
First Report of Small Shelly Fossils from the Cambrian Miaolingian Limestones (Zhangxia and Hsuzhuang Formations) in Yiyang County, Henan Province of North China



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction

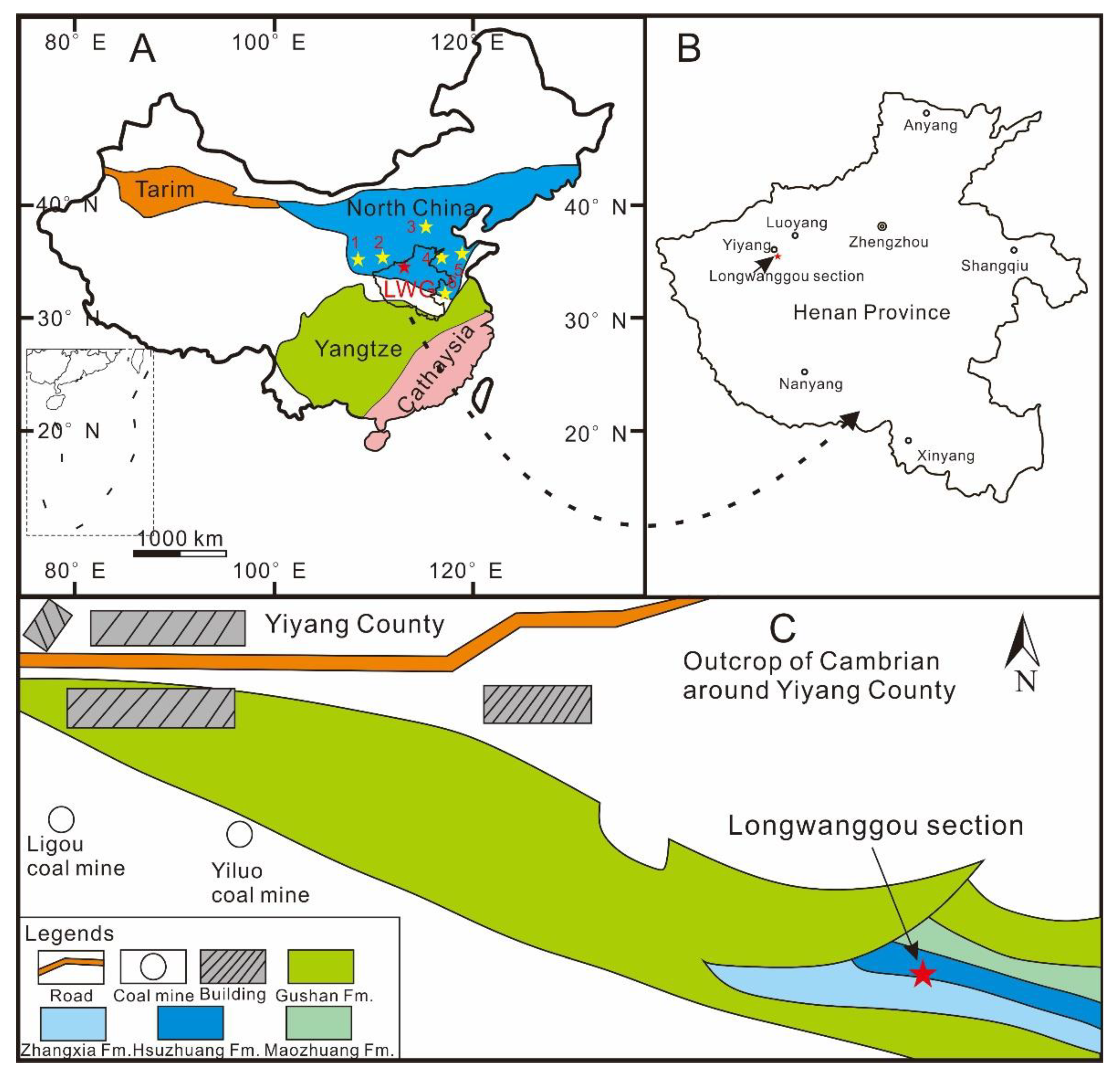
2. Geological Background
