Using Behavioral Instability to Investigate Behavioral Reaction Norms in Captive Animals: Theoretical Implications and Future Perspectives

 ,
,
Abstract
:1. Introduction
Aim of the Paper
2. Methods
2.1. Animals and Setting
2.2. Data Collection
2.3. Analysis
2.4. Proportion of Time Each Individual Spent on Each Behavior
2.5. Reaction Norms for Testing Differences between Individuals and Between Treatments
3. Results
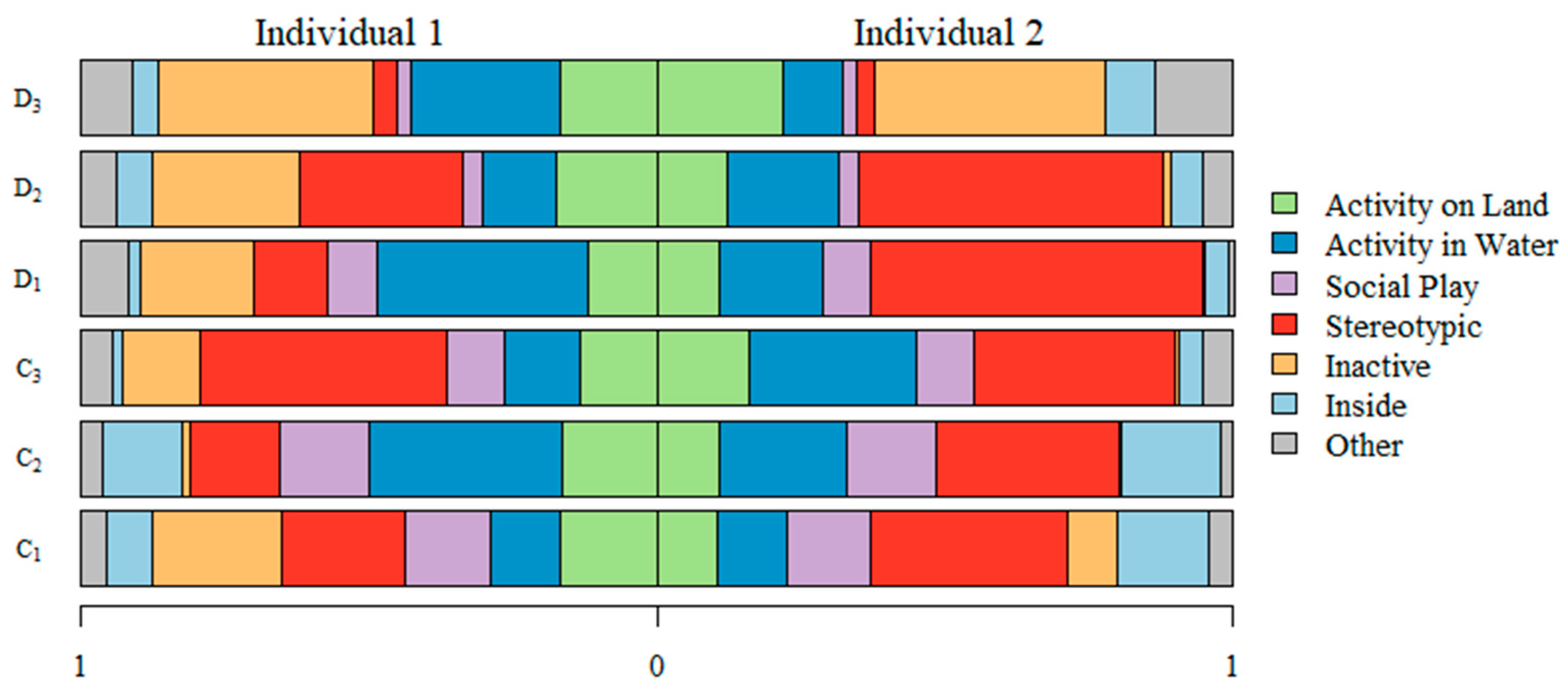
3.1. Proportion of Time Each Individual Spent on Each Behavior
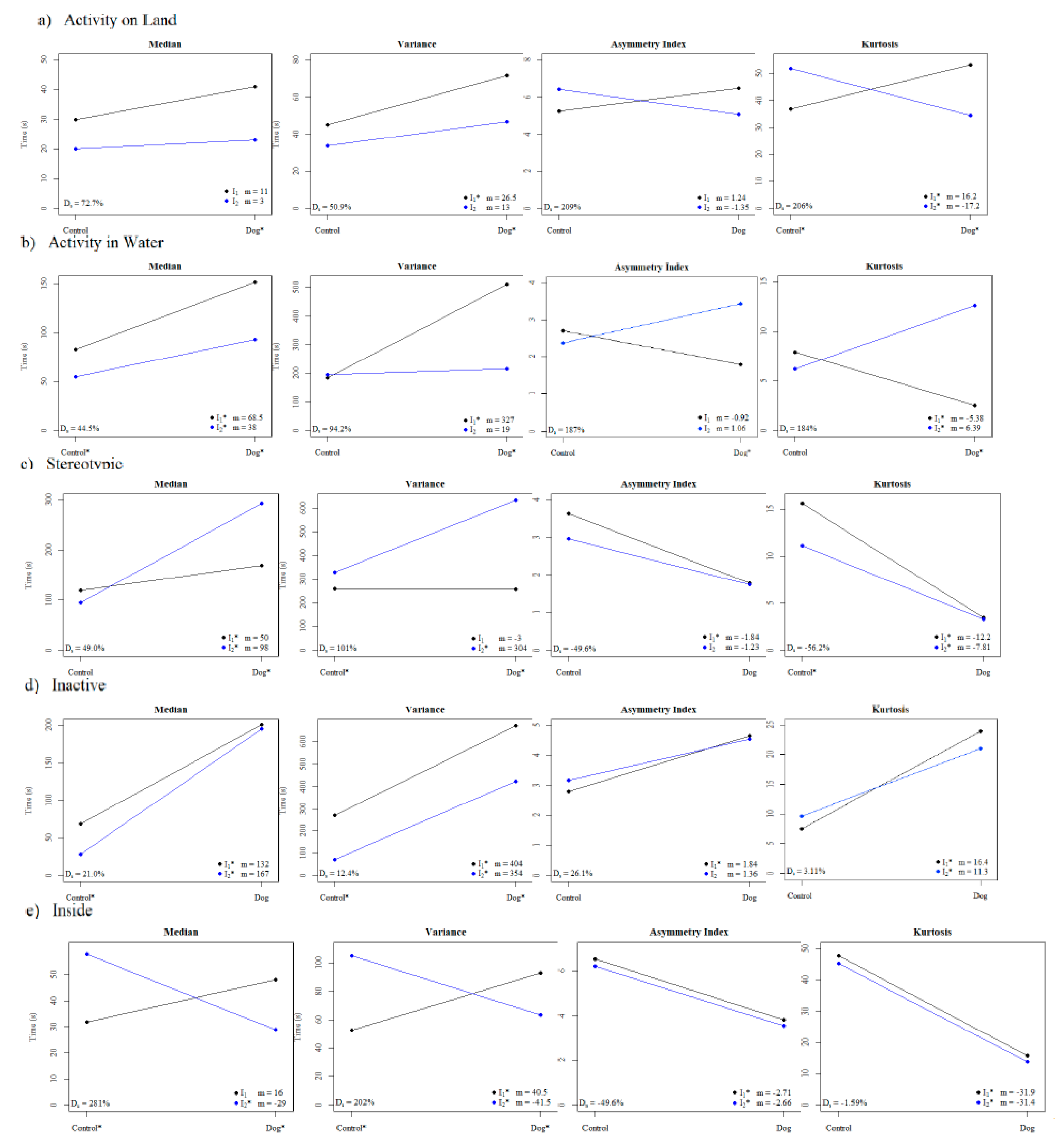
3.2. Reaction Norms for Testing Differences between Individuals and Treatments
4. Discussion
4.1. Results of the Case Study
4.2. Reliability of Results
4.3. Considerations when Removing Outliers
4.4. Applying Behavioral Instability to Behavioral Investigations
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

Appendix B
χ2 Test of Total Time
| Behavior | χ2 Test Comparing Individuals | χ2 Test Comparing Treatments | ||
| Activity on land | χ2 = 0.3542 | χ2 = 6.730 × 10−3 | χ2 = 1.388 × 10−4 | χ2 = 0.2510 |
| p = 0.5518 | p = 0.9346 | p = 0.9906 | p = 0.6164 | |
| Activity in water | χ2 = 0.05417 | χ2 = 2.058 | χ2 = 0.6791 | χ2 = 0.7223 |
| p = 0.8160 | p = 0.1514 | p = 0.4099 | p = 0.3954 | |
| Social play | χ2 = 2.666 × 10−4 | |||
| p = 0.9870 | ||||
| Stereotypic | χ2 = 0.8345 | χ2 = 9.695 | χ2 = 3.104 | χ2 = 0.2462 |
| p = 0.3610 | p = 0.001848 | p = 0.07808 | p = 0.6197 | |
| Inactive | χ2 = 4.501 | χ2 = 5.736 | ||
| p = 0.03388 | p = 0.01661 | |||
| Inside | χ2 = 1.118 | χ2 = 2.169 | ||
| p = 0.2904 | p = 0.1408 | |||
Appendix C
Appendix C.1. Slopes of Medians, Variances, Asymmetry Indices and Kurtoses
Appendix C.2. Dataset where Outliers were Removed outside IQR





Appendix C.3. Dataset where Outliers were Removed Using the MAD Method





Appendix D
χ2 Test for Distribution of Statistics
| Behavior | χ2 Test Comparing Individuals | χ2 Test Comparing Treatments | ||
| Activity on land | χ2 = 2.000 | χ2 = 5.063 | χ2 = 1.704 | χ2 = 0.2093 |
| p = 0.1573 | p = 0.02445 | p = 0.1917 | p = 0.6473 | |
| Activity in water | χ2 = 5.681 | χ2 = 14.00 | χ2 = 20.01 | χ2 = 9.757 |
