
Effects of Tillage and Winter Cover Management in a Maize Soybean Rotation on Soil Bacterial and Fungal Community Composition


Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Experimental Design and Treatment Description
2.3. Soil Sampling
2.4. DNA Extraction, Sequencing, Data Processing and Analysis
2.5. Analyses of Soil Properties
2.6. Statistical Analysis
3. Results
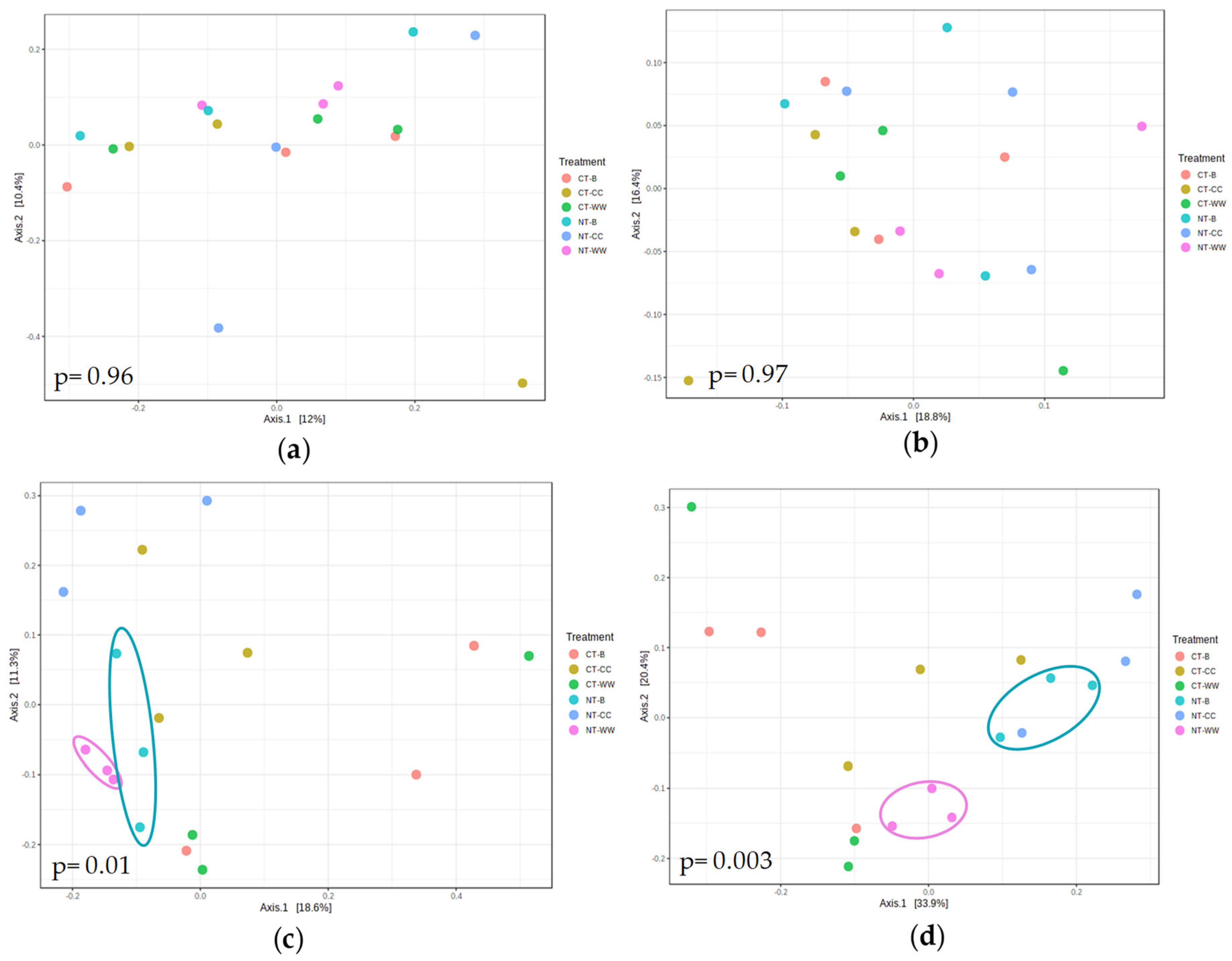
3.1. Microbial Alpha and Beta Diversity Differences in the Different Management Systems
3.2. Bacterial and Fungal Community Composition in the Different Management Systems
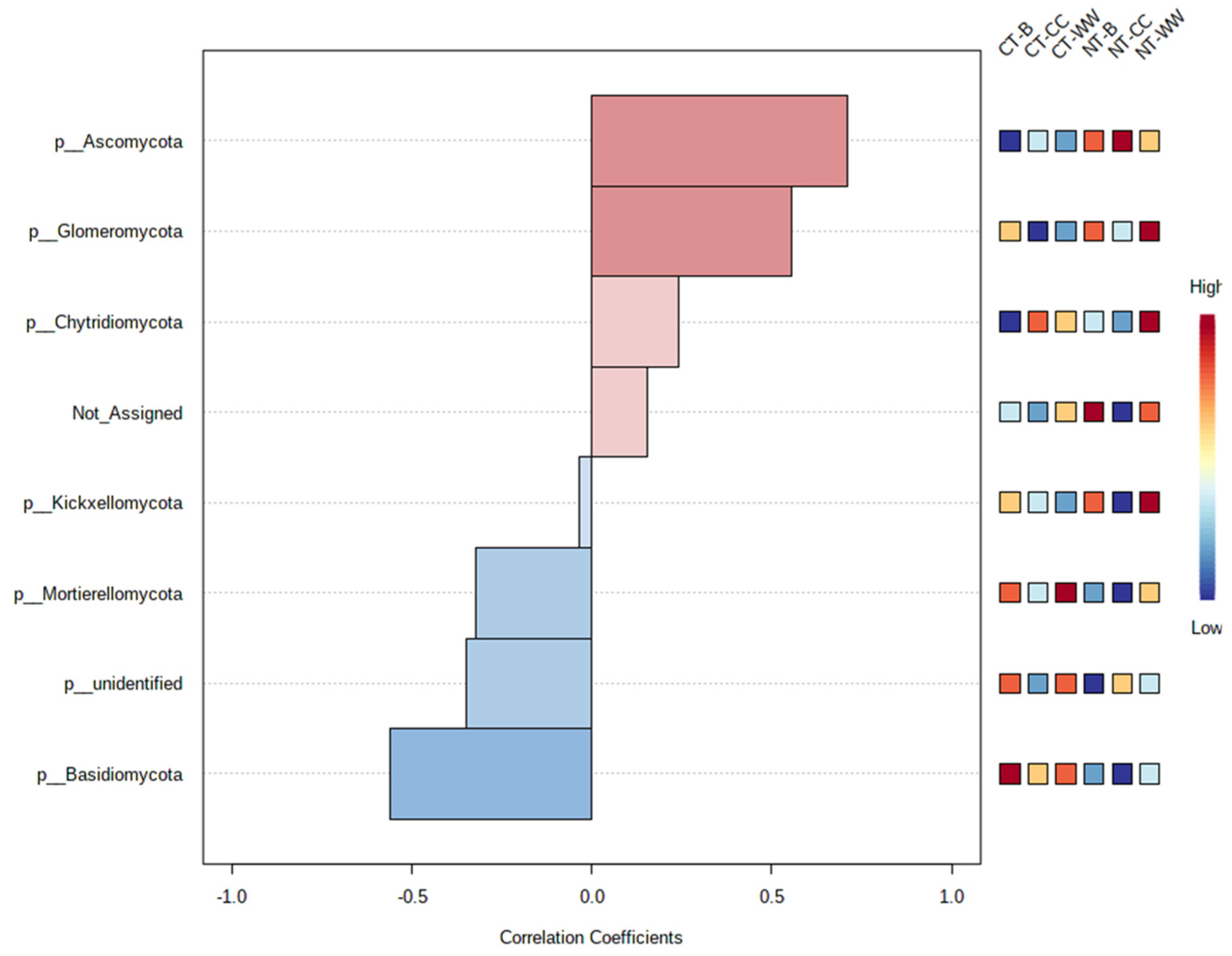
3.3. Microbial Community Profiling and Correlation Analysis
3.4. Soil Properties
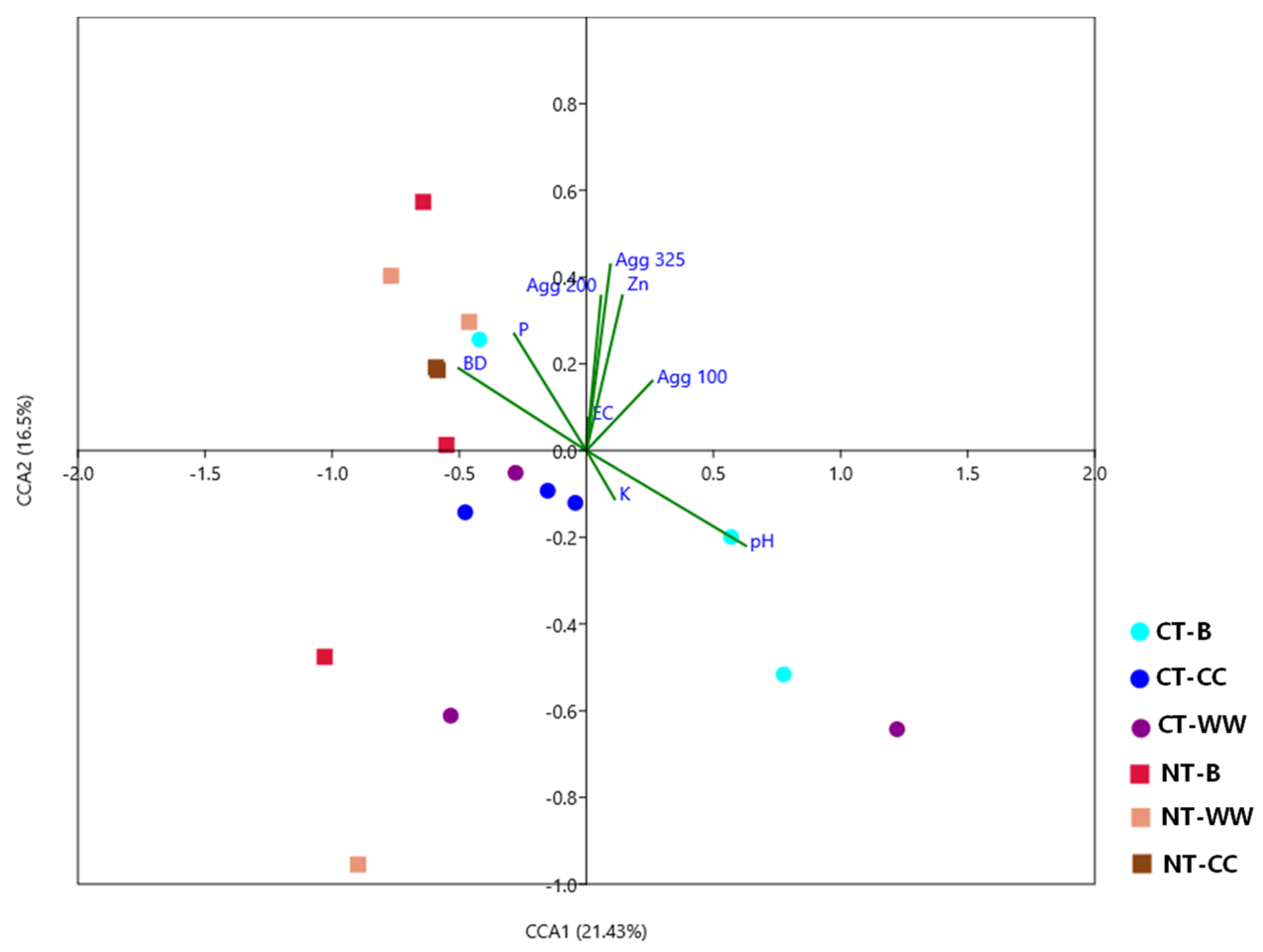
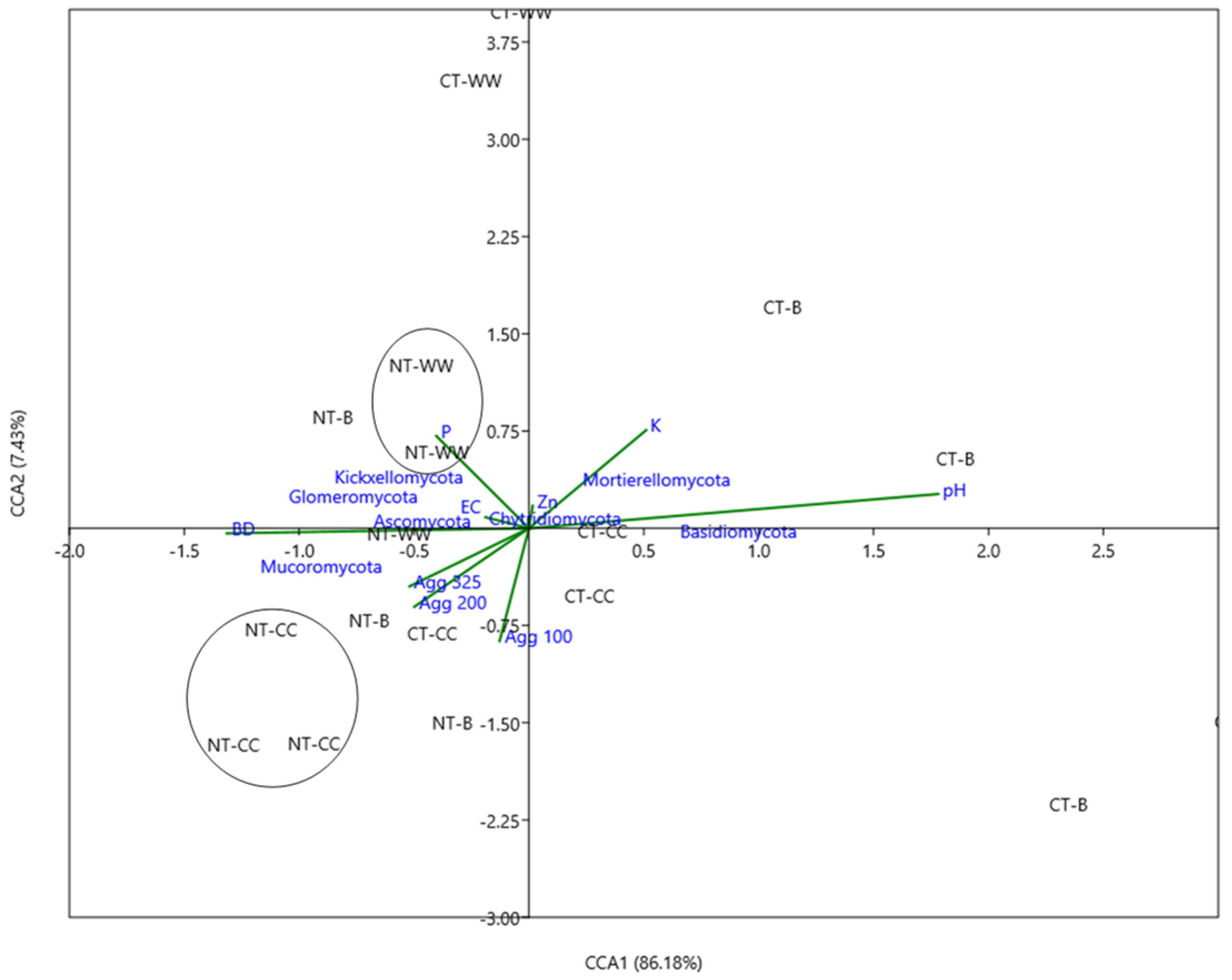
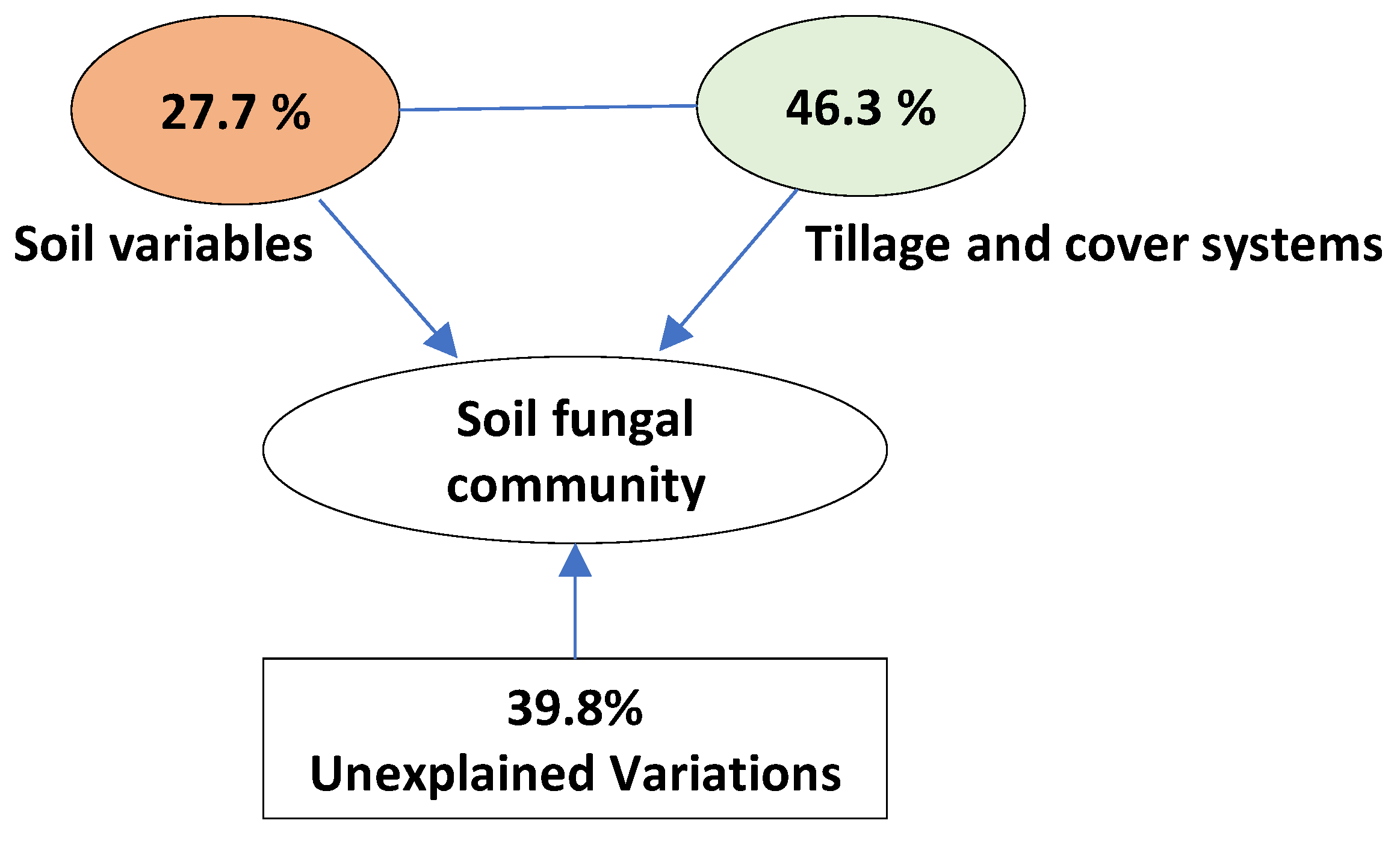
3.5. Relationship between Soil Properties and Fungal Community Composition
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ranjard, L.; Poly, F.; Nazaret, S. Monitoring Complex Bacterial Communities Using Culture-Independent Molecular Techniques: Application to Soil Environment. Res. Microbiol. 2000, 151, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Sjöberg, G.; Nilsson, S.I.; Persson, T.; Karlsson, P. Degradation of Hemicellulose, Cellulose and Lignin in Decomposing Spruce Needle Litter in Relation to N. Soil Biol. Biochem. 2004, 36, 1761–1768. [Google Scholar] [CrossRef]
- White, P.M.; Rice, C.W. Tillage Effects on Microbial and Carbon Dynamics during Plant Residue Decomposition. Soil Sci. Soc. Am. J. 2009, 73, 138–145. [Google Scholar] [CrossRef]
