
The Wetlands of Northeastern Algeria (Guelma and Souk Ahras): Stakes for the Conservation of Regional Biodiversity
 , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
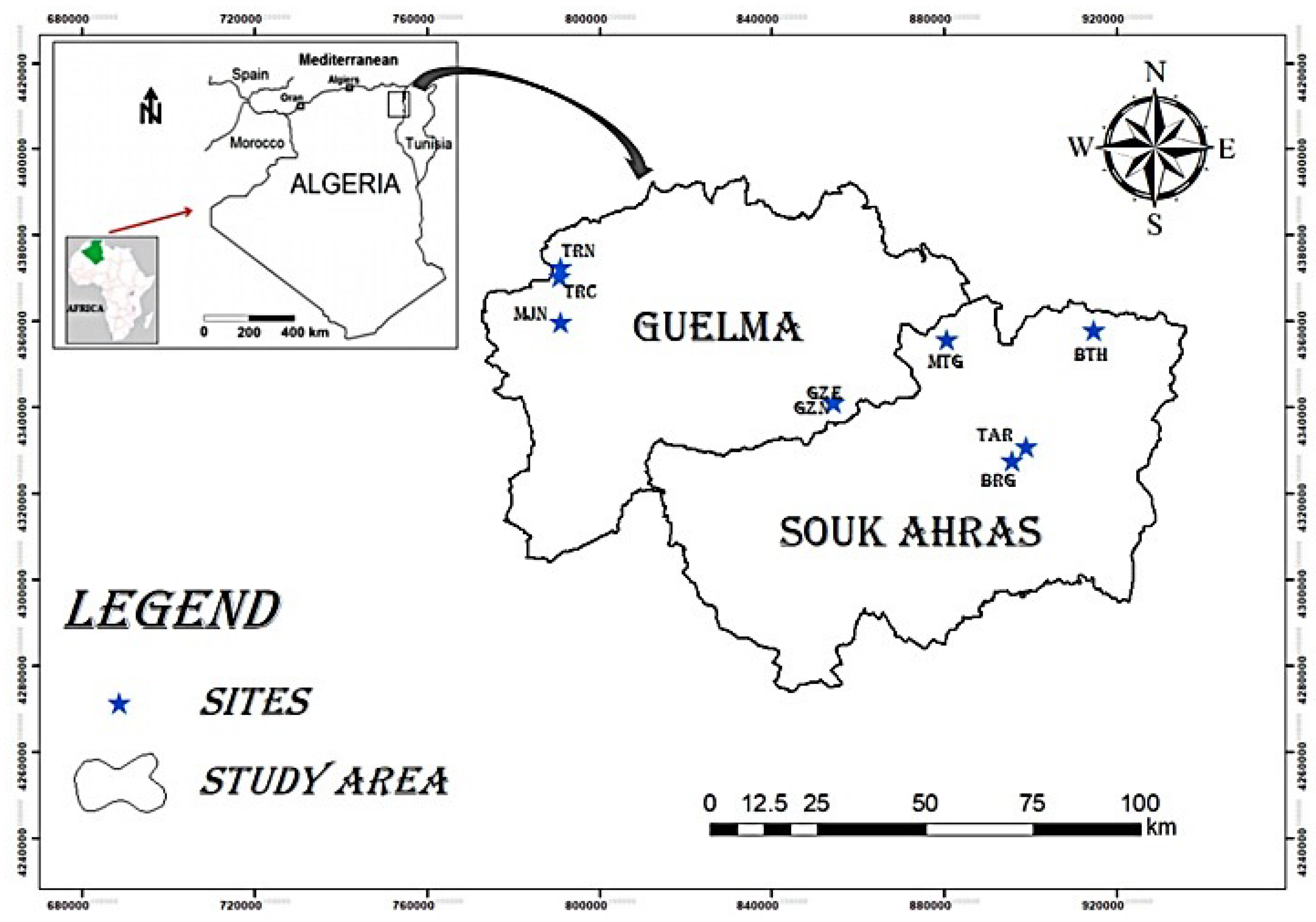
2.1. Study Area
2.2. Methodology
2.2.1. Soil Analysis
2.2.2. Water Analysis
2.2.3. Measurement of Other Environmental Variables
2.2.4. Floristic Study
2.2.5. Statistical Analysis
3. Results
3.1. Hydro-Edaphic Characteristics
3.2. Other Environmental Variables Measurement
3.3. Floristic Diversity
- Forestry species, such as Quercus suber L., Erica arborea L., Cytisus villosus Pourr., Crataegus monogyna Jacq., Myrtus communis L., Pistacia lentiscus L. and Olea europaea L.
- Lawn species, including both temporary-pond species (Cyperus rotundus L. subsp. rotundus, Silene laeta (Aiton) Godr., Isoetes histrix Durieu ex Bory, Typha domingensis Pers., Potentilla reptans L. and Illecebrum verticillatum L.) and therophitic pelouse species (Catapodium rigidum (L.) C.E. Hubb., Cerastium glomeratum Thuill., Ranunculus muricatus L., Medicago murex Willd., Trifolium arvense L., Plantago lanceolata L., Stellaria media (L.) Vill. and Geranium dissectum L.).
- Hydrophytes species, such as Lemna minor L., Ranunculus aquatilis L. and Callitriche obtusangula Le Gall.
- Hygrophytic species, such as Nasturtium officinale R. Br., Lythrum junceum Banks & Sol., Schoenus nigricans L., Juncus bufonius L., Cotula coronopifolia L., Mentha aquatica L. and Mentha pulegium L.
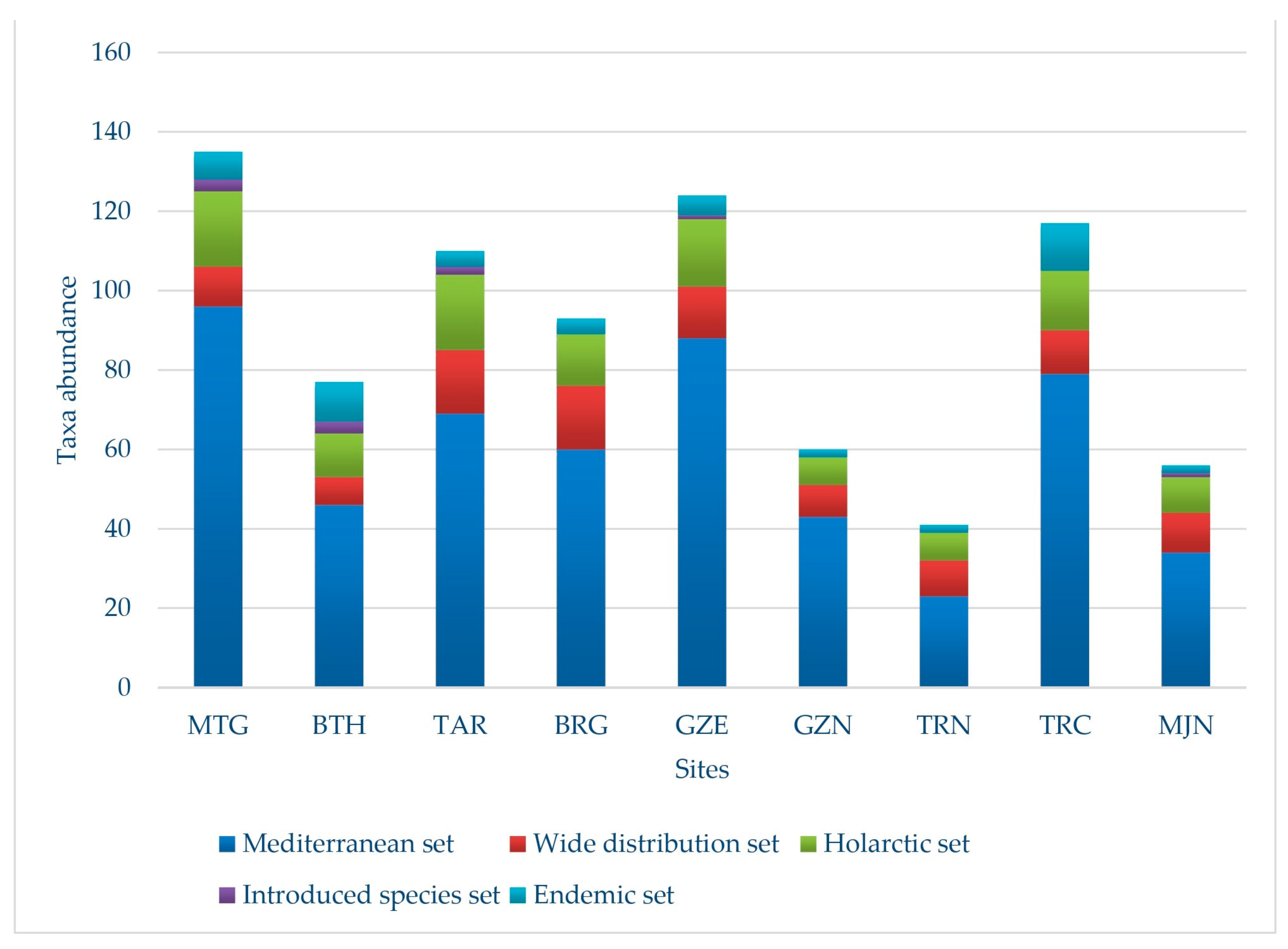
3.4. Biogeographic Distribution
- The Mediterranean set: the most numerous group with 210 taxa (66.25%), of which 165 correspond to strictly Mediterranean elements, 34 to euri-Mediterranean connecting elements, and 11 to Mediterranean-Atlantic connecting elements. In this group, the richest families are best represented, such as Fabaceae and Poaceae, with 26 and 15 taxa respectively, compared to the 31 taxa of the Asteraceae family.