| p = 0.01715 | p = 0.0001831 | p = 7.705 × 10−6 | p = 0.001787 | |
| Stereotypic | χ2 = 2.692 | χ2 = 33.28 | χ2 = 8.681 | χ2 = 101.0 |
| p = 0.1009 | p = 7.974 × 10−9 | p = 0.003216 | p < 2.2 × 10−16 | |
| Inactive | χ2 = 17.33 | χ2 = 0.090909 | χ2 = 64.53 | χ2 = 125.1 |
| p = 3.142 × 10−5 | p = 0.7630 | p = 9.491 × 10−16 | p < 2.2 × 10−16 | |
| Inside | χ2 = 7.511 | χ2 = 4.6883 | χ2 = 3.200 | χ2 = 9.667 |
| p = 0.006130 | p = 0.03037 | p = 0.07364 | p = 0.001876 | |
| Behavior | χ2 Test Comparing Individuals | χ2 Test Comparing Treatments | ||
| Cpooled | Dpooled | |||
| Activity on land | χ2 = 1.607 | χ2 = 5.1802 | χ2 = 6.0279 | χ2 = 2.0994 |
| p = 0.2049 | p = 0.02285 | p = 0.01408 | p = 0.1474 | |
| Activity in water | χ2 = 0.3485 | χ2 = 120.68 | χ2 = 153.85 | χ2 = 0.92631 |
| p = 0.5550 | p < 2.2 × 10−16 | p < 2.2 × 10−16 | p = 0.3358 | |
| Stereotypic | χ2 = 8.000 | χ2 = 160.06 | χ2 = 0.02381 | χ2 = 96.901 |
| p = 0.004677 | p < 2.2 × 10−16 | p = 0.8774 | p < 2.2 × 10−16 | |
| Inactive | χ2 = 117.65 | χ2 = 56.668 | χ2 = 172.9 | χ2 = 254.14 |
| p < 2.2 × 10−16 | p = 5.15910−14 | p < 2.2 × 10−16 | p < 2.2 × 10−16 | |
| Inside | χ2 = 17.5 | χ2 = 5.5607 | χ2 = 11.273 | χ2 = 10.221 |
| p = 2.873 × 10−5 | p = 0.01837 | p = 0.0007863 | p = 0.001388 | |
| Behavior | χ2 Test Comparing Individuals | χ2 Test Comparing Treatments | ||
| Cpooled | Dpooled | |||
| Activity on land | χ2 = 1.201 | χ2 = 1.713 | χ2 = 1.311 | χ2 = 1.588 |
| p = 0.2731 | p = 0.1905 | p = 0.2523 | p = 0.2076 | |
| Activity in water | χ2 = 0.2064 | χ2 = 5.254 | χ2 = 1.886 | χ2 = 1.934 |
| p = 0.6496 | p = 0.0219 | p = 0.1697 | p = 0.1644 | |
| Stereotypic | χ2 = 0.6759 | χ2 = 0.007748 | χ2 = 6.280 | χ2 = 3.216 |
| p = 0.411 | p = 0.9299 | p = 0.01221 | p = 0.07293 | |
| Inactive | χ2 = 0.2501 | χ2 = 0.01137 | χ2 = 4.58 | χ2 = 2.384 |
| p = 0.617 | p = 0.9151 | p = 0.03235 | p = 0.1226 | |
| Inside | χ2 = 0.07902 | χ2 = 0.09389 | χ2 = 7.104 | χ2 = 7.228 |
| p = 0.7786 | p = 0.7593 | p = 0.007693 | p = 0.007178 | |
| Behavior | χ2 Test Comparing Individuals | χ2 Test Comparing Treatments | ||