- Stefan, L.; Hartmann, M.; Engbersen, N.; Six, J.; Schöb, C. Positive Effects of Crop Diversity on Productivity Driven by Changes in Soil Microbial Composition. Front. Microbiol. 2021, 12, 660749. [Google Scholar] [CrossRef] [PubMed]
- Chaparro, J.M.; Sheflin, A.M.; Manter, D.K.; Vivanco, J.M. Manipulating the Soil Microbiome to Increase Soil Health and Plant Fertility. Biol. Fertil. Soils. 2012, 48, 489–499. [Google Scholar] [CrossRef]
- Ceja-Navarro, J.A.; Rivera-Orduña, F.N.; Patiño-Zúñiga, L.; Vila-Sanjurjo, A.; Crossa, J.; Govaerts, B.; Dendooven, L. Phylogenetic and Multivariate Analyses To Determine the Effects of Different Tillage and Residue Management Practices on Soil Bacterial Communities. Appl. Environ. Microbiol. 2010, 76, 3685–3691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choudhary, M.; Sharma, P.C.; Jat, H.S.; Dash, A.; Rajashekar, B.; McDonald, A.J.; Jat, M.L. Soil Bacterial Diversity under Conservation Agriculture-Based Cereal Systems in Indo-Gangetic Plains. 3 Biotech 2018, 8, 304. [Google Scholar] [CrossRef]
- Kim, N.; Zabaloy, M.C.; Guan, K.; Villamil, M.B. Do Cover Crops Benefit Soil Microbiome? A Meta-Analysis of Current Research. Soil Biol. Biochem. 2020, 142, 107701. [Google Scholar] [CrossRef]
- Miguez, F.E.; Bollero, G.A. Review of Corn Yield Response under Winter Cover Cropping Systems Using Meta-Analytic Methods. Crop Sci. 2005, 45, 2318–2329. [Google Scholar] [CrossRef] [Green Version]
- Plastina, A.; Liu, F.; Sawadgo, W.; Miguez, F.; Carlson, S.; Marcillo, G. Annual Net Returns to Cover Crops in Iowa. J. Agric. Appl. Econ. 2018, 2, 2. [Google Scholar] [CrossRef]
- Huang, M.; Jiang, L.; Zou, Y.; Xu, S.; Deng, G. Changes in Soil Microbial Properties with No-Tillage in Chinese Cropping Systems. Biol. Fertil. Soils 2013, 49, 373–377. [Google Scholar] [CrossRef]