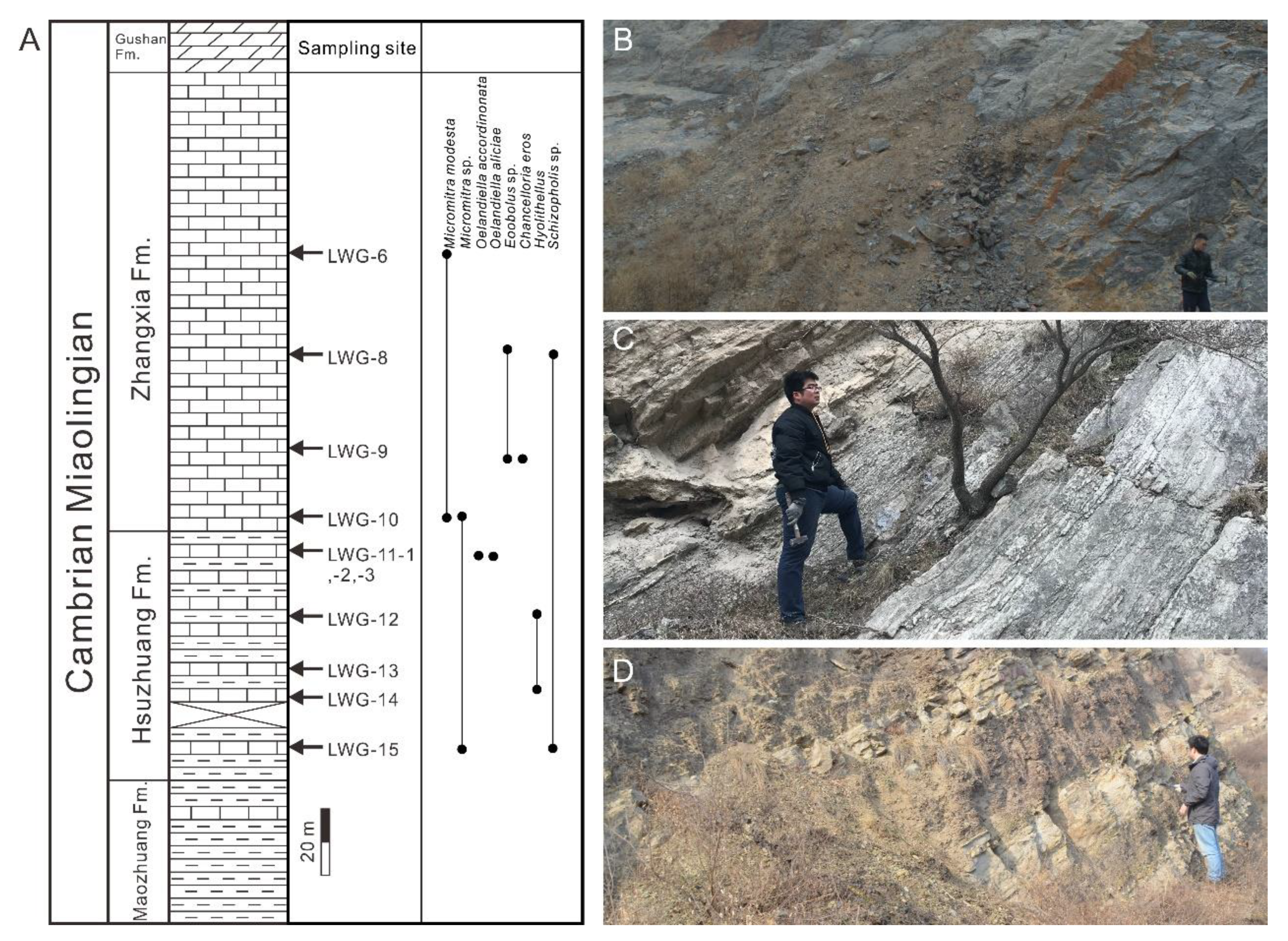
3. Methods and Material
4. Previous Paleontological Work on the Zhangxia and Hsuzhuang Formations of Henan Province
5. Fossil Assemblage from the Zhangxia and Hsuzhuang Formations at the Longwanggou Section and Global Correlations
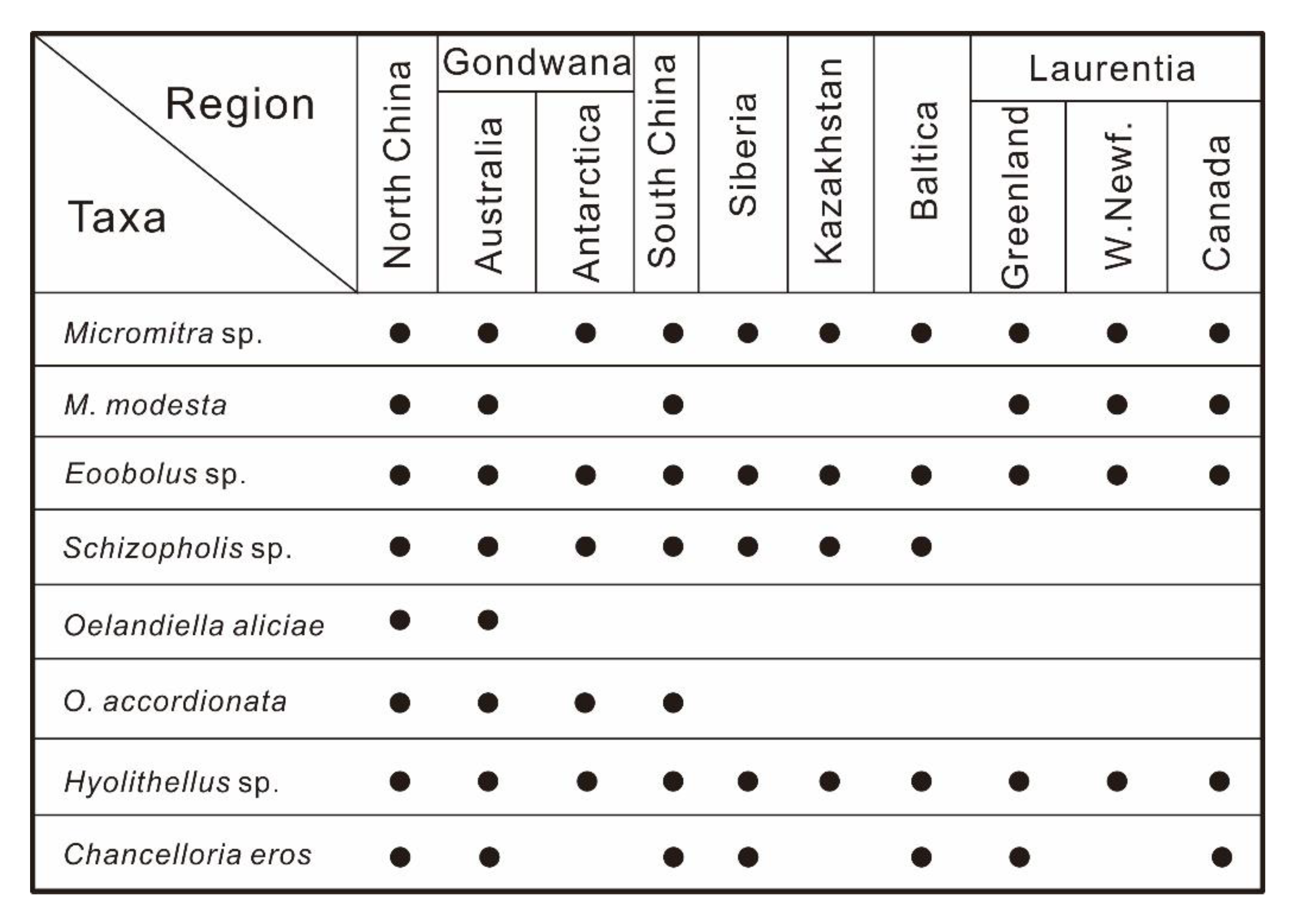