- The widespread distribution set: Within this classification, there are 31 taxa, constituting 9.78% of the studied flora. Among them, 12 are cosmopolitan taxa and 19 are subcosmopolitan taxa.
- The Holarctic set: these species represent 12.93% (41 taxa) of the total flora. The paleotemperate element, with 24 taxa, is followed by the Holarctic element with 8 taxa; 6 taxa are Eurasian, and finally, 3 taxa originate from the tropics.
- The introduced species set: 2.21% belong to this group, totaling 7 species, of which 5 are listed as introduced species, one as naturalized and another as cultivated.
- The endemic species set: 28 regional endemic taxa are found in the study ponds, accounting for 8.83% of the total. With 11 species, i.e., 42.3% of the region’s endemic flora, Algerian-Tunisian endemic taxa are the most abundant. The families Apaiceae, Asteraceae, and Lamiaceae have the best representation with 4 taxa each, followed by Hyacinthaceae with 3 endemic species. Nine families are identified with a single endemic taxon.
3.5. Shannon Index and Its Relation to Ecological Variables
3.6. Botanical Heritage
3.7. Taxonomic Distinctness (TD)
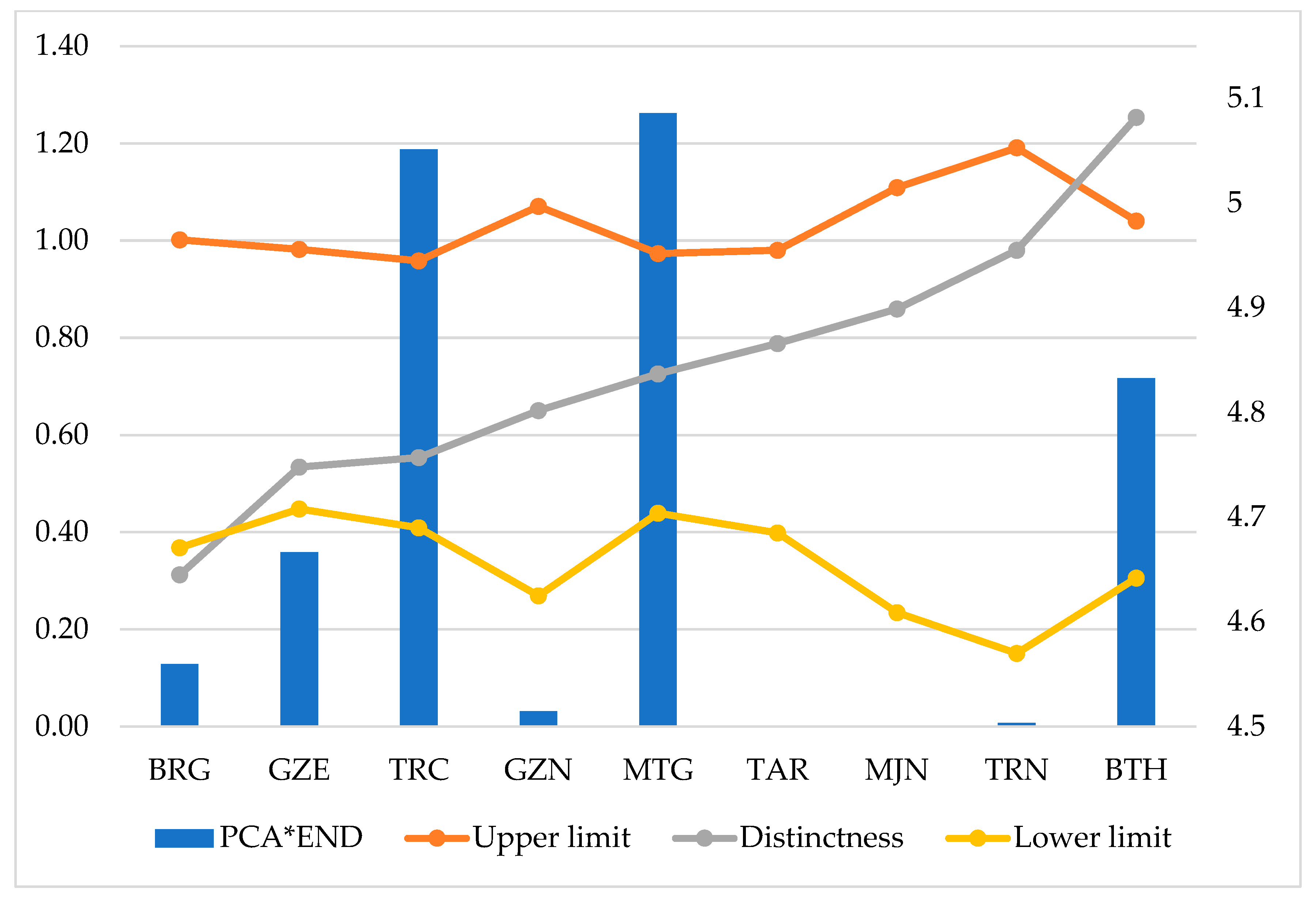
3.8. Priority Conservation Areas Based on Endemism (PCA_END)
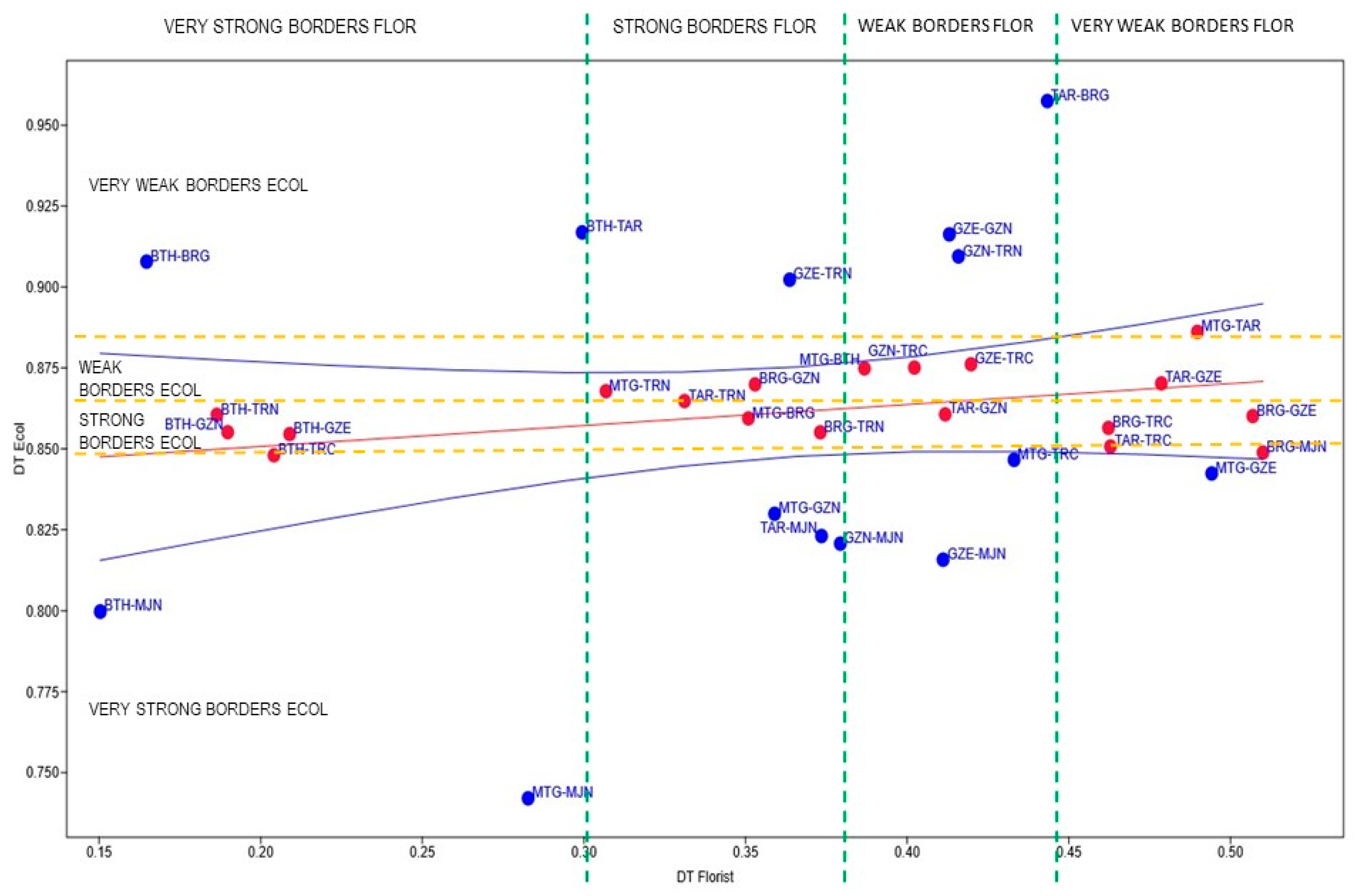
3.9. Estimation of Ecofloristic Boundaries
4. Discussion
4.1. Influence of Hydropedological Variables on Floristic Diversity
4.2. Floristic and Phytogeographical Diversity
4.3. Conservation of Plant Heritage
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Véla, E.; Benhouhou, S. Évaluation d’un nouveau point chaud de biodiversité végétale dans le Bassin méditerranéen (Afrique du Nord). Comptes Rendus Biol. 2007, 330, 589–605. [Google Scholar] [CrossRef] [PubMed]
- Samraoui, B.; de Bélair, G. The Guerbes-Senhadja wetlands. Part I: An overview. Ecologie 1997, 28, 233–250. [Google Scholar]
- Médail, F.; Michaud, H.; Molina, J.; Paradis, G.; Loisel, R. Conservation de la flore et de la végétation des mares temporaires dulçaquicoles et oligotrophes de France méditerranéenne. Ecol. Mediterr. 1998, 24, 119–134. [Google Scholar] [CrossRef]
- Ghrabi-Gammar, Z.; Daoud-Bouattour, A.; Ferchichi, H.; Gammar, A.M.; Muller, S.D.; Rhazi, L.; Limam-Ben Saad, S. Flore vasculaire rare, endémique et menacée des zones humides de Tunisie. Rev. DÉcologie Terre Vie 2009, 64, 19–40. [Google Scholar] [CrossRef]
- Rouissi, M.; Boix, D.; Muller, S.D.; Gascon, S.; Ruhi, A.; Sala, J.; Bouattour, A.; Ben Haj Jilani, I.; Ghrabi-Gammar, Z.; Ben Saad-Limam, S.; et al. Spatio-temporal variability of faunal and floral assemblages in Mediterranean temporary wetlands. Comptes Rendus Biol. 2014, 337, 695–708. [Google Scholar] [CrossRef] [PubMed]
- Zouaidia, H.; de Bélair, G.; Samraoui, B. Biodiversité et écologie des mares temporaires de la Numidie orientale; Nord-est Algérien: Cas du complexe des mares Gauthier. Agrobiologia 2021, 11, 2457–2469. [Google Scholar]
- Rifai, N.; Khattabi, A.; Moukrim, S.; Arahou, M.; Rhazi, L. Évaluation de la dynamique de l’occupation du sol dans la zone humide RAMSAR de Tahaddart (Nord-Ouest du Maroc). Rev. DÉcologie Terre Vie 2018, 73, 142–152. [Google Scholar] [CrossRef]
- Chillasse, L.; Dakki, M. Potentialités et statuts de conservation des zones humides du Moyen-Atlas (Maroc), avec référence aux influences de la sécheresse. Sécheresse 2004, 15, 337–345. [Google Scholar]
- Perennou, C.; Beltrame, C.; Guelmami, A.; Tomàs Vives, P.; Caessteker, P. Existing areas and past changes of wetland extent in the Mediterranean region: An overview. Ecol. Mediterr. 2012, 38, 53–66. [Google Scholar] [CrossRef]
- Davidson, N.C. How much wetland has the world lost? Long-term and recent trends in global wetland area. Mar. Freshw. Res. 2014, 65, 934–941. [Google Scholar]
- Chedad, A.; Bendjoudi, D.; Guezoul, O. Biodiversité de l’avifaune aquatique d’une zone humide artificielle à Kef Doukhane (Ghardaia, Sahara Algérien). Bull. Société. Zool. Fr. 2020, 145, 383–400. [Google Scholar]
- Benzina, I.; Si Bachir, A.; Perennou, C. Waterbird diversity at the Sebkhet of Aures wetlands complex, North East Algeria. Ornithol. Beob. 2022, 119, 318–329. [Google Scholar]
- Quézel, P.; Santa, S. Nouvelle Flore de L’algérie et des Régions Désertiques Méridionales, 2nd ed.; CNRS: Paris, France, 1962–1963; pp. 1–1170. [Google Scholar]
- Rhazi, L.; Rhazi, M.; Grillas, P.; Khyari, D.E. Richness and structure of plant communities in temporary pools from western Morocco: Influence of human activities. Hydrobiologia 2006, 570, 197–203. [Google Scholar] [CrossRef]
- Ferchichi Ben Jamaa, H.; Serge, D.; Muller, S.D.; Ghrabi-Gammar, Z.; Rhazi, L.; Soulié-Märsche, I.; Gammar, A.M.; Ouali, M.; Ben Saad-Limam, S.; Daoud-Bouattour, A. Influence du pâturage sur la structure, la composition et la dynamique de la végétation de mares temporaires méditerranéennes (Tunisie septentrionale). Rev. DÉcologie Terre Vie 2014, 69, 196–213. [Google Scholar] [CrossRef]
