
Highly Specific Monoclonal Antibody Targeting the Botulinum Neurotoxin Type E Exposed SNAP-25 Neoepitope

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Peptides
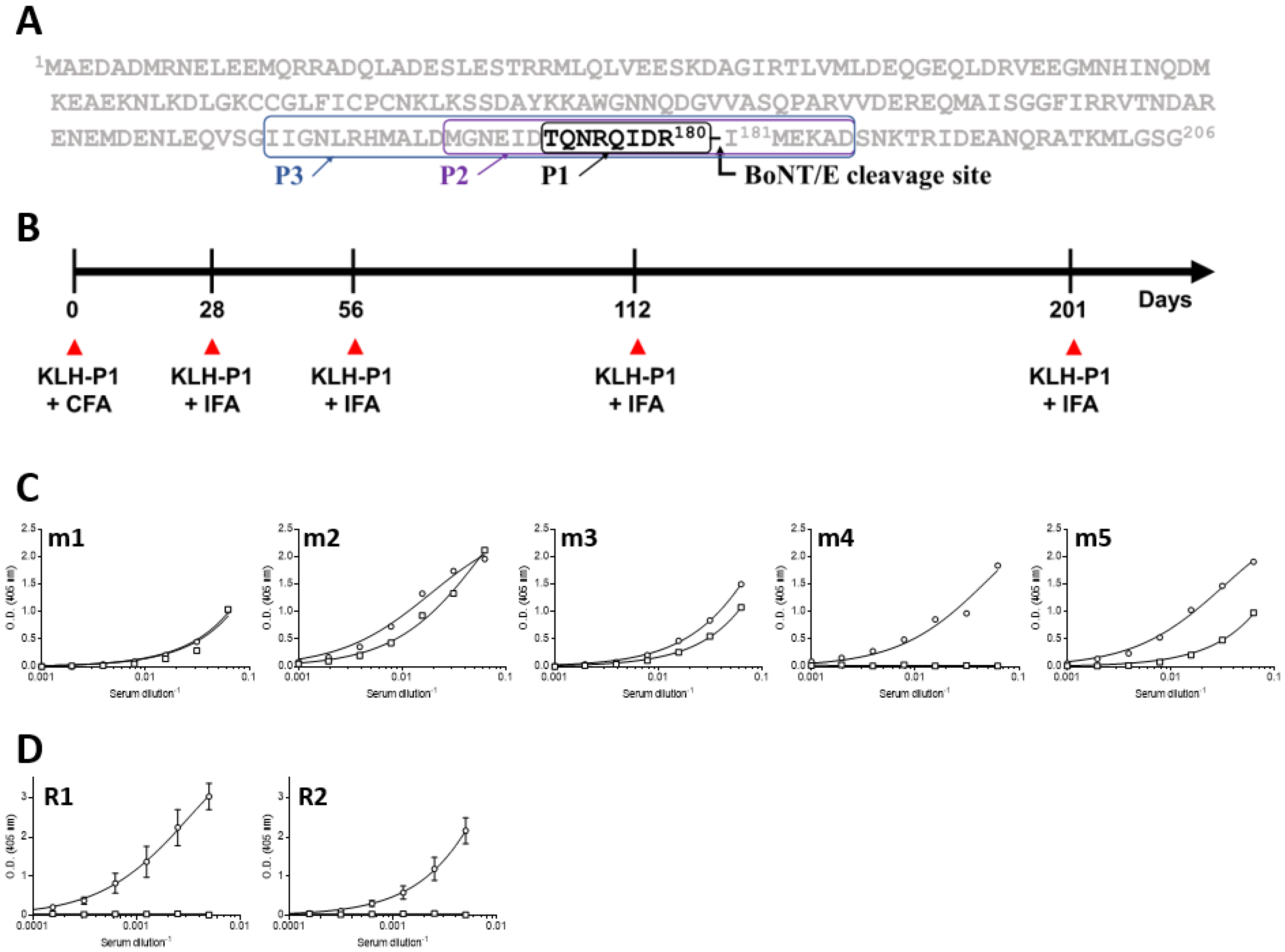
2.2. Animal Immunization
2.3. ScFv Library Construction and Screening
2.4. Production of Chimeric Antibodies
2.5. ELISA
2.6. Affinity and Specificity Measurements
2.7. Bacteria and Toxins
2.8. Exposure of SiMa cells to BoNT
2.9. Western-Blot
2.10. Antibody Variable Domain Structure Prediction
3. Results
3.1. Immunization and Characterization of the Elicited Antibody Response
3.2. Library Construction and Panning
3.3. Characterization of the Chimeric SNAP1 Antibody
3.4. Binding Characteristics of SNAP1 to the Cleaved SNAP-25 Product
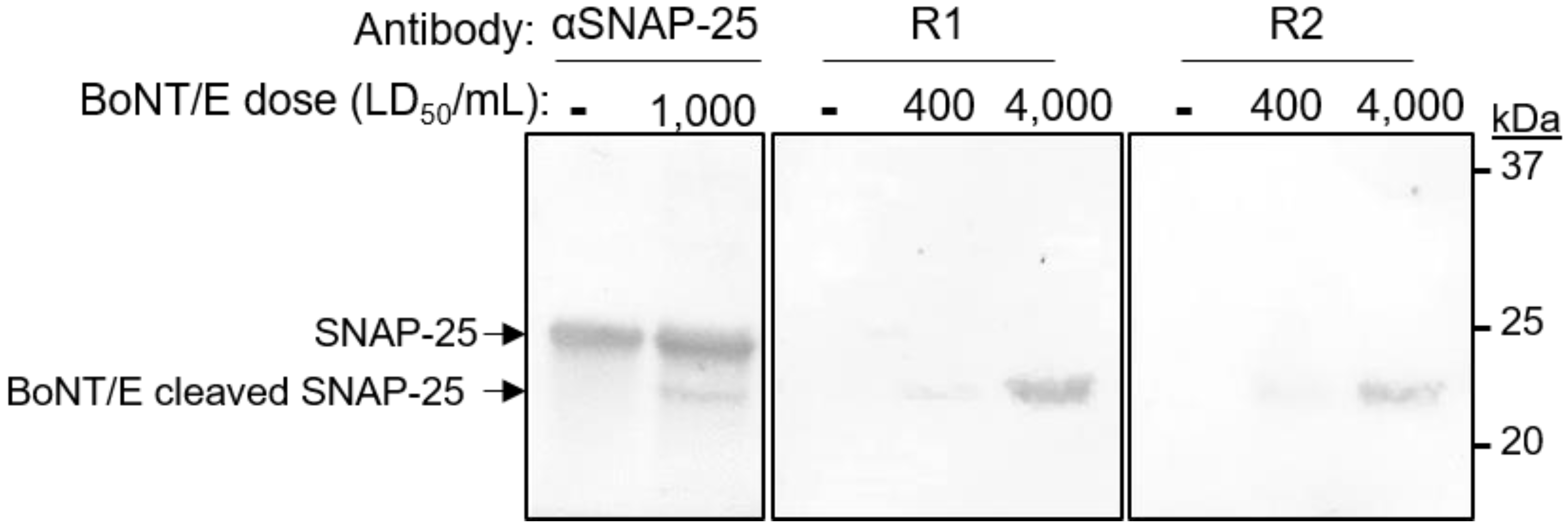
3.5. Selective Detection of BoNT/E Mediated Cleavage of SNAP-25
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Paddle, B.M. Therapy and prophylaxis of inhaled biological toxins. J. Appl. Toxicol. 2003, 23, 139–170. [Google Scholar] [CrossRef] [PubMed]
- Pirazzini, M.; Rossetto, O.; Eleopra, R.; Montecucco, C. Botulinum Neurotoxins: Biology, Pharmacology, and Toxicology. Pharmacol. Rev. 2017, 69, 200–235. [Google Scholar] [CrossRef] [PubMed]
- Lacy, D.B.; Stevens, R.C. Sequence homology and structural analysis of the clostridial neurotoxins. J. Mol. Biol. 1999, 291, 1091–1104. [Google Scholar] [CrossRef] [PubMed]
- Simpson, L.L. Identification of the major steps in botulinum toxin action. Annu. Rev. Pharmacol. Toxicol. 2004, 44, 167–193. [Google Scholar] [CrossRef] [PubMed]
- Schiavo, G.; Matteoli, M.; Montecucco, C. Neurotoxins affecting neuroexocytosis. Physiol. Rev. 2000, 80, 717–766. [Google Scholar] [CrossRef] [PubMed]
- Montal, M. Botulinum Neurotoxin: A Marvel of Protein Design. Annu. Rev. Biochem. 2010, 79, 591–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rusnak, J.M.; Smith, L.A. Botulinum neurotoxin vaccines: Past history and recent developments. Hum. Vaccines 2009, 5, 794–805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, L.A.; Rusnak, J.M. Botulinum Neurotoxin Vaccines: Past, Present, and Future. Crit. Rev. Immunol. 2007, 27, 303–318. [Google Scholar] [CrossRef] [PubMed]
- Binz, T.; Blasi, J.; Yamasaki, S.; Baumeister, A.; Link, E.; Südhof, T.C.; Jahn, R.; Niemann, H. Proteolysis of SNAP-25 by types E and A botulinal neurotoxins. J. Biol. Chem. 1994, 269, 1617–1620. [Google Scholar] [CrossRef]
