terrestrial habitats and improving connectivity [37,38].
Therefore, our knowledge about the effects of agricultural management practices on Mediterranean amphibian communities is incomplete. Further studies are needed to better characterize the effects of the transformation of rainfed into irrigated agriculture on amphibians and other associated biotic communities. Investigating the effects of agricultural intensification and irrigation on amphibian communities is required to identify and mitigate the negative effects of particular management regimes and to provide guidelines for the conservation of diverse amphibian communities in agricultural areas.
We used a metacommunity approach to compare the amphibian communities of two contiguous agricultural areas in northern Iberia. Both areas were managed as rainfed crops (cereal) until the 1990s, when part of the area was transformed into intensively managed irrigated crops (corn), and have thus been subject to different agricultural management regimes for several amphibian generations. We explore the effects of this process of agricultural intensification on local amphibian community structure, beta diversity and patterns of species occurrence and abundance. Our expectation is that amphibian communities in the irrigated area will be altered as a result of habitat homogenization at the landscape scale, and this will be reflected in differences in patterns of amphibian occupancy, abundance and community structure across differentially managed areas. Our a priori hypothesis is that irrigated ponds will host less diverse amphibian communities with an altered community structure.
2. Materials and Methods
2.1. Study Area
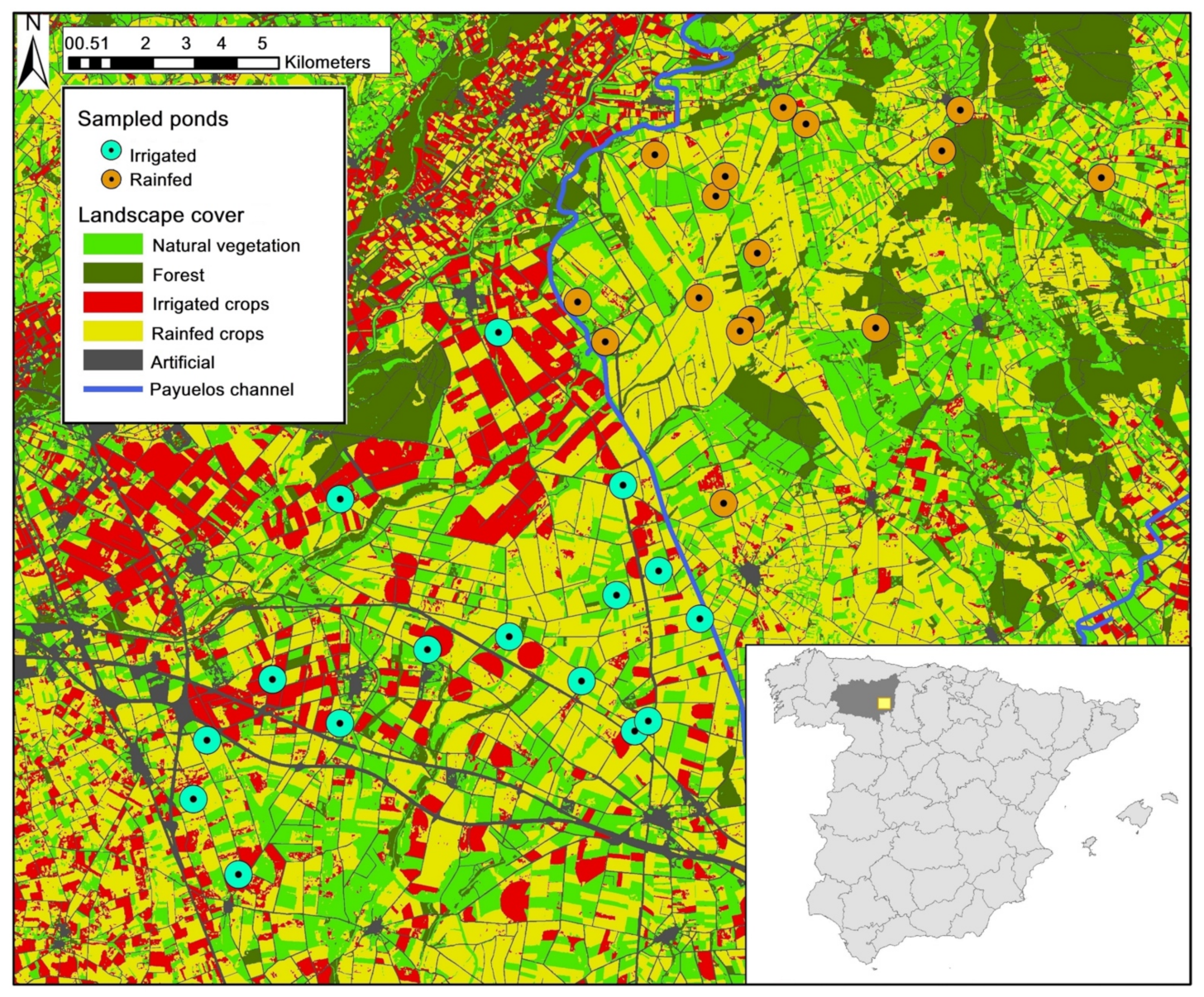
The study area is located in SE León Province (Castilla y León, Spain), in the Iberian North Plateau, near the Esla river, part of the Duero basin (Figure 1). The climate is Mediterranean with continental influence, with cold winters and dry and hot summers. Rainfall concentrates in the spring and autumn. Until the mid-1990s, the landscape was dominated by a traditional agro-ecosystem, a mixture of rainfed crops (mostly cereal), oak forest patches (Quercus pyrenaica) and meadows used for pasture. A considerable number of natural ponds occur in this area, favored by local lithological features, mostly comprising thin post-Tertiary deposits from the nearby Cantabrian Mountains [39]. All sampled ponds are located in a formerly homogeneous area, part of which was transformed into intensive irrigated agriculture around 1998, following the construction of the Payuelos irrigation channel. Studies carried out in 1994 and 1995, a few years before the shift to irrigation, showed that all ponds were homogeneous in their hydrochemistry [40] and in their aquatic macrophyte and invertebrate communities [41], with eutrophication and hydroperiod mainly shaping biotic differences among ponds. This is the first study about amphibian communities in this region.
We selected two groups of 16 ponds with similar characteristics in each agricultural management regime; that is, 16 ponds in the rainfed area and 16 in the irrigated area (Table S1 in Supplementary Materials, Figure 1). These ponds are mostly shallow and temporary, drying up during the summer months (July to September). Mean pond area was 8551 m2 (range: 416–42,201 m2), and mean depth was 74 cm (range: 35–140 cm). Only natural ponds were sampled; other wetlands, such as excavated ponds, inundated gravel pits and irrigation reservoirs, were excluded from analyses. Surveys in these artificial sites and in other natural ponds around our study area show they share a common amphibian species pool with ponds in our study area (unpublished data).
2.2. Amphibian Sampling
Amphibians were sampled at night, during their breeding period (February to May, 2019). Each pond was sampled monthly during this period, resulting in four visits per pond, covering the breeding phenology of all species expected to occur in the area. We selected nights with optimal conditions for amphibian activity (temperature > 5 °C, humidity > 70% or raining) for sampling. Sampling took place from sunset until 4–5 h later. On each visit, we first conducted an acoustic monitoring survey for five to ten minutes, with no artificial lights, in order to detect and, if possible, quantify all male anurans of the different species calling. Then, we surveyed through the entire surface of each pond, except in the largest pond (>40,000 m2), where we surveyed a 5 m band along the full perimeter of the pond, where most breeding individuals concentrate. Amphibians were located directly with a flashlight or through their eye shine. For capture, when necessary, we used a dip-net.
We recorded amphibian species as “present” in a sampled pond when we were able to confirm reproduction of that species in the pond. In the case of newts, reproduction was confirmed with the observation of adults in their aquatic phase, larvae or eggs, and in anurans, by the observation of adults with nuptial pads, calling males, amplectant pairs, larvae or eggs. Individual counts were performed for all species except for Pelophylax perezi, due to the high abundance of this aquatic frog in some ponds. As a surrogate of amphibian abundance in each pond we took the maximum count across all visits, considering only adults. Most Iberian amphibians are explosive breeders, so the maximum number of individuals recorded in a pond is expected to reflect the breeding peak of each species, thus representing a good surrogate for population size. We standardized this value across all ponds relative to pond area, obtaining a general value of abundance for each species and pond in units of individuals/1000 m2.
2.3. Environmental Variables
Following the ecological metacommunity approach [42], we considered environmental predictors at the local and landscape scales for the study of amphibian community patterns on both agricultural management groups. At the local scale, we selected variables affecting amphibian dynamics in temperate regions as reported in previous studies [43,44,45,46], including pond area (m2), medium (average across visits) and maximum depth (cm), and helophyte vegetation cover (%). Pond area and helophyte cover were measured directly from satellite images (the more recent images available from the Spanish “Centro Nacional de Información Geográfica”, CNIG) using ArcGIS 10.7 (®ESRI). Pond depth was recorded on each visit using a graduated stick, at the point of maximum depth detected. Landscape variables were considered at two spatial scales, 500 m and 1000 m from each pond, accounting for recorded dispersal distances of amphibian species in the area [47,48]. We recorded the following landscape types: natural forest, forest plantation, grassland, scrub, bare soil, wetlands, irrigated crops and non-irrigated (i.e., rainfed) crops. The importance of each landscape type on a pond was calculated as the proportion cover (%) of a circular buffer of 500 m or 1000 m radio centered in the pond centroid. Land cover data was extracted from the database of “Mapa de Cultivos y Superficies Naturales de Castilla y León” (MCSNCyL, Junta de Castilla y León). Proportion cover was calculated with ArcGIS.