- Pannkuk, C.D.; Papendick, R.I.; Saxton, K.E. Fallow Management Effects on Soil Water Storage and Wheat Yields in the Pacific Northwest. Agron. J. 1997, 89, 386–391. [Google Scholar] [CrossRef]
- Lange, M.; Habekost, M.; Eisenhauer, N.; Roscher, C.; Bessler, H.; Engels, C.; Oelmann, Y.; Scheu, S.; Wilcke, W.; Schulze, E.-D.; et al. Biotic and Abiotic Properties Mediating Plant Diversity Effects on Soil Microbial Communities in an Experimental Grassland. PLoS ONE 2014, 9, e96182. [Google Scholar] [CrossRef] [Green Version]
- Cambardella, C.A.; Elliott, E.T. Particulate Soil Organic-Matter Changes across a Grassland Cultivation Sequence. Soil Sci. Soc. Am. J. 1992, 56, 777–783. [Google Scholar] [CrossRef]
- Shanmugam, S.G.; Buehring, N.W.; Prevost, J.D.; Kingery, W.L. Soil Bacterial Community Diversity and Composition as Affected by Tillage Intensity Treatments in Corn-Soybean Production Systems. Microbiol. Res. Int. 2021, 12, 157–172. [Google Scholar] [CrossRef]
- Shah, A.N.; Tanveer, M.; Shahzad, B.; Yang, G.; Fahad, S.; Ali, S.; Bukhari, M.A.; Tung, S.A.; Hafeez, A.; Souliyanonh, B. Soil Compaction Effects on Soil Health and Cropproductivity: An Overview. Environ. Sci. Pollut. Res. 2017, 24, 10056–10067. [Google Scholar] [CrossRef] [PubMed]
- Acharya, B.S.; Dodla, S.; Gaston, L.A.; Darapuneni, M.; Wang, J.J.; Sepat, S.; Bohara, H. Winter Cover Crops Effect on Soil Moisture and Soybean Growth and Yield under Different Tillage Systems. Soil Tillage Res. 2019, 195, 104430. [Google Scholar] [CrossRef]
- Pietikäinen, J.; Pettersson, M.; Bååth, E. Comparison of Temperature Effects on Soil Respiration and Bacterial and Fungal Growth Rates. FEMS Microbiol. Ecol. 2005, 52, 49–58. [Google Scholar] [CrossRef]