- Schiavo, G.; Santucci, A.; Dasgupta, B.R.; Mehta, P.P.; Jontes, J.; Benfenati, F.; Wilson, M.C.; Montecucco, C. Botulinum neurotoxins serotypes A and E cleave SNAP-25 at distinct COOH-terminal peptide bonds. FEBS Lett. 1993, 335, 99–103. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Meng, J.; Lawrence, G.W.; Zurawski, T.H.; Sasse, A.; Bodeker, M.O.; Gilmore, M.A.; Fernández-Salas, E.; Francis, J.; Steward, L.E.; et al. Novel chimeras of botulinum neurotoxins A and E unveil contributions from the binding, translocation, and protease domains to their functional characteristics. J. Biol. Chem. 2008, 283, 16993–17002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dembek, Z.F.; Smith, L.A.; Rusnak, J.M. Botulism: Cause, effects, diagnosis, clinical and laboratory identification, and treatment modalities. Disaster Med. Public Health Prep. 2007, 1, 122–134. [Google Scholar] [CrossRef] [PubMed]
- MMWR. Investigational heptavalent botulinum antitoxin (HBAT) to replace licensed botulinum antitoxin AB and investigational botulinum antitoxin E. MMWR Morb. Mortal. Wkly. Rep. 2010, 59, 299. [Google Scholar]
- Infant Botulism Treatment and Prevention Program. Available online: http://www.infantbotulism.org (accessed on 3 October 2021).
- European Directorate for the Quality of Medicines and Healthcare. Botulinum Toxin Type A/Type B for Injection, 10th ed.; European Pharmacopoeia: Strasbourg, France, 2019. [Google Scholar]
- Torgeman, A.; Schwartz, A.; Diamant, E.; Baruchi, T.; Dor, E.; Ben David, A.; Pass, A.; Barnea, A.; Tal, A.; Rosner, A.; et al. Studying the differential efficacy of postsymptom antitoxin treatment in type A versus type B botulism using a rabbit spirometry model. Dis. Model. Mech 2018, 11, dmm035089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behrensdorf-Nicol, H.A.; Wild, E.; Bonifas, U.; Klimek, J.; Hanschmann, K.M.; Krämer, B.; Kegel, B. In vitro potency determination of botulinum neurotoxin serotype A based on its receptor-binding and proteolytic characteristics. Toxicol. Int. J. Publ. Assoc. BIBRA 2018, 53, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Dressler, D.; Dirnberger, G. Botulinum toxin antibody testing: Comparison between the immunoprecipitation assay and the mouse diaphragm assay. Eur. Neurol. 2001, 45, 257–260. [Google Scholar] [CrossRef] [PubMed]
- Hanna, P.A.; Jankovic, J. Mouse bioassay versus Western blot assay for botulinum toxin antibodies: Correlation with clinical response. Neurology 1998, 50, 1624–1629. [Google Scholar] [CrossRef] [PubMed]
- Lindsey, C.Y.; Smith, L.A.; West, M.W.; Boles, J.W.; Brown, J.E. Evaluation of a botulinum fragment C-based ELISA for measuring the humoral immune response in primates. Biologicals 2003, 31, 17–24. [Google Scholar] [CrossRef]