For subsequent analyses, environmental variables were filtered to account for multicollinearity using Pearson’s pairwise correlation as calculated with R package stats [49] (Table S2). We found high correlation between all landscape variables at the 500 and 1000 m spatial scales (Pearson’s Correlation Coefficient, r > 0.7), so we retained only one value for each landscape spatial category, selecting the one less correlated on average with the rest of environmental predictors. Retained variables included natural forest, artificial surfaces, bare soil, scrub, irrigated and non-irrigated crops for the 500 m scale, and plantation forest, grassland and wetland cover for the 1000 m scale (Table S3). We also found a high correlation between maximum and medium pond depth (r = 0.93) so we retained only medium depth as the predictor less correlated with the remaining variables.
2.4. Statistical Analysis
In a first level of inference, we tested for differences in species richness, abundance and patterns of co-occurrence between the amphibian communities of the two agricultural management groups. We first tested for environmental differences using local and landscape environmental variables. Levene’s normality test was significant for almost all variables, so we used the non-parametric Mann–Whitney test. To compare alpha diversity (richness) and species abundance estimates between ponds in the two management groups we used an Analysis of Similarities (ANOSIM). If significant, we looked for the species with the highest contribution to inter-group differences using a Similarity Percentages (SIMPER) test with 9999 permutations. Finally, we tested for differences in species by species paired occurrences in irrigated and rainfed ponds using Fisher’s exact test. All analyses were performed with packages stats and vegan in R.
For the second level of inference, we quantified differences in community composition among and within agricultural management groups. We calculated beta diversity following the additive partition approach [50], which decomposes total beta diversity (calculated by the Sørensen index, βsor) in two components explaining different measures of inter-site differences: spatial turnover (measured by Simpson’s index, βsim) and nestedness (βnes). Differences due to turnover are caused by the replacement of some species by others between two sites, and nestedness occurs when the species composition of a community is a subset of that of another richer community. We used presence/absence matrices to calculate the three components of beta diversity for all ponds and separately for each group using the function ‘beta.multi’ implemented in R package betapart [51]. Then, we calculated the three beta components for each pair of ponds within the groups using the function ‘beta.pair’ in betapart to test for differences in beta diversity between irrigated and rainfed ponds. Pairwise approaches have been used in many ecological comparisons of multi-site data because they provide a good surrogate of average differences between paired samples [52,53]. Pairwise values were pooled for each group using the function ‘dist2list’ in R library spaa [54]. Since pairwise beta value distributions did not fit a normal distribution, we used Mann–Whitney’s test to assess differences between irrigated and rainfed amphibian communities in all three beta diversity components.
The third inference level focused on differences in amphibian community structure between ponds and agricultural management groups. We followed the “Elements of Metacommunity Structure” (EMS) approach [55], which uses analyses of coherence, turnover and boundary clumping to distinguish several idealized metacommunity patterns (Evenly Spaced, Gleasonian, Clementsian, Nested, Checkerboard and Random). Prior to this analysis, species by site matrices were ordered by reciprocal averaging following the main axis of community variation. Coherence is assessed by counting the number of embedded absences, and significance is assessed by comparing against a null distribution created by random reordering of the community matrix. Then, turnover is calculated with the number of replacements between species among sites and Morisita’s Index is used for the evaluation of Boundary Clumping [56]. The distinct metacommunity structure patterns represent a gradient from metacommunities driven by environmental species sorting (Clementsian, Checkerboard) to other patterns driven by species dispersal (Nested) or intermediate situations (Evenly Spaced, Gleasonian, Random). This analysis was performed with package metacom in R [57].
Finally, we explored the main drivers of individual species preferences using both an ordination analysis via redundancy analysis (RDA) and a binomial regression analysis, with species composition per site as the response variable and all environmental predictors as factors. For the regression analysis we summarized environmental variables via Principal Component Analysis (PCA), which reduced the set of predictor variables to six principal components reflecting the main gradients of environmental variation among ponds (Table S4). Then, we conducted logistic regression analyses with these principal components as the new environmental variables. Using this approach decreases the complexity of fitting regression models [37] and also reduces problems of perfect separation (which could be detected and thus excluded from the final models). Model selection was based on Akaike’s Information Criterion (AIC) using a stepwise procedure. Analyses were performed with package vegan in R. For the RDA analyses we used software Canoco 4.5 CanoDraw4 to analyze and plot results [58].
3. Results
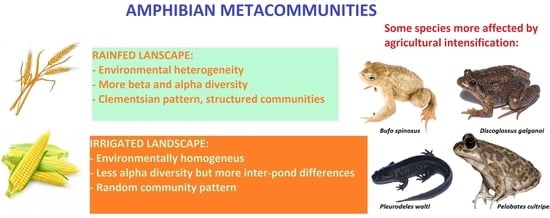
The environmental comparison between ponds in the two agricultural areas showed significant differences for many landscape variables, but not for the local ones (Table S3). The overall landscape was dominated by grasslands (≈20% cover), and rainfed crops (>40%). Natural vegetation cover, excluding grasslands, was significantly higher in the traditionally managed landscape (>12% versus <1%), with more proportion of natural and planted forest, bare ground and scrub. As expected, the landscape surrounding ponds in the irrigated area showed a significantly higher proportion cover of irrigated crops and artificial surfaces. Wetland cover, reflecting pond density, was not different between both agricultural areas. The irrigated area was mainly characterized by a greater diversity of crops, both irrigated and rainfed, but with less proportion of natural cover (forest and pastures). Therefore, the rainfed area has a greater level of landscape heterogeneity, with a mixture of crops and natural areas, whereas the irrigated area is more homogenous, containing mostly crops.
We found eight amphibian species in the study area: marbled newt (Triturus marmoratus), Iberian ribbed newt (Pleurodeles waltl), western spadefoot toad (Pelobates cultripes), Iberian painted frog (Discoglossus galganoi), spiny toad (Bufo spinosus), natterjack toad (Epidalea calamita), Iberian treefrog (Hyla molleri) and Iberian green frog (Pelophylax perezi). All species were found at least once in both irrigated and rainfed areas except for the painted frog, which was only found in five ponds in the rainfed area. All species were more frequent in the rainfed area, except for the marbled newt and natterjack toad, which occurred in the same number of ponds in the two areas. Fisher’s test showed significant differences in the occurrences of P. cultripes (p = 0.015) and D. galganoi (p = 0.043) between the two pond groups, with both species being more frequent in the rainfed area (Table S5).
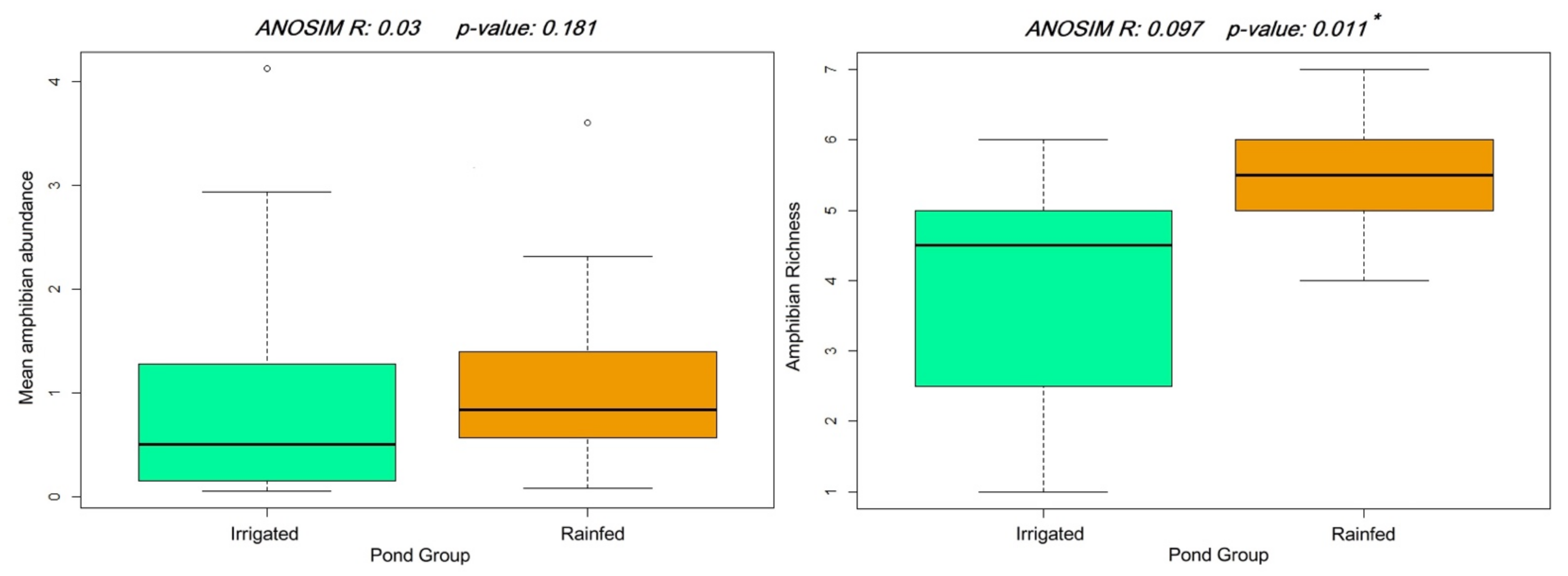
Species richness ranged from one to seven amphibian species across ponds. Mean species richness by pond also differed between the two areas, with ponds in the rainfed area having significantly more amphibian species on average (ANOSIM R: 0.097, p < 0.02; Figure 2). The SIMPER analysis showed P. cultripes, E. calamita and P. waltl to be responsible for more than 50% of inter-group differences in species richness (Table S6). Overall, amphibian abundance was more similar between agricultural management groups than species richness (Figure 2), and abundance estimates for the different species were not significantly different between irrigated and rainfed ponds (Table S7), although P. cultripes and P. waltl were considerably more abundant in rainfed ponds (>40% more).