- Young, I.M.; Ritz, K. Tillage, Habitat Space and Function of Soil Microbes. Soil Tillage Res. 2000, 53, 201–213. [Google Scholar] [CrossRef]
- Smith, C.R.; Blair, P.L.; Boyd, C.; Cody, B.; Hazel, A.; Hedrick, A.; Kathuria, H.; Khurana, P.; Kramer, B.; Muterspaw, K.; et al. Microbial Community Responses to Soil Tillage and Crop Rotation in a Corn/Soybean Agroecosystem. Ecol. Evol. 2016, 6, 8075–8084. [Google Scholar] [CrossRef]
- Van Eerd, L.L.; Congreves, K.A.; Hayes, A.; Verhallen, A.; Hooker, D.C. Long-Term Tillage and Crop Rotation Effects on Soil Quality, Organic Carbon, and Total Nitrogen. Can. J. Soil Sci. 2014, 94, 303–315. [Google Scholar] [CrossRef]
- McDaniel, M.D.; Tiemann, L.K.; Grandy, A.S. Does Agricultural Crop Diversity Enhance Soil Microbial Biomass and Organic Matter Dynamics? A Meta-Analysis. Ecol. Appl. 2014, 24, 560–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mondal, S.; Mishra, J.S.; Poonia, S.P.; Kumar, R.; Dubey, R.; Kumar, S.; Verma, M.; Rao, K.K.; Ahmed, A.; Dwivedi, S.; et al. Can Yield, Soil C and Aggregation Be Improved under Long-Term Conservation Agriculture in the Eastern Indo-Gangetic Plain of India? Eur. J. Soil Sci. 2021, 72, 1742–1761. [Google Scholar] [CrossRef]
- Hartman, K.; van der Heijden, M.G.A.; Wittwer, R.A.; Banerjee, S.; Walser, J.-C.; Schlaeppi, K. Cropping Practices Manipulate Abundance Patterns of Root and Soil Microbiome Members Paving the Way to Smart Farming. Microbiome 2018, 6, 14. [Google Scholar] [CrossRef]