- Palace, J.; Nairne, A.; Hyman, N.; Doherty, T.V.; Vincent, A. A radioimmuno-precipitation assay for antibodies to botulinum A. Neurology 1998, 50, 1463–1466. [Google Scholar] [CrossRef]
- Rosen, O.; Ozeri, E.; Barnea, A.; David, A.B.; Zichel, R. Development of an Innovative in Vitro Potency Assay for Anti-Botulinum Antitoxins. Toxins 2016, 8, 276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wild, E.; Bonifas, U.; Klimek, J.; Trösemeier, J.H.; Krämer, B.; Kegel, B.; Behrensdorf-Nicol, H.A. In vitro potency determination of botulinum neurotoxin B based on its receptor-binding and proteolytic characteristics. Toxicol. Int. J. Publ. Assoc. BIBRA 2016, 34, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Salas, E.; Wang, J.; Molina, Y.; Nelson, J.B.; Jacky, B.P.; Aoki, K.R. Botulinum neurotoxin serotype A specific cell-based potency assay to replace the mouse bioassay. PLoS ONE 2012, 7, e49516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torgeman, A.; Diamant, E.; Levin, L.; Ben David, A.; Epstein, E.; Girshengorn, M.; Mazor, O.; Rosenfeld, R.; Zichel, R. An in vitro cell-based potency assay for pharmaceutical type A botulinum antitoxins. Vaccine 2017, 35, 7213–7216. [Google Scholar] [CrossRef] [PubMed]
- Bak, N.; Rajagopal, S.; Stickings, P.; Sesardic, D. SiMa Cells for a Serotype Specific and Sensitive Cell-Based Neutralization Test for Botulinum Toxin A and E. Toxins 2017, 9, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leveque, C.; Ferracci, G. An optical biosensor assay for rapid dual detection of Botulinum neurotoxins A and E. Sci. Rep. 2015, 5, 17953. [Google Scholar] [CrossRef] [PubMed]
- von Berg, L.; Stern, D.; Pauly, D.; Mahrhold, S.; Weisemann, J.; Jentsch, L.; Hansbauer, E.M.; Muller, C.; Avondet, M.A.; Rummel, A.; et al. Functional detection of botulinum neurotoxin serotypes A to F by monoclonal neoepitope-specific antibodies and suspension array technology. Sci. Rep. 2019, 9, 5531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mechaly, A.; Alcalay, R.; Noy-Porat, T.; Epstein, E.; Gal, Y.; Mazor, O. Novel Phage Display-Derived Anti-Abrin Antibodies Confer Post-Exposure Protection against Abrin Intoxication. Toxins 2018, 10, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mechaly, A.; Elia, U.; Alcalay, R.; Cohen, H.; Epstein, E.; Cohen, O.; Mazor, O. Inhibition of Francisella tularensis phagocytosis using a novel anti-LPS scFv antibody fragment. Sci. Rep. 2019, 9, 11418. [Google Scholar] [CrossRef] [Green Version]