5.1. Fossil Assemblages
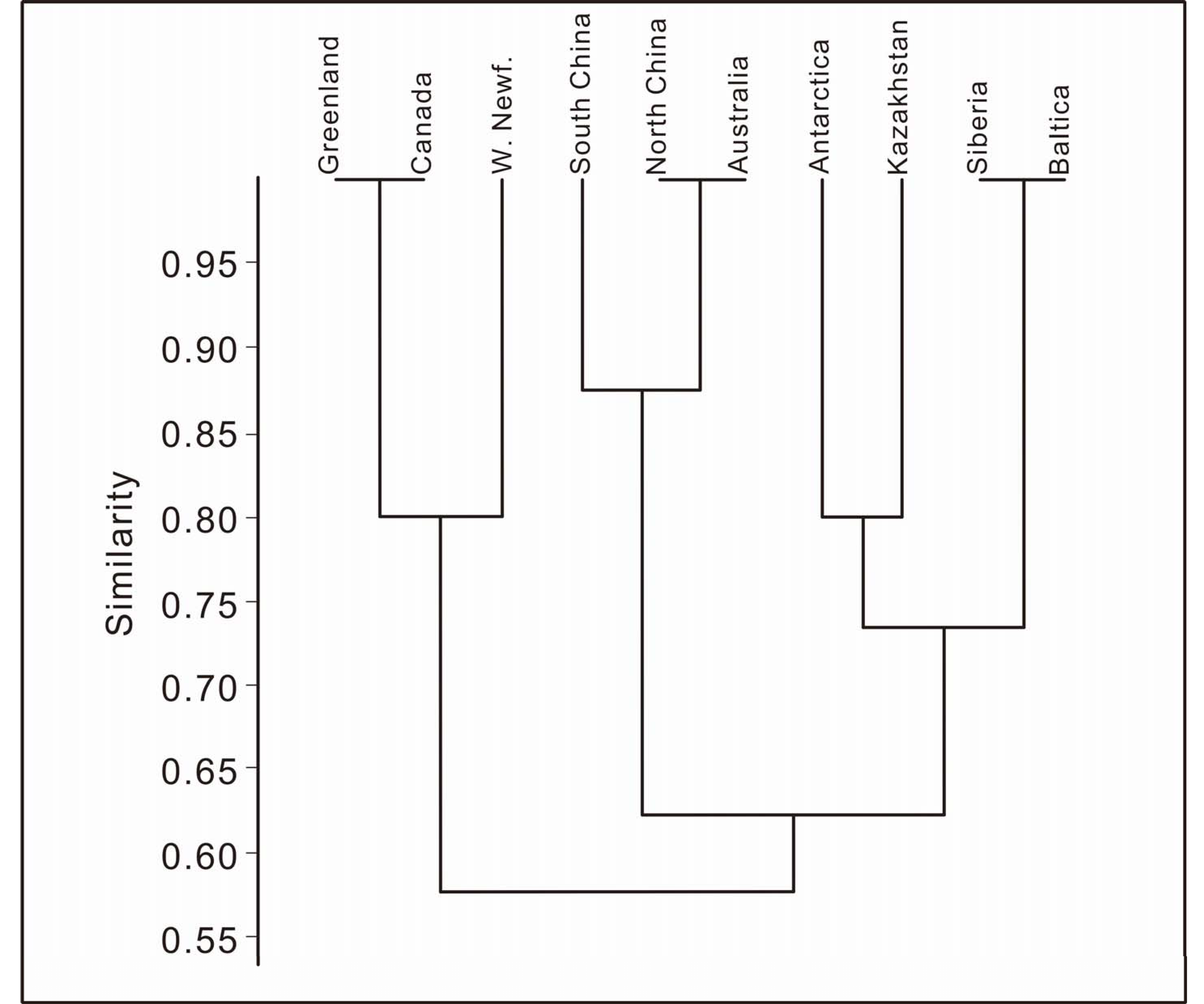
5.2. Global Correlations
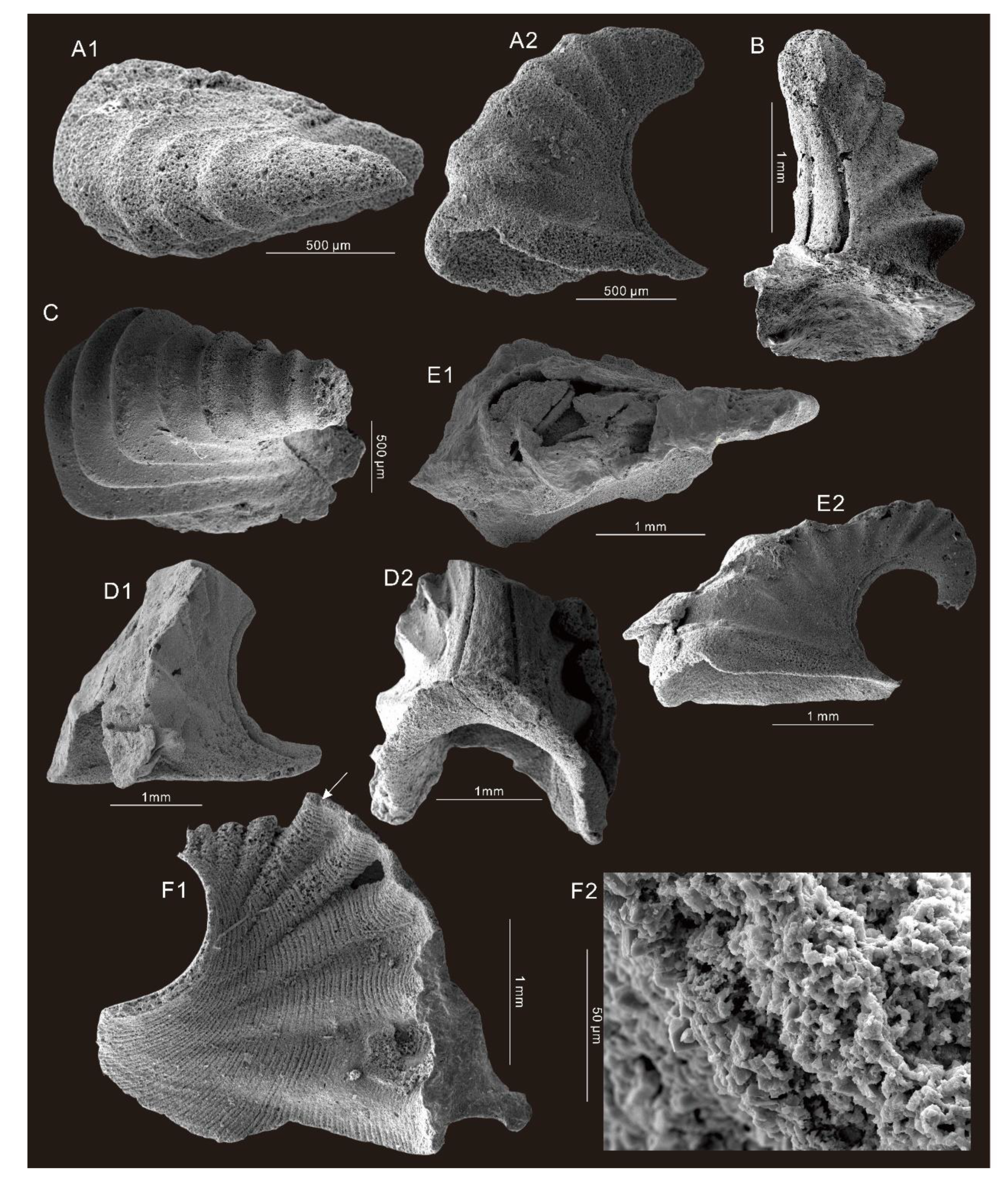
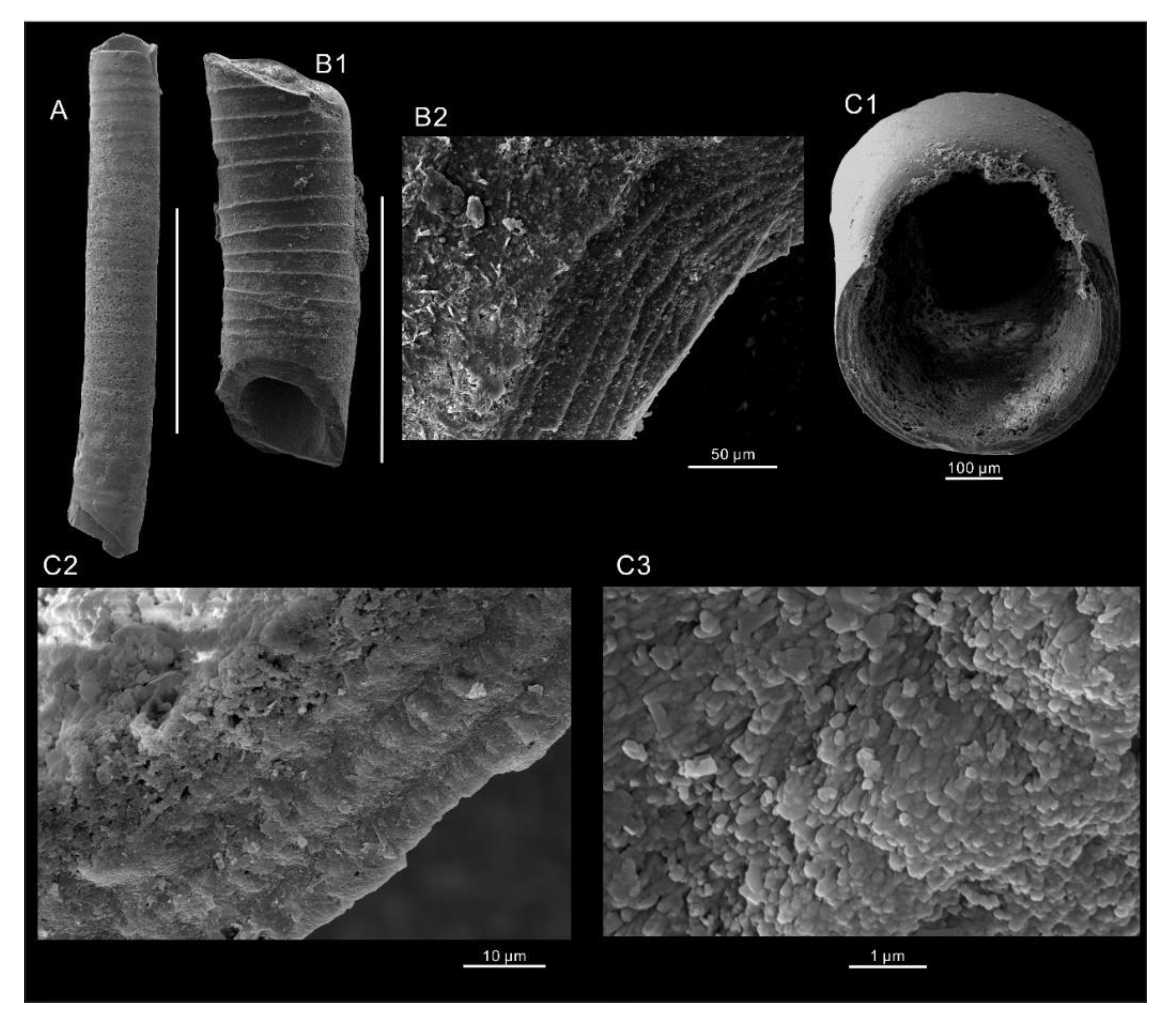
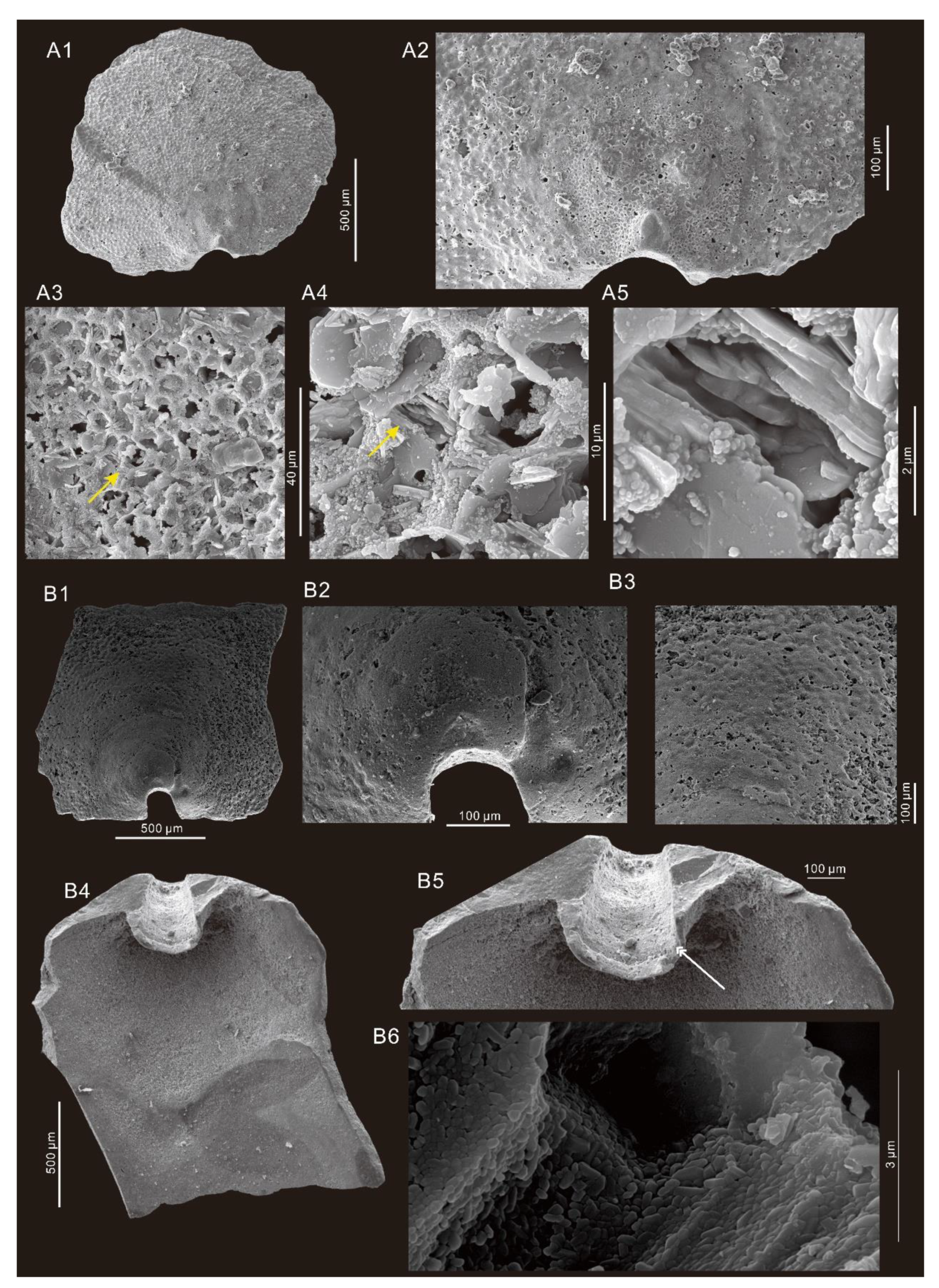
6. Systematic Paleontology
- Phylum Brachiopoda Duméril, 1806
- Class Paterinata Williams et al., 1996
- Order Paterinida Rowell, 1965
- Superfamily Paterinoidea Schuchert, 1893
- Family Paterinidae Schuchert 1893
- Genus Micromitra Meek, 1873
- Micromitra modesta Lochman, 1940.
- Micromitra sp.
- Order Lingulida Waagen, 1885
- Family Eoobolidae Holmer et al., 1996
- Genus Eoobolus Matthew, 1902
- Eoobolus sp.