- Sun, R.; Li, W.; Dong, W.; Tian, Y.; Hu, C.; Liu, B. Tillage Changes Vertical Distribution of Soil Bacterial and Fungal Communities. Front. Microbiol. 2018, 9, 699. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Sun, J.; Li, L.; Wang, X.; Li, X.; Qu, J. Effect of Soybean and Maize Rotation on Soil Microbial Community Structure. Agronomy 2019, 9, 42. [Google Scholar] [CrossRef] [Green Version]
- Kodadinne Narayana, N.; Kingery, W.L.; Shankle, M.W.; Ganapathi Shanmugam, S. Differential Response of Soil Microbial Diversity and Community Composition Influenced by Cover Crops and Fertilizer Treatments in a Dryland Soybean Production System. Agronomy 2022, 12, 618. [Google Scholar] [CrossRef]
- Hannula, S.E.; Di Lonardo, D.P.; Christensen, B.T.; Crotty, F.V.; Elsen, A.; van Erp, P.J.; Hansen, E.M.; Rubæk, G.H.; Tits, M.; Toth, Z.; et al. Inconsistent Effects of Agricultural Practices on Soil Fungal Communities across 12 European Long-Term Experiments. Eur. J. Soil Sci. 2021, 72, 1902–1923. [Google Scholar] [CrossRef]
- Grenada Series. Grenada Silt Loam Official Soil Series Description. National Cooperative Soil Survey Official Series Description. Available online: https://soilseries.sc.egov.usda.gov/OSD_Docs/G/GRENADA.html. (accessed on 6 September 2022).
- NOAA. Climate Normals. National Centers for Environmental Information. Available online: https://www.ncdc.noaa.gov/data-access/land-based-station-data/land-based-datasets/climate-normals/1981-2010-normals-data. (accessed on 6 September 2022).
- Jacobs, A.; Barrett, R.; Williams, M.; Garner, E.R. A Tool for Selecting Cover Crops for Row Crop Rotations in the Southeast. Available online: https://www.nrcs.usda.gov/wps/portal/nrcs/detail/plantmaterials/home/?cid=nrcseprd1340654. (accessed on 26 September 2022).