- Noy-Porat, T.; Rosenfeld, R.; Ariel, N.; Epstein, E.; Alcalay, R.; Zvi, A.; Kronman, C.; Ordentlich, A.; Mazor, O. Isolation of Anti-Ricin Protective Antibodies Exhibiting High Affinity from Immunized Non-Human Primates. Toxins 2016, 8, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azriel-Rosenfeld, R.; Valensi, M.; Benhar, I. A human synthetic combinatorial library of arrayable single-chain antibodies based on shuffling in vivo formed CDRs into general framework regions. J. Mol. Biol. 2004, 335, 177–192. [Google Scholar] [CrossRef]
- Rosenfeld, R.; Marcus, H.; Ben-Arie, E.; Lachmi, B.; Mechaly, A.; Reuveny, S.; Gat, O.; Mazor, O.; Ordentlich, A. Isolation and Chimerization of a Highly Neutralizing Antibody conferring Passive Protection against Lethal B. anthracis Infection. PLoS ONE 2009, 4, e6351. [Google Scholar] [CrossRef] [PubMed]
- Bujak, E.; Matasci, M. (Eds.) Reformatting of scFv Antibodies into the scFv-Fc Format and Their Downstream Purification; Springer: New York, NY, USA, 2014; Volume 1131. [Google Scholar]
- Noy-Porat, T.; Alcalay, R.; Mechaly, A.; Peretz, E.; Makdasi, E.; Rosenfeld, R.; Mazor, O. Characterization of antibody-antigen interactions using biolayer interferometry. STAR Protoc. 2021, 2, 100836. [Google Scholar] [CrossRef] [PubMed]
- Whelan, S.M.; Elmore, M.J.; Bodsworth, N.J.; Atkinson, T.; Minton, N.P. The complete amino acid sequence of the Clostridium botulinum type-E neurotoxin, derived by nucleotide-sequence analysis of the encoding gene. Eur. J. Biochem. 1992, 204, 657–667. [Google Scholar] [CrossRef] [PubMed]
- Binz, T.; Kurazono, H.; Wille, M.; Frevert, J.; Wernars, K.; Niemann, H. The complete sequence of botulinum neurotoxin type A and comparison with other clostridial neurotoxins. J. Biol. Chem. 1990, 265, 9153–9158. [Google Scholar] [CrossRef]
- Diamant, E.; Torgeman, A.; Epstein, E.; Mechaly, A.; David, A.B.; Levin, L.; Schwartz, A.; Dor, E.; Girshengorn, M.; Barnea, A.; et al. A cell-based alternative to the mouse potency assay for pharmaceutical type E botulinum antitoxins. ALTEX 2022, 39, 113–122. [Google Scholar] [CrossRef]
- Mirdita, M.; Schütze, K.; Moriwaki, Y.; Heo, L.; Ovchinnikov, S.; Steinegger, M. ColabFold—Making protein folding accessible to all. bioRxiv 2021. bioRxiv:2021.2008.2015.456425. [Google Scholar]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zidek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Evans, R.; O’Neill, M.; Pritzel, A.; Antropova, N.; Senior, A.; Green, T.; Žídek, A.; Bates, R.; Blackwell, S.; Yim, J.; et al. Protein complex prediction with AlphaFold-Multimer. bioRxiv 2021. bioRxiv:2021.2010.2004.463034. [Google Scholar]