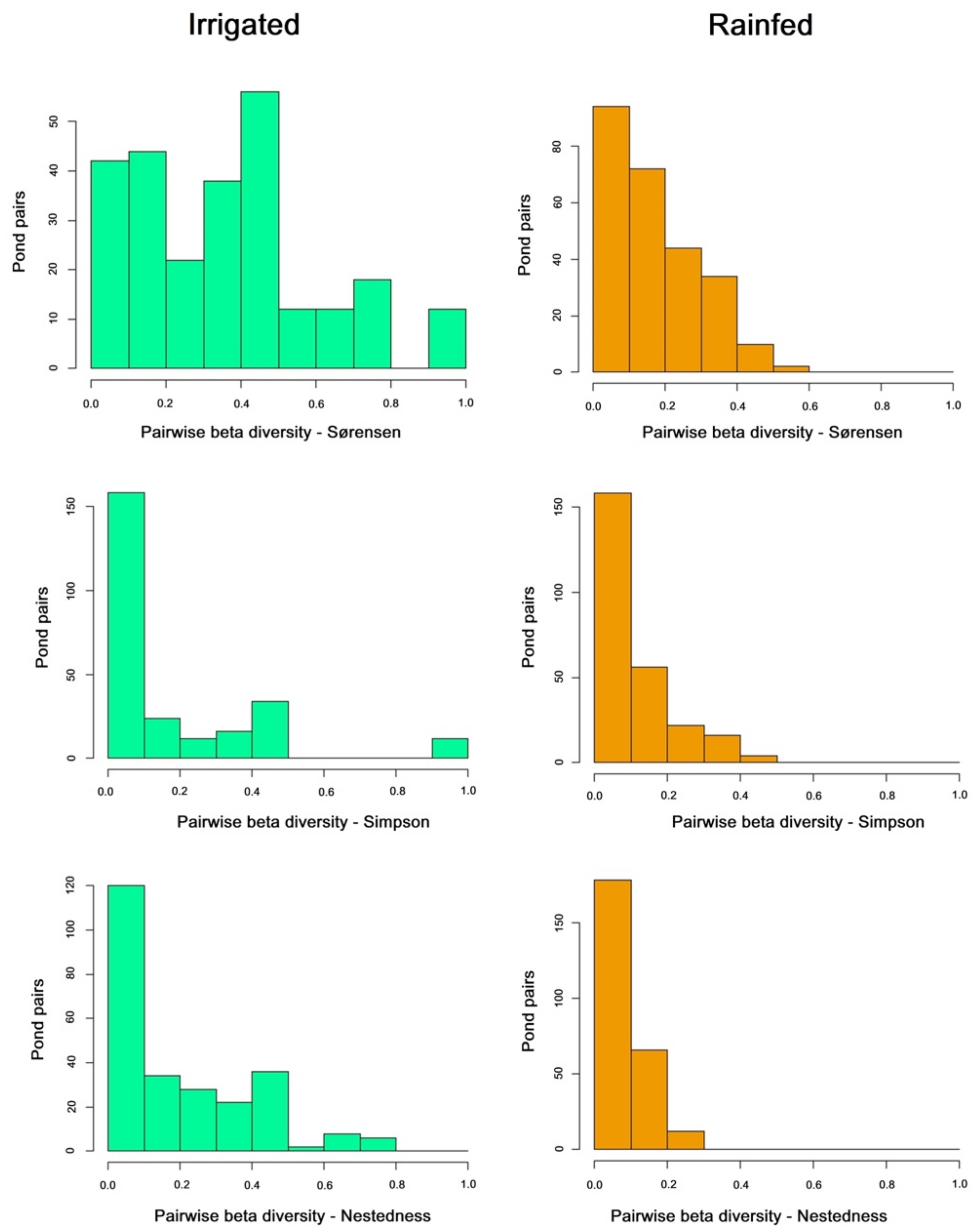
Partition of beta diversity (Table 1) showed higher total beta diversity in rainfed ponds, with higher values of the nestedness and turnover components in comparison with the irrigated area. Turnover was more important than nestedness in both areas, but the partition also showed stronger differences between the two components for irrigated than for rainfed ponds. Rainfed ponds hosted more amphibian species on average, with some of them being present in only a few ponds (E. calamita, B. spinosus, D. galganoi). Thus, they are expected to have higher total beta diversity values than irrigated ponds, which have fewer species and similar occurrences.
The density distribution of pairwise values is shown in Figure 3. Beta diversity among all ponds was higher in the rainfed area, but pairwise beta diversity reached significantly higher values for irrigated ponds both in total beta diversity and in the nestedness component (Table 1), highlighting the existence of some pairs of well-differentiated ponds in their community composition (Figure 3). Based on pairwise comparisons, mean nestedness was higher than turnover in the irrigated group. Rainfed ponds showed less difference between nestedness and turnover beta diversity on the mean pairwise values than on the overall beta value. In spite of the major importance of turnover for total beta diversity, pairwise measures showed some pairs of ponds with high nestedness in the irrigated group, with these nestedness pairwise values being significantly higher than in the rainfed group (Figure 3, Table 1). Thus, in the irrigated group there are some species-poor ponds whose species composition is a subset of that found in other ponds.
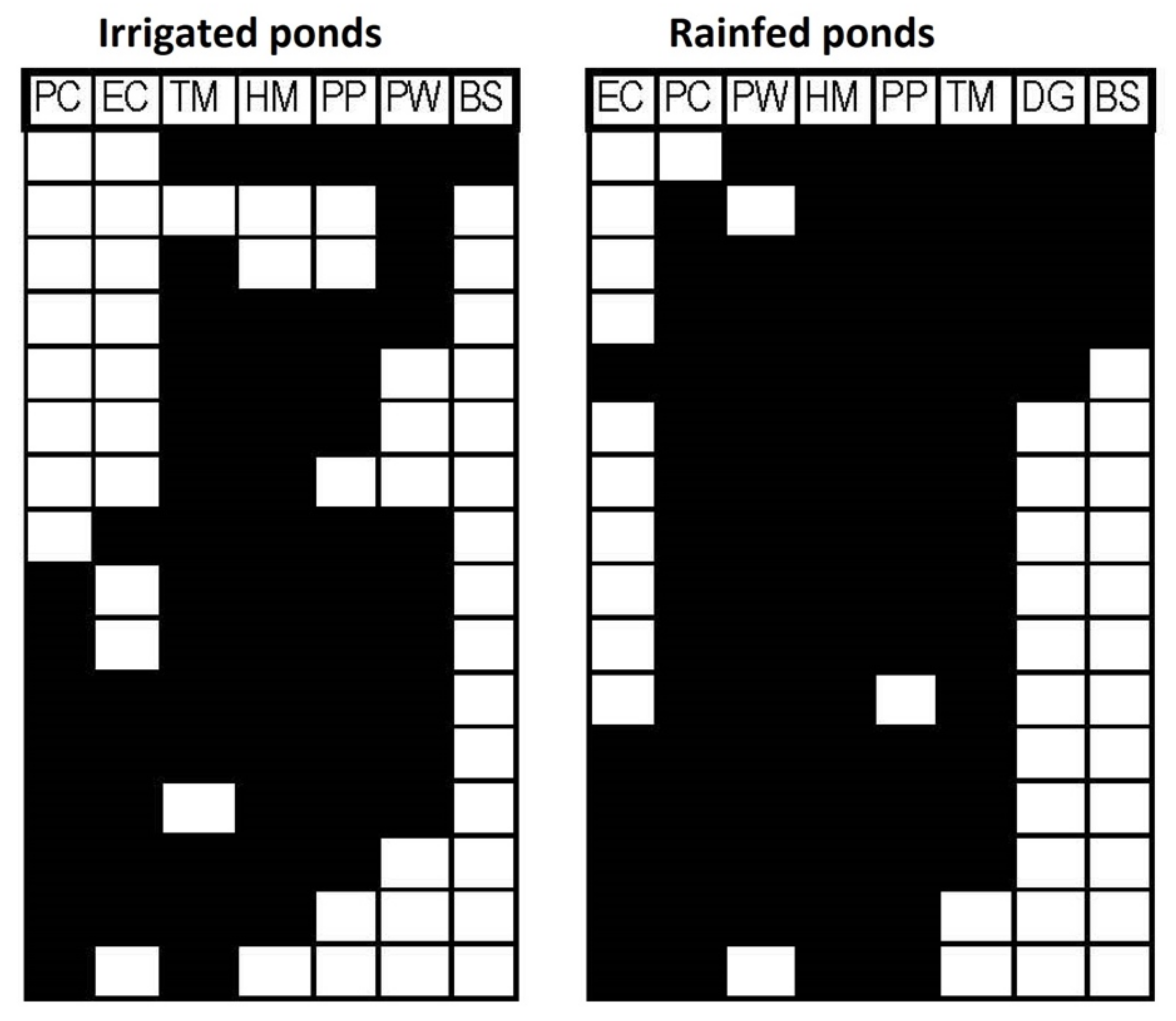
The results of the EMS analysis are summarized in Table 2. EMS showed a random pattern for the community structure of ponds in the irrigated area and also when ponds of the two areas were considered jointly. Rainfed ponds showed a significant coherence pattern and a Clementsian-type community, characterized by positive turnover and significant boundary clumping. Species by site matrices ordered by reciprocal averaging for each management group are shown in Figure 4.