| Cpooled | Dpooled | |||
| Activity on land | χ2 = 25.15 | χ2 = 39.02 | χ2 = 29.34 | χ2 = 34.19 |
| p = 5.305 × 10−7 | p = 4.187 × 10−10 | p = 6.09 × 10−8 | p = 4.996 × 10−9 | |
| Activity in water | χ2 = 2.062 | χ2 = 66.84 | χ2 = 27.65 | χ2 = 21.70 |
| p = 0.1511 | p = 2.945e-16 | p = 1.452 × 10−7 | p = 3.196 × 10−6 | |
| Stereotypic | χ2 = 7.574 | χ2 = 0.02510 | χ2 = 77.88 | χ2 = 42.31 |
| p = 0.005922 | p = 0.8741 | p < 2.2 × 10−16 | p = 7.788 × 10−11 | |
| Inactive | χ2 = 2.824 | χ2 = 1.915 | χ2 = 86.12 | χ2 = 41.71 |
| p = 0.09284 | p = 0.1664 | p < 2.2 × 10−16 | p = 1.059 × 10−10 | |
| Inside | χ2 = 0.6862 | χ2 = 1.432 | χ2 = 159.7 | χ2 = 167.1 |
| p = 0.4075 | p = 0.2315 | p < 2.2 × 10−16 | p < 2.2 × 10−16 | |
| Behavior | χ2 Test Comparing Individuals | χ2 Test Comparing Treatments | ||
| Activity on land | χ2 = 1.581 | χ2 = 5.400 | χ2 = 5.924 | χ2 = 1.882 |
| p = 0.2086 | p = 0.02014 | p = 0.01486 | p = 0.1701 | |
| Activity in water | χ2 = 0.5436 | χ2 = 22.42 | χ2 = 41.12 | χ2 = 6.208 |
| p = 0.4609 | p = 2.194 × 10−6 | p = 1.429 × 10−10 | p = 0.0127 | |
| Stereotypic | χ2 = 0.6429 | χ2 = 130.9 | χ2 = 1.766 | χ2 = 141.3 |
| p = 0.4227 | p < 2.2 × 10−16 | p = 0.1839 | p < 2.2 × 10−16 | |
| Inactive | χ2 = 36.92 | χ2 = 1.313 | χ2 = 48.00 | χ2 = 124.5 |
| p = 1.234 × 10−9 | p = 0.2519 | p = 4.255 × 10−12 | p < 2.2 × 10−16 | |
| Inside | χ2 = 11.77 | χ2 = 15.99 | χ2 = 20.56 | χ2 = 8.25 |
| p = 0.0006032 | p = 6.351 × 10−5 | p = 5.771 × 10−6 | p = 0.004075 | |
| Behavior | χ2 Test Comparing Individuals | χ2 Test Comparing Treatments | ||
| Activity on land | χ2 = 0.9885 | χ2 = 1.231 | χ2 = 0.1648 | χ2 = 0.0821 |
| p = 0.3201 | p = 0.2673 | p = 0.6848 | p = 0.7745 | |
| Activity in water | χ2 = 0.07778 | χ2 = 1.211 | χ2 = 0.1620 | χ2 = 0.9592 |
| p = 0.7803 | p = 0.2711 | p = 0.6873 | p = 0.3274 | |
| Stereotypic | χ2 = 0.518 | χ2 = 1.044 | χ2 = 0.3828 | χ2 = 5.069 |
| p = 0.4715 | p = 0.3069 | p = 0.5361 | p = 0.02436 | |
| Inactive | χ2 = 0.0781 | χ2 = 2.675 | χ2 = 0.2099 | χ2 = 0.8363 |
| p = 0.7799 | p = 0.1019 | p = 0.6468 | p = 0.3605 | |
| Inside | χ2 = 4.791 | χ2 = 0.04612 | χ2 = 2.024 | χ2 = 0.3942 |