- Superfamily Acrotheloidea Walcott and Schuchert in Walcott, 1908
- Family Botsfordiidae Schindewolf, 1955
- Genus Schizopholis Waagen, 1885
- 1885 Discinolepis Waagen
- 1986 Karathele Koneva
- Schizopholis sp.
- Phylum Mollusca Cuvier, 1795
- Class Helcionelloida Peel, 1991
- Order Helcionellida Geyer, 1994
- Family Helcionellidae Wenz, 1938
- Genus Oelandiella Vostokova, 1962
- Oelandiella accordionata Runnegar and Jell, 1976.
- Oelandiella aliciae Brock, 1998.
- 1976 Latouchella accordionata Runnegar & Jell, figs. 10C.
- 1998 Latouchella aliciae Brock, figs. 3.7–12; 4.1–3.
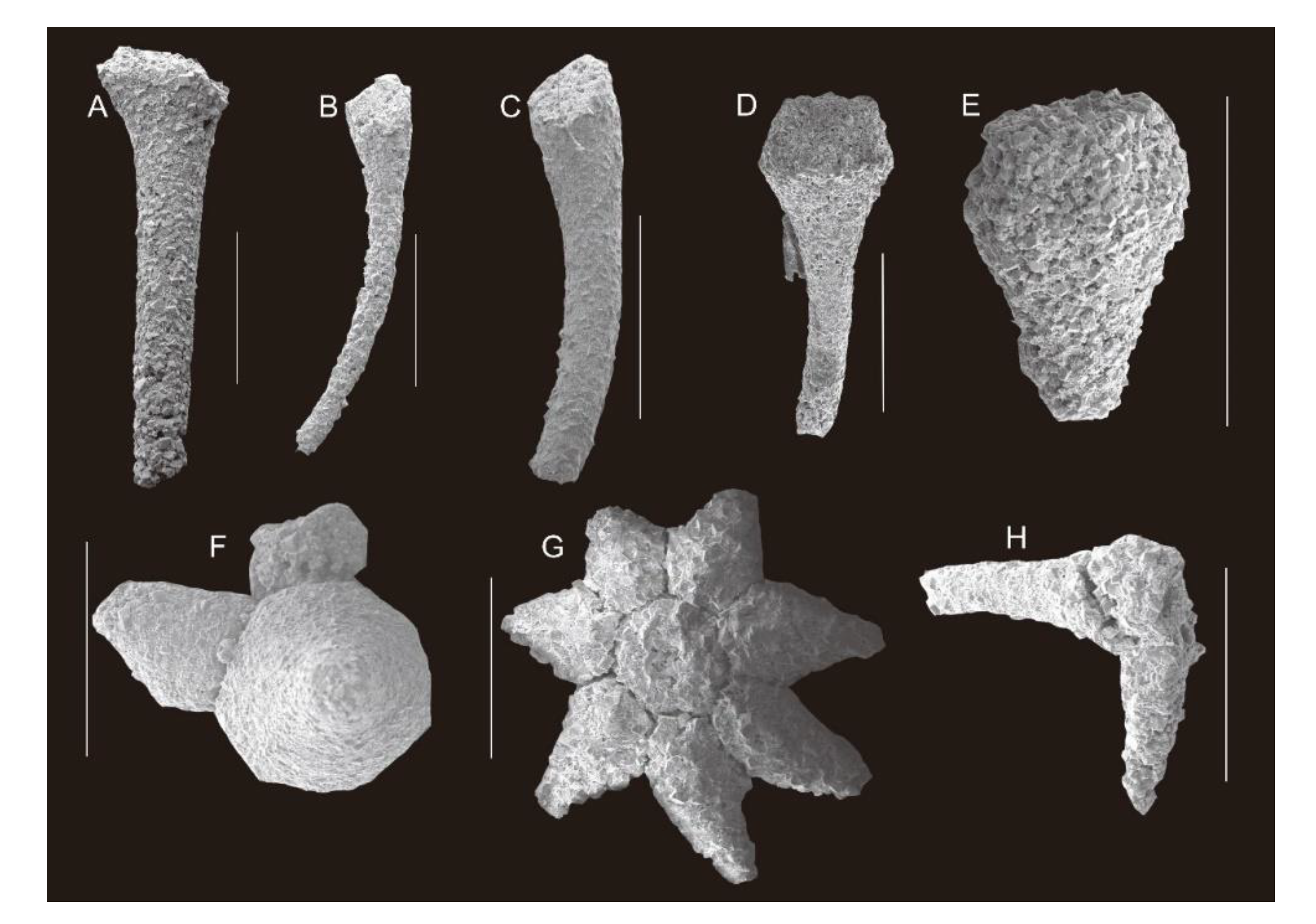
- Order Hyolithelminthida Fisher, 1962
- Family Hyolithellidae Walcott, 1886
- Genus Hyolithellus Billings, 1871
- Hyolithellus sp.
- Phylum Uncertain
- Class Hyolitha Marek, 1963
- Hyolithid sp. Indet
- Phylum Echinodermata Bruguière, 1791
- INDET. Echinoderm ossicles
- Phylum Porifera Grant, 1872
- Class, Order, Family, Genus and Species unassigned
- Sponge spicule
- Phylum and Class uncertain
- Order Chancelloriida Walcott, 1920
- Genus Chancelloria Walcott, 1920
- Chancelloria eros Walcott, 1920
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Z.F.; Zhang, Z.L.; Li, G.X.; Holmer, L.E. The Cambrian Brachiopod Fauna from the First-Trilobite Age Shuijingtuo Formation in the Three Gorges Area of China. Palaeoworld 2016, 25, 333–355. [Google Scholar] [CrossRef]
- Zhang, Z.; Ghobadi Pour, M.; Popov, L.E.; Holmer, L.E.; Chen, F.; Chen, Y.; Brock, G.A.; Zhang, Z. The Oldest Cambrian Trilobite—Brachiopod Association in South China. Gondwana Res. 2021, 89, 147–167. [Google Scholar] [CrossRef]
- Pei, F. First Discovery of Yochelcionella from the Lower Cambrian in China and Its Significance. Acta Micropalaeontol. Sin. 1985, 2, 395–401. [Google Scholar]
- Li, L.; Zhang, X.; Skovsted, C.B.; Yun, H.; Pan, B.; Li, G. Homologous Shell Microstructures in Cambrian Hyoliths and Molluscs. Palaeontology 2019, 62, 515–532. [Google Scholar] [CrossRef]
- Li, L.; Zhang, X.; Skovsted, C.B.; Yun, H.; Li, G.; Pan, B. Shell Microstructures of the Helcionelloid Mollusc Anabarella australis from the Lower Cambrian (Series 2) Xinji Formation of North China. J. Syst. Palaeontol. 2019, 17, 1479–1489. [Google Scholar] [CrossRef]
- Li, L.; Skovsted, C.B.; Yun, H.; Betts, M.J.; Zhang, X. New Insight into the Soft Anatomy and Shell Microstructures of Early Cambrian Orthothecids (Hyolitha): Anatomy and Biomineralization. Proc. R. Soc. B 2020, 287, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhang, X.; Skovsted, C.B.; Yun, H.; Pan, B.; Li, G. Revisiting the Molluscan Fauna from the Cambrian (Series 2, Stages 3–4) Xinji Formation of North China. Pap. Palaeontol. 2021, 7, 521–564. [Google Scholar] [CrossRef] [Green Version]
- Pan, B.; Skovsted, C.B.; Brock, G.A.; Topper, T.P.; Holmer, L.E.; Li, L.Y.; Li, G.X. Early Cambrian Organophosphatic Brachiopods from the Xinji Formation, at Shuiyu Section, Shanxi Province, North China. Palaeoworld 2020, 29, 512–533. [Google Scholar] [CrossRef]
- Chen, F.; Zhang, Z.; Betts, M.J.; Zhang, Z.; Liu, F. First Report on Guanshan Biota (Cambrian Stage 4) at the Stratotype Area of Wulongqing Formation in Malong County, Eastern Yunnan, China. Geosci. Front. 2019, 10, 1459–1476. [Google Scholar] [CrossRef]
- Fu, D.; Tong, G.; Dai, T.; Liu, W.; Yang, Y.; Zhang, Y.; Cui, L.; Li, L.; Yun, H.; Wu, Y.; et al. The Qingjiang Biota—A Burgess Shale—Type Fossil Lagerstätte from the Early Cambrian of South China. Science 2019, 1342, 1338–1342. [Google Scholar] [CrossRef]
- Shu, D.; Han, J. Core Value of the Chengjiang Fauna: Formation of the Animal Kingdom and the Birth of Basic Human Organs. Earth Sci. Front. 2020, 27, 382–412. [Google Scholar] [CrossRef]
- Duan, X.; Liang, Y.; Holmer, L.E.; Zhang, Z. First Report of Acrotretoid Brachiopod Shell Beds in the Lower Cambrian (Stage 4) Guanshan Biota of Eastern Yunnan, South China. J. Paleontol. 2021, 95, 40–55. [Google Scholar] [CrossRef]
- Zhang, Z.; Strotz, L.C.; Topper, T.P.; Chen, F.; Chen, Y.; Liang, Y.; Zhang, Z.; Skovsted, C.B.; Brock, G.A. An Encrusting Kleptoparasite-Host Interaction from the Early Cambrian. Nat. Commun. 2020, 11, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Holmer, L.E.; Liang, Y.; Chen, Y.; Duan, X. The Oldest ’Lingulellotreta’ (Lingulata, Brachiopoda) from China and Its Phylogenetic Significance: Integrating New Material from the Cambrian Stage 3–4 Lagerstätten in Eastern Yunnan, South China. J. Syst. Palaeontol. 2020, 18, 945–973. [Google Scholar] [CrossRef]
- Peng, J.; Zhao, Y.; Wu, Y.; Yuan, J.; Tai, T. The Balang Fauna—A New Early Cambrian Fauna from Kaili City, Guizhou Province. Chin. Sci. Bul. 2005, 50, 1055–1057. (In Chinese) [Google Scholar] [CrossRef]