Regarding the environmental preferences of individual species, regression analyses (Table S8) mostly showed weak and non-significant responses of amphibian species to the main gradients of environmental variation. Discoglossus galganoi and P. cultripes showed preference for large ponds in heterogeneous landscapes with low irrigation, in accordance with their lower abundance and occurrence in the irrigated area.
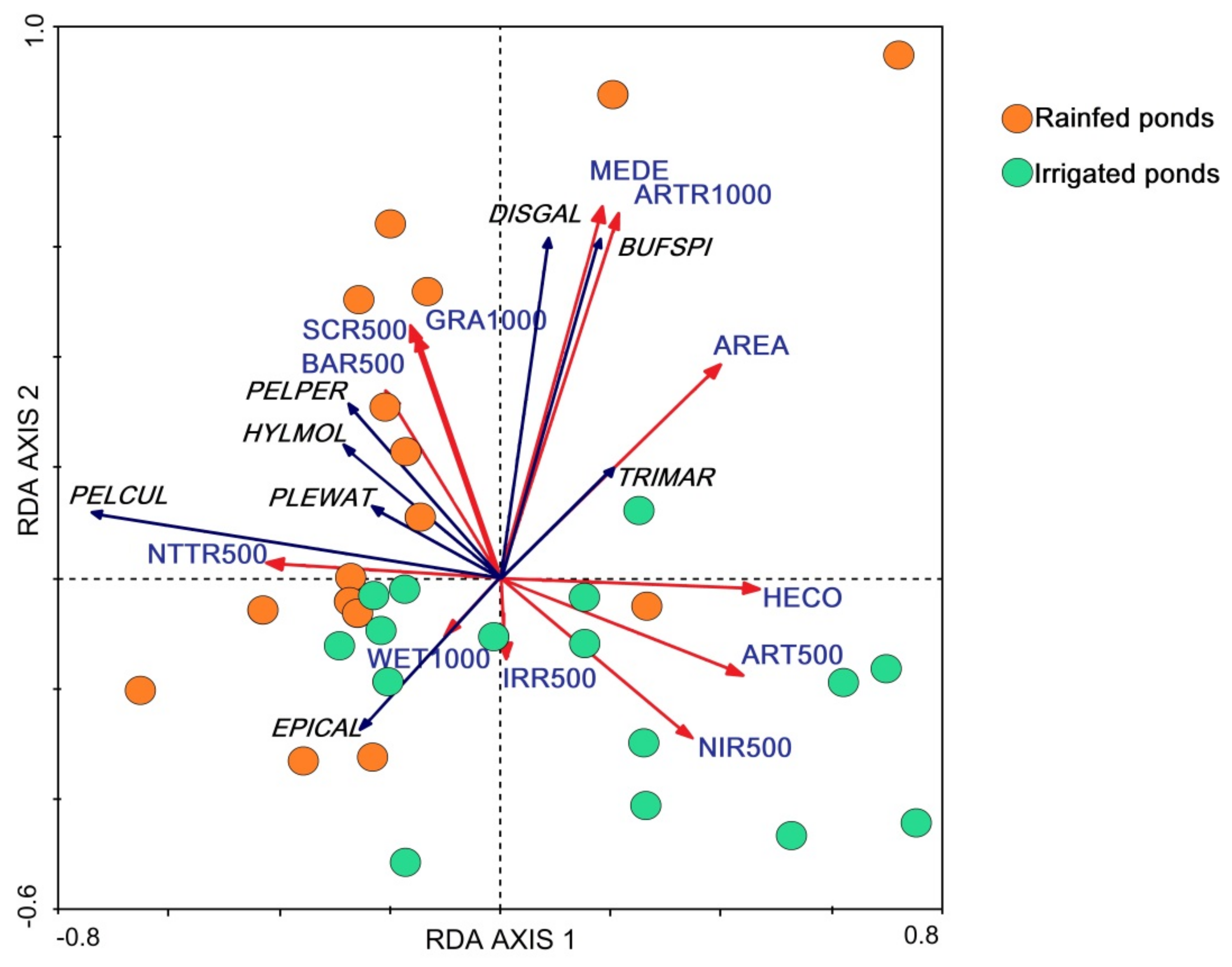
On the other hand, the RDA analysis combining landscape and local variables showed clear differences between rainfed and irrigated ponds (Figure 5). The main two axes of the redundancy analysis explained up to 61.9% of amphibian community variation, with similar importance (Axis 1: 32.6%; Axis 2: 29.3%). Both axes were driven by a combination of local and landscape features and thus represent landscape heterogeneity. Axis 1 can be interpreted as a gradient from vegetated ponds in intensive agricultural sites (high axis values), to more insolated ponds in heterogeneous landscapes with a greater proportion of natural forest. Axis 2 mostly represents pond size and landscape heterogeneity, with negative values correlating with the amount of irrigated and non-irrigated crops. Overall, the RDA plot (Figure 5) shows separation between ponds in the two agricultural management regimes, with landscape variables being most discriminative. Irrigated ponds are associated with high scores on axis 1 and markedly low scores on axis 2 (corresponding to smaller ponds in agricultural landscapes).
Species responses were in general positively correlated with natural vegetation and pond size, except for E. calamita, which preferred smaller ponds close to other wetlands (Figure 5). RDA also offers an environmental explanation for the community turnover pattern among rainfed ponds. The responses to environmental variables were similar for B. spinosus and D. galganoi, both showing a strong preference for large and deep ponds. On the other hand, the presence of E. calamita decreased with pond area and depth and increased with the vicinity of wetlands. The presence of P. cultripes was highly correlated with natural forests and low agricultural cover at the landscape scale, preferring insolated ponds (with low helophyte cover). Triturus marmoratus was more frequent in large ponds with aquatic vegetation. Finally, the responses of P. perezi, H. molleri and P. waltl were similar and positively correlated with the presence of natural vegetation at the landscape scale and negatively affected by both types of croplands.
RDA found no clear differences in the effects of irrigated and rainfed crops on amphibian presence. Both types of crops showed negative effects on all species, except on E. calamita, which seems to be slightly favored by irrigated crops. The effect of rainfed crop cover on species occurrences seems to be stronger than that of irrigated crops, but this could be a consequence of the higher abundance of rainfed crops in the study area. The species which seemed more negatively affected by crop cover were also the scarcest in the intensively managed area: D. galganoi and B. spinosus. Other species, like H. molleri, P. perezi, P. cultripes, and P. waltl were also negatively correlated with crop cover, as the amount of crop cover was also correlated with lower cover of natural vegetation.
4. Discussion
Our analyses showed significant differences between the two agricultural management groups in the terrestrial landscape matrix surrounding sampled ponds, and in their associated amphibian communities, which differed in structure and distribution patterns. These differences are linked to the effect of landscape (e.g., higher natural vegetation in rainfed area), but not local variables across management groups. This suggests that the transformation of rainfed crops into irrigated agriculture has resulted in an overall homogenization of the landscape with negative consequences for amphibian communities.
As we predicted, amphibian species richness was significantly higher in rainfed than in irrigated ponds (Figure 2), in accordance with previous studies supporting a positive relationship between landscape heterogeneity and amphibian richness [43,59]. Species composition was similar in irrigated and rainfed areas, with the exception of D. galganoi, which was only present in the latter, albeit in a small number of ponds. Overall, total species abundance was not significantly different in the two agro-systems, but some species were markedly less abundant in the irrigated area, including B. spinosus, P. cultripes and P. waltl. Pelobates showed the most contrasting patterns of occupancy between the two areas, as shown by our SIMPER analysis. This species has low dispersal capacity compared to other sympatric amphibian species [44], and is strongly dependent on non-compacted soils, where they burrow for refuge [60]. Traditionally managed areas could favor the presence of the species by maintaining a larger extent of bare soil and fallow. Negative effects of intensive agriculture on this species were also reported by [33], who found a strong negative correlation between agricultural cover and larval abundance. On the other hand, Discoglossus usually breeds in shallow and ephemeral puddles [61]; the loss of inundated grassland habitats surrounding ponds in the irrigated area may have negatively affected this species. Deleterious effects of agrochemicals have been reported for both Pelobates and Discoglossus in their larval stages [62,63], suggesting that reduced abundance in intensified areas could be associated with the increased use of fertilizers and pesticides around irrigated ponds. A previous study [64] found important concentrations of pesticides and fertilizers in the study area, with a positive relationship between the intensity of agricultural management and the amount of polycyclic aromatic hydrocarbons (PAHs, a family of pollutants) in the ponds.