- Fetnaci, I.; Beddiar, A.; Hamel, T. Le lac Fetzara (Nord-Est algérien): Biodiversité floristique et menaces potentielles. Flora Mediterr. 2019, 29, 227–245. [Google Scholar]
- Carabal, N.; Segura, M.; Puche, E.; Rojo, C.; Rodrigo, M.A. How the diversity of constructed wetlands improves the plankton communities discharged into a protected Mediterranean wetland. Hydrobiologia 2023, 851, 243–259. [Google Scholar] [CrossRef]
- Sissaoui, A.; Baba, R.; Chabi, D. Climate Change and Wetlands in Algeria: Vulnerability and Conservation issues (the case of Beni-Bélaïd wetland-Jijel). J. New Sci. CSIEA 2017, 11, 2678–2683. [Google Scholar]
- Allem, M.; Hamel, T.; Tahraoui, C.; Boulemtafes, A. Diversité floristique des mares temporaires de la région d’Annaba (Nord-Est Algérien). Int. J. Environ. Stud. 2017, 75, 405–424. [Google Scholar] [CrossRef]
- Boukeng, J.E.D. Dynamique et Services des Écosystèmes Agroforestiers Autour D’une Aire Protégée du Cameroun. Ph.D. Thesis, Laval Québec University, Québec City, QC, Canada, 2021. [Google Scholar]
- Belouahem-Abed, D.; Belouahem, F.; Benslama, M.; de Bélair, G.; Muller, S.D. Les aulnaies de Numidie (NE algérien): Biodiversité floristique, vulnérabilité et conservation. Comptes Rendus Biol. 2011, 334, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Boulemtafes, A. Contribution à L’étude de la Végétation Littorale Numidienne: Phytoécologie et Conservation. Ph.D. Thesis, Badji-Mokhtar Annaba University, Annaba, Algeria, 2020. [Google Scholar]
- Hamel, T.; Hidalgo Triana, N.; Meddad-Hamza, A.; Boulemtafes, A.; Souilah, N.; de Bélair, G.; Salvo Tierra, A.E. Analysis of taxonomic distinctness and priority conservation areas as a basis for heritage enhancement of floristic diversity: The case of the ‘hotspot’ of the islands of Numidia (North-eastern Algeria). Mediterr. Bot. 2022, 43, 1–24. [Google Scholar] [CrossRef]
- Hamel, T.; Seridi, R.; de Bélair, G.; Slimani, A.; Babali, B. Flore vasculaire rare et endémique de la péninsule de l’Edough (Nord-Est algerien). Rev. Sci. Technol. Synth. 2013, 26, 65–74. [Google Scholar]
- de Bélair, G.; Véla, E.; Boussouak, R. Inventaire des orchidées de la Numidie (NE Algérie) sur vingt années. J. Eur. Orch. 2005, 37, 291–401. [Google Scholar]
- Hamel, T.; Boulemtafes, A. Découverte d’une endémique tyrrhénienne Soleirolia soleirolii (Urticaceae) en Algérie (Afrique du Nord). Flora Mediterr. 2017, 27, 185–193. [Google Scholar]
- Boulemtafes, A.; Hamel, T.; de Bélair, G.; Véla, E. Nouvelles données sur la distribution et l’écologie de seize taxons végétaux du littoral de la péninsule de l’Edough (Nord–Est algérien). Bull. Soc. Linn. Provence 2018, 69, 1–18. [Google Scholar]
- Hamel, T.; de Bélair, G.; Slimani, A.; Telailia, L.B.; Telailia, S. Nouvelle station de Pteris vittata L. (Pteridaceae) en Numidie (Algérie orientale). Acta Bot. Malacit. 2020, 45, 127–129. [Google Scholar] [CrossRef]
- Saci, A.; Boussaada, Z.; Hamel, T.; de Bélair, G. Première observation d’une endémique tyrrhénienne (Leucojum aestivum subsp. pulchellum, Amaryllidaceae) sur le continent africain (Algérie). Flora Mediterr. 2021, 31, 123–128. [Google Scholar]
- Touati, L.; Hamel, T.; Meddad-Hamza, A.; de Bélair, G. Analysis of rare and endemic flora in northeastern Algeria: The case of the wilaya of Souk Ahras. Bull. Société R. Sci. Liège. 2021, 90, 213–240. [Google Scholar] [CrossRef]
- Seltzer, P.; Auberty, E. Le Climat de L’algérie, 1st ed.; The Institute of Meteorology and Global Physics of Algeria: Algies, Algeria, 1946. [Google Scholar]
- Tlidjane, A.; Menaa, M.; Rebbah, A.C.; Telailia, S.; Seddik, S.; Chefrour, A.; Maazi, M.C. La richesse et la distribution des Amphibiens dans la région de Souk Ahras (Nord-Est de l’Algérie). Bull. Soc. Zool. 2019, 144, 179–201. [Google Scholar]
- Kafi, F.; Hanane, S.; Bensouilah, T.; Zeraoula, A.; Brahmia, H.; Houhamdi, M. Les facteurs déterminant la réussite de la reproduction des tourterelles des bois (Streptopelia turtur) dans un milieu agricole Nord-Africain. Rev. DÉcologie Terre Vie 2015, 70, 271–279. [Google Scholar] [CrossRef]
- Boukehili, K.; Boutabia, L.; Telailia, S.; Menaa, M.; Tlidjane, A.; Maazi, M.C.; Chefrour, A.; Saheb, M.; Véla, E. Les orchidées de la wilaya de souk-ahras (nord-est algérien): Inventaire, écologie, répartition et enjeux de conservation. Rev. DÉcologie Terre Vie 2018, 73, 167–179. [Google Scholar] [CrossRef]
- Jeannot, R.; Lemière, B.; Chiron, S.; Augustin, F.; Darmendrail, D. Guide Méthodologique Pour L’analyse des Sols Pollués, 1st ed.; BRGM: Orléans, France, 2000; p. 41. [Google Scholar]