- Jacobs, A.A.; Evans, R.S.; Allison, J.K.; Garner, E.R.; Kingery, W.L.; McCulley, R.L. Cover Crops and No-Tillage Reduce Crop Production Costs and Soil Loss, Compensating for Lack of Short-Term Soil Quality Improvement in a Maize and Soybean Production System. Soil Tillage Res. 2022, 218, 105310. [Google Scholar] [CrossRef]
- Caporaso, J.; Lauber, C.; Walters, W.; Berg-Lyons, D.; Lozupone, C.; Turnbaugh, P.; Fierer, N.; Knight, R. Global Patterns of 16S RRNA Diversity at a Depth of Millions of Sequences per Sample. Proc. Natl. Acad. Sci. USA 2011, 108, 4516–4522. [Google Scholar] [CrossRef]
- Orgiazzi, A.; Lumini, E.; Nilsson, R.H.; Girlanda, M.; Vizzini, A.; Bonfante, P.; Bianciotto, V. Unravelling Soil Fungal Communities from Different Mediterranean Land-Use Backgrounds. PLoS ONE 2012, 7, e34847. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; Sankaran, K.; Fukuyama, J.A.; McMurdie, P.J.; Holmes, S.P. Bioconductor Workflow for Microbiome Data Analysis: From Raw Reads to Community Analyses 2016. F1000Research 2016, 5, 1492. [Google Scholar] [CrossRef] [PubMed]
- Dhariwal, A.; Chong, J.; Habib, S.; King, I.L.; Agellon, L.B.; Xia, J. MicrobiomeAnalyst: A Web-Based Tool for Comprehensive Statistical, Visual and Meta-Analysis of Microbiome Data. Nucleic Acids Res. 2017, 45, W180–W188. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.; Liu, P.; Zhou, G.; Xia, J. Using MicrobiomeAnalyst for Comprehensive Statistical, Functional, and Meta-Analysis of Microbiome Data. Nat. Protoc. 2020, 15, 799–821. [Google Scholar] [CrossRef] [PubMed]
- Sikora, F.J.; Kissel, D.E. Soil pH. In Soil Test Methods from the Southeastern United States; Sikora, F.J., Moore, K.P., Eds.; University of Georgia: Athens, GA, USA, 2014; p. 48. [Google Scholar]
- Wang, J.J.; Provin, T.; Zhang, H. Measurement of Soil Salinity and Sodicity. In Soil Test Methods from the Southeastern United States; Sikora, F.J., Moore, K.P., Eds.; University of Georgia: Athens, GA, USA, 2014; p. 185. [Google Scholar]
- Zhang, H.; Hardy, D.H.; Mylavarapu, R.; Wang, J.J. Mehlich-3. In Soil Test Methods from the Southeastern United States; Sikora, F.J., Moore, K.P., Eds.; University of Georgia: Athens, GA, USA, 2014; p. 101. [Google Scholar]
- Zhang, H.; Wang, J.J. Loss on Ignition. In Soil Test Methods from the Southeastern United States; Sikora, F.J., Moore, K.P., Eds.; University of Georgia: Athens, GA, USA, 2014; p. 155. [Google Scholar]
- Barnhisel, R.I.; Bertsch, P.M. Chlorites and Hydroxy-Interlayered Vermiculite and Smectite. In Minerals in Soil Environments; Soil Science Society of America Inc.: Madison, WI, USA, 1989; pp. 729–788. [Google Scholar]
- Jensen, J.L.; Christensen, B.T.; Schjønning, P.; Watts, C.W.; Munkholm, L.J. Converting Loss-on-Ignition to Organic Carbon Content in Arable Topsoil: Pitfalls and Proposed Procedure. Eur. J. Soil Sci. 2018, 69, 604–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynn, W.C.; Ahrens, R.J.; Smith, A.L. Soil Minerals, Their Geographic Distribution, and Soil Taxonomy. In Soil Mineralogy with Environmental Applications; Soil Science Society of America Inc.: Madison, WI, USA, 2002; pp. 691–709. [Google Scholar]
- Huluka, G.; Miller, R. Particle Size Determination by Hydrometer Method. In Soil Test Methods from the Southeastern United States; Sikora, F.J., Moore, K.P., Eds.; University of Georgia: Athens, GA, USA, 2014; pp. 180–184. [Google Scholar]
- Blaud, A.; Menon, M.; van der Zaan, B.; Lair, G.J.; Banwart, S.A. Effects of Dry and Wet Sieving of Soil on Identification and Interpretation of Microbial Community Composition. In Advances in Agronomy; Banwart, S.A., Sparks, D.L., Eds.; Academic Press: Cambridge, MA, USA, 2017; Volume 142, pp. 119–142. [Google Scholar]
- He, Y.; Zhou, B.-J.; Deng, G.-H.; Jiang, X.-T.; Zhang, H.; Zhou, H.-W. Comparison of Microbial Diversity Determined with the Same Variable Tag Sequence Extracted from Two Different PCR Amplicons. BMC Microbiol. 2013, 13, 208. [Google Scholar] [CrossRef] [Green Version]
- Lupwayi, N.Z.; Clayton, G.W.; O’Donovan, J.T.; Harker, K.N.; Turkington, T.K.; Rice, W.A. Soil Microbiological Properties during Decomposition of Crop Residues under Conventional and Zero Tillage. Can. J. Soil. Sci. 2004, 84, 411–419. [Google Scholar] [CrossRef]
- Wipf, H.M.-L.; Xu, L.; Gao, C.; Spinner, H.B.; Taylor, J.; Lemaux, P.; Mitchell, J.; Coleman-Derr, D. Agricultural Soil Management Practices Differentially Shape the Bacterial and Fungal Microbiomes of Sorghum Bicolor. Appl. Environ. Microbiol. 2021, 87, e02345-20. [Google Scholar] [CrossRef]