| p = 0.02861 | p = 0.8300 | p = 0.1548 | p = 0.5300 | |
| Behavior | χ2 Test Comparing Individuals | χ2 Test Comparing Treatments | ||
| Activity on land | - | - | - | - |
| Activity in water | - | - | - | - |
| Stereotypic | - | - | - | - |
| Inactive | χ2 = 4.858 | χ2 = 34.39 | χ2 = 3.096 | χ2 = 8.407 |
| p = 0.02752 | p = 4.518 × 10−9 | p = 0.07851 | p = 0.003738 | |
| Inside | - | - | - | - |
| Behavior | χ2 Test Comparing Individuals | χ2 Test Comparing Treatments | ||
| Activity on land | χ2 = 2.814 | χ2 = 5.333 | χ2 = 0.4237 | χ2 = 0 |
| p = 0.09345 | p = 0.02092 | p = 0.5151 | p = 1 | |
| Activity in water | χ2 = 2.919 | χ2 = 1.651 | χ2 = 3.6552 | χ2 = 5.434 |
| p = 0.08753 | p = 0.1988 | p = 0.05589 | p = 0.01975 | |
| Stereotypic | χ2 = 11.21 | χ2 = 46.41 | χ2 = 3.435 | χ2 = 131.1 |
| p = 0.0008153 | p = 9.609 × 10−12 | p = 0.06385 | p < 2.2 × 10−16 | |
| Inactive | χ2 = 9.1351 | χ2 = 15.125 | χ2 = 23.11 | χ2 = 116.5 |
| p = 0.0002507 | p = 0.0001469 | p = 1.528 × 10−6 | p < 2.2 × 10−16 | |
| Inside | χ2 = 2.522 | χ2 = 0.8649 | χ2 = 2.882 | χ2 = 0.6712 |
| p = 0.1122 | p = 0.3524 | p = 0.08956 | p = 0.4126 | |
| Behavior | χ2 Test Comparing Individuals | χ2 Test Comparing Treatments | ||
| Activity on land | χ2 = 4.373 | χ2 = 9.727 | χ2 = 4.580 | χ2 = 1.142 |
| p = 0.03873 | p = 0.001816 | p = 0.03234 | p = 0.2853 | |
| Activity in water | χ2 = 6.0945 | χ2 = 11.48 | χ2 = 19.16 | χ2 = 11.99 |
| p = 0.01356 | p = 0.000704 | p = 1.203 × 10−5 | p = 0.000536 | |
| Stereotypic | χ2 = 3.647 | χ2 = 152.4 | χ2 = 4.179 | χ2 = 247.5 |
| p = 0.05616 | p < 2.2 × 10−16 | p = 0.04093 | p < 2.2 × 10−16 | |
| Inactive | χ2 = 22.38 | χ2 = 59.73 | χ2 = 51.22 | χ2 = 298.2 |
| p = 2.36 × 10−6 | p = 1.091 × 10−14 | p = 8.25 × 10−13 | p < 2.2 × 10−16 | |
| Inside | χ2 = 21.59 | χ2 = 9.940 | χ2 = 24.30 | χ2 = 8.127 |
| p = 3.383 × 10−6 | p = 0.001618 | p = 8.229 × 10−7 | p = 0.00436 | |
| Behavior | χ2 Test Comparing Individuals | χ2 Test Comparing Treatments | ||
| Activity on land | χ2 = 0.1273 | χ2 = 0.01984 | χ2 = 0.1 | χ2 = 0.01004 |
| p = 0.7212 | p = 0.888 | p = 0.7518 | p = 0.9202 | |