- Robinson, D.A.; Thomas, A.; Reinsch, S.; Lebron, I.; Feeney, C.J.; Maskell, L.C.; Wood, C.M.; Seaton, F.M.; Emmett, B.A.; Cosby, B.J. Analytical modelling of soil porosity and bulk density across the soil organic matter and land-use continuum. Sci. Rep. 2022, 12, 7085. [Google Scholar] [CrossRef] [PubMed]
- Allison, L.E.; Moodie, C.D. Carbonate. Methods of Soil Analysis: Part 2 Chemical and Microbiological Properties. Am. Soc. Agron. 1983, 9, 595–624. [Google Scholar]
- Liu, Y.; Kumar, M.; Katul, G.G.; Feng, X.; Konings, A.G. Plant hydraulics accentuates the effect of atmospheric moisture stress on transpiration. Nat. Clim. Chang. 2020, 10, 691–695. [Google Scholar] [CrossRef]
- Lv, J.; Huang, Z.; Luo, L.; Zhang, S.; Wang, Y. Advances in Molecular and Microscale Characterization of Soil Organic Matter: Current Limitations and Future Prospects. Environ. Sci. Technol. 2022, 56, 12793–12810. [Google Scholar] [CrossRef] [PubMed]
- Gobat, J.M.; Aragno, M.; Matthey, W. Le sol Vivant: Bases de Pédologie—Biologie des Sols, 3rd ed.; Polytechnic and University Presses Romande: Romande, Italy, 2010; pp. 59–362. [Google Scholar]
- Teghidet, H. Etude de la Cristallisation Contrôlée de la Calcite par Voie Électrochimique. Effet des Ions Étrangers au Système Calcocarbonique sur la Nucléation-Croissance de la Calcite. Ph.D. Thesis, Pierre et Marie Curie-Paris VI University, Paris, France, 2012. [Google Scholar]
- Pousset, J. Engrais Verts et Fertilité des Sols, 2nd ed.; French Agricultural Group: Paris, France, 2002; pp. 53–57. [Google Scholar]
- Ismayilov, A.I.; Mamedov, A.I.; Fujimaki, H.; Tsunekawa, A.; Levy, G.J. Soil salinity type effects on the relationship between the electrical conductivity and salt content for 1:5 soil-to-water extract. Sustainability 2021, 13, 3395. [Google Scholar] [CrossRef]
- Hartemink, A.E.; Barrow, N.J. Soil pH-nutrient relationships: The diagram. Plant Soil 2023, 486, 209–215. [Google Scholar] [CrossRef]
- Baaloudj, A.; Ouarab, S.; Kerfouf, A.; Bouriach, M.; Hussein, A.A.; Hammana, C.; Djeneba, H.D. Use of macro invertebrates to assess the quality of Seybouse River (North-East of Algeria). Ukr. J. Ecol. 2020, 10, 60–66. [Google Scholar]
- Gal, F.; Ghestem, J.P. Détermination du Potentiel D’oxydoréduction dans les Eaux Proposition de Document Normative; Aquaref: Paris France, 2019; pp. 9–11. [Google Scholar]
- Shrestha, A.K.; Basnet, N. The Correlation and Regression Analysis of Physicochemical Parameters of River Water for the Evaluation of Percentage Contribution to Electrical Conductivity. J. Chem. 2018, 2018, 8369613. [Google Scholar] [CrossRef]
- Derwich, E.; Benaabidate, L.; Zian, A.; Sadki, O.; Belghity, D. Caractérisation physico-chimique des eaux de la nappe alluviale du haut Sebou en aval de sa confluence avec oued Fès. Lahryss J. 2010, 8, 101–112. [Google Scholar]
- Aminot, A.; Kérouel, R. Hydrologie des Écosystèmes Marins: Paramètres et Analyses; Ifremer: Brest, France, 2004; pp. 44–303. [Google Scholar]
- Al-hadithi, M. Application of water quality index to assess suitability of groundwater quality for drinking purposes in Ratmao–Pathri Rao watershed, Haridwar District, India. Am. J. Sci. Ind. Res. 2012, 3, 395–402. [Google Scholar] [CrossRef]
- Gaid, K. Traitement de L’eau poTable 1: Qualité des Eaux et Clarification, 1st ed.; ISTE Edition Ltd.: London, UK, 2022; pp. 12–178. [Google Scholar]
- Frontier, S. Stratégies D’échantillonnage en Écologie; Masson: Paris, France, 1983; pp. 50–494. [Google Scholar]
- Battandier, J.A. Flore de L’algérie: Dicotylédones; Typographie Adolphe Jourdan: Paris, France, 1888–1890; pp. 1–866. [Google Scholar]
- Battandier, J.A.; Trabut, L. Flore D’algérie, Contenant la Description de Toutes les Plantes Signalées Jusqu’à ce Jour Comme Spontanées en Algérie et Catalogue des Plantes du Maroc: Monocotylédones; Typographie Adolphe Jourdan: Paris, France, 1895; pp. 1–256. [Google Scholar]
- Maire, R. Flore de l’Afrique du Nord (Maroc, Algérie, Tunisie, Tripolitaine, Cyrénaïque et Sahara), 18th ed.; Paul Lechevalier: Paris, France, 1952–1987; pp. 1–307. [Google Scholar]
- Pignatti, S. Flora d’Italia, 3rd ed.; Edagricole: Bologna, Italy, 1982; pp. 1–1329. [Google Scholar]
- Blanca, G.; Cabezudo, B.; Cueto, M.; Lopez, C.F.; Torres, C.M. Flora Vasculair de Andalucía Oriental, 1st–4th ed.; Consejeria de Medio Ambiente. Junta de Andalucia: Seville, Spain, 2009; pp. 23–1848. [Google Scholar]
- Dobignard, A.; Chatelain, C. Index Synonymique de la Flore d’Afrique du Nord, 1st–5th ed.; Conservatory and Botanical Gardens of Geneve: Geneve, Switzerland, 2010–2013; pp. 8–2236. [Google Scholar]
- APD: African Plants Database; Conservatoire et Jardin botaniques de la Ville de Genève: Geneve, Switzerland; South African National Biodiversity Institute: Pretoria, South Africa, 2023; Available online: https://africanplantdatabase.ch/ (accessed on 25 November 2023).