- Navarro-Noya, Y.E.; Gómez-Acata, S.; Montoya-Ciriaco, N.; Rojas-Valdez, A.; Suárez-Arriaga, M.C.; Valenzuela-Encinas, C.; Jiménez-Bueno, N.; Verhulst, N.; Govaerts, B.; Dendooven, L. Relative Impacts of Tillage, Residue Management and Crop-Rotation on Soil Bacterial Communities in a Semi-Arid Agroecosystem. Soil Biol. Biochem. 2013, 65, 86–95. [Google Scholar] [CrossRef]
- Schmidt, R.; Gravuer, K.; Bossange, A.V.; Mitchell, J.; Scow, K. Long-Term Use of Cover Crops and No-till Shift Soil Microbial Community Life Strategies in Agricultural Soil. PLoS ONE 2018, 13, e0192953. [Google Scholar] [CrossRef] [PubMed]
- Cloutier, M.L.; Murrell, E.; Barbercheck, M.; Kaye, J.; Finney, D.; García-González, I.; Bruns, M.A. Fungal Community Shifts in Soils with Varied Cover Crop Treatments and Edaphic Properties. Sci. Rep. 2020, 10, 6198. [Google Scholar] [CrossRef] [Green Version]
- Madegwa, Y.M.; Uchida, Y. Land Use and Season Drive Changes in Soil Microbial Communities and Related Functions in Agricultural Soils. Environ. DNA 2021, 3, 1214–1228. [Google Scholar] [CrossRef]
- Broeckling, C.D.; Broz, A.K.; Bergelson, J.; Manter, D.K.; Vivanco, J.M. Root Exudates Regulate Soil Fungal Community Composition and Diversity. Appl. Environ. Microbiol. 2008, 74, 738–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haichar, F.e.Z.; Marol, C.; Berge, O.; Rangel-Castro, J.I.; Prosser, J.I.; Balesdent, J.; Heulin, T.; Achouak, W. Plant Host Habitat and Root Exudates Shape Soil Bacterial Community Structure. ISME J. 2008, 2, 1221–1230. [Google Scholar] [CrossRef] [PubMed]
- Benitez, M.-S.; Ewing, P.M.; Osborne, S.L.; Lehman, R.M. Rhizosphere Microbial Communities Explain Positive Effects of Diverse Crop Rotations on Maize and Soybean Performance. Soil Biol. Biochem. 2021, 159, 108309. [Google Scholar] [CrossRef]
- Ritz, K.; Young, I.M. Interactions between Soil Structure and Fungi. Mycologist 2004, 18, 52–59. [Google Scholar] [CrossRef] [Green Version]
- Schlatter, D.C.; Kahl, K.; Carlson, B.; Huggins, D.R.; Paulitz, T. Fungal Community Composition and Diversity Vary with Soil Depth and Landscape Position in a No-till Wheat-Based Cropping System. FEMS Microbiol. Ecol. 2018, 94, fiy098. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Liu, L.; Chen, Q.; Wen, X.; Liao, Y. Conservation Tillage Increases Soil Bacterial Diversity in the Dryland of Northern China. Agron. Sustain. Dev. 2016, 36, 28. [Google Scholar] [CrossRef] [Green Version]
- Lozupone, C.; Knight, R. UniFrac: A New Phylogenetic Method for Comparing Microbial Communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef]
- Lozupone, C.A.; Hamady, M.; Kelley, S.T.; Knight, R. Quantitative and Qualitative β Diversity Measures Lead to Different Insights into Factors That Structure Microbial Communities. Appl. Environ. Microbiol. 2007, 73, 1576–1585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, X.; Tao, B.; Guo, J.; Li, J.; Chen, G. Changes in the Microbial Community Structure and Soil Chemical Properties of Vertisols Under Different Cropping Systems in Northern China. Front. Environ. Sci. 2018, 6, 132. [Google Scholar] [CrossRef] [Green Version]
- Thomas, G.A.; Dalal, R.C.; Standley, J. No-till Effects on Organic Matter, PH, Cation Exchange Capacity and Nutrient Distribution in a Luvisol in the Semi-Arid Subtropics. Soil Tillage Res. 2007, 94, 295–304. [Google Scholar] [CrossRef]
- López-Fando, C.; Pardo, M.T. Changes in Soil Chemical Characteristics with Different Tillage Practices in a Semi-Arid Environment. Soil Tillage Res. 2009, 104, 278–284. [Google Scholar] [CrossRef]
- Li, Y.; Li, Z.; Cui, S.; Zhang, Q. Trade-off between Soil PH, Bulk Density and Other Soil Physical Properties under Global No-Tillage Agriculture. Geoderma 2020, 361, 114099. [Google Scholar] [CrossRef]
- Vukicevich, E.; Lowery, T.; Bowen, P.; Úrbez-Torres, J.R.; Hart, M. Cover Crops to Increase Soil Microbial Diversity and Mitigate Decline in Perennial Agriculture. A Review. Agron. Sustain. Dev. 2016, 36, 48. [Google Scholar] [CrossRef] [Green Version]
- Ullah, S.; Ai, C.; Ding, W.; Jiang, R.; Zhao, S.; Zhang, J.; Zhou, W.; Hou, Y.; He, P. The Response of Soil Fungal Diversity and Community Composition to Long-Term Fertilization. Appl. Soil Ecol. 2019, 140, 35–41. [Google Scholar] [CrossRef]
- Rousk, J.; Brookes, P.C.; Bååth, E. Contrasting Soil PH Effects on Fungal and Bacterial Growth Suggest Functional Redundancy in Carbon Mineralization. Appl. Environ. Microbiol. 2009, 75, 1589–1596. [Google Scholar] [CrossRef] [Green Version]