| Activity in water | χ2 = 0.03937 | χ2 = 0.3240 | χ2 = 0.003371 | χ2 = 0.09825 |
| p = 0.8427 | p = 0.5692 | p = 0.9537 | p = 0.7539 | |
| Stereotypic | χ2 = 0.09412 | χ2 = 0.02645 | χ2 = 0.003557 | χ2 = 0.2793 |
| p = 0.759 | p = 0.8708 | p = 0.9524 | p = 0.5972 | |
| Inactive | χ2 = 0.4181 | χ2 = 1.288 | χ2 = 0.2054 | χ2 = 0.001563 |
| p = 0.5179 | p = 0.2564 | p = 0.6504 | p = 0.9685 | |
| Inside | χ2 = 0.0004049 | χ2 = 0.2091 | χ2 = 0.03389 | χ2 = 0.08620 |
| p = 0.9839 | p = 0.6475 | p = 0.8539 | p = 0.7691 | |
| Behavior | χ2 Test Comparing Individuals | χ2 Test Comparing Treatments | ||
| Activity on land | χ2 = 31.44 | χ2 = 0 | χ2 = 31.44 | χ2 = 0 |
| p = 2.055 × 10−8 | p = 1 | p = 2.055 × 10−8 | p = 1 | |
| Activity in water | χ2 = 71.67 | χ2 = 31.14 | χ2 = 5.4724 | χ2 = 1.140 |
| p < 2.2 × 10−16 | p = 2.391 × 10−8 | p = 0.01932 | p = 0.2858 | |
| Stereotypic | χ2 = 11.406 | χ2 = 1.034 | χ2 = 1.199 | χ2 = 29.12 |
| p = 0.0007321 | p = 0.3092 | p = 0.2736 | p = 6.79 × 10−8 | |
| Inactive | χ2 = 108.5 | χ2 = 166.4 | χ2 = 31.51 | χ2 = 9.219 |
| p < 2.2 × 10−16 | p < 2.2 × 10−16 | p = 1.981 × 10−8 | p = 0.002395 | |
| Inside | χ2 = 0 | χ2 = 81 | χ2 = 16.12 | χ2 = 28.47 |
| p = 1 | p < 2.2 × 10−16 | p = 5.932 × 10−5 | p = 9.513 × 10−8 | |
Appendix E
Appendix E.1. Moving Medians






Appendix E.2. Moving Variances






References
- Myers, P.; Young, J. Consistent individual behavior: Evidence of personality in black bears. J. Ethol. 2018, 36, 117–124. [Google Scholar] [CrossRef] [Green Version]
- Santicchia, F.; Wauters, L.A.; Dantzer, B.; Westrick, S.E.; Ferrari, N.; Romeo, C.; Palme, R.; Preatoni, D.G.; Martinoli, A. Relationships between personality traits and the physiological stress response in a wild mammal. Curr. Zool. 2019, 66, 197–204. [Google Scholar] [CrossRef]
- Thys, B.; Lambreghts, Y.; Pinxten, R.; Eens, M. Nest defence behavioral reaction norms: Testing life-history and parental investment theory predictions. R. Soc. Open Sci. 2019, 6, 182180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montiglio, P.-O.; Garant, D.; Pelletier, F.; Re´ale, D. Personality differences are related to long-term stress reactivity in a population of wild eastern chipmunks, Tamias striatus. Anim. Behav. 2012, 84, 1071–1079. [Google Scholar] [CrossRef]