Beta diversity also differed across management groups, while the turnover component dominated on both irrigated and rainfed pond groups, as in other amphibian studies [65,66] (but see [67]). The latter showed higher beta diversity on all three components. The pairwise analysis showed a “harmonic” pattern on the rainfed side, with a majority of pond pairs showing small differences in community composition, in contrast with irrigated ponds, where we found a non-normal pattern, with more pronounced community differences among ponds. This may be the result of local extinction processes in the irrigated pond metacommunity, with some irrigated ponds having only three or less species. Local extinctions can increase pairwise beta diversity due to turnover among impoverished communities (subtractive heterogenization [53]). Pairwise beta diversity due to nestedness is also expected to increase among pairs of ponds with different levels of degradation, as the most degraded will have lost a higher number of species. Pleurodeles waltl, B. spinosus, H. molleri, P. perezi and especially P. cultripes occur less frequently in irrigated than in rainfed ponds, and could thus be considered to be more sensitive towards intensification, driving observed inter-pond differences. On the other hand, E. calamita and T. marmoratus show an equal number of occurrences on both irrigated and rainfed areas, and can therefore be regarded as more tolerant to changes in agricultural practices. This decrease in beta diversity associated with agricultural intensification has not been described in amphibians or in Mediterranean environments so far, but it has been reported in other taxonomic groups in different geographic regions [68,69,70].
To our knowledge, this is the first study to apply the EMS approach for the characterization of amphibian metacommunities. Our results suggest that amphibian communities in areas affected by agricultural intensification are more randomly structured, whereas in traditionally managed areas, species arrange in communities formed by species with similar environmental preferences. The analysis shows rainfed ponds display a Clementsian metacommunity pattern, whereas no clear pattern could be associated with irrigated ponds, as their incidence matrices were not significantly coherent [56]. Clementsian patterns are named after the classic text by Clements [71], and represent discrete species assemblages that replace each other along an environmental gradient [55]. Each assemblage can be referred to as a compartment [72]; based on ordered matrices we can distinguish three assemblages in rainfed ponds: ponds with D. galganoi and B. spinosus, ponds with E. calamita, and ponds hosting only the most generalist species. This result is in accordance with the higher importance of the turnover beta diversity component on the rainfed side, both for total beta and for pairwise values. The absence of D. galganoi and the extreme rarity of B. spinosus in the irrigated area, along with the greater scarcity of species that are common in the rainfed area, probably explain the random metacommunity structure of irrigated ponds. Differences in community structure could thus represent the disassembly of a recognizable metacommunity pattern (Clementsian, Gleasonian or Nested) into a random pattern following anthropic disturbances [73,74].
Our analysis for individual amphibian species showed lower potential to detect possible effects of agricultural management regimes compared to analyses at the community-level. This shows the advantage of metacommunity approaches when addressing large scale ecological perturbations associated with human activities. Specifically, logistic regressions were not very informative, probably because many species were present in most of the ponds, while others appeared in a small number of ponds. This reduces the power of this type of analysis to relate the presence of each species to environmental variables. On the other hand, RDA provided further insights on the role of different factors in shaping community differences across management groups. Specifically, these analyses revealed a positive association of all amphibian species with the amount of natural vegetation, and negative responses to crop cover. The exception is E. calamita, which seems to be more dependent on local variables, especially pond area and depth [75], and thus more indifferent to agricultural management regimes. A positive relationship between amphibian richness and natural vegetation cover has been found in previous studies [45,76,77,78]. Amphibian species in our study area showed different responses to anthropogenic alterations driven by agricultural intensification, as found in previous studies [40,79,80], and while some do not appear to be sensitive to landscape homogenization, those most affected are Iberian endemics of conservation concern (D. galganoi, P. cultripes), included in the EU Habitats Directive.
Our results have important implications for amphibian conservation in Mediterranean croplands. First, local variables seem to be more important than landscape variables in driving the observed turnover (i.e., Clementsian) pattern among rainfed ponds. In this context, conservation planning is recommended to target multiple ponds, not only those with higher species richness [53]. In contrast, conservation efforts should focus on the richest sites at the expense of species-poorer communities when nested patterns are found, as in some ponds in the irrigated area. In Mediterranean pond-breeding amphibian communities, which are characterized by strong rainfall variation and unpredictability and thus with high probability of local extinctions due to stochastic factors, these general guidelines should be complemented with explicit assessments of inter-pond connectivity to account for possible rescue effects [81].
Agricultural intensification has been shown to promote population fragmentation in amphibians, but crop compositional heterogeneity can have positive effects for some species [59]. In our study area, however, diversification of crop types with the introduction of irrigated crops seems to have negatively affected amphibian communities. Moreover, the increase of the water content of soils and the presence of channels with excess water from irrigation in the irrigated area did not have a positive effect on amphibian communities, in contrast with the significant negative effect of reduced natural vegetation patches (forest, scrub and grassland). Our study supports the application of environmental measures for biodiversity conservation in European agricultural policies, including preserving strips of forest or scrub across crop plots to favor the persistence of diverse amphibian communities. Patches of natural grassland and bare soil, especially near water bodies, can also provide adequate foraging and terrestrial refugial habitats for some species. Regulations like the Common Agriculture Policy should stress the role of patches of natural vegetation and promote the construction and maintenance of small ponds and wetlands, providing breeding habitat, terrestrial corridors and refugial areas for amphibians.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/land10090924/s1, Table S1: Location of the sampled 32 ponds and agricultural management groups (Datum: WGS84); Table S2: Pearson’s correlation between environmental variables from sampled ponds. In brown, removed variables due to high correlation. Local variables are AREA: pond area (m2), MXDE: maximum pond depth on all visits (cm), MEDE: medium pond depth on all visits (cm), HECO: proportion of helophyte cover over pond area (0–1). Landscape variables (proportion of surface on a 500 m or 1000 m buffer around the pond) are ARTR: man-made forest, poplar or pine, GRA: grasslands, WET: wetlands other than the sampled pond, NTTR: natural oak forest, ART: artificial surfaces, BAR: bare ground, SCR: scrublands; Table S3: Means and range of values for environmental variables recorded in ponds in the two agricultural management areas, with statistical significance of differences between irrigated and rainfed groups assessed with Mann-Whitney tests. Variable names as in Table S2; Table S4: Environmental variables summarized via Principal Components Analysis (PCA) showing the scores for each principal component (PC) and environmental variable; Table S5: Occurrences of amphibian species (number and proportion of occupied ponds) in irrigated vs. rainfed ponds. Statistical significance of differences was assessed by means of Fisher’s test; Table S6: Results of the SIMPER analysis between irrigated and rainfed pond communities; Table S7: Abundances of amphibian species (individuals/1000 m2) in irrigated vs. rainfed ponds. Statistical significance of differences between the two agricultural areas was assessed with ANOSIM test.
Author Contributions
Conceptualization, L.A., Í.M.-S. and E.B.; methodology, L.A., A.A.; formal analysis, L.A.; resources, M.L., E.B.; writing—original draft preparation, L.A.; writing—review and editing, all authors; supervision, Í.M.-S., E.B.; project administration, E.B.; funding acquisition, E.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Spanish Ministerio de Economía, Industria y Competitividad, project Metaponds, (ref: CGL2017-84176-R); and by Fundación Biodiversidad, Ministerio para la Transición Ecológica y el Reto Demográfico (project BT-2019). L. Albero is funded by a Phd grant from Universidad de León (ULE).
Institutional Review Board Statement
All experimental protocols and sampling permits were approved by the regional authority (Consejería de Medio Ambiente, Junta de Castilla y León, reference: AUES/CYL/693/2019). We followed ethical guidelines provided by Junta de Castilla y León and León University (Universidad de León). No amphibians were harmed during sampling; and handling, when necessary, involved the minimum time required for species identification. We disinfected field equipment between ponds to prevent transmission of infectious diseases.