- Walter, K.S.; Gillett, H.J. 1997 IUCN Red List of Threatened Plants. The IUCN Species Survival Commission: Gland, Switzerland; Cambridge, UK, 1998; pp. 10–862. [Google Scholar]
- IUCN. IUCN Red List of Threatened Plants; Version 2022-2; IUCN: Gland, Switzerland, 2023; Available online: https://www.iucnredlist.org/ (accessed on 25 November 2023).
- ORA, J. Décret exécutif du 18 janvier, complétant la liste des espèces végétales non cultivées et protégées. J. Off. République Algérienne 2012, 51, 1–44. [Google Scholar]
- Raunkiaer, C. The Life Forms of Plants and Statistical Plant Geography; Oxford University Press: London, UK, 1934; pp. 1–632. [Google Scholar]
- Øyvind, H.; David, A.T.H.; Paul, D.R. Past: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- IBM Corp. IBM SPSS Statistics for Windows; Version 28.0; IBM Corp.: Armonk, NY, USA, 2021. [Google Scholar]
- Pérez Hernández, C.X. Distintividad taxonómica: Evaluación de la diversidad en la estructura taxonómica en los ensambles. In La Biodiversidad en un Mundo Cambiante: Fundamentos Teóricos y Metodológicos Para su Estudio; Moreno, C.E., Ed.; Universidad Autónoma del Estado de Hidalgo/Libermex: Ciudad de México, Mexico, 2019; pp. 285–306. [Google Scholar]
- Clarke, K.R.; Warwick, R.M. A taxonomic distinctness index and its statistical properties. J. Appl. Ecol. 1998, 35, 523–531. [Google Scholar] [CrossRef]
- Pereña-Ortiz, J.F.; Salvo-Tierra, Á.E.; Sánchez-Mata, D. Application of Phytosociological Information in the Evaluation of the Management of Protected Areas. Plants 2023, 12, 406. [Google Scholar] [CrossRef]
- Vane-Wright, R.I.; Humphries, C.J.; Williams, P.H. What to protect? Systematics and the agony of choice. Biol. Conserv. 1991, 55, 235–254. [Google Scholar] [CrossRef]
- Marre, A. Le Tell Oriental Algérien de Collo à la Frontière Tunisienne, Étude Géomorphologique; University Publications Office UPO: Algiers, Algeria, 1992; pp. 1–177. [Google Scholar]
- Aibeche, C.; Sidhoum, W.; Djabeur, A.; Kaid-Harche, M. Effet des caractéristiques physico-chimiques sur la charge microbienne de l’eau des zones humides du nord-ouest algérien: Cas du lac Télamine et de Dayet Oum Ghellaz, Oran. Alger. Alger. J. Environ. Sci. Technol. 2020, 6, 1647–1657. [Google Scholar]
- Hade, A. Nos Lacs, les Connaître Pour Mieux les Protéger, 1st ed.; Fides: Montreal, QC, Canada, 2003; pp. 210–221. [Google Scholar]
- Quezel, P. La région méditerranéenne française et ses essences forestières, signification écologique dans le contexte circum-méditerranéen. Forêt Méditerranéenne 1979, 1, 7–18. [Google Scholar]
- Laala, A.; Alatou, D.; Adimi, A. Predicting potential habitat suitability of Quercus suber L. in Algeria under climate change scenarios. Afr. J. Ecol. 2021, 59, 976–987. [Google Scholar] [CrossRef]
- Bennadja, S.; de Bélair, G.; Tlili Ait Kaki, Y. La subéraie de la Numidie orientale: Une source de biodiversité. Quad. Bot. Amb. Appl. 2013, 24, 49–53. [Google Scholar]
- Rhazi, L.; Grillas, P.; Mounirou Touré, A.; Tan Ham, L. Impact of land use and activities on water, sediment and vegetation of temporary pools in Morocco. Comptes Rendus Académie Sci. Life Sci.—Ser. III—Sci. Vie 2001, 324, 165–177. [Google Scholar]
- Jamaa, H.F.B.; Muller, S.D.; Daoud-Bouattour, A.; Ghrabi-Gammar, Z.; Rhazi, L.; Soulié-Märsche, I.; Ouali, M.; Saad-Limam, S.B. Structures de végétation et conservation des zones humides temporaires méditerranéennes: La région des Mogods (Tunisie septentrionale). Comptes Rendus Biol. 2010, 333, 265–279. [Google Scholar] [CrossRef]
- Bouahim, S.; Rhazi, L.; Amami, B.; Waterkeyn, A.; Rhazi, M.; Saber, E.R.; Zouahri, A.; Van Den Broeck, M.; Muller, S.D.; Brendonck, L.; et al. Unravelling the impact of anthropogenic pressure on plant communities in Mediterranean temporary ponds. Mar. Freshw. Res. 2014, 65, 918–929. [Google Scholar] [CrossRef]
- Oertli, B.; Frossard, P.A. Mares et Étangs: Ecologie, Gestion, Aménagement et Valorisation, 1st ed.; Polytechnic and University Presses of Romandy: Romandy, Italy, 2013; pp. 49–72. [Google Scholar]
- Calvet, R. Le Sol: Propriétés et Fonctions—Constitution et Structure, Phénomènes aux Interfaces, 1st ed.; France Agricole-Dunod: Paris, France, 2003; pp. 40–250. [Google Scholar]
- Tahiri, A.; Destain, J.; Druart, P.; Thonart, P. Propriétés physico-chimiques et biologiques des substances humiques en relation avec le développement végétal. Biotechnol. Agron. Soc. Environ. 2014, 18, 436–445. [Google Scholar]
- Montaigne, W.; Debon, H.; Domenach, A.M.; Roggy, J.C. Gestion durable de la fertilité des sols par l’utilisation de matières organiques: Retours d’expérience en Guyane française. Innov. Agron. 2018, 64, 71–82. [Google Scholar]
- Freschet, G.T.; Violle, C.; Roumet, C.; Garnier, E. Interactions Entre le sol et la Végétation: Structure des Communautés de Plantes et Fonctionnement du sol. Les Sols au Cœur de la Zone Critique: Écologie; Lemanceau, P., Blouin, M., Eds.; ISTE: London, UK, 2018; pp. 83–104. [Google Scholar]
- Adi, N.; Amrani, S.; Hirche, A.; Boughani, A.; Nedjraoui, D. Diversité biologique et phytogéographique pour des niveaux différents de salinité dans la région du Chott-Ech-Chergui (sud-ouest de l’Algérie). Rev. DÉcologie Terre Vie 2016, 4, 342–355. [Google Scholar] [CrossRef]
- Hosseini-Nasr, F.; Etesami, H.; Alikhani, H.A. Silicon Improves Plant Growth-Promoting Effect of Nodule Non-Rhizobial Bacterium on Nitrogen Concentration of Alfalfa Under Salinity Stress. J. Soil Sci. Plant Nutr. 2023, 23, 496–513. [Google Scholar] [CrossRef]
- Benslama, M. Les Sols du Complexe Humide de l’Algérie Nord Orientale (Formation, Organisation et Evolution). Ph.D. Thesis, National Agronomic Institute of El Harrach-Algiers, Alger, Algeria, 2007. [Google Scholar]
- Ali Tatar, B.; Touati, L.; Hamel, T.; Mechentel, E.; Badouna, B.; Benslama, M. Phytoécologie, dynamique et reconstitution pollinique de deux mares tourbeuses (Aoural et Ain Salhat) de la petite Kabylie orientale: Cas du bassin versant d’Oued Zhor (Nord-Est algérien). Flora Mediterr. 2023, 33, 193–213. [Google Scholar]