- Zhang, X.; Liu, W.; Zhao, Y. Cambrian Burgess Shale-Type Lagerstätten in South China: Distribution and Significance. Gondwana Res. 2008, 18, 255–262. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, Z.; Holmer, L.E.; Li, G. First Report of Linguloid Brachiopods with Soft Parts from the Lower Cambrian (Series 2, Stage 4) of the Three Gorges Area, South China. Annales Paléontologie 2015, 101, 167–177. [Google Scholar] [CrossRef] [Green Version]
- Duan, X.; Betts, M.J.; Holmer, L.E.; Chen, Y.; Liu, F.; Liang, Y.; Zhang, Z. Early Cambrian (Stage 4) Brachiopods from the Shipai Formation in the Three Gorges Area of South China. J. Paleontol. 2021, 95, 497–526. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, D.; Lieberman, B.S. New Isoxys (Arthropoda) from the Cambrian Mantou Formation, Shandong Province. Acta Palaeontol. Sin. 2010, 49, 398–406, (In Chinese with English Abstract). [Google Scholar]
- Han, Y.; Wang, P.; Mao, Y.; Gao, J. New Material of Nisusia Walcott, 1905 from the Manto Formation (2-3 Series, Cambrian), Tangshan, Hebei Province. Acta Palaeontol. Sin. 2018, 57, 295–303, (In Chinese with English Abstract). [Google Scholar]
- Han, Q.; Li, G.; Wang, P.; Gao, J.; Zhang, T.; Sun, Z. New Material of Chancelloriids from Cambrian, North China. Geol. J. China Univ. 2019, 25, 633–640, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Liu, Y.; Sun, Z.; Zeng, H.; Zhao, F. Brachiopods from the Upper Part of the Mantou Formation (Cambrian, Miaolingian, Wuliuan) in Weifang, Shandong, North China. Acta Palaeontol. Sin. 2021, 60, 150–165, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Sun, Z.; Zeng, H.; Zhao, F. Digestive Structures in Cambrian Miaolingian Trilobites from Shandong. Acta Palaeontol. Sin. 2021, 60, 166–175, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Mei, S. Middle and Upper Cambrian Inarticulate Brachiopods from Wanxian, Hebei, North China. Acta Palaeontol. Sin. 1993, 32, 400–439, (In Chinese with English Abstract). [Google Scholar]
- Park, T.Y.; Woo, J.; Lee, D.J.; Lee, D.C.; Lee, S.B.; Han, Z.; Chough, S.K.; Choi, D.K. A Stem-Group Cnidarian described from the mid-Cambrian of China and its significance for Cnidarian evolution. Nat. Commun. 2011, 2, 442. [Google Scholar] [CrossRef] [PubMed]
- Yun, H.; Zhang, X.; Li, L.; Zhang, M.; Liu, W. Skeletal Fossils and Microfacies Analysis of the Lowermost Cambrian in the Southwestern Margin of the North China Platform. J. Asian Earth Sci. 2016, 129, 54–66. [Google Scholar] [CrossRef]
- Pan, B.; Skovsted, C.B.; Sun, H.; Li, G. Biostratigraphical and Palaeogeographical Implications of Early Cambrian Hyoliths from the North China Platform. Alcheringa 2019, 43, 351–380. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electro. 2001, 4, 1–9. [Google Scholar]
- Pei, F. The Cambrian of North China Type in Henan Province. Reg. Geol. China 1991, 3, 210–220. [Google Scholar]
- Pei, F. Division and Correlation of the North China Type Cambrian Biostratigraphic Units of Henan Province. Henan Geol. 2000, 18, 97–106. [Google Scholar]
- Liu, Q.; Huang, D.; Gong, Y. Sponge Fossils from the Cambrian Mantou Formation of Hebei, Henan, Central China. Earth Sci. J. China Univ. Geosci. 2012, 37, 129–135, (In Chinese with English Abstract). [Google Scholar]
- Yun, H.; Zhang, X.; Brock, G.A.; Li, L.; Li, G. Biomineralization of the Cambrian Chancelloriids. Geology 2021, 49, 623–628. [Google Scholar] [CrossRef]
- Pan, B.; Brock, G.A.; Skovsted, C.B.; Betts, M.J.; Topper, T.P.; Li, G. Paterimitra pyramidalis Laurie, 1986, the First Tommotiid Discovered from the Early Cambrian of North China. Gondwana Res. 2018, 63, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Pan, B.; Topper, T.P.; Skovsted, C.B.; Miao, L.; Li, G. Occurrence of Microdictyon from the Lower Cambrian Xinji Formation along the Southern Margin of the North China Platform. J. Paleontol. 2018, 92, 59–70. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Zhang, X.; Yun, H.; Li, G. Complex Hierarchical Microstructures of Cambrian Mollusk Pelagiella: Insight into Early Biomineralization and Evolution. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmer, L.E.; Popov, L.E. Class Lingulata. In Treatise on Invertebrate Paleontology, Part H, Brachiopoda (Revised), Part 2; Williams, A., Brunton, C.H.C., Carlson, S.J., Eds.; University of Kansas Press: Lawrence, KS, USA, 2000; pp. 20–146. [Google Scholar]
- Engelbretsen, M.J. Middle Cambrian Lingulate Brachiopods from the Murrawong Creek Formation, Northeastern New South Wales. Hist. Biol. 1996, 11, 69–99. [Google Scholar] [CrossRef]
- Zell, M.G.; Rowell, A.J. Brachiopods of the Holm Dal Formation (Late Middle Cambrian), Central North Grenland. Meddr. Grønland. Geosci. 1988, 20, 119–144. [Google Scholar]
- Robson, S.P.; Pratt, B.R. Cambrian and Ordovician Linguliform Brachiopods from the Shallow Bay Formation (Cow Head Group), Western Newfoundland. J. Paleontol. 2001, 75, 241–260. [Google Scholar] [CrossRef]