- Wrammert, J.; Smith, K.; Miller, J.; Langley, W.A.; Kokko, K.; Larsen, C.; Zheng, N.; Mays, I.; Garman, L.; Helms, C.; et al. Rapid cloning of high-affinity human monoclonal antibodies against influenza virus. Nature 2008, 453, 667–668. [Google Scholar] [CrossRef] [Green Version]
- Greenfield, E.A. Standard Immunization of Rabbits. Cold Spring Harb. Protoc. 2020, 2020, 100305. [Google Scholar] [CrossRef] [PubMed]
- Overduin, L.A.; van Dongen, J.J.M.; Visser, L.G. The Cellular Immune Response to Rabies Vaccination: A Systematic Review. Vaccines 2019, 7, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webster, R.G. The immune response to influenza virus. II. Effect of the route and schedule of vaccination on the quantity and avidity of antibodies. Immunology 1968, 14, 29–37. [Google Scholar] [PubMed]
- Kodangattil, S.; Huard, C.; Ross, C.; Li, J.; Gao, H.; Mascioni, A.; Hodawadekar, S.; Naik, S.; Min-debartolo, J.; Visintin, A.; et al. The functional repertoire of rabbit antibodies and antibody discovery via next-generation sequencing. mAbs 2014, 6, 628–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavinder, J.J.; Hoi, K.H.; Reddy, S.T.; Wine, Y.; Georgiou, G. Systematic characterization and comparative analysis of the rabbit immunoglobulin repertoire. PLoS ONE 2014, 9, e101322. [Google Scholar] [CrossRef] [PubMed]
- Rief, N.; Waschow, C.; Nastainczyk, W.; Montenarh, M.; Gotz, C. Production and characterization of a rabbit monoclonal antibody against human CDC25C phosphatase. Hybridoma 1998, 17, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Wang, X.; Jin, H. Rabbit monoclonal antibody: Potential application in cancer therapy. Am. J. Transl. Res. 2011, 3, 269–274. [Google Scholar] [PubMed]
- Li, Y.; Cockburn, W.; Kilpatrick, J.B.; Whitelam, G.C. High affinity ScFvs from a single rabbit immunized with multiple haptens. Biochem. Biophys. Res. Commun. 2000, 268, 398–404. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Han, Z.; Lu, L.; Wang, L.; Ni, G.; Zhao, Z.; Wu, A.; Zheng, X. Development of a new rabbit monoclonal antibody and its based competitive indirect enzyme-linked immunosorbent assay for rapid detection of sulfonamides. J. Sci. Food Agric. 2013, 93, 667–673. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Zhao, Z.; Tan, Y.; Lu, L.; Wang, L.; Liao, Y.; Beloglazova, N.; De Saeger, S.; Zheng, X.; Wu, A. Simultaneous Raising of Rabbit Monoclonal Antibodies to Fluoroquinolones with Diverse Recognition Functionalities via Single Mixture Immunization. Anal. Chem. 2016, 88, 1246–1252. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Kriegel, A.M.; Boustany, C.A.; Blake, D.A. Single-chain variable fragment (scFv) antibodies optimized for environmental analysis of uranium. Anal. Chem. 2011, 83, 3717–3724. [Google Scholar] [CrossRef]