- Williams, D.D. The Biology of Temporary Waters; Oxford University Press: London, UK, 2005. [Google Scholar]
- Megharbi, A.; Abdoun, F.; Belgherbi, B. Diversité floristique en relation avec les gradients abiotiques dans la zone humide de la Macta (Ouest d’Algérie). Rev. DÉcologie Terre Vie 2016, 71, 142–155. [Google Scholar] [CrossRef]
- Belouahem-Abed, D.; Belouahem, F.; de Bélair, G. Biodiversité floristique et vulnérabilité des aulnaies glutineuses de la Numidie algérienne (NE Algérien). Eur. J. Sci. Res. 2009, 32, 329–361. [Google Scholar]
- Djebbouri, M.; Terras, M. Floristic diversity with particular reference to endemic, rare or endangered flora in forest formations of Saïda (Algeria). Int. J. Environ. Stud. 2019, 76, 990–1003. [Google Scholar] [CrossRef]
- Quézel, P. Réflexions sur L’évolution de la Flore et de la Végétation au Maghreb Méditerranéen; Ibis Press: Paris, France, 2000. [Google Scholar]
- Bonnet, B.; Aulong, S.; Goyet, S.; Lutz, M.; Mathevet, R. Gestion Intégrée des Zones Humides Méditerranéennes: Conservation des Zones Humides; Tour de Vallet: Arles, France, 2005. [Google Scholar]
- Bagella, S.; Caria, M.C.; Filigheddu, R.S.; Farris, E.E. Phytosociological analysis in Sardinian Mediterranean temporary wet habitats. Fitosocio 2009, 46, 11–26. [Google Scholar]
- Hamel, T.; Azzouz, Z.; Bellili, A.M.; Boutabia, L.; Telailia, S. L’arctothèque souci (Arctotheca calendula): Une nouvelle espèce exotique pour la flore algérienne. Flora Mediterr. 2020, 30, 137–142. [Google Scholar]
- Vennetier, M. Forêts et changement climatique. Le constat en région méditerranéenne. Sci. Eaux Territ. 2020, 3, 18–25. [Google Scholar] [CrossRef]
- Zouaidia, H.; de Bélair, G.; Benslama, M.; Souile-Maesch, I.; Muller, S.D. Intérêt des Characeae comme bio indicateur de la qualité des eaux: Le cas des zones humides de Numidie (Nord-Est algérien). Rev. DÉcologie Terre Vie 2015, 70, 121–133. [Google Scholar] [CrossRef]
- Grillas, P.; Gauthier, P.; Yavercovski, N.; Perennou, C. Les Mares Temporaires Méditerranéennes, Enjeux de Conservation, Fonctionnement et Gestion, 1st ed.; Tour du Valat: Arles, France, 2004; pp. 1–120. [Google Scholar]
- Zedam, A.; Khoudour, D.; Mimeche, F.A. RAMSAR wetland’s endemic flora. inventory and conservation of Chott El Hodna’s plants (Northeastern Algeria). Pak. J. Bot. 2022, 54, 1025–1032. [Google Scholar] [CrossRef] [PubMed]
- Meddour-Sahar, O.; Bouisset, C. Les grands incendies de forêt en Algérie: Problèmes humains et politiques publiques dans la gestion des risques. Méditerranée 2013, 121, 33–40. [Google Scholar] [CrossRef]
- Francisco, L.; Paul, Z. The effect of forest fire on vegetation. In Fire Effects on Soils and Restoration Strategies; Science Publishers: Enfield, NH, USA, 2009; pp. 321–322. [Google Scholar]
- Laala, A.; Alatou, D. Interpolation spatiale des températures mensuelles en Algérie orientale. La Météorologie 2017, 98, 45–51. [Google Scholar] [CrossRef]
- Ricklefs, R.E.; Relyea, R. Écologie: L’économie de la Nature, 1st ed.; De Boeck Supérieur: Louvain-la-Neuve, Belgium, 2019; pp. 190–195. [Google Scholar]
- Rhazi, L.; Grillas, P.; Rhazi, M.; Aznar, J.C. Ten-year dynamics of vegetation in a Mediterranean temporary pool in western Morocco. Hydrobiologia 2010, 634, 185–194. [Google Scholar] [CrossRef]
- Chauvel, B.; Darmency, H.; Munier-Jolain, N.; Rodriguez, A. Gestion Durable de la Flore Adventice des Cultures; Quae: Versailles, France, 2018; pp. 188–193. [Google Scholar]
- Médail, F.; Diadema, K. Biodiversité végétale méditerranéenne et anthropisation: Approches macro et micro-régionales. Ann. Géographie 2006, 651, 618–640. [Google Scholar] [CrossRef]
- Grillas, P.; Roché, J. Végétation des Marais Temporaires: Ecologie et Gestion: Conservation des Zones Humides Méditerranéennes, 8th ed.; MedWet, Tour du Valat: Arles, France, 1997; pp. 1–86. [Google Scholar]
- Bouahim, S.; Rhazi, L.; Amami, B.; Sahib, N.; Rhazi, M.; Waterkeyn, A.; Zouahri, A.; Mesleard, F.; Muller, S.D.; Grillas, P. Impact of grazing on the species richness of plant communities in Mediterranean temporary pools (western Morocco). Comptes Rendus Biol. 2010, 333, 670–679. [Google Scholar] [CrossRef]
- Hamel, T.; Boulemtafes, A.; Bellili, A.M. L’impact de surpâturage sur les subéraies de la péninsule de l’Edough (Nord-Est algérien). Geo-Eco-Trop 2019, 43, 119–128. [Google Scholar]
- Lefebvre, T.; Gallet, C. Impacts des grands herbivores sur la végétation des prairies et conséquences sur la décomposition de la litière. INRA Prod. Anim. 2018, 30, 455–464. [Google Scholar] [CrossRef]
- Touati, L. Potentiel de la Flore rare et Endémique dans le Nord-Est Algérien: Cas de la Wilaya de Souk Ahras. Utilisation de Champignons Mycorhiziens sur Opuntia maxima. Ph.D. Thesis, Badji-Mokhtar Annaba University, Annaba, Algeria, 2021. [Google Scholar]
- Benhouhou, S.; Yahi, N.; Véla, E. Algeria (chapter 3” Key Biodiversity Areas (KBAs) for plants in the Mediterranean region”). In Conserving Wild Plants in the South and East Mediterranean Region, 3rd ed.; IUCN & IUCN Centre for Mediterranean Cooperation: Gland, Switzerland, 2018; pp. 53–60. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Site | GPS Coordinates | Biogeographical Sector [13] | Altitude (m) | Surface Area (m2) |
|---|---|---|---|---|---|
| MTG | El-Matlegue | 36°23′34″ N; 7°53′42″ E | K3 | 947 | 76 |
| BTH | El-Batha | 36°25′24″ N; 8°14′39″ E | K3 | 835 | 3543 |
| TAR | Taoura | 36°09′13″ N; 7°59′28″ E | H2 | 855 | 6352 |
| BRG | Burgas Lake | 36°09′10″ N; 7°58′59″ E | H2 | 835 | 2162 |
| GZE | Guelta Zarga Effesus | 36°17′35″ N; 7°38′42″ E | C1 | 725 | 955 |
| GZN | Guelta Zarga Nigricans | 36°17′34″ N; 7°38′37″ E | C1 | 743 | 1500 |
| TRN | Taya Ranuncule | 36°30′55″ N; 7°05′37″ E | C1 | 940 | 386 |
| TRC | Taya Rockery | 36°30′32″ N; 7°05′32″ E | C1 | 1082 | 180 |