- Tedersoo, L.; Bahram, M.; Põlme, S.; Kõljalg, U.; Yorou, N.S.; Wijesundera, R.; Ruiz, L.V.; Vasco-Palacios, A.M.; Thu, P.Q.; Suija, A.; et al. Global Diversity and Geography of Soil Fungi. Science 2014, 346, 1256688. [Google Scholar] [CrossRef] [Green Version]
- Kemmitt, S.J.; Lanyon, C.V.; Waite, I.S.; Wen, Q.; Addiscott, T.M.; Bird, N.R.A.; O’Donnell, A.G.; Brookes, P.C. Mineralization of Native Soil Organic Matter Is Not Regulated by the Size, Activity or Composition of the Soil Microbial Biomass—A New Perspective. Soil Biol. Biochem. 2008, 40, 61–73. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Alpha Diversity Indices | ||
| Treatments | Shannon | Richness (Chao1) |
| CT-B | 4.14 (0.18) | 78.41 (13.79) |
| CT-CC | 3.89 (0.60) | 71.91 (47.63) |
| CT-WW | 4.25 (0.34) | 90.01 (31.04) |
| NT-B | 4.40 (0.36) | 108.03 (39.83) |
| NT-CC | 3.82 (0.19) | 53.33 (11.23) |
| NT-WW | 4.14 (0.20) | 77.33 (15.04) |
| p-value | 0.29 | 0.32 |
| Fungal Alpha Diversity Indices | ||
| CT-B | 3.33 (0.47) | 117.66 (9.38) |
| CT-CC | 4.06 (0.19) | 140.09 (20.14) |
| CT-WW | 3.48 (0.86) | 113.92 (25.12) |
| NT-B | 3.71 (0.34) | 115.34 (10.55) |
| NT-CC | 3.52 (0.87) | 129.03 (13.71) |
| NT-WW | 4.07 (0.15) | 132.21 (10.95) |
| p-value | 0.37 | 0.30 |
| Treatments | Ascomycota (%) | Basidiomycota (%) | Mortierellomycota (%) | Chytridiomycota (%) | Glomeromycota (%) |
|---|---|---|---|---|---|
| CT-B | 39.50 (6.36) d | 44.23 (11.99) | 10.23 (4.93) ab | 1.38 (0.84) | 0.37 (0.29) |
| CT-CC | 62.82 (4.91) bc | 21.88 (2.92) | 5.99 (1.47) bc | 3.77 (1.03) | 0.36 (0.15) |
| CT-WW | 48.06 (21.15) cd | 29.72 (30.62) | 13.90 (4.95) a | 2.89 (2.15) | 0.61 (0.54) |
| NT-B | 71.90 (1.42) ab | 11.86 (5.37) | 4.82 (2.74) bc | 2.42 (1.23) | 1.13 (0.59) |
| NT-CC | 84.08 (2.27) a | 7.13 (1.61) | 2.83 (0.94) c | 1.87 (0.57) | 0.66 (0.68) |
| NT-WW | 66.59 (2.40) b | 12.14 (0.99) | 6.70 (1.58) bc | 6.08 (3.00) | 1.36 (0.56) |
| p-value | <0.001 *** | 0.05 * | 0.013 * | 0.056 | 0.171 |
| Treatment Systems | pH | EC (dS/cm) | Total C (%) | P (mg/kg) | K (mg/kg) | Zn (mg/kg) | Sand (%) | Silt (%) | Clay (%) | Bulk Density (g cm−3) | Agg. Size 100 Mesh (g/g) | Agg. Size 100 Mesh (g/g) | Agg. Size 100 Mesh (g/g) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CT-B | 6.3 (0.6) a | 0.3 (0.1) | 0.5 (0.3) | 21.0 (7.0) | 69.8 (22.2) | 1.3 (0.4) | 24 (7) | 59 (5) | 17 (3) | 1.26 (0.07) bc | 5.00 (1.23) | 1.40 (0.28) | 2.45 (0.53) |
| CT-CC | 6.0 (0.3) ab | 0.2 (0.1) | 0.5 (0.1) | 22.0 (2.9) | 62.3 (12.1 | 1.3 (0.2) | 16 (8) | 67 (4) | 18 (3) | 1.20 (0.03) c | 6.40 (1.72) | 1.15 (0.34) | 2.30 (0.62) |
| CT-WW | 6.2 (0.6) a | 0.3 (0.1) | 0.4 (0.0) | 19.5 (3.0) | 68.5 (8.4) | 1.2 (0.2) | 25 (10) | 58 (9) | 18 (2) | 1.30 (0.06) bc | 5.10 (0.26) | 1.15 (0.34) | 2.20 (0.33) |
| NT-B | 6.0 (0.3) ab | 0.3 (0.1) | 0.4 (0.0) | 23.5 (2.6) | 62.8 (14.8) | 1.4 (0.3) | 15 (5) | 67 (5) | 19 (1) | 1.43 (0.04) a | 5.30 (1.29) | 1.10 (0.26) | 2.30 (0.20) |
| NT-CC | 5.8 (0.4) b | 0.2 (0.1) | 0.6 (0.1) | 19.5 (3.1) | 57.8 (4.3) | 1.3 (0.2) | 23 (13) | 59 (11) | 19 (2) | 1.36 (0.07) ab | 4.90 (0.35) | 1.60 (0.37) | 2.70 (0.26) |
| NT-WW | 6.2 (0.3) ab | 0.3 (0.1) | 0.5 (0.1) | 23.0 (5.6) | 70.5 (26.4) | 1.2 (0.3) | 16 (4) | 67 (4) | 18 (2) | 1.46 (0.09) a | 5.45 (0.96) | 1.25 (0.34) | 2.50 (0.20) |
| p-value | 0.005 ** | 0.475 | 0.167 | 0.426 | 0.821 | 0.767 | 0.355 | 0.209 | 0.934 | <0.001 *** | 0.319 | 0.323 | 0.314 |
| pH | EC | C | P | K | Zn | Bulk Density | Aggregate Stability | |
|---|---|---|---|---|---|---|---|---|
| R | 0.336 | −0.141 | 0.179 | −0.001 | −0.053 | 0.041 | 0.032 | −0.071 |
| p value | 0.014 * | 0.301 | 0.333 | 0.989 | 0.667 | 0.683 | 0.719 | 0.633 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Narayana, N.K.; Kingery, W.L.; Jacobs, A.A.; Allison, J.K.; Ganapathi Shanmugam, S. Effects of Tillage and Winter Cover Management in a Maize Soybean Rotation on Soil Bacterial and Fungal Community Composition. Land 2022, 11, 2259. https://0-doi-org.brum.beds.ac.uk/10.3390/land11122259
Narayana NK, Kingery WL, Jacobs AA, Allison JK, Ganapathi Shanmugam S. Effects of Tillage and Winter Cover Management in a Maize Soybean Rotation on Soil Bacterial and Fungal Community Composition. Land. 2022; 11(12):2259. https://0-doi-org.brum.beds.ac.uk/10.3390/land11122259
Chicago/Turabian StyleNarayana, Nisarga Kodadinne, William L. Kingery, Alayna A. Jacobs, Jon K. Allison, and Shankar Ganapathi Shanmugam. 2022. "Effects of Tillage and Winter Cover Management in a Maize Soybean Rotation on Soil Bacterial and Fungal Community Composition" Land 11, no. 12: 2259. https://0-doi-org.brum.beds.ac.uk/10.3390/land11122259