- Robson, S.P.; Nowlan, G.S.; Pratt, B.R. Middle to Upper Cambrian Linguliformean Brachiopods from the Deadwood Formation of Subsurface Alberta and Saskatchewan, Canada. J. Paleontol. 2003, 77, 201–211. [Google Scholar] [CrossRef]
- Engelbretsen, M.J.; Peng, S. Middle Cambrian (Wulingian) Linguliformean Brachiopods from the Paibi Section, Huaqiao Formation, Hunan Province, South China. Mem. Assoc. Australas. Palaeontol. 2007, 34, 311–329. [Google Scholar]
- Brock, G.A. Middle Cambrian Molluscs from the Southern New England Fold Belt, New South Wales, Australia. Geobios 1998, 31, 571–586. [Google Scholar] [CrossRef]
- Runnegar, B.; Jell, P.A. Australian Middle Cambrian Molluscs and Their Bearing on Early Molluscan Evolution. Alcheringa 1976, 1, 109–138. [Google Scholar] [CrossRef]
- Pojeta, J.; Simes, J.E.; Cooper, R.A. New Zealand Cambrian and Ordovician Micromolluscs. Mem. Assoc. Australas. Palaeontol. 2014, 1, 1–16. [Google Scholar]
- Yang, Y.; Zhao, Y.; Peng, J.; Yang, X. Oelandiella from the Cambrian Tsinghsutung Formation of Jianhe County, Guizhou. Acta Palaeontol. Sin. 2012, 51, 308–319, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Duan, X.; Yang, X.; Wang, Y. Oelandiella from the Cambrian Jialao Formation of Guizhou, China. Acta Palaeontol. Sin. 2017, 56, 529–537. [Google Scholar] [CrossRef]
- Percival, I.G.; Quinn, C.D.; Brock, G.A. A Review of Cambrian and Ordovician Stratigraphy in New South Wales. Q. Notes Geol. Surv. N. S. Wales 2011, 137, 1–39. [Google Scholar] [CrossRef]
- Zhao, H.; Zhang, S.; Zhu, M.; Ding, J.; Li, H.; Yang, T.; Wu, H. Paleomagnetic Insights into the Cambrian Biogeographic Conundrum: Did the North China Craton Link Laurentia and East Gondwana? Geology 2021, 49, 372–376. [Google Scholar] [CrossRef]
- Pang, K.; Wu, C.; Sun, Y.; Ouyang, Q.; Yuan, X.; Shen, B.; Lang, X.; Wang, R.; Chen, Z.; Zhou, C. New Ediacara-Type Fossils and Late Ediacaran Stratigraphy from the Northern Qaidam Basin (China): Paleogeographic Implications. Geology 2021, XX, 1–5. [Google Scholar] [CrossRef]
- McKenzie, N.R.; Hughes, N.C.; Myrow, P.M.; Choi, D.K.; Park, T.Y. Trilobites and Zircons Link North China with the Eastern Himalaya during the Cambrian. Geology 2011, 39, 591–594. [Google Scholar] [CrossRef]
- Han, Y.; Zhao, G.; Cawood, P.A.; Sun, M.; Eizenhöfer, P.R.; Hou, W.; Zhang, X.; Liu, Q. Tarim and North China Cratons Linked to Northern Gondwana through Switching Accretionary Tectonics and Collisional Orogenesis. Geology 2016, 44, 95–98. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Zhang, Z.; Popov, L.E.; Holmer, L.E. Earliest Ontogeny of Early Cambrian Acrotretoid Brachiopods—First Evidence for Metamorphosis and Its Implications. BMC Evol. Biol. 2018, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laurie, J.R. Paterinide Musculature and Vascular Systems.Pdf. BMR J. Aust. Geol. Geophy. 1987, 10, 261–265. [Google Scholar]
- Babcock, L.E.; Robison, R.A.; Rees, M.N.; Peng, S.; Saltzman, M.R. The Global Boundary Stratotype Section and Point (GSSP) of the Drumian Stage (Cambrian) in the Drum Mountains, Utah, USA. Episodes 2007, 30, 85–95. [Google Scholar] [CrossRef] [Green Version]
- Popov, L.E.; Bassett, M.G.; Zhemchuzhnikov, V.G.; Holmer, L.E.; Klishevich, I.A. Gondwanan Faunal Signatures from Early Palaeozoic Terranes of Kazakhstan and Central Asia: Evidence and Tectonic Implications. Geo. Soc. Lond. Spec. Pub. 2009, 325, 23–64. [Google Scholar] [CrossRef]
- Betts, M.J.; Paterson, J.R.; Jago, J.B.; Jacquet, S.M.; Skovsted, C.B.; Topper, T.P.; Brock, G.A. A New Lower Cambrian Shelly Fossil Biostratigraphy for Australia. Gondwana Res. 2016, 36, 176–208. [Google Scholar] [CrossRef] [Green Version]
- Bassett-Butt, L. Systematics, Biostratigraphy and Biogeography of Brachiopods and Other Fossils from the Middle Cambrian Nelson Limestone, Antarctica. GFF 2016, 138, 377–392. [Google Scholar] [CrossRef]
- Ushatinskaya, G.T. Protegulum and Brephic Shell of the Earliest Organophosphatic Brachiopods. Paleontol. J. 2016, 50, 37–47. [Google Scholar] [CrossRef]
- Williams, A.; Popov, L.E.; Holmer, L.E.; Cusack, M. The Diversity and Phylogeny of the Paterinate Brachiopods. Paleontology 1998, 41, 221–262. [Google Scholar]
- Popov, L.E.; Holmer, L.E.; Hughes, N.C.; Ghobadi Pour, M.; Myrow, P.M. Himalayan Cambrian Brachiopods. Pap. Palaeontol. 2015, 1, 345–399. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Jin, Y.; Fan, D. Brachiopod Fossils of China; Science Press: Beijing, China, 1964; ISBN 1261000043520. (In Chinese) [Google Scholar]
- Nemliher, J. A New Type of Shell Structure in a Phosphatic Brachiopod from the Cambrian of Estonia. Proc. Est. Acad. Sci. Geol. 2006, 55, 259–268. [Google Scholar]