Acknowledgments
We wish to thank all people who helped during field sampling: O. Rodríguez, A. Casanova, M. Borrego, V. Ezquerra, P. Oviedo, A. Cachorro, R. de Prado and specially A. Benito. We also thank Fundación SCAYLE for their support with bioinformatics.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Ceballos, G.; Ehrlich, P.R. The misunderstood sixth mass extinction. Science 2018, 360, 1080–1081. [Google Scholar] [CrossRef]
- Pimm, S.L.; Jenkins, C.N.; Abell, R.; Brooks, T.M.; Gittleman, J.L.; Joppa, L.N.; Raven, P.H.; Roberts, C.M.; Sexton, J.O. The biodiversity of species and their rates of extinction, distribution, and protection. Science 2014, 344. [Google Scholar] [CrossRef] [PubMed]
- Grant, E.H.C.; Muths, E.; Schmidt, B.R.; Petrovan, S.O. Amphibian conservation in the Anthropocene. Biol. Conserv. 2019, 236, 543–547. [Google Scholar] [CrossRef]
- Foley, J.A.; DeFries, R.; Asner, G.P.; Barford, C.; Bonan, G.; Carpenter, S.R.; Chapin, F.S.; Coe, M.T.; Daily, G.C.; Gibbs, H.K.; et al. Global consequences of land use. Science 2005, 309, 570–574. [Google Scholar] [CrossRef] [Green Version]
- Martin-Guay, M.O.; Paquette, A.; Dupras, J.; Rivest, D. The new Green Revolution: Sustainable intensification of agriculture by intercropping. Sci. Total Environ. 2018, 615, 767–772. [Google Scholar] [CrossRef] [PubMed]
- Pingali, P.L. Green revolution: Impacts, limits, and the path ahead. Proc. Natl. Acad. Sci. USA 2012, 109, 12302–12308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balestrieri, A.; Gazzola, A.; Formenton, G.; Canova, L. Long-term impact of agricultural practices on the diversity of small mammal communities: A case study based on owl pellets. Environ. Monit. Assess. 2019, 191. [Google Scholar] [CrossRef]
- Habel, J.C.; Ulrich, W.; Biburger, N.; Seibold, S.; Schmitt, T. Agricultural intensification drives butterfly decline. Insect Conserv. Divers. 2019, 12, 289–295. [Google Scholar] [CrossRef]
- Morgado, R.; Santana, J.; Porto, M.; Sánchez-Oliver, J.S.; Reino, L.; Herrera, J.M.; Rego, F.; Beja, P.; Moreira, F. A Mediterranean silent spring? The effects of olive farming intensification on breeding bird communities. Agric. Ecosyst. Environ. 2020, 288, 106694. [Google Scholar] [CrossRef]
- Uchida, K.; Ushimaru, A. Biodiversity declines due to abandonment and intensification of agricultural lands: Patterns and mechanisms. Ecol. Monogr. 2014, 84, 637–658. [Google Scholar] [CrossRef]
- Betts, M.G.; Wolf, C.; Pfeifer, M.; Banks-Leite, C.; Arroyo-Rodríguez, V.; Bandini Ribeiro, D.; Barlow, J.; Eigenbrod, F.; Faria, D.; Fletcher, R.J., Jr.; et al. Extinction filters mediate the global effects of habitat fragmentation on animals. Science 2019, 366, 1236–1239. [Google Scholar] [CrossRef] [Green Version]
- Fahrig, L. Ecological Responses to Habitat Fragmentation Per Se. Annu. Rev. Ecol. Evol. Syst. 2017, 48, 1–23. [Google Scholar] [CrossRef]
- Benton, T.G.; Vickery, J.A.; Wilson, J.D. Farmland biodiversity: Is habitat heterogeneity the key? Trends Ecol. Evol. 2003, 18, 182–188. [Google Scholar] [CrossRef]
- Fahrig, L.; Baudry, J.; Brotons, L.; Burel, F.G.; Crist, T.O.; Fuller, R.J.; Sirami, C.; Siriwardena, G.M.; Martin, J.L. Functional landscape heterogeneity and animal biodiversity in agricultural landscapes. Ecol. Lett. 2011, 14, 101–112. [Google Scholar] [CrossRef]
- Córdova-Lepe, F.; Del Valle, R.; Ramos-Jiliberto, R. The process of connectivity loss during habitat fragmentation and their consequences on population dynamics. Ecol. Model. 2018, 376, 68–75. [Google Scholar] [CrossRef]
- Lewis-Phillips, J.; Brooks, S.J.; Sayer, C.D.; Patmore, I.R.; Hilton, G.M.; Harrison, A.; Robson, H.; Axmacher, J.C. Ponds as insect chimneys: Restoring overgrown farmland ponds benefits birds through elevated productivity of emerging aquatic insects. Biol. Conserv. 2020, 241, 108253. [Google Scholar] [CrossRef]
- Leibold, M.A.; Holyoak, M.; Mouquet, N.; Amarasekare, P.; Chase, J.M.; Hoopes, M.F.; Holt, R.D.; Shurin, J.B.; Law, R.; Tilman, D.; et al. The metacommunity concept: A framework for multi-scale community ecology. Ecol. Lett. 2004, 7, 601–613. [Google Scholar] [CrossRef]
- Heino, J.; Melo, A.S.; Siqueira, T.; Soininen, J.; Valanko, S.; Bini, L.M. Metacommunity organisation, spatial extent and dispersal in aquatic systems: Patterns, processes and prospects. Freshw. Biol. 2015, 60, 845–869. [Google Scholar] [CrossRef]
- Chaparro, G.; Horváth, Z.; O’Farrell, I.; Ptacnik, R.; Hein, T. Plankton metacommunities in floodplain wetlands under contrasting hydrological conditions. Freshw. Biol. 2018, 60, 380–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Álvarez Cobelas, M.; Rojo, C.; Angeler, D.C. Mediterranean Limnology: Current status, gaps and the future. J. Limnol. 2005, 64, 13–29. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, M.; Beja, P. Mediterranean amphibians and the loss of temporary ponds: Are there alternative breeding habitats? Biol. Conserv. 2013, 165, 179–186. [Google Scholar] [CrossRef]
- Ruhí, A.; Sebastian, O.S.; Feo, C.; Franch, M.; Gascón, S.; Richter-Boix, À.; Boix, D.; Llorente, G. Man-made Mediterranean temporary ponds as a tool for amphibian conservation. Ann. Limnol. Int. J. Limnol. 2012, 48, 81–93. [Google Scholar] [CrossRef]
- Bolle, H.J. (Ed.) Mediterranean Climate. Variability and Trends; Springer: Berlin, Germany, 2003; 320p. [Google Scholar]
- Beklioglu, M.; Romo, S.; Kagalou, I.; Quintana, X.; Becares, E. State of the art in the functioning of shallow Mediterranean lakes: Workshop conclusions. Hydrobiologia 2007, 584, 317–326. [Google Scholar] [CrossRef]
- Harmanny, K.S.; Malek, Ž. Adaptations in irrigated agriculture in the Mediterranean region: An overview and spatial analysis of implemented strategies. Reg. Environ. Change 2019, 19, 1401–1416. [Google Scholar] [CrossRef] [Green Version]
- Giralt, D.; Pantoja, J.; Morales, M.B.; Traba, J.; Bota, G. Landscape-Scale Effects of Irrigation on a Dry Cereal Farmland Bird Community. Front. Ecol. Evol. 2021, 9, 1–8. [Google Scholar] [CrossRef]
- De Frutos, A.; Olea, P.P.; Mateo-Tomás, P. Responses of medium- and large-sized bird diversity to irrigation in dry cereal agroecosystems across spatial scales. Agric. Ecosyst. Environ. 2015, 207, 141–152. [Google Scholar] [CrossRef]
- Clemente-Orta, G.; Madeira, F.; Batuecas, I.; Sossai, I.; Juárez-Escario, A.; Albajes, R. Changes in landscape composition influence the abundance of insects on maize: The role of fruit orchards and alfalfa crops. Agric. Ecosyst. Environ. 2020, 291, 106805. [Google Scholar] [CrossRef]
- Cox, N.; Chanson, J.; Stuart, S. The Status and Distribution of Reptiles and Amphibians of the Mediterranean Basin; IUCN: Gland, Switzerland; Cambridge, UK, 2006; 42p. [Google Scholar]
- Beja, P.; Alcazar, R. Conservation of Mediterranean temporary ponds under agricultural intensification: An evaluation using amphibians. Biol. Conserv. 2003, 114, 317–326. [Google Scholar] [CrossRef]
- Fortuna, M.A.; Gómez-Rodríguez, C.; Bascompte, J. Spatial network structure and amphibian persistence in stochastic environments. Proc. R. Soc. Lond. B Biol. Sci. 2005, 273, 1429–1434. [Google Scholar] [CrossRef] [Green Version]
- Caballero-Díaz, C.; Sánchez-Montes, G.; Butler, H.M.; Vredenburg, V.T.; Martínez-Solano, Í. The role of artificial breeding sites in amphibian conservation: A case study in rural areas in central Spain. Herpetol. Conserv. Biol. 2020, 15, 87–104. [Google Scholar]
- Valdez, J.W.; Gould, J.; Garnham, J.I. Global assessment of artificial habitat use by amphibian species. Biol. Conserv. 2021, 257, 109129. [Google Scholar] [CrossRef]
- Jakob, C.; Poizat, G.; Veith, M.; Seitz, A.; Crivelli, A.J. Breeding phenology and larval distribution of amphibians in a Mediterranean pond network with unpredictable hydrology. Hydrobiologia 2003, 499, 51–61. [Google Scholar] [CrossRef]
- Richter-Boix, A.; Llorente, G.A.; Montori, A. Structure and dynamics of an amphibian metacommunity in two regions. J. Anim. Ecol. 2007, 76, 607–618. [Google Scholar] [CrossRef] [PubMed]
- Fuentes-Rodríguez, F.; Juan, M.; Gallego, I.; Lusi, M.; Fenoy, E.; León, D.; Peñalver, P.; Toja, J.; Casas, J.J. Diversity in Mediterranean farm ponds: Trade-offs and synergies between irrigation modernisation and biodiversity conservation. Freshw. Biol. 2003, 58, 63–78. [Google Scholar] [CrossRef]
- Aspe, C.; Gilles, A.; Jacqué, M. Irrigation canals as tools for climate change adaptation and fish biodiversity management in Southern France. Reg. Environ. Change 2014, 16, 1975–1984. [Google Scholar] [CrossRef] [Green Version]