- Luo, L.; Reimert, I.; de Haas, E.N.; Kemp, B.; Bolhuis, J.E. Effects of early and later life environmental enrichment and personality on attention bias in pigs (Sus scrofa domesticus). Anim. Cogn. 2019, 22, 959–972. [Google Scholar] [CrossRef] [Green Version]
- Dingemanse, N.J.; Kazem, A.J.N.; Réale, D.; Wright, J. Behavioural reaction norms: Animal personality meets individual plasticity. Trends Ecol. Evol. 2009, 25, 82–89. [Google Scholar] [CrossRef]
- Wilson, V.; Guenther, A.; Øverli, Ø.; Seltmann, M.W.; Altschul, D. Future directions for personality research: Contributing new insights to the understanding of animal behavior. Animals 2019, 9, 240. [Google Scholar] [CrossRef] [Green Version]
- Clubb, R.; Mason, G.J. Animal welfare: Captivity effects on wide-ranging carnivores. Nature 2003, 425, 473–474. [Google Scholar] [CrossRef]
- Mason, G.J.; Latham, N.R. Can’t stop, Won’t stop: Is stereotypy a reliable animal welfare indicator? Anim. Welf. 2004, 13, 57–69. [Google Scholar]
- Shepherdson, D.; Lewis, K.D.; Carlstead, K.; Bauman, J.; Perrin, N. Individual and environmental factors associated with stereotypic behavior and fecal glucocorticoid metabolite levels in zoo housed polar bears. Appl. Anim. Behav. Sci. 2013, 147, 268–277. [Google Scholar] [CrossRef]
- Carlstead, K.; Seidensticker, J. Seasonal variation in stereotypic pacing in an american black bear Ursus americanus. Behav. Process. 1991, 25, 155–161. [Google Scholar] [CrossRef]
- Ross, S.R. Issues of choice and control in the behaviour of a pair of captive polar bears Ursus maritimus. Behav. Process. 2006, 73, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Shyne, A. Meta-analytic review of the effects of enrichment on stereotypic behavior in zoo mammals. Zoo Biol. 2006, 25, 317–337. [Google Scholar] [CrossRef]
- Cless, I.T.; Voss-Hoynes, H.A.; Ritzmann, R.E.; Lukas, K.E. Defining pacing quantitatively: A comparison of gait characteristics between pacing and non-repetitive locomotion in zoo-housed polar bears. Appl. Anim. Behav. Sci. 2015, 169, 78–85. [Google Scholar] [CrossRef]