- Miller, W.; Rosenbloom, K.; Hardison, R.C.; Hou, M.; Taylor, J.; Raney, B.; Burhans, R.; King, D.C.; Baertsch, R.; Blankenberg, D.; et al. 28-way vertebrate alignment and conservation track in the UCSC Genome Browser. Genome Res. 2007, 17, 1797–1808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosner, K.; Winter, D.B.; Tarone, R.E.; Skovgaard, G.L.; Bohr, V.A.; Gearhart, P.J. Third complementarity-determining region of mutated VH immunoglobulin genes contains shorter V, D, J, P, and N components than non-mutated genes. Immunology 2001, 103, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Fernandes-Salas, E.; Wang, J.; Kei, R. Immuno-Based Botulinum Toxin Serotype A Activity Assays. U.S. Patent No. US 12/403,531, 6 September 2012. [Google Scholar]
- Chen, X.; Tomchick, D.R.; Kovrigin, E.; Arac, D.; Machius, M.; Sudhof, T.C.; Rizo, J. Three-dimensional structure of the complexin/SNARE complex. Neuron 2002, 33, 397–409. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Heavy Chain | Light Chain | ||||||
|---|---|---|---|---|---|---|---|
| Ab | Source | CDR1 1 | CDR2 | CDR3 | CDR1 | CDR2 | CDR3 |
| SNAP1 | Rabbit 2 | GIDLSDSA | IYIGSGSA | ARAWDI | ENIYNN | RAS | QSSYVGTHVNYGND |
| SNAP2 | Rabbit 1 | GINLSSSA | IYAGSGNT | ATAWDI | QSIGSN | RAS | QSSYVGTHINYGNG |
| SNAP3 | Rabbits 1 + 2 | GIDFSDNA | TYVGSGRT | ARTWDI | ENIYNN | RAS | QSSYVGTTINYGNG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mechaly, A.; Diamant, E.; Alcalay, R.; Ben David, A.; Dor, E.; Torgeman, A.; Barnea, A.; Girshengorn, M.; Levin, L.; Epstein, E.; et al. Highly Specific Monoclonal Antibody Targeting the Botulinum Neurotoxin Type E Exposed SNAP-25 Neoepitope. Antibodies 2022, 11, 21. https://0-doi-org.brum.beds.ac.uk/10.3390/antib11010021
Mechaly A, Diamant E, Alcalay R, Ben David A, Dor E, Torgeman A, Barnea A, Girshengorn M, Levin L, Epstein E, et al. Highly Specific Monoclonal Antibody Targeting the Botulinum Neurotoxin Type E Exposed SNAP-25 Neoepitope. Antibodies. 2022; 11(1):21. https://0-doi-org.brum.beds.ac.uk/10.3390/antib11010021
Chicago/Turabian StyleMechaly, Adva, Eran Diamant, Ron Alcalay, Alon Ben David, Eyal Dor, Amram Torgeman, Ada Barnea, Meni Girshengorn, Lilach Levin, Eyal Epstein, and et al. 2022. "Highly Specific Monoclonal Antibody Targeting the Botulinum Neurotoxin Type E Exposed SNAP-25 Neoepitope" Antibodies 11, no. 1: 21. https://0-doi-org.brum.beds.ac.uk/10.3390/antib11010021