- Skovsted, C.B.; Holmer, L.E. The Early Cambrian (Botomian) Stem Group Brachiopod Mickwitzia from Northeast Greenland. Acta Palaeontol. Pol. 2003, 48, 1–20. [Google Scholar]
- Lochman, C. Fauna of the Basal Bonneterre Dolomite (Upper Cambrian) of Southeastern Missouri. J. Paleontol. 1940, 14, 1–53. [Google Scholar]
- Percival, I.G.; Kruse, P.D. Middle Cambrian Brachiopods from the Southern Georgina Basin of Central Australia. Mem. Assoc. Australas. Palaeontol. 2014, 1, 349–402. [Google Scholar]
- Kruse, P. Cambrian Fauna of the Top Springs Limestone, Georgina Basin. Beagle Rec. Mus. Art Galleries North. Territ. 1991, 8, 169–188. [Google Scholar] [CrossRef]
- Holmer, L.; Popov, L.; Wrona, R. Early Cambrian Lingulate Brachiopods from Glacial Erratics of King George Island (South Shetland Islands), Antarctica. Acta Palaeontol. Pol. 1996, 55, 37–50. [Google Scholar]
- Li, G.; Holmer, L.E. Early Cambrian Lingulate Brachiopods from the Shaanxi Province, China. GFF 2004, 126, 193–211. [Google Scholar] [CrossRef]
- Balthasar, U. The Brachiopod Eoobolus from the Early Cambrian Mural Formation (Canadian Rocky Mountains). Paläontologische Zeitschrift 2009, 83, 407–418. [Google Scholar] [CrossRef]
- Ushatinskaya, G.T.; Korovnikov, I.V. Revision of the Early-Middle Cambrian Lingulida (Brachiopoda) from the Siberian Platform. Paleontol. J. 2014, 48, 26–40. [Google Scholar] [CrossRef]
- Streng, M.; Holmer, L.E.; Popov, L.E.; Budd, G.E. Columnar Shell Structures in Early Linguloid Brachiopods—New Data from the Middle Cambrian of Sweden. Earth Environ. Sci. Trans. R. Soc. Edinb. 2007, 98, 221–232. [Google Scholar] [CrossRef]
- Griesshaber, E.; Schmahl, W.W.; Neuser, R.; Pettke, T.; Blüm, M.; Mutterlose, J.; Brand, U. Crystallographic Texture and Microstructure of Terebratulide Brachiopod Shell Calcite: An Optimized Materials Design with Hierarchical Architecture. Am. Mineral. 2007, 92, 722–734. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, Z.; Holmer, L.E. Studies on the shell ultrastructure and ontogeny of the oldest Acrotretid Brachiopods from South China. Acta Palaeontol. Sin. 2017, 56, 483–503, (In Chinese with English abstract). [Google Scholar] [CrossRef]
- Pérez-Huerta, A.; Cusack, M.; McDonald, S.; Marone, F.; Stampanoni, M.; MacKay, S. Brachiopod Punctae: A Complexity in Shell Biomineralisation. J. Struct. Biol. 2009, 167, 62–67. [Google Scholar] [CrossRef]
- Betts, M.J.; Claybourn, T.M.; Brock, G.A.; Jago, J.B.; Skovsted, C.B.; Paterson, J.R. Shelly Fossils from the Lower Cambrian White Point Conglomerate, Kangaroo Island, Australia. Acta Palaeontol. Pol. 2019, 64, 489–522. [Google Scholar] [CrossRef]
- Poulsen, V. Notes on an Ordovician Acrotretacean Brachiopod from the Oslo Region. Bull. Geol. Soc. Den. 1971, 20, 265–278. [Google Scholar]
- Liu, Y.J.; Peng, J.; Zhao, Y.L.; Mao, Y.Q. First Report of Schizopholis (Lingulata, Brachiopoda) from the Tsinghsutung Formation (Cambrian Series 2, Stage 4) in Guizhou, China. Palaeoworld 2021, 30, 422–429. [Google Scholar] [CrossRef]
- Claybourn, T.M.; Skovsted, C.B.; Holmer, L.E.; Pan, B.; Myrow, P.M.; Topper, T.P.; Brock, G.A. Brachiopods from the Byrd Group (Cambrian Series 2, Stage 4) Central Transantarctic Mountains, East Antarctica: Biostratigraphy, Phylogeny and Systematics. Pap. Palaeontol. 2020, 6, 349–383. [Google Scholar] [CrossRef]
- Gubanov, A.P.; Peel, J.S. Oelandiella, the Earliest Cambrian Helcionelloid Mollusc from Siberia. Palaeontology 1999, 42, 211–222. [Google Scholar] [CrossRef]
- Skovsted, C.B.; Peel, J.S. Hyolithellus in Life Position from the Lower Cambrian of North Greenland. J. Paleontol. 2011, 85, 37–47. [Google Scholar] [CrossRef]
- Skovsted, C.B.; Balthasar, U.; Vinther, J.; Sperling, E.A. Small Shelly Fossils and carbon isotopes from the Early Cambrian (Stages 3–4) Mural Formation of Western Laurentia. Pap. Palaeontol. 2021, 7, 951–983. [Google Scholar] [CrossRef]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, Y.; Holmer, L.E.; Liang, Y.; Duan, X.; Zhang, Z. First Report of Small Shelly Fossils from the Cambrian Miaolingian Limestones (Zhangxia and Hsuzhuang Formations) in Yiyang County, Henan Province of North China. Minerals 2021, 11, 1104. https://0-doi-org.brum.beds.ac.uk/10.3390/min11101104
Hu Y, Holmer LE, Liang Y, Duan X, Zhang Z. First Report of Small Shelly Fossils from the Cambrian Miaolingian Limestones (Zhangxia and Hsuzhuang Formations) in Yiyang County, Henan Province of North China. Minerals. 2021; 11(10):1104. https://0-doi-org.brum.beds.ac.uk/10.3390/min11101104
Chicago/Turabian StyleHu, Yazhou, Lars E. Holmer, Yue Liang, Xiaolin Duan, and Zhifei Zhang. 2021. "First Report of Small Shelly Fossils from the Cambrian Miaolingian Limestones (Zhangxia and Hsuzhuang Formations) in Yiyang County, Henan Province of North China" Minerals 11, no. 10: 1104. https://0-doi-org.brum.beds.ac.uk/10.3390/min11101104