- Carlstead, K.; Seidensticker, J.; Baldwin, R. Environmental enrichment for zoo bears. Zoo Biol. 1991, 10, 3–16. [Google Scholar] [CrossRef]
- Rose, P.E.; Nash, S.M.; Riley, L.M. To pace or not to pace? A Review of what abnormal repetitive behavior tells us about zoo animal management. J. Vet. Behav. 2017, 20, 11–21. [Google Scholar] [CrossRef]
- Altmann, J. Observational study of behavior: Sampling methods. Behaviour 1974, 49, 227–267. [Google Scholar] [CrossRef] [Green Version]
- Bashaw, M.J.; Kelling, A.S.; Bloomsmith, M.A.; Maple, T.L. Environmental effects on the behavior of zoo-housed lions and tigers, with a case study of the effects of a visual barrier on pacing. J. Appl. Anim. Welf. Sci. 2007, 10, 95–109. [Google Scholar] [CrossRef]
- Pertoldi, C.; Bahrndorff, S.; Novicic, Z.K.; Rohde, P.D. The novel concept of ”behavioral instability” and its potential applications. Symmetry 2016, 8, 135. [Google Scholar] [CrossRef] [Green Version]
- Bech-Hansen, M.; Kallehauge, R.M.; Bruhn, D.; Castenschiold, J.H.F.; Gehrlein, J.B.; Laubek, B.; Jensen, L.F.; Pertoldi, C. Effect of landscape elements on the symmetry and variance of the spatial distribution of individual birds within foraging flocks of geese. Symmetry 2019, 11, 1103. [Google Scholar] [CrossRef] [Green Version]
- Clark, F.; King, A.J. A critical review of zoo-based olfactory enrichment. In Chemical Signals in Vertebrates 11; Hurst, J.L., Beynon, R.J., Roberts, S.C., Wyatt, T.D., Eds.; Springer: New York, NY, USA, 2008. [Google Scholar]
- RStudio Team. RStudio: Integrated Development for R; RStudio, Inc.: Boston, MA, USA, 2016. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Leys, C.; Ley, C.; Klein, O.; Bernard, P.; Licata, L. Detecting outliers: Do not use standard deviation around the mean, use absolute deviation around the median. J. Exp. Soc. Psychol. 2013, 49, 764–766. [Google Scholar] [CrossRef] [Green Version]
- Zar, J.H. Biostatistical Analysis, 4th ed.; Pearson: London, UK; Northern Illinois University: DeKalb, IL, USA, 1999. [Google Scholar]