| MJN | Madjen Belahriti | 36°26′09″ N; 7°05′12″ E | C1 | 501 | 5155 |
| Soil Parameters | Abbreviation | Unit | Definition |
|---|---|---|---|
| Moisture | H | % | The amount of water in the soil that plants need for development, often correlated with air humidity. Lower soil moisture is usually associated with dry winds [38]. |
| Organic matter | OM | % | We can also name it organic carbon (OC). The mixture of animal and/or plant remains in various stages of decomposition defines soil organic matter (SOM), and its percentage of organic carbon directly affects its physicochemical qualities [39]. |
| Porosity | P | % | Depending on the soil’s moisture levels, air or water fills most of its spaces. Together, they represent porosity, a characteristic quantifying the volume of soil gaps in proportion to the total volume [40]. |
| Total limestone | TL | % | Limestone, primarily constituted of calcium carbonate crystallizing in the form of rhombohedral-symmetric calcite [41], provides the calcium that enables the soil to hold nutrient reserves [42]. |
| Electrical conductivity | EC | mS/cm | Determined by the concentration of ions and all soluble salts present [43]. |
| Hydrogen potential | pH | Affects the availability of most nutrients. It shows the soil’s acidity through the concentration of H+ ions [44]. |
| Water Parameters | Abbreviation | Unit | Definition |
|---|---|---|---|
| Hydrogen potential | pH | Affected by various natural factors, such as rock type, soil structure, and human activity [45]. | |
| Oxidation-reduction potential | ORP | Mv | Reflected in the redox potential of water, which establishes the ability of a chemical species to gain or lose electrons. It helps to improve the understanding of aquatic chemistry and to estimate the equilibrium behavior of chemical species [46]. |
| Dissolved oxygen | DO | mg/L | Measures the naturally dispersed oxygen in water. Its values should be above 4 mg/L to ensure a healthy aquatic environment and support the activity of microorganisms [47]. |
| Electrical conductivity | EC | µS/cm | Measures the ease with which water can transport an electric current, therefore; it is a crucial factor in stagnant water dynamics. It increases with the concentration of ionized minerals [48] |
| Resistivity | ρ | MΩ·cm | A measure in which electric charges in particles within a medium move under the influence of a potential difference, so is, sometimes, preferred to a conductivity value in low-conductivity measurements, such as ultrapure water [49]. |
| Total dissolved solids | TDS | mg/L | Naturally occurring pollutants that affect the color, total alkalinity and water conductivity in aquatic ecosystems which could be harmful with higher values [50]. |
| Salinity | S | PSU | Represents the weight and content of dissolved salts in the water and is the ideal indicator of freshwater and saltwater mixtures, all of them using a professional multiparameter device, the HANNA HI9829 |
| Temperature | T | °C | Affected by various natural factors, such as rock type, soil structure and human activity [45]. |
| Turbidity | T | NTU | According to Gaid (2022) [51], it is linked to the concentration of suspended solids. It is a characteristic that fluctuates depending on colloidal substances (clays, rock fragments, bacteria, etc.) or humic acids (plant degradation). Significant water turbidity reduces transparency, subsequently decreasing the amount of solar energy that can enter the water and be utilized for photosynthesis by aquatic life. It was measured using an AQUA LYTIC AL450T-IR turbidimeter. |
| Characteristic | Abbreviation | Scale and Ranks |
|---|---|---|
| Temporality and facies | Tmp and facs | st = temporary stagnation (1), sp = permanent stagnation (2), cp = permanent current (3), ct = temporary current (4). |
| Perimeter (m) | Per (m) | 1 = <100, 2 = 100–200, 3 = 201–300, 4 = 301–500, 5 = >500 |
| Surface area (m2) | Sur (m2) | 1 = <300, 2 = 300–700, 3 = 701–1500, 4 = 1501–2000, 5 = >2000 |
| Altitude (m) | Alti (m) | 1 = <300, 2 = 300–600, 3 = 601–800, 4 = 801–1000, 5 = >1000 |
| Slope | Slo | 1: none, 2: <15%, 3:15–30%, 4: >30% |
| Depth (cm) | Dep (cm) | 1 = < 25, 2 = 25–50, 3 = 51–75, 4 = 76–100, 5 = >100 |
| Exposure | Expo | 1 = North, 2 = South, 3 = West, 4 = North-east, 5 = North-west, 6 = South-east |
| Woody and herbaceous species cover rate | Wspr | 1 = <5%, 2 = 5–10%, 3 = 10–25%, 4 = 25–50%, 5 = 50–100% |
| Grazing, fire and agricultural activity | Gra, Fire and Agr | 1 = absent, 2 = minimal activity, 3 = slight activity, 4 = medium activity, 5 = very significant activity |
| Site | H (%) | OM (%) | P (%) | TC (%) | TL (%) | EC (mS/cm) | pH |
|---|---|---|---|---|---|---|---|
| MTG | 2.234 | 10.175 | 24.69 | 0.738 | 22.075 | 0.1 | 6.3 |
| BTH | 2.881 | 13.284 | 23.58 | 0.924 | 22.118 | 0.1 | 6.72 |
| TAR | 4.786 | 13.908 | 24.91 | 3.567 | 24.783 | 0.8 | 7.53 |
| BRG | 4.91 | 13.334 | 26.27 | 3.444 | 24.55 | 1.15 | 7.48 |
| GZE | 4.314 | 8.423 | 26.36 | 7.749 | 24.95 | 0.4 | 7.51 |
| GZN | 6.13 | 7.223 | 28.83 | 0.615 | 24.775 | 0.3 | 7.66 |
| TRN | 3.81 | 8.209 | 29.6 | 1.107 | 20.875 | 0.1 | 6.54 |
| TRC | 4.556 | 16.439 | 28.23 | 5.412 | 21.275 | 0.3 | 6.75 |
| MJN | 7.568 | 10.516 | 28.11 | 3.321 | 0 | 2.6 | 7.35 |
| Site | pH | ORP (mV) | DO (mg/L) | EC (µS/cm) | ρ (MΩ·cm) | TDS (mg/L) | S (PSU) | T (°C) | T (NTU) |
|---|---|---|---|---|---|---|---|---|---|
| MTG | 7.17 | 210.3 | 1.58 | 124 | 0.0081 | 62 | 0.06 | 20.01 | 11.6 |
| BTH | 7.2 | 248 | 1.44 | 147 | 0.0041 | 71 | 0.11 | 14.19 | 10.27 |
| TAR | 7.58 | 308.4 | 1.55 | 529.67 | 0.002 | 264.33 | 0.26 | 22.97 | 19.26 |
| BRG | 7.79 | 157.5 | 2.09 | 490.5 | 0.002 | 245 | 0.24 | 23.03 | 64.2 |
| GZE | 7.71 | 110.5 | 4.54 | 1325 | 0.0008 | 662 | 0.67 | 15.76 | 2.38 |
| GZN | 7.74 | 118.6 | 4.72 | 988 | 0.001 | 494 | 0.49 | 15.28 | 146 |
| TRN | 7.6 | 86.6 | 4.28 | 418 | 0.0024 | 209 | 0.2 | 14.86 | 5.07 |