- Riedener, E.; Rusterholz, H.P.; Baur, B. Land-use abandonment owing to irrigation cessation affects the biodiversity of hay meadows in an arid mountain region. Agric. Ecosyst. Environ. 2016, 185, 144–152. [Google Scholar] [CrossRef]
- Fernández Aláez, M.; Fernández Aláez, C.; Rodríguez, S.; Bécares, E. Evaluation of the state of conservation of shallow ponds in the province of Leon (Northwest Spain) using botanical criteria. Limnetica 1999, 17, 107–117. [Google Scholar]
- Fernandez-Aláez, C.; Fernández-Aláez, M.; Trigal, C.; Luis, B. Hydrochemistry of northwest Spain ponds and its relationships to groundwaters. Limnetica 2006, 25, 433–452. [Google Scholar]
- Pozo, R.; Fernandez-Aláez, M.; Fernández-Aláez, C. Composición de las comunidades de macrófitos y establecimiento del estado de conservación de charcas y lagunas de la Depresión del Duero (noroeste de España) en base a criterios botánicos. Limnetica 2012, 31, 47–58. [Google Scholar]
- Logue, J.B.; Mouquet, N.; Peter, H.; Hillebrand, H. Empirical approaches to metacommunities: A review and comparison with theory. Trends Ecol. Evol. 2011, 26, 482–491. [Google Scholar] [CrossRef]
- Boissinot, A.; Besnard, A.; Lourdais, O. Amphibian diversity in farmlands: Combined influences of breeding-site and landscape attributes in western France. Agric. Ecosyst. Environ. 2019, 269, 51–61. [Google Scholar] [CrossRef]
- Hecnar, S.J.; M’Closkey, R.T. Species richness patterns of amphibians in southwestern Ontario ponds. J. Biogeogr. 1998, 25, 763–772. [Google Scholar] [CrossRef]
- Piha, H.; Luoto, M.; Merila, J. Amphibian Occurrence Is Influenced by Current and Historic Landscape Characteristics. Ecol. Appl. 2007, 17, 2298–2309. [Google Scholar] [CrossRef] [PubMed]
- Indermaur, L.; Schaub, M.; Jokela, J.; Tockner, K.; Schmidt, B.R. Differential response to abiotic conditions and predation risk rather than competition avoidance determine breeding site selection by anurans. Ecography 2010, 33, 887–895. [Google Scholar] [CrossRef]
- Cayuela, H.; Valenzuela-Sánchez, A.; Teulier, L.; Martínez-Solano, Í.; Léna, J.P.; Merilä, J.; Muths, E.; Shine, R.; Quay, L.; Denoël, M.; et al. Determinants and consequences of dispersal in vertebrates with complex life cycles: A review of pond-breeding amphibians. Quart. Rev. Biol. 2020, 95, 1–36. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Montes, G.; Wang, J.; Ariño, A.H.; Martínez-Solano, Í. Mountains as barriers to gene flow in amphibians: Quantifying the differential effect of a major mountain ridge on the genetic structure of four sympatric species with different life history traits. J. Biogeogr. 2018, 45, 318–331. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014; Available online: http://www.R-project.org/ (accessed on 30 January 2021).
- Baselga, A. Partitioning the turnover and nestedness components of beta diversity. Glob. Ecol. Biogeogr. 2010, 19, 134–143. [Google Scholar] [CrossRef]
- Baselga, A.; Orme, C.D.L. Betapart: An R package for the study of beta diversity. Methods Ecol. Evol. 2012, 3, 808–812. [Google Scholar] [CrossRef]
- Marion, Z.H.; Fordyce, J.A.; Fitzpatrick, B.M. Pairwise beta diversity resolves an underappreciated source of confusion in calculating species turnover. Ecology 2017, 98, 933–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Socolar, J.B.; Gilroy, J.J.; Kunin, W.E.; Edwards, D.P. How Should Beta-Diversity Inform Biodiversity Conservation? Trends Ecol. Evol. 2016, 31, 67–80. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J. Species Association Analysis. 2016. Available online: https://cran.r-project.org/web/packages/spaa/spaa.pdf (accessed on 30 January 2021).
- Leibold, M.A.; Mikkelson, G.M. Coherence, species turnover, and boundary clumping: Elements of meta-community structure. Oikos 2002, 97, 237–250. [Google Scholar] [CrossRef] [Green Version]
- Presley, S.J.; Higgins, C.L.; Willig, M.R. A comprehensive framework for the evaluation of metacommunity structure. Oikos 2010, 119, 908–917. [Google Scholar] [CrossRef]
- Dallas, T. Metacom: An R package for the analysis of metacommunity structure. Ecography 2014, 37, 402–405. [Google Scholar] [CrossRef] [Green Version]
- Ter Braak, C.J.F.; Smilauer, P. CANOCO Reference Manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination (Version 4.5); Microcomputer Power: Ithaca, NY, USA, 2002; 500p. [Google Scholar]
- Collins, S.J.; Fahrig, L. Responses of anurans to composition and configuration of agricultural landscapes. Agric. Ecosyst. Environ. 2017, 239, 399–409. [Google Scholar] [CrossRef]
- Recuero, E. Sapo de Espuelas—Pelobates cultripes. In Enciclopedia Virtual de los Vertebrados Españoles; Salvador, A., Martínez Solano, I., Eds.; Museo Nacional de Ciencias Naturales: Madrid, Spain, 2014. Available online: http://www.vertebradosibericos.org/ (accessed on 30 January 2021).
- Martínez-Solano, I. Sapillo pintojo ibérico—Discoglossus galganoi. In Enciclopedia Virtual de los Vertebrados Españoles; Salvador, A., Martínez-Solano, I., Eds.; Museo Nacional de Ciencias Naturales: Madrid, Spain, 2014. Available online: http://www.vertebradosibericos.org/ (accessed on 30 January 2021).
- Ortiz, M.E.; Marco, A.; Saiz, N.; Lizana, M. Impact of ammonium nitrate on growth and survival of six European amphibians. Arch. Environ. Contam. Toxicol. 2004, 47, 234–239. [Google Scholar] [CrossRef]
- Polo-Cavia, N.; Burraco, P.; Gomez-Mestre, I. Low levels of chemical anthropogenic pollution may threaten amphibians by impairing predator recognition. Aquat. Toxicol. 2016, 172, 30–35. [Google Scholar] [CrossRef]
- Hijosa-Valsero, M.; Bécares, E.; Fernández-Aláez, C.; Fernández-Aláez, M.; Mayo, R.; Jiménez, J.J. Chemical pollution in inland shallow ponds in the Mediterranean region (NW Spain): PAHs, insecticides and herbicides in water and sediments. Sci. Total Environ. 2016, 544, 797–810. [Google Scholar] [CrossRef]
- Tavares, H.N.; Da Silva, F.R. Species turnover drives the spatial distribution of frog beta diversity in farmland ponds. J. Trop. Ecol. 2019, 35, 199–202. [Google Scholar] [CrossRef]
- Boix, D.; Caria, M.C.; Gascón, S.; Mariani, M.A.; Sala, J.; Ruhí, A.; Compte, J.; Bagella, S. Contrasting intra-annual patterns of six biotic groups with different dispersal mode and ability in Mediterranean temporary ponds. Mar. Freshw. Res. 2017, 68, 1044–1060. [Google Scholar] [CrossRef]
- Ficetola, G.F.; De Bernardi, F. Amphibians in a human-dominated landscape: The community structure is related to habitat features and isolation. Biol. Conserv. 2004, 119, 219–230. [Google Scholar] [CrossRef]
- Santana, J.; Porto, M.; Reino, L.; Moreira, F.; Ribeiro, P.F.; Santos, J.L.; Rotenberry, J.T.; Beja, P. Using beta diversity to inform agricultural policies and conservation actions on Mediterranean farmland. J. Appl. Ecol. 2017, 54, 1825–1835. [Google Scholar] [CrossRef] [Green Version]
- Sueyoshi, M.; Ishiyama, N.; Nakamura, F. β-diversity decline of aquatic insects at the microhabitat scale associated with agricultural land use. Landsc. Ecol. Eng. 2016, 12, 187–196. [Google Scholar] [CrossRef]
- Johnson, R.K.; Angeler, D.G. Effects of agricultural land use on stream assemblages: Taxon-specific responses of alpha and beta diversity. Ecol. Indic. 2014, 45, 386–393. [Google Scholar] [CrossRef]
- Clements, F.E. Plant Succession. An Analysis of the Development of Vegetation; Carnegie Institution of Washington: Washington, DC, USA, 1917. [Google Scholar] [CrossRef] [Green Version]
- Lewinsohn, T.M.; Inácio Prado, P.; Jordano, P.; Bascompte, J.; Olesen, J.M. Structure in plant-animal interaction assemblages. Oikos 2006, 113, 174–184. [Google Scholar] [CrossRef] [Green Version]
- Brasil, L.S.; Vieira, T.B.; de Oliveira-Junior, J.M.B.; Dias-Silva, K.; Juen, L. Elements of metacommunity structure in Amazonian Zygoptera among streams under different spatial scales and environmental conditions. Ecol. Evol. 2019, 7, 3190–3200. [Google Scholar] [CrossRef] [PubMed]
- Leavitt, D.J.; Fitzgerald, L.A. Disassembly of a dune-dwelling lizard community due to landscape fragmentation. Ecosphere 2013, 4, 1–15. [Google Scholar] [CrossRef]
- Gómez-Mestre, I. Sapo corredor—Epidalea calamita. In Enciclopedia Virtual de los Vertebrados Españoles; Salvador, A., Martínez-Solano, I., Eds.; Museo Nacional de Ciencias Naturales: Madrid, Spain, 2014. Available online: http://www.vertebradosibericos.org/ (accessed on 30 January 2021).