- Yosef, R.; Raz, M.; Ben-Baruch, N.; Shmueli, L.; Kosicki, L.J.Z.; Fratczak, M.; Tryjanowski, P. Directional preferences of dogs’ changes in the presence of a bar magnet: Educational experiments in Israel. J. Vet. Behav. 2019, in press. [Google Scholar] [CrossRef]
- Rousseeuw, P.J.; Croux, C. Alternatives to the median absolute deviation. J. Am. Stat. Assoc. 1993, 88, 1273–1283. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Behavior | Description |
|---|---|
| Activity on land | Locomotion and interaction with objects while on land |
| Activity in water | Locomotion and interaction with objects while submerged in water |
| Social play | Individuals interacting playfully of fighting with each other, possibly while interacting with objects. |
| Stereotypic | Repeating a specific walking pattern or movement aimlessly |
| Inactive | Resting or sleeping; laying down or sitting with minimal movement |
| Inside | Inside the den and therefore out of sight |
| Other | Eating, drinking, urinating, defecating, maintenance of coat (e.g., by rolling in gravel) and out of sight due to blind camera angles |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cathrine Linder, A.; Gottschalk, A.; Lyhne, H.; Gade Langbak, M.; Hammer Jensen, T.; Pertoldi, C. Using Behavioral Instability to Investigate Behavioral Reaction Norms in Captive Animals: Theoretical Implications and Future Perspectives. Symmetry 2020, 12, 603. https://0-doi-org.brum.beds.ac.uk/10.3390/sym12040603
Cathrine Linder A, Gottschalk A, Lyhne H, Gade Langbak M, Hammer Jensen T, Pertoldi C. Using Behavioral Instability to Investigate Behavioral Reaction Norms in Captive Animals: Theoretical Implications and Future Perspectives. Symmetry. 2020; 12(4):603. https://0-doi-org.brum.beds.ac.uk/10.3390/sym12040603
Chicago/Turabian StyleCathrine Linder, Anne, Anika Gottschalk, Henriette Lyhne, Marie Gade Langbak, Trine Hammer Jensen, and Cino Pertoldi. 2020. "Using Behavioral Instability to Investigate Behavioral Reaction Norms in Captive Animals: Theoretical Implications and Future Perspectives" Symmetry 12, no. 4: 603. https://0-doi-org.brum.beds.ac.uk/10.3390/sym12040603