| TRC | 7.43 | 108.6 | 4.77 | 215 | 0.0047 | 108 | 0.1 | 12.38 | 68.1 |
| MJN | 10.18 | 30.3 | 4.56 | 4197 | 0.0002 | 2099 | 2.25 | 17.81 | 5.31 |
| Site | Temp and Facs | Per | Sur | Alti | Slop | Dpth | Exp | Wspr | Graz | Agr | Fir |
|---|---|---|---|---|---|---|---|---|---|---|---|
| MTG | 1 | 1 | 1 | 4 | 2 | 3 | 1 | 5 | 3 | 1 | 3 |
| BTH | 2 | 3 | 5 | 4 | 1 | 1 | 4 | 4 | 5 | 2 | 2 |
| TAR | 3 | 3 | 5 | 4 | 2 | 3 | 2 | 5 | 5 | 1 | 2 |
| BRG | 2 | 4 | 5 | 4 | 2 | 4 | 2 | 5 | 5 | 2 | 2 |
| GZE | 1 | 2 | 3 | 3 | 3 | 1 | 6 | 4 | 3 | 1 | 2 |
| GZN | 1 | 2 | 3 | 3 | 1 | 3 | 6 | 4 | 4 | 2 | 2 |
| TRN | 1 | 1 | 2 | 4 | 4 | 3 | 5 | 3 | 4 | 1 | 2 |
| TRC | 3 | 1 | 1 | 5 | 4 | 3 | 5 | 4 | 3 | 4 | 2 |
| MJN | 2 | 3 | 5 | 2 | 2 | 5 | 3 | 4 | 5 | 4 | 2 |
| Chorological Types | MTG | BTH | TAR | BRG | GZE | GZN | TRN | TRC | MJN |
|---|---|---|---|---|---|---|---|---|---|
| Mediterranean | 96 | 46 | 69 | 60 | 88 | 43 | 23 | 79 | 34 |
| Widespread distribution | 10 | 7 | 16 | 16 | 13 | 8 | 9 | 11 | 10 |
| Holarctic | 19 | 11 | 19 | 13 | 17 | 7 | 7 | 15 | 9 |
| Introduced species | 3 | 3 | 2 | 0 | 1 | 0 | 0 | 0 | 1 |
| Endemic | 7 | 10 | 4 | 4 | 5 | 2 | 2 | 12 | 2 |
| Site | MTG | GZE | TRC | TAR | BRG | BTH | GZN | MJN | TRN |
|---|---|---|---|---|---|---|---|---|---|
| Shannon_H (S = ACE) | 38.65 | 35.82 | 34.53 | 32.2 | 27.78 | 23.59 | 19.09 | 18.02 | 13.96 |
| H (%) | OM (%) | P (%) | C (%) | CL (%) | CE (mS/cm) | pH | |
|---|---|---|---|---|---|---|---|
| Correl Shannon_H | −0.45794 | 0.40123 | −0.55393 | 0.49178 | 0.16373 | −0.27482 | −0.10743 |
| PH | ORP (mV) | OD (mg/L) | CE (µS/cm) | ρ (MΩ·m) | TDS (mg/L) | S (PSU) | T (°C) | T (NTU) | |
|---|---|---|---|---|---|---|---|---|---|
| Correl Shannon_H | −0.44011 | 0.43611 | −0.33878 | −0.37657 | 0.52755 | −0.37636 | −0.37926 | 0.25246 | −0.14978 |
| BRG | GZE | TRC | GZN | MTG | TAR | MJN | TRN | BTH | |
|---|---|---|---|---|---|---|---|---|---|
| Upper limit | 4.965 | 4.956 | 4.945 | 4.997 | 4.952 | 4.955 | 5.015 | 5.053 | 4.983 |
| Distinctness | 4.645 | 4.748 | 4.757 | 4.802 | 4.837 | 4.866 | 4.899 | 4.955 | 5.082 |
| Lower limit | 4.671 | 4.708 | 4.69 | 4.625 | 4.704 | 4.685 | 4.609 | 4.57 | 4.642 |
| Fir | Exp | Alti | Agra | Per | Temp & Facs | Sur | Wspr | Slop | Dpth | Graz | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Correl DT index | −0.5822 | 0.4278 | −0.4310 | −0.7474 | 0.0045 | 0.2315 | 0.0839 | −0.5108 | 0.3505 | 0.4927 | −0.1144 |
| p value | 0.0999 | 0.2506 | 0.2467 | 0.0206 | 0.9907 | 0.5489 | 0.83 | 0.1599 | 0.3551 | 0.1778 | 0.7694 |
| Florist | MTG | BTH | TAR | BRG | GZE | GZN | TRN | TRC | MJN |
|---|---|---|---|---|---|---|---|---|---|
| MTG | 1 | 0.38679245 | 0.48979592 | 0.35087719 | 0.49420849 | 0.35897436 | 0.30681818 | 0.43307087 | 0.28272251 |
| BTH | 0.38679245 | 1 | 0.29946524 | 0.16470588 | 0.20895522 | 0.18978102 | 0.18644068 | 0.20408163 | 0.15037594 |
| TAR | 0.48979592 | 0.29946524 | 1 | 0.44334975 | 0.47863248 | 0.41176471 | 0.33112583 | 0.4628821 | 0.37349398 |
| BRG | 0.35087719 | 0.16470588 | 0.44334975 | 1 | 0.50691244 | 0.35294118 | 0.37313433 | 0.46226415 | 0.51006711 |
| GZE | 0.49420849 | 0.20895522 | 0.47863248 | 0.50691244 | 1 | 0.41304348 | 0.36363636 | 0.41975309 | 0.41111111 |
| GZN | 0.35897436 | 0.18978102 | 0.41176471 | 0.35294118 | 0.41304348 | 1 | 0.41584158 | 0.40223464 | 0.37931034 |
| TRN | 0.30681818 | 0.18644068 | 0.33112583 | 0.37313433 | 0.36363636 | 0.41584158 | 1 | 0.4 | 0.43298969 |
| TRC | 0.43307087 | 0.20408163 | 0.4628821 | 0.46226415 | 0.41975309 | 0.40223464 | 0.4 | 1 | 0.4 |
| MJN | 0.28272251 | 0.15037594 | 0.37349398 | 0.51006711 | 0.41111111 | 0.37931034 | 0.43298969 | 0.4 | 1 |
| Ecolog | MTG | BTH | TAR | BRG | GZE | GZN | TRN | TRC | MJN |
| MTG | 1 | 0.87485649 | 0.88617289 | 0.85939661 | 0.84237389 | 0.82990935 | 0.86783652 | 0.8466044 | 0.74205644 |
| BTH | 0.87485649 | 1 | 0.91690337 | 0.90788013 | 0.85456294 | 0.85513673 | 0.86054515 | 0.848 | 0.79972772 |
| TAR | 0.88617289 | 0.91690337 | 1 | 0.95751396 | 0.87023449 | 0.86066447 | 0.86477987 | 0.85072167 | 0.8230553 |
| BRG | 0.85939661 | 0.90788013 | 0.95751396 | 1 | 0.86013635 | 0.8699316 | 0.85511029 | 0.85645933 | 0.84884628 |
| GZE | 0.84237389 | 0.85456294 | 0.87023449 | 0.86013635 | 1 | 0.91629956 | 0.9022948 | 0.87613543 | 0.81573479 |
| GZN | 0.82990935 | 0.85513673 | 0.86066447 | 0.8699316 | 0.91629956 | 1 | 0.90948068 | 0.87511478 | 0.82068933 |
| TRN | 0.86783652 | 0.86054515 | 0.86477987 | 0.85511029 | 0.9022948 | 0.90948068 | 1 | 0.91221595 | 0.78929205 |
| TRC | 0.8466044 | 0.848 | 0.85072167 | 0.85645933 | 0.87613543 | 0.87511478 | 0.91221595 | 1 | 0.78720497 |
| MJN | 0.74205644 | 0.79972772 | 0.8230553 | 0.84884628 | 0.81573479 | 0.82068933 | 0.78929205 | 0.78720497 | 1 |
| VSBF | SBF | WBF | VWBF | |
|---|---|---|---|---|
| VWBE | MTG-TAR | |||
| WBE | MTG-TRN/BRG-GZN | MTG-BTH/GZN-TRC/GZE TRC | TAR-GZE | |
| SBE | BTH-TRN/BTH-GZN/BTH-GZE | TAR-TRN/MTG-BRG/BRG-TRN | TAR-GZN | BRG-TRC/BRG-GZE |
| VSBE | BTH-TRC | TAR-TRC/BRG-MJN |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hammana, C.; Pereña-Ortiz, J.F.; Meddad-Hamza, A.; Hamel, T.; Salvo-Tierra, Á.E. The Wetlands of Northeastern Algeria (Guelma and Souk Ahras): Stakes for the Conservation of Regional Biodiversity. Land 2024, 13, 210. https://0-doi-org.brum.beds.ac.uk/10.3390/land13020210
Hammana C, Pereña-Ortiz JF, Meddad-Hamza A, Hamel T, Salvo-Tierra ÁE. The Wetlands of Northeastern Algeria (Guelma and Souk Ahras): Stakes for the Conservation of Regional Biodiversity. Land. 2024; 13(2):210. https://0-doi-org.brum.beds.ac.uk/10.3390/land13020210
Chicago/Turabian StyleHammana, Chayma, Jaime F. Pereña-Ortiz, Amel Meddad-Hamza, Tarek Hamel, and Ángel Enrique Salvo-Tierra. 2024. "The Wetlands of Northeastern Algeria (Guelma and Souk Ahras): Stakes for the Conservation of Regional Biodiversity" Land 13, no. 2: 210. https://0-doi-org.brum.beds.ac.uk/10.3390/land13020210