- Price, S.J.; Marks, D.R.; Howe, R.W.; Hanowski, J.A.M.; Niemi, G.J. The importance of spatial scale for conservation and assessment of anuran populations in coastal wetlands of the western Great Ponds, USA. Landsc. Ecol. 2005, 20, 441–454. [Google Scholar] [CrossRef]
- Rubbo, M.J.; Kiesecker, J.M. Amphibian Breeding Distribution in an Urbanized Landscape. Conserv. Biol. 2005, 19, 504–511. [Google Scholar] [CrossRef]
- Couto, A.P.; Ferreira, E.; Torres, R.T.; Fonseca, C. Local and landscape drivers of pond-breeding amphibian diversity at the northern edge of the Mediterranean. Herpetologica 2017, 73, 10–17. [Google Scholar] [CrossRef]
- Pyron, R.A. Global amphibian declines have winners and losers. Proc. Natl. Acad. Sci. USA 2018, 115, 3739–3741. [Google Scholar] [CrossRef] [Green Version]
- Nowakowski, A.J.; Thompson, M.E.; Donnelly, M.A.; Todd, B.D. Amphibian sensitivity to habitat modification is associated with population trends and species traits. Glob. Ecol. Biogeogr. 2017, 26, 700–712. [Google Scholar] [CrossRef] [Green Version]
- Capellà-Marzo, B.; Sánchez-Montes, G.; Martínez-Solano, I. Contrasting demographic trends and asymmetric migration rates in a spatially structured amphibian population. Integr. Zool. 2020, 15, 482–497. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Map of the study area showing its location in León Province (Spain, inset), landscape cover types and sampled ponds in each agricultural management group. The blue line represents Payuelos Channel. “Natural vegetation” includes scrub, grassland and bare soil; “Artificial” includes roads and urban areas.
Figure 1.
Map of the study area showing its location in León Province (Spain, inset), landscape cover types and sampled ponds in each agricultural management group. The blue line represents Payuelos Channel. “Natural vegetation” includes scrub, grassland and bare soil; “Artificial” includes roads and urban areas.

Figure 2.
Comparison of mean amphibian abundance (left) and species richness (right) between irrigated and rainfed areas. * Significant under α = 0.05.
Figure 2.
Comparison of mean amphibian abundance (left) and species richness (right) between irrigated and rainfed areas. * Significant under α = 0.05.

Figure 3.
Distribution of pairwise beta diversity values for each component of beta diversity in irrigated (left) and rainfed (right) ponds, following the additive partition approach.
Figure 3.
Distribution of pairwise beta diversity values for each component of beta diversity in irrigated (left) and rainfed (right) ponds, following the additive partition approach.

Figure 4.
Occurrence matrices of all amphibian species in irrigated (left) and rainfed (right) ponds, ordered by maximum coherence following EMS analysis. Black boxes indicate species presence (columns) in each pond (rows). EC: Epidalea calamita, PC: Pelobates cultripes, PW: Pleurodeles waltl, HM: Hyla molleri, PP: Pelophylax perezi, TM: Triturus marmoratus, DG: Discoglossus galganoi, BS: Bufo spinosus.
Figure 4.
Occurrence matrices of all amphibian species in irrigated (left) and rainfed (right) ponds, ordered by maximum coherence following EMS analysis. Black boxes indicate species presence (columns) in each pond (rows). EC: Epidalea calamita, PC: Pelobates cultripes, PW: Pleurodeles waltl, HM: Hyla molleri, PP: Pelophylax perezi, TM: Triturus marmoratus, DG: Discoglossus galganoi, BS: Bufo spinosus.

Figure 5.
RDA ordination plot showing the main two redundancy axes and their relationships with local and landscape environmental variables (red arrows) and patterns of amphibian species occurrence (blue arrows). PLEWAT: Pleurodeles waltl; TRIMAR: Triturus marmoratus; PELCUL: Pelobates cultripes; DISGAL: Discoglossus galganoi; BUFSPI: Bufo spinosus; EPICAL: Epidalea calamita; HYLMOL: Hyla molleri; PELPER: Pelophylax perezi. Environmental variables at the local scale: pond area in m2 (AREA); medium pond depth (MEDE); helophyte cover in % pond area (HECO). Landscape variables in % cover at a 1000 m scale from the pond: man-made forest, poplar or pine (ARTR1000); grasslands (GRA1000); wetlands (WET1000). In % cover at a 500 m scale from the pond: natural oak forest (NTTR500), artificial surfaces (ART500), bare ground (BAR500), scrub (SCR500).
Figure 5.
RDA ordination plot showing the main two redundancy axes and their relationships with local and landscape environmental variables (red arrows) and patterns of amphibian species occurrence (blue arrows). PLEWAT: Pleurodeles waltl; TRIMAR: Triturus marmoratus; PELCUL: Pelobates cultripes; DISGAL: Discoglossus galganoi; BUFSPI: Bufo spinosus; EPICAL: Epidalea calamita; HYLMOL: Hyla molleri; PELPER: Pelophylax perezi. Environmental variables at the local scale: pond area in m2 (AREA); medium pond depth (MEDE); helophyte cover in % pond area (HECO). Landscape variables in % cover at a 1000 m scale from the pond: man-made forest, poplar or pine (ARTR1000); grasslands (GRA1000); wetlands (WET1000). In % cover at a 500 m scale from the pond: natural oak forest (NTTR500), artificial surfaces (ART500), bare ground (BAR500), scrub (SCR500).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Partition of total and pairwise beta diversity in irrigated and rainfed ponds, following the additive partition approach. The statistical significance of differences between groups was assessed with Mann–Whitney tests. * Significant under α = 0.05.
Table 1.
Partition of total and pairwise beta diversity in irrigated and rainfed ponds, following the additive partition approach. The statistical significance of differences between groups was assessed with Mann–Whitney tests. * Significant under α = 0.05.
| Sorensen | Simpson | Nestedness | |
|---|---|---|---|
| All ponds Rainfed | 0.75 | 0.5 | 0.25 |
| All ponds Irrigated | 0.6 | 0.42 | 0.18 |
| All ponds Rainfed + Irrigated | 0.81 | 0.61 | 0.2 |
| Mean Pairwise Rainfed | 0.177 | 0.092 | 0.084 |
| Mean Pairwise Irrigated | 0.368 | 0.166 | 0.202 |
| Pairwise U Mann-Whitney | 16,996 | 30,014 | 21,802 |
| Pairwise p-value | 2.20 × 10−16 * | 0.06 | 3.478 × 10−11 * |
Table 2.
Results of the Elements of Metacommunity Structure Analysis. * Significant under α = 0.05.
| Coherence z | Coherence p-Value | Turnover z | Turnover p-Value | Morisita Index | Morisita p-Value | Structure | |
|---|---|---|---|---|---|---|---|
| Rainfed | 2.05 | 0.04 * | 1.78 | 0.075 | 2 | 0.0012 * | Clementsian |
| Irrigated | 0.62 | 0.53 | 1.7 | 0.09 | NA | NA | Random |
| All ponds | 0.96 | 0.34 | 2.41 | 0.015 * | NA | NA | Random |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Albero, L.; Martínez-Solano, Í.; Arias, A.; Lizana, M.; Bécares, E. Amphibian Metacommunity Responses to Agricultural Intensification in a Mediterranean Landscape. Land 2021, 10, 924. https://0-doi-org.brum.beds.ac.uk/10.3390/land10090924
AMA Style
Albero L, Martínez-Solano Í, Arias A, Lizana M, Bécares E. Amphibian Metacommunity Responses to Agricultural Intensification in a Mediterranean Landscape. Land. 2021; 10(9):924. https://0-doi-org.brum.beds.ac.uk/10.3390/land10090924
Chicago/Turabian StyleAlbero, Luis, Íñigo Martínez-Solano, Ana Arias, Miguel Lizana, and Eloy Bécares. 2021. "Amphibian Metacommunity Responses to Agricultural Intensification in a Mediterranean Landscape" Land 10, no. 9: 924. https://0-doi-org.brum.beds.ac.uk/10.3390/land10